
NUSAP1 Binds ILF2 to Modulate R-Loop Accumulation and DNA Damage in Prostate Cancer
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
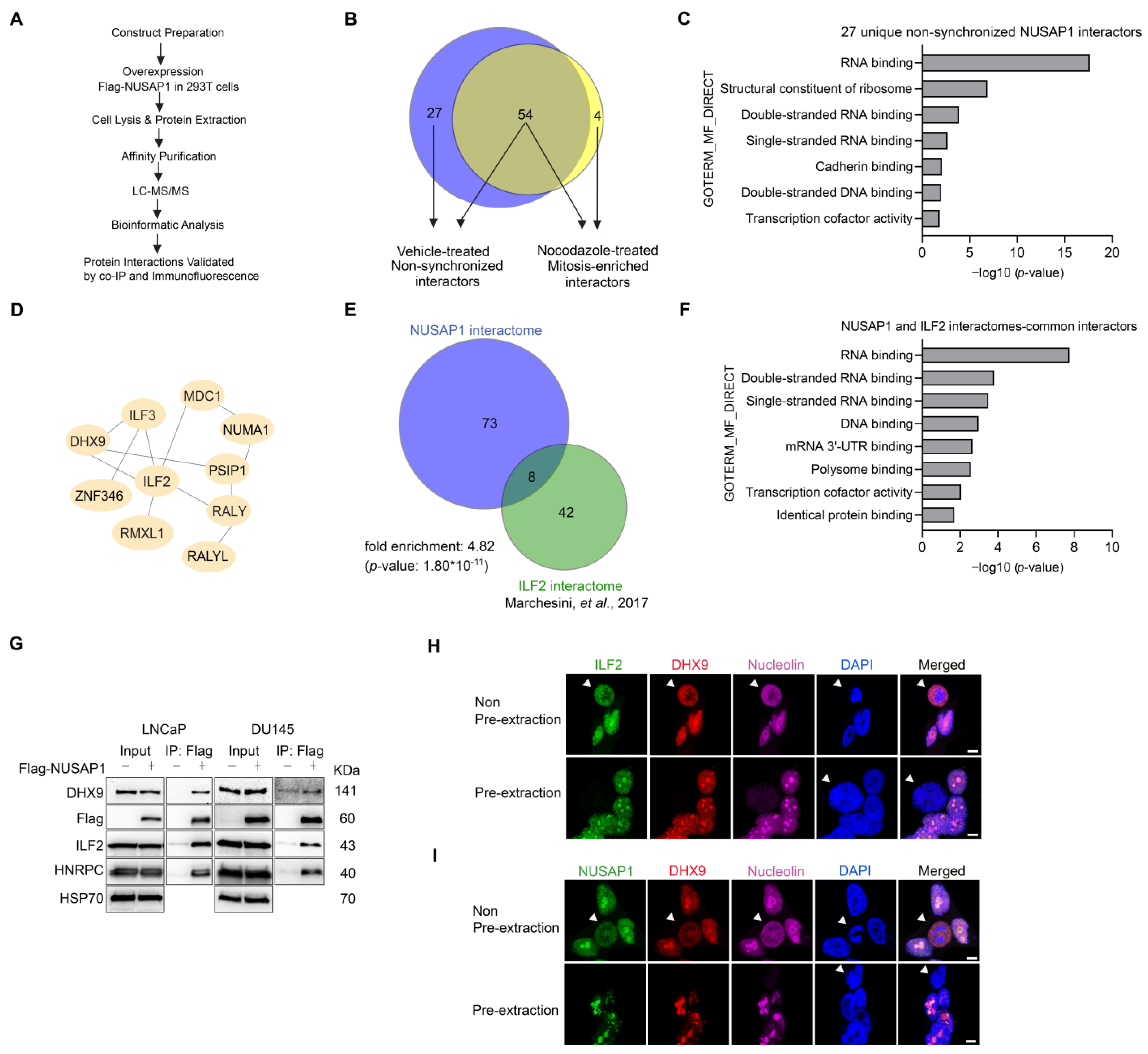
2.1. AP–MS Identifies NUSAP1 Interactions with ILF2 and DHX9
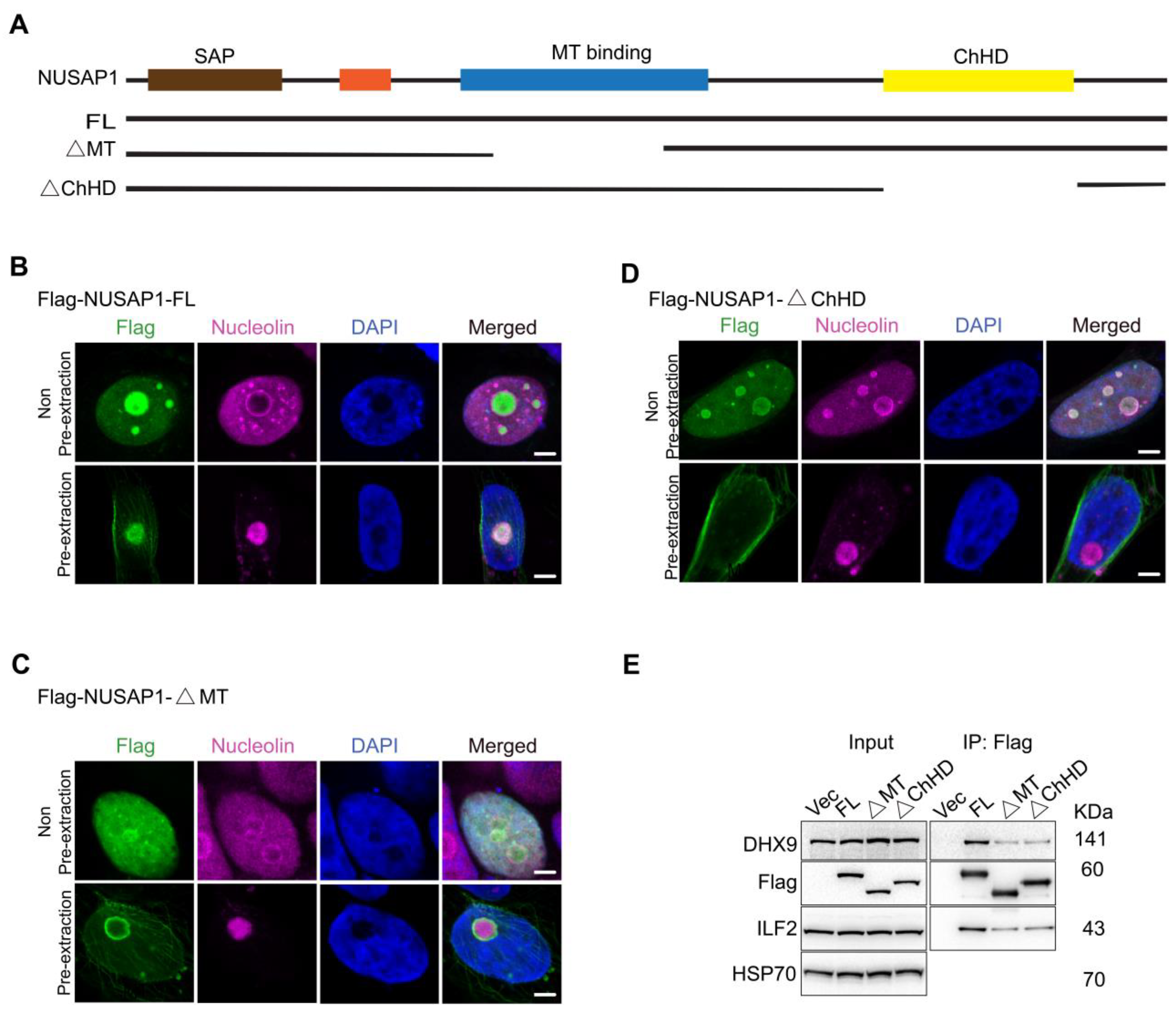
2.2. NUSAP1 MT and ChHD Domains Are Important for ILF2 and DHX9 Interactions
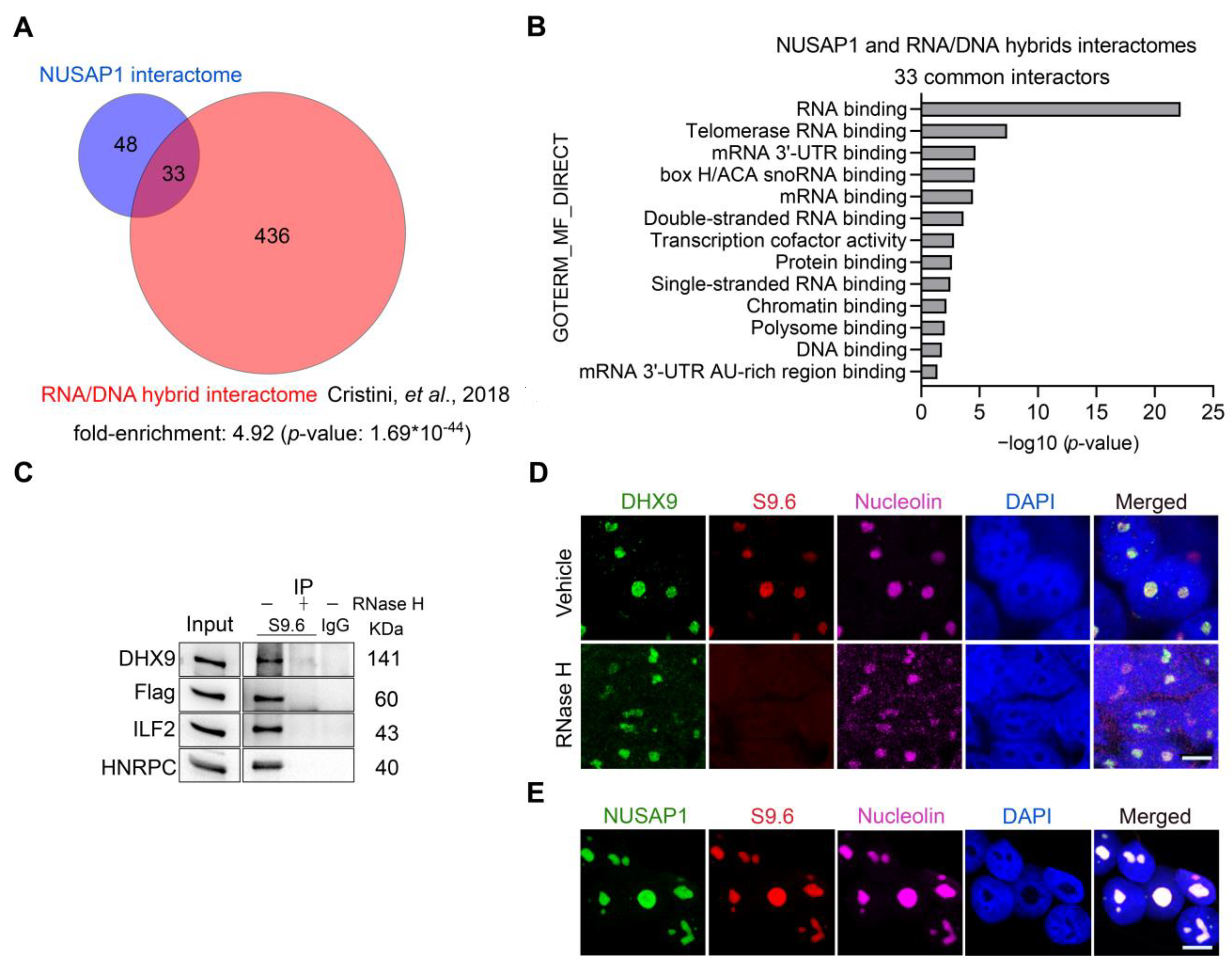
2.3. NUSAP1, ILF2 and DHX9 Associate with R-Loops
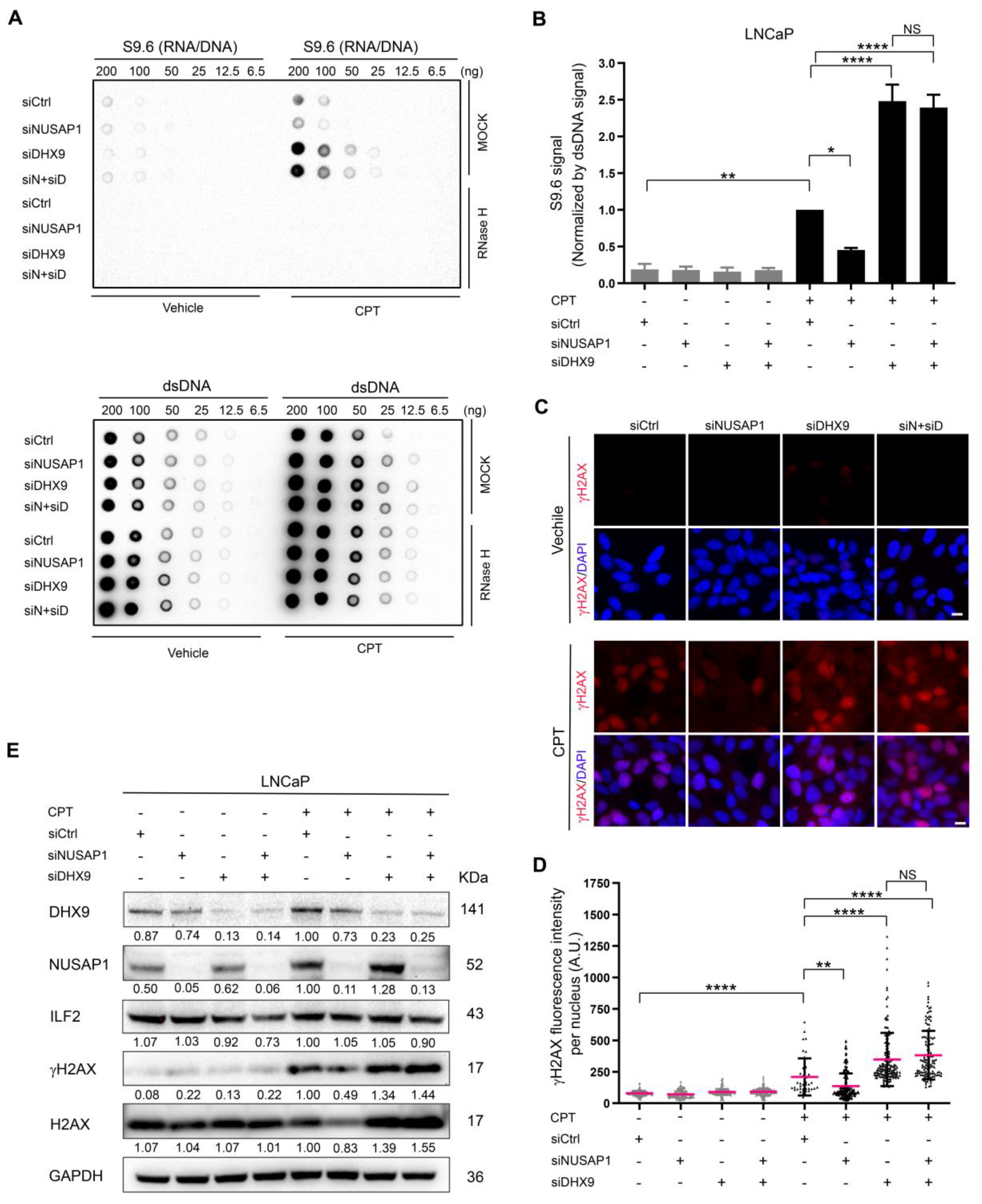
2.4. NUSAP1 Depletion Reduces CPT-Induced R-Loop Accumulation and DNA Damage but Does Not Compensate for DHX9 Depletion
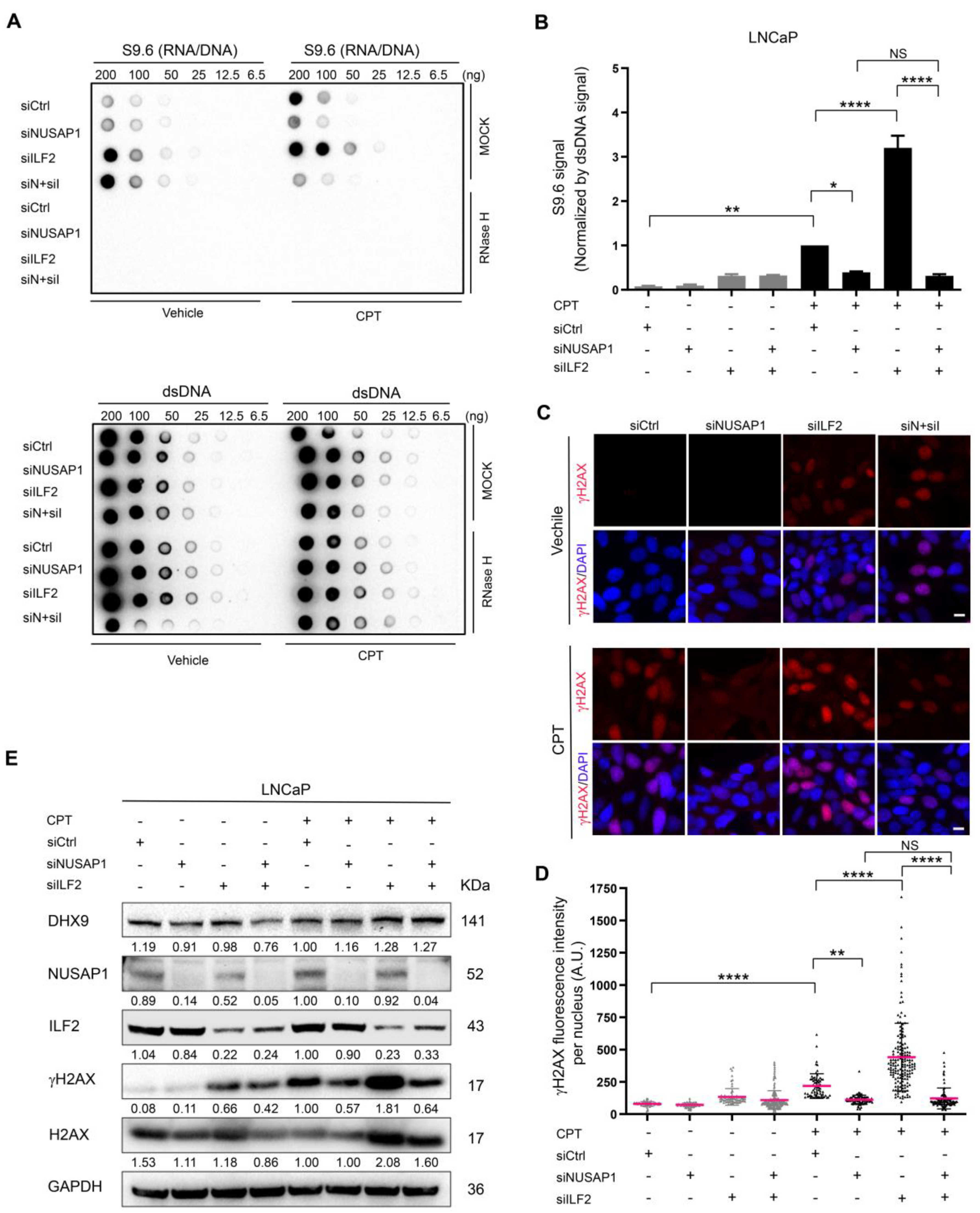
2.5. ILF2 Depletion Increases R-Loop Accumulation and DNA Damage That Is Abolished by Depletion of NUSAP1
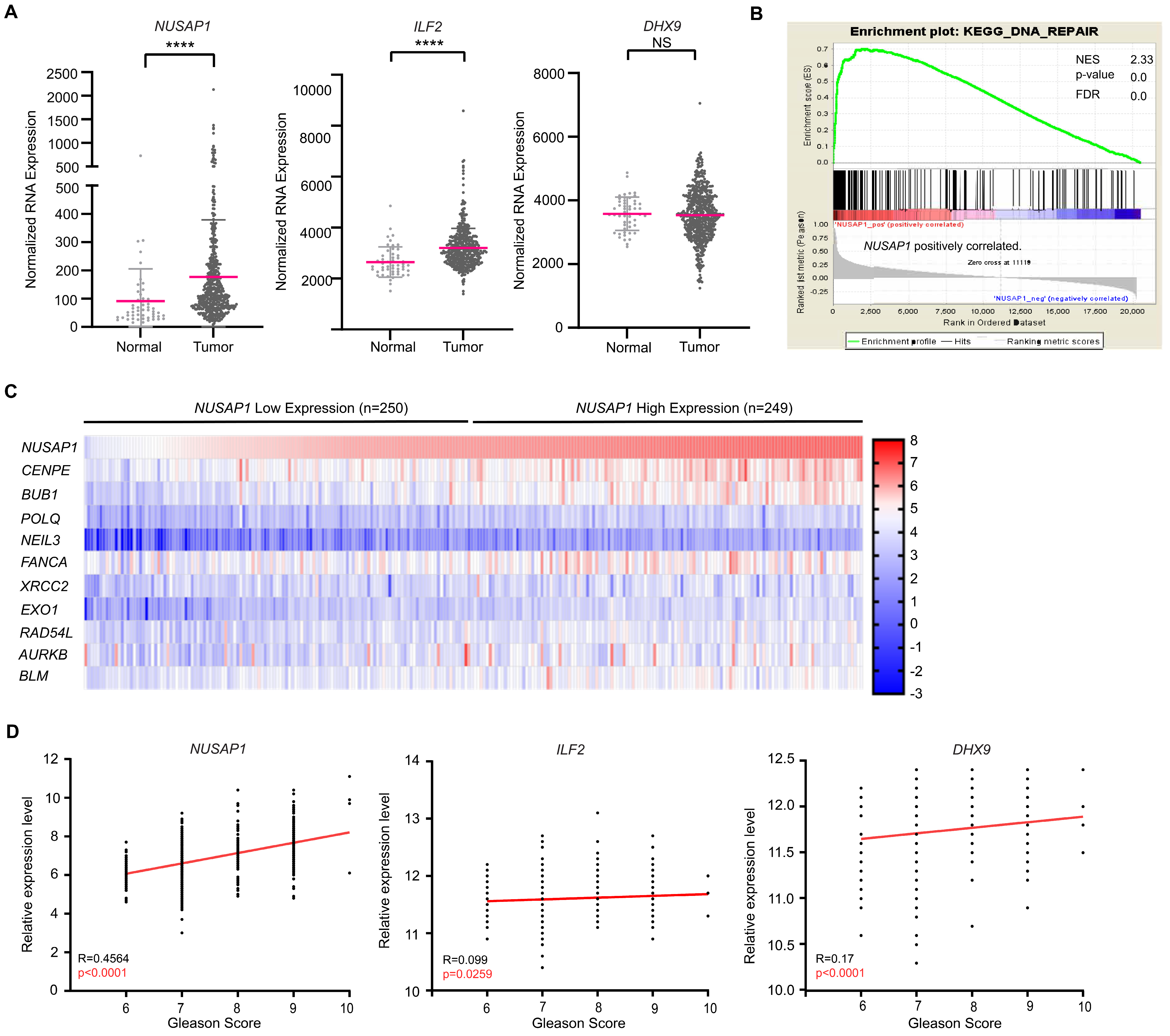
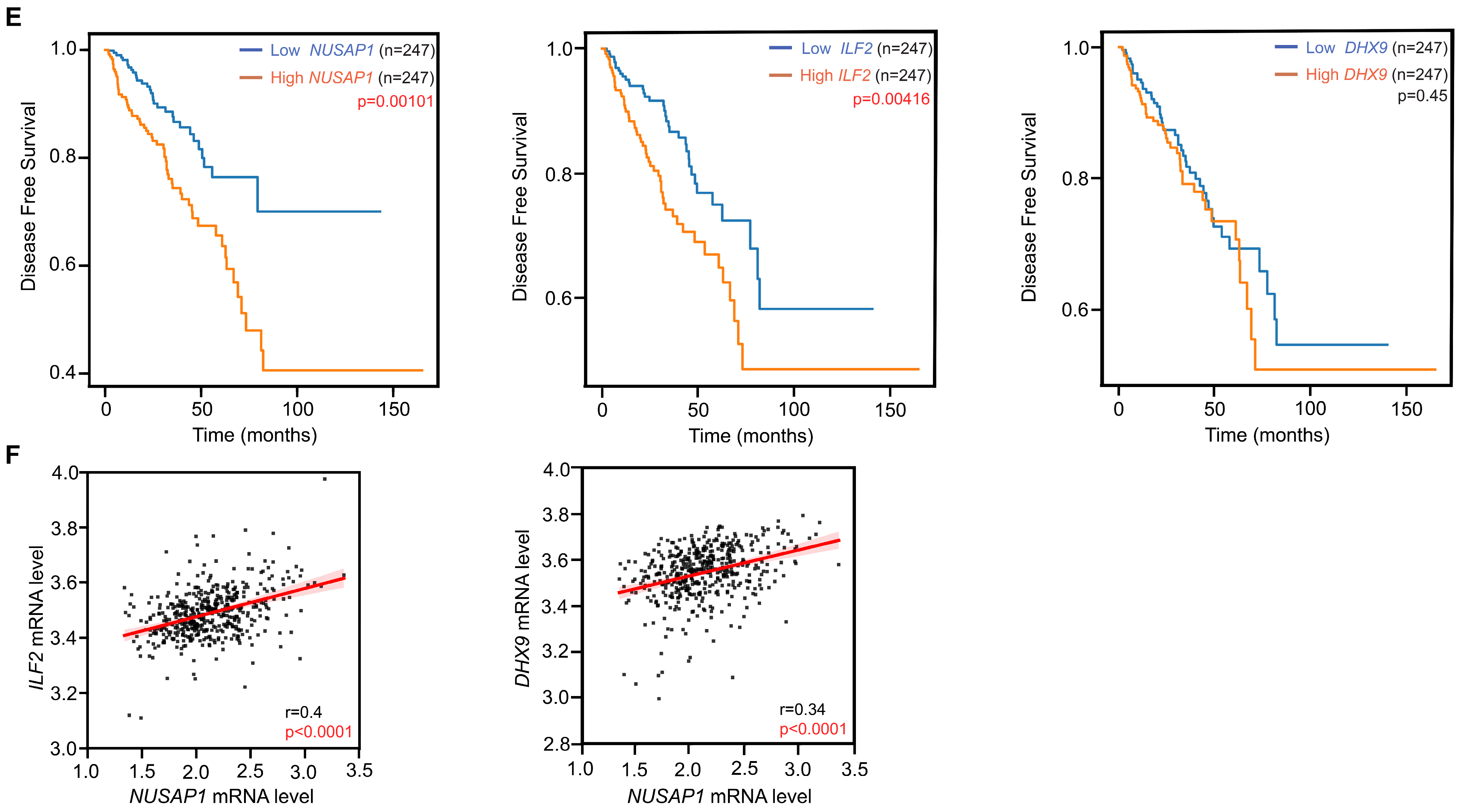
2.6. High NUSAP1 Expression Correlated with DNA Repair Pathways and Poor Clinical Outcome in Prostate Adenocarcinoma
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. siRNA
4.3. Plasmids
4.4. Transient Transfections and Drug Treatment
4.5. Immunoprecipitation for Mass Spectrometry Analyses
4.6. Mass Spectrometry (MS)
4.7. Immunoprecipitation
4.8. Immunofluorescence Staining and Image Analyses
4.9. Protein Lysate Preparation and Immunoblotting
4.10. R-Loop Immunoprecipitation
4.11. R-Loop Analysis by Dot-Blot
4.12. The Cancer Genome Atlas (TCGA) Data Analysis
4.13. Gene Set Enrichment Analysis (GSEA)
4.14. Quantification and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Gulzar, Z.G.; McKenney, J.K.; Brooks, J.D. Increased expression of NuSAP in recurrent prostate cancer is mediated by E2F1. Oncogene 2013, 32, 70–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, T.; Wei, D.; Li, Q.; Yang, X.; Han, Y.; Luo, Y.; Jiang, Y. Four Novel Prognostic Genes Related to Prostate Cancer Identified Using Co-expression Structure Network Analysis. Front. Genet. 2021, 12, 584164. [Google Scholar] [CrossRef]
- Cooperberg, M.R.; Simko, J.P.; Cowan, J.E.; Reid, J.E.; Djalilvand, A.; Bhatnagar, S.; Gutin, A.; Lanchbury, J.S.; Swanson, G.P.; Stone, S.; et al. Validation of a cell-cycle progression gene panel to improve risk stratification in a contemporary prostatectomy cohort. J. Clin. Oncol. 2013, 31, 1428–1434. [Google Scholar] [CrossRef]
- Cuzick, J.; Swanson, G.P.; Fisher, G.; Brothman, A.R.; Berney, D.M.; Reid, J.E.; Mesher, D.; Speights, V.O.; Stankiewicz, E.; Foster, C.S.; et al. Prognostic value of an RNA expression signature derived from cell cycle proliferation genes in patients with prostate cancer: A retrospective study. Lancet Oncol. 2011, 12, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Ryu, B.; Kim, D.S.; Deluca, A.M.; Alani, R.M. Comprehensive expression profiling of tumor cell lines identifies molecular signatures of melanoma progression. PLoS ONE 2007, 2, e594. [Google Scholar] [CrossRef] [Green Version]
- Bogunovic, D.; O’Neill, D.W.; Belitskaya-Levy, I.; Vacic, V.; Yu, Y.L.; Adams, S.; Darvishian, F.; Berman, R.; Shapiro, R.; Pavlick, A.C.; et al. Immune profile and mitotic index of metastatic melanoma lesions enhance clinical staging in predicting patient survival. Proc. Natl. Acad. Sci. USA 2009, 106, 20429–20434. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Shi, C.; Liu, S.; Zhang, E.; Yan, L.; Ji, C.; Zhao, Y. Overexpression of NuSAP1 is predictive of an unfavourable prognosis and promotes proliferation and invasion of triple-negative breast cancer cells via the Wnt/beta-catenin/EMT signalling axis. Gene 2020, 747, 144657. [Google Scholar] [CrossRef]
- Chen, L.; Yang, L.; Qiao, F.; Hu, X.; Li, S.; Yao, L.; Yang, X.L.; Shao, Z.M. High Levels of Nucleolar Spindle-Associated Protein and Reduced Levels of BRCA1 Expression Predict Poor Prognosis in Triple-Negative Breast Cancer. PLoS ONE 2015, 10, e0140572. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Zhou, L.; Chen, J.; Su, J.; Shen, W.; Liu, B.; Zhou, J.; Yu, S.; Qian, J. A four-gene signature for prognosis in breast cancer patients with hypermethylated IL15RA. Oncol. Lett. 2019, 17, 4245–4254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colak, D.; Nofal, A.; Albakheet, A.; Nirmal, M.; Jeprel, H.; Eldali, A.; Al-Tweigeri, T.; Tulbah, A.; Ajarim, D.; Malik, O.A.; et al. Age-specific gene expression signatures for breast tumors and cross-species conserved potential cancer progression markers in young women. PLoS ONE 2013, 8, e63204. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Q.; Butler, W.; Zhou, Y.; Zhang, H.; Tang, L.; Perkinson, K.; Chen, X.; Jiang, X.S.; McCall, S.J.; Inman, B.A.; et al. Pre-existing Castration-resistant Prostate Cancer-like Cells in Primary Prostate Cancer Promote Resistance to Hormonal Therapy. Eur. Urol. 2022, 81, 446–455. [Google Scholar] [CrossRef]
- Xie, Q.; Ou-Yang, W.; Zhang, M.; Wang, H.; Yue, Q. Decreased Expression of NUSAP1 Predicts Poor Overall Survival in Cervical Cancer. J. Cancer 2020, 11, 2852–2863. [Google Scholar] [CrossRef]
- Espinosa, A.M.; Alfaro, A.; Roman-Basaure, E.; Guardado-Estrada, M.; Palma, I.; Serralde, C.; Medina, I.; Juarez, E.; Bermudez, M.; Marquez, E.; et al. Mitosis is a source of potential markers for screening and survival and therapeutic targets in cervical cancer. PLoS ONE 2013, 8, e55975. [Google Scholar] [CrossRef]
- Qian, Z.; Li, Y.; Ma, J.; Xue, Y.; Xi, Y.; Hong, L.; Dai, X.; Zhang, Y.; Ji, X.; Chen, Y.; et al. Prognostic value of NUSAP1 in progression and expansion of glioblastoma multiforme. J. Neurooncol. 2018, 140, 199–208. [Google Scholar] [CrossRef]
- Zhu, T.; Xie, P.; Gao, Y.F.; Huang, M.S.; Li, X.; Zhang, W.; Zhou, H.H.; Liu, Z.Q. Nucleolar and spindle-associated protein 1 is a tumor grade correlated prognosis marker for glioma patients. CNS Neurosci. Ther. 2018, 24, 178–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Xu, B.; Yang, C.; Wang, W.; Zhong, D.; Zhao, Z.; He, L.; Hu, Y.; Jiang, L.; Li, J.; et al. Nucleolar and spindle associated protein 1 promotes the aggressiveness of astrocytoma by activating the Hedgehog signaling pathway. J. Exp. Clin. Cancer Res. 2017, 36, 127. [Google Scholar] [CrossRef] [Green Version]
- Dai, Q.; Liu, T.; Gao, Y.; Zhou, H.; Li, X.; Zhang, W. Six genes involved in prognosis of hepatocellular carcinoma identified by Cox hazard regression. BMC Bioinform. 2021, 22, 167. [Google Scholar] [CrossRef]
- Chen, H.; Wu, J.; Lu, L.; Hu, Z.; Li, X.; Huang, L.; Zhang, X.; Chen, M.; Qin, X.; Xie, L. Identification of Hub Genes Associated With Immune Infiltration and Predict Prognosis in Hepatocellular Carcinoma via Bioinformatics Approaches. Front. Genet. 2020, 11, 575762. [Google Scholar] [CrossRef]
- Liu, L.; Chen, A.; Chen, S.; Song, W.; Yao, Q.; Wang, P.; Zhou, S. CCNB2, NUSAP1 and TK1 are associated with the prognosis and progression of hepatocellular carcinoma, as revealed by co-expression analysis. Exp. Ther. Med. 2020, 19, 2679–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.E.; Shang, X.; Nie, K.C.; Xu, Q.; Chen, N.B.; Zhu, Z.Z. Identification of potential crucial genes associated with the pathogenesis and prognosis of pancreatic adenocarcinoma. Oncol. Lett. 2020, 20, 60. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Ma, H.; Hao, J.; Xie, Q.; Zhao, R. MCM2 and NUSAP1 Are Potential Biomarkers for the Diagnosis and Prognosis of Pancreatic Cancer. Biomed. Res. Int. 2020, 2020, 8604340. [Google Scholar] [CrossRef]
- Guo, H.; Zou, J.; Zhou, L.; Zhong, M.; He, Y.; Huang, S.; Chen, J.; Li, J.; Xiong, J.; Fang, Z.; et al. NUSAP1 Promotes Gastric Cancer Tumorigenesis and Progression by Stabilizing the YAP1 Protein. Front. Oncol. 2020, 10, 591698. [Google Scholar] [CrossRef]
- Ge, Y.; Li, Q.; Lin, L.; Jiang, M.; Shi, L.; Wang, B.; Yang, L.; Xu, Z. Downregulation of NUSAP1 suppresses cell proliferation, migration, and invasion via inhibiting mTORC1 signalling pathway in gastric cancer. Cell. Biochem. Funct. 2020, 38, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, K.; Cai, H.; Chen, S.; Sun, D.; Jiang, P. The role of nucleolar spindle-associated protein 1 in human ovarian cancer. J. Cell. Biochem. 2020, 121, 4397–4405. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Yu, S.; Sun, X.; Yin, M.; Fei, J.; Zhou, J. Identification of key biomarkers associated with development and prognosis in patients with ovarian carcinoma: Evidence from bioinformatic analysis. J. Ovarian Res. 2019, 12, 110. [Google Scholar] [CrossRef] [Green Version]
- Ling, B.; Wei, P.; Xiao, J.; Cen, B.; Wei, H.; Feng, X.; Ye, G.; Li, S.; Zhang, Z.; Liang, W.; et al. Nucleolar and spindleassociated protein 1 promotes nonsmall cell lung cancer progression and serves as an effector of myocyte enhancer factor 2D. Oncol. Rep. 2021, 45, 1044–1058. [Google Scholar] [CrossRef]
- Zeng, H.; Ji, J.; Song, X.; Huang, Y.; Li, H.; Huang, J.; Ma, X. Stemness Related Genes Revealed by Network Analysis Associated With Tumor Immune Microenvironment and the Clinical Outcome in Lung Adenocarcinoma. Front. Genet. 2020, 11, 549213. [Google Scholar] [CrossRef] [PubMed]
- Verbakel, W.; Carmeliet, G.; Engelborghs, Y. SAP-like domain in nucleolar spindle associated protein mediates mitotic chromosome loading as well as interphase chromatin interaction. Biochem. Biophys. Res. Commun. 2011, 411, 732–737. [Google Scholar] [CrossRef]
- Raemaekers, T.; Ribbeck, K.; Beaudouin, J.; Annaert, W.; Van Camp, M.; Stockmans, I.; Smets, N.; Bouillon, R.; Ellenberg, J.; Carmeliet, G. NuSAP, a novel microtubule-associated protein involved in mitotic spindle organization. J. Cell. Biol. 2003, 162, 1017–1029. [Google Scholar] [CrossRef]
- Gordon, C.A.; Gulzar, Z.G.; Brooks, J.D. NUSAP1 expression is upregulated by loss of RB1 in prostate cancer cells. Prostate 2015, 75, 517–526. [Google Scholar] [CrossRef]
- Gordon, C.A.; Gong, X.; Ganesh, D.; Brooks, J.D. NUSAP1 promotes invasion and metastasis of prostate cancer. Oncotarget 2017, 8, 29935–29950. [Google Scholar] [CrossRef] [Green Version]
- Kotian, S.; Banerjee, T.; Lockhart, A.; Huang, K.; Catalyurek, U.V.; Parvin, J.D. NUSAP1 influences the DNA damage response by controlling BRCA1 protein levels. Cancer Biol. Ther. 2014, 15, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; He, J.; Li, Y.; Lv, S.; Cui, H. NUSAP1 potentiates chemoresistance in glioblastoma through its SAP domain to stabilize ATR. Signal. Transduct. Target. Ther. 2020, 5, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Zhang, W.; Yan, M.; Qiu, J.; Chen, J.; Sun, X.; Chen, X.; Song, L.; Zhang, Y. Nucleolar and spindle associated protein 1 promotes metastasis of cervical carcinoma cells by activating Wnt/beta-catenin signaling. J. Exp. Clin. Cancer Res. 2019, 38, 33. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Liu, H.; Gong, Y.; Zhang, B.; Chen, W. ANKRD22 enhances breast cancer cell malignancy by activating the Wnt/beta-catenin pathway via modulating NuSAP1 expression. Bosn. J. Basic. Med. Sci. 2021, 21, 294–304. [Google Scholar] [CrossRef]
- Li, X.; Fu, Y.; Xia, X.; Zhang, X.; Xiao, K.; Zhuang, X.; Zhang, Y. Knockdown of SP1/Syncytin1 axis inhibits the proliferation and metastasis through the AKT and ERK1/2 signaling pathways in non-small cell lung cancer. Cancer Med. 2019, 8, 5750–5759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blajeski, A.L.; Phan, V.A.; Kottke, T.J.; Kaufmann, S.H. G(1) and G(2) cell-cycle arrest following microtubule depolymerization in human breast cancer cells. J. Clin. Investig. 2002, 110, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.; Larsen, B.; Lin, Z.Y.; Breitkreutz, A.; Mellacheruvu, D.; Fermin, D.; Qin, Z.S.; Tyers, M.; Gingras, A.C.; Nesvizhskii, A.I. SAINT: Probabilistic scoring of affinity purification-mass spectrometry data. Nat. Methods 2011, 8, 70–73. [Google Scholar] [CrossRef]
- Marchesini, M.; Ogoti, Y.; Fiorini, E.; Aktas Samur, A.; Nezi, L.; D’Anca, M.; Storti, P.; Samur, M.K.; Ganan-Gomez, I.; Fulciniti, M.T.; et al. ILF2 Is a Regulator of RNA Splicing and DNA Damage Response in 1q21-Amplified Multiple Myeloma. Cancer Cell. 2017, 32, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Cristini, A.; Groh, M.; Kristiansen, M.S.; Gromak, N. RNA/DNA Hybrid Interactome Identifies DXH9 as a Molecular Player in Transcriptional Termination and R-Loop-Associated DNA Damage. Cell. Rep. 2018, 23, 1891–1905. [Google Scholar] [CrossRef] [Green Version]
- Brickner, J.R.; Garzon, J.L.; Cimprich, K.A. Walking a tightrope: The complex balancing act of R-loops in genome stability. Mol. Cell. 2022, 82, 2267–2297. [Google Scholar] [CrossRef]
- Nadel, J.; Athanasiadou, R.; Lemetre, C.; Wijetunga, N.A.; Broin, P.Ó.; Sato, H.; Zhang, Z.; Jeddeloh, J.; Montagna, C.; Golden, A.; et al. RNA:DNA hybrids in the human genome have distinctive nucleotide characteristics, chromatin composition, and transcriptional relationships. Epigenetics Chromatin 2015, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos-Pereira, J.M.; Aguilera, A. R loops: New modulators of genome dynamics and function. Nat. Rev. Genet. 2015, 16, 583–597. [Google Scholar] [CrossRef]
- Crossley, M.P.; Bocek, M.; Cimprich, K.A. R-Loops as Cellular Regulators and Genomic Threats. Mol. Cell. 2019, 73, 398–411. [Google Scholar] [CrossRef] [Green Version]
- Taylor, B.S.; Schultz, N.; Hieronymus, H.; Gopalan, A.; Xiao, Y.; Carver, B.S.; Arora, V.K.; Kaushik, P.; Cerami, E.; Reva, B.; et al. Integrative genomic profiling of human prostate cancer. Cancer Cell. 2010, 18, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapointe, J.; Li, C.; Giacomini, C.P.; Salari, K.; Huang, S.; Wang, P.; Ferrari, M.; Hernandez-Boussard, T.; Brooks, J.D.; Pollack, J.R. Genomic profiling reveals alternative genetic pathways of prostate tumorigenesis. Cancer Res. 2007, 67, 8504–8510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- The Cancer Genome Atlas Research Network. The Molecular Taxonomy of Primary Prostate Cancer. Cell 2015, 163, 1011–1025. [Google Scholar] [CrossRef] [Green Version]
- Toubiana, S.; Selig, S. DNA:RNA hybrids at telomeres—When it is better to be out of the (R) loop. FEBS J. 2018, 285, 2552–2566. [Google Scholar] [CrossRef] [Green Version]
- Marinello, J.; Bertoncini, S.; Aloisi, I.; Cristini, A.; Malagoli Tagliazucchi, G.; Forcato, M.; Sordet, O.; Capranico, G. Dynamic Effects of Topoisomerase I Inhibition on R-Loops and Short Transcripts at Active Promoters. PLoS ONE 2016, 11, e0147053. [Google Scholar] [CrossRef] [Green Version]
- Sordet, O.; Redon, C.E.; Guirouilh-Barbat, J.; Smith, S.; Solier, S.; Douarre, C.; Conti, C.; Nakamura, A.J.; Das, B.B.; Nicolas, E.; et al. Ataxia telangiectasia mutated activation by transcription- and topoisomerase I-induced DNA double-strand breaks. EMBO Rep. 2009, 10, 887–893. [Google Scholar] [CrossRef] [Green Version]
- Dou, P.; Li, Y.; Sun, H.; Xie, W.; Zhang, X.; Zhang, X.; Zhang, D.; Qiao, S.; Ci, Y.; Nie, H.; et al. C1orf109L binding DHX9 promotes DNA damage depended on the R-loop accumulation and enhances camptothecin chemosensitivity. Cell. Prolif. 2020, 53, e12875. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.; Grosse, F. Human DHX9 helicase preferentially unwinds RNA-containing displacement loops (R-loops) and G-quadruplexes. DNA Repair. (Amst) 2011, 10, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Bustos, M.A.; Gross, R.; Ramos, R.I.; Takeshima, T.L.; Mills, G.B.; Yu, Q.; Hoon, D.S.B. Interleukin enhancer-binding factor 2 promotes cell proliferation and DNA damage response in metastatic melanoma. Clin. Transl. Med. 2021, 11, e608. [Google Scholar] [CrossRef] [PubMed]
- Haley, B.; Paunesku, T.; Protic, M.; Woloschak, G.E. Response of heterogeneous ribonuclear proteins (hnRNP) to ionising radiation and their involvement in DNA damage repair. Int. J. Radiat. Biol. 2009, 85, 643–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anantha, R.W.; Alcivar, A.L.; Ma, J.; Cai, H.; Simhadri, S.; Ule, J.; Konig, J.; Xia, B. Requirement of heterogeneous nuclear ribonucleoprotein C for BRCA gene expression and homologous recombination. PLoS ONE 2013, 8, e61368. [Google Scholar] [CrossRef] [Green Version]
- Salvi, J.S.; Mekhail, K. R-loops highlight the nucleus in ALS. Nucleus 2015, 6, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, M.; Toth, R.; Vandermoere, F.; Morrice, N.A.; Rouse, J. Identification and characterization of FUS/TLS as a new target of ATM. Biochem. J. 2008, 415, 297–307. [Google Scholar] [CrossRef] [Green Version]
- Rulten, S.L.; Rotheray, A.; Green, R.L.; Grundy, G.J.; Moore, D.A.; Gomez-Herreros, F.; Hafezparast, M.; Caldecott, K.W. PARP-1 dependent recruitment of the amyotrophic lateral sclerosis-associated protein FUS/TLS to sites of oxidative DNA damage. Nucleic Acids Res. 2014, 42, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Adamson, B.; Smogorzewska, A.; Sigoillot, F.D.; King, R.W.; Elledge, S.J. A genome-wide homologous recombination screen identifies the RNA-binding protein RBMX as a component of the DNA-damage response. Nat. Cell. Biol. 2012, 14, 318–328. [Google Scholar] [CrossRef] [Green Version]
- Zheng, T.; Zhou, H.; Li, X.; Peng, D.; Yang, Y.; Zeng, Y.; Liu, H.; Ren, J.; Zhao, Y. RBMX is required for activation of ATR on repetitive DNAs to maintain genome stability. Cell. Death Differ. 2020, 27, 3162–3176. [Google Scholar] [CrossRef]
- Groh, M.; Gromak, N. Out of balance: R-loops in human disease. PLoS Genet. 2014, 10, e1004630. [Google Scholar] [CrossRef] [Green Version]
- Drews, R.M.; Hernando, B.; Tarabichi, M.; Haase, K.; Lesluyes, T.; Smith, P.S.; Morrill Gavarro, L.; Couturier, D.L.; Liu, L.; Schneider, M.; et al. A pan-cancer compendium of chromosomal instability. Nature 2022, 606, 976–983. [Google Scholar] [CrossRef]
- Ribbeck, K.; Groen, A.C.; Santarella, R.; Bohnsack, M.T.; Raemaekers, T.; Kocher, T.; Gentzel, M.; Gorlich, D.; Wilm, M.; Carmeliet, G.; et al. NuSAP, a mitotic RanGTP target that stabilizes and cross-links microtubules. Mol. Biol. Cell. 2006, 17, 2646–2660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribbeck, K.; Raemaekers, T.; Carmeliet, G.; Mattaj, I.W. A role for NuSAP in linking microtubules to mitotic chromosomes. Curr. Biol. 2007, 17, 230–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, P.; Li, L.; Xing, G.; Tian, C.; Yin, Y.; He, F.; Zhang, L. ATM-mediated NuSAP phosphorylation induces mitotic arrest. Biochem. Biophys. Res. Commun. 2011, 404, 413–418. [Google Scholar] [CrossRef] [PubMed]
- McKenney, J.K.; Wei, W.; Hawley, S.; Auman, H.; Newcomb, L.F.; Boyer, H.D.; Fazli, L.; Simko, J.; Hurtado-Coll, A.; Troyer, D.A.; et al. Histologic Grading of Prostatic Adenocarcinoma Can Be Further Optimized: Analysis of the Relative Prognostic Strength of Individual Architectural Patterns in 1275 Patients From the Canary Retrospective Cohort. Am. J. Surg. Pathol. 2016, 40, 1439–1456. [Google Scholar] [CrossRef]
- Stumpf, C.R.; Ruggero, D. The cancerous translation apparatus. Curr. Opin. Genet. Dev. 2011, 21, 474–483. [Google Scholar] [CrossRef] [Green Version]
- Li, C.G.; Mahon, C.; Sweeney, N.M.; Verschueren, E.; Kantamani, V.; Li, D.; Hennigs, J.K.; Marciano, D.P.; Diebold, I.; Abu-Halawa, O.; et al. PPARgamma Interaction with UBR5/ATMIN Promotes DNA Repair to Maintain Endothelial Homeostasis. Cell. Rep. 2019, 26, 1333–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Chiu, C.L.; Lo, E.; Lee, Y.J.; Yamada, S.; Lo, S.H. Hyperactivity of Mek in TNS1 knockouts leads to potential treatments for cystic kidney diseases. Cell. Death Dis. 2019, 10, 871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez, P.; Crouch, R.J.; Cheung, V.G.; Grunseich, C. R-Loop Analysis by Dot-Blot. J. Vis. Exp. 2021, 167. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstrale, M.; Laurila, E.; et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Garcia-Gutierrez, P.; Garcia-Dominguez, M. BETting on a Transcriptional Deficit as the Main Cause for Cornelia de Lange Syndrome. Front. Mol. Biosci. 2021, 8, 709232. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, C.-L.; Li, C.G.; Verschueren, E.; Wen, R.M.; Zhang, D.; Gordon, C.A.; Zhao, H.; Giaccia, A.J.; Brooks, J.D. NUSAP1 Binds ILF2 to Modulate R-Loop Accumulation and DNA Damage in Prostate Cancer. Int. J. Mol. Sci. 2023, 24, 6258. https://doi.org/10.3390/ijms24076258
Chiu C-L, Li CG, Verschueren E, Wen RM, Zhang D, Gordon CA, Zhao H, Giaccia AJ, Brooks JD. NUSAP1 Binds ILF2 to Modulate R-Loop Accumulation and DNA Damage in Prostate Cancer. International Journal of Molecular Sciences. 2023; 24(7):6258. https://doi.org/10.3390/ijms24076258
Chicago/Turabian StyleChiu, Chun-Lung, Caiyun G. Li, Erik Verschueren, Ru M. Wen, Dalin Zhang, Catherine A. Gordon, Hongjuan Zhao, Amato J. Giaccia, and James D. Brooks. 2023. "NUSAP1 Binds ILF2 to Modulate R-Loop Accumulation and DNA Damage in Prostate Cancer" International Journal of Molecular Sciences 24, no. 7: 6258. https://doi.org/10.3390/ijms24076258