
Physiological Benefits of Novel Selenium Delivery via Nanoparticles

{kind=link}
{kind=link}
Abstract
:1. Introduction of Selenium
1.1. Selenoproteins
1.2. Chemical Forms of Selenium
2. Selenium Nanoparticles
2.1. Mechanisms of Bioactivity for Selenium Nanoparticles
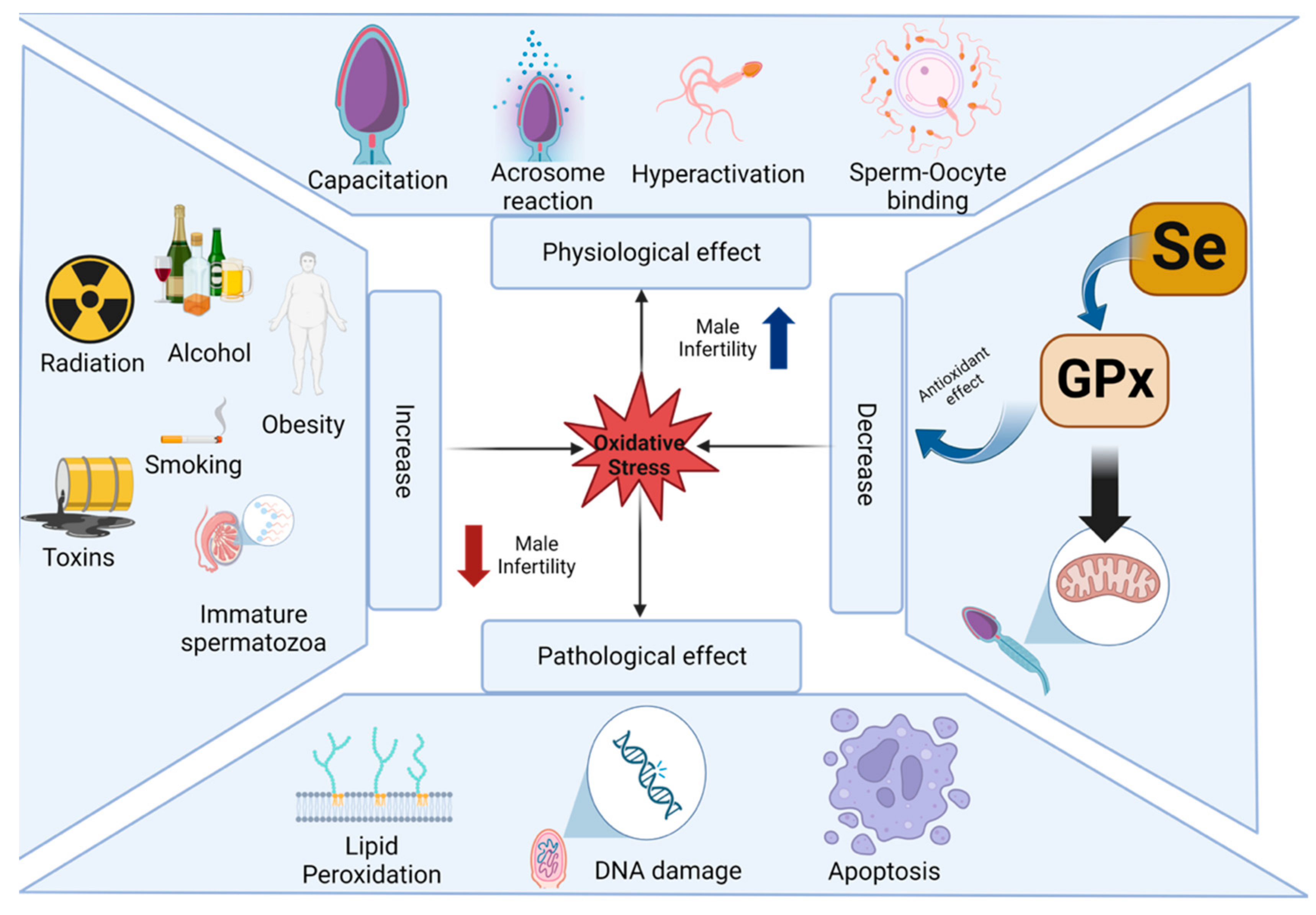
2.2. Impacts of Selenium Nanoparticles on Fertility
2.3. Impacts of Selenium Nanoparticles on Growth
2.4. Impacts of Selenium Nanoparticles on Diseases and Human Health
2.5. Synthesis of Selenium Nanoparticles
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Mojadadi, A.; Au, A.; Salah, W.; Witting, P.; Ahmad, G. Role for Selenium in Metabolic Homeostasis and Human Reproduction. Nutrients 2021, 13, 3256. [Google Scholar] [CrossRef] [PubMed]
- Fordyce, F.M. Selenium deficiency and toxicity in the environment. In Essentials of Medical Geology; Springer: Berlin/Heidelberg, Germany, 2013; pp. 375–416. [Google Scholar]
- Mal, J.; Veneman, W.J.; Nancharaiah, Y.V.; van Hullebusch, E.D.; Peijnenburg, W.J.G.M.; Vijver, M.G.; Lens, P.N.L. A comparison of fate and toxicity of selenite, biogenically, and chemically synthesized selenium nanoparticles to zebrafish (Danio rerio) embryogenesis. Nanotoxicology 2017, 11, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krysiak, R.; Kowalcze, K.; Okopień, B. Selenomethionine potentiates the impact of vitamin D on thyroid autoimmunity in euthyroid women with Hashimoto’s thyroiditis and low vitamin D status. Pharmacol. Rep. 2019, 71, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, X.; Pedram, P.; Shahidi, M.; Du, J.; Yi, Y.; Gulliver, W.; Zhang, H.; Sun, G. Significant Beneficial Association of High Dietary Selenium Intake with Reduced Body Fat in the CODING Study. Nutrients 2016, 8, 24. [Google Scholar] [CrossRef]
- Winther, K.H.; Rayman, M.P.; Bonnema, S.J.; Hegedüs, L. Selenium in thyroid—Essential knowledge for clinicians. Nat. Rev. Endocrinol. 2020, 16, 165–176. [Google Scholar] [CrossRef]
- Mutonhodza, B.; Joy, E.J.M.; Bailey, E.H.; Lark, M.R.; Kangara, M.G.M.; Broadley, M.R.; Matsungo, T.M.; Chopera, P. Linkages between soil, crop, livestock, and human selenium status in Sub-Saharan Africa: A scoping review. Int. J. Food Sci. Technol. 2022, 57, 6336–6349. [Google Scholar] [CrossRef]
- Wang, N.; Tan, H.-Y.; Li, S.; Xu, Y.; Guo, W.; Feng, Y. Supplementation of Micronutrient Selenium in Metabolic Diseases: Its Role as an Antioxidant. Oxidative Med. Cell. Longev. 2017, 2017, 7478523. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of Mammalian Selenoproteomes. Science 2003, 300, 1439. [Google Scholar] [CrossRef] [Green Version]
- Fontenelle, L.C.; Feitosa, M.M.; Freitas, T.E.C.; Severo, J.S.; Morais, J.B.S.; Henriques, G.S.; Oliveira, F.E.; Moita Neto, J.M.; Marreiro, D.d.N. Selenium status and its relationship with thyroid hormones in obese women. Clin. Nutr. Eur. Soc. Clin. Nutr. Metab. 2021, 41, 398–404. [Google Scholar] [CrossRef]
- Santos, A.d.C.; Passos, A.F.F.; Holzbach, L.C.; Cominetti, C. Selenium Intake and Glycemic Control in Young Adults with Normal-Weight Obesity Syndrome. Front. Nutr. 2021, 8, 696325. [Google Scholar] [CrossRef]
- Dhillon, V.S.; Deo, P.; Fenech, M. Plasma Micronutrient Profile of Prostate Cancer Cases Is Altered Relative to Healthy Controls-Results of a Pilot Study in South Australia. Cancers 2022, 15, 77. [Google Scholar] [CrossRef]
- Steevens, J.; van den Brandt, P.A.; Goldbohm, R.A.; Schouten, L.J. Selenium status and the risk of esophageal and gastric cancer subtypes: The Netherlands cohort study. Gastroenterology 2010, 138, 1704–1713. [Google Scholar] [CrossRef]
- Xue, G.; Liu, R. Association between dietary selenium intake and bone mineral density in the US general population. Ann. Transl. Med. 2022, 10, 869. [Google Scholar] [CrossRef]
- Kang, D.; Lee, J.; Wu, C.; Guo, X.; Lee, B.J.; Chun, J.-S.; Kim, J.-H. The role of selenium metabolism and selenoproteins in cartilage homeostasis and arthropathies. Exp. Mol. Med. 2020, 52, 1198–1208. [Google Scholar] [CrossRef]
- Avery, J.C.; Hoffmann, P.R. Selenium, Selenoproteins, and Immunity. Nutrients 2018, 10, 1203. [Google Scholar] [CrossRef] [Green Version]
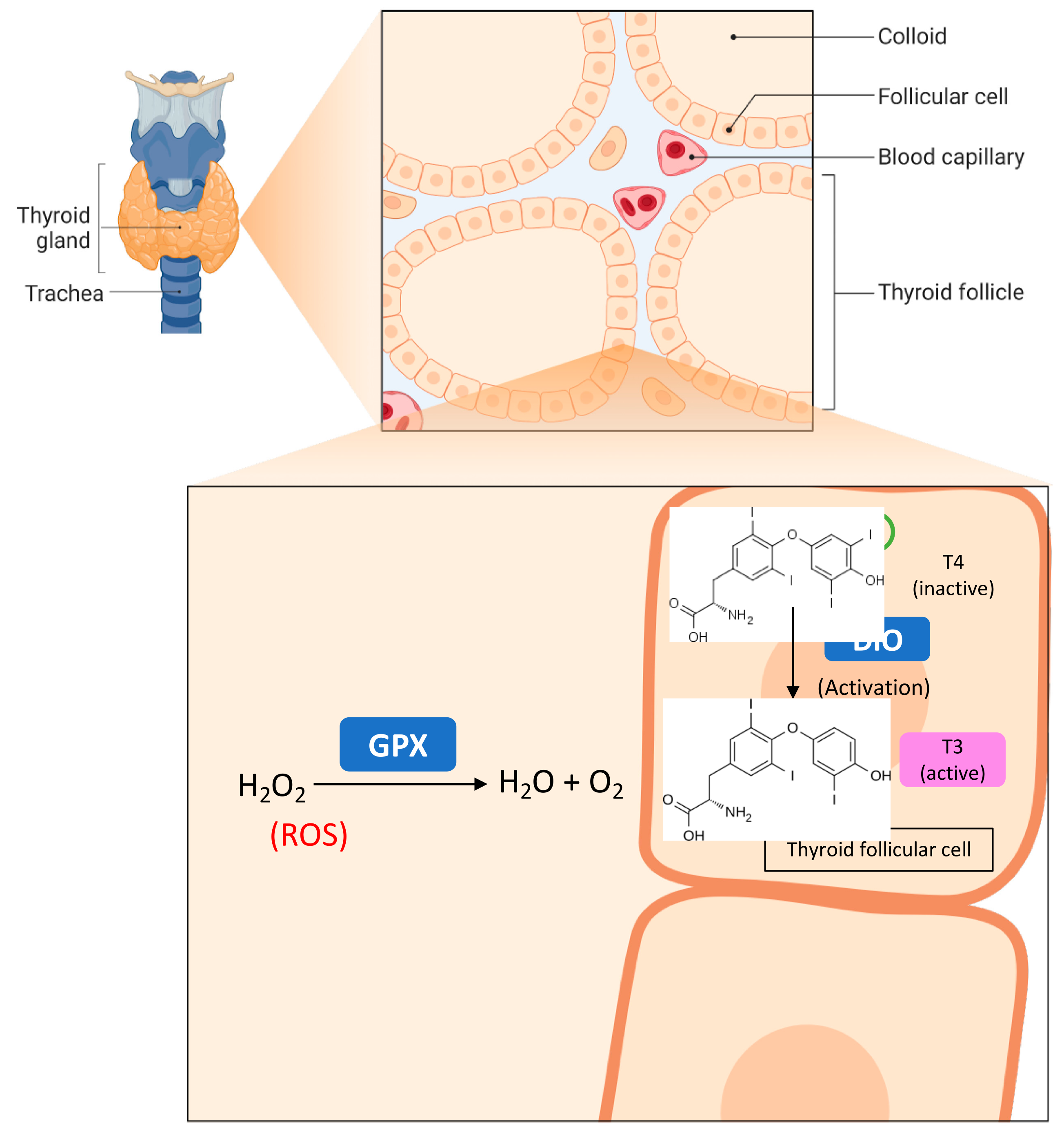
- Miot, F.; Dupuy, C.; Dumont, J.; Rousset, B. Chapter 2 Thyroid Hormone Synthesis and Secretion. In Endotext; MDText. com, Inc.: South Dartmouth, MA, USA, 2000; pp. 2000–2015. [Google Scholar]
- Yang, X.; Tang, Z.; Li, J.; Jiang, J. Esophagus cancer and essential trace elements. Front. Public Health 2022, 10, 1038153. [Google Scholar] [CrossRef]
- Cai, X.; Wang, C.; Yu, W.; Fan, W.; Wang, S.; Shen, N.; Wu, P.; Li, X.; Wang, F. Selenium Exposure and Cancer Risk: An Updated Meta-analysis and Meta-regression. Sci. Rep. 2016, 6, 19213. [Google Scholar] [CrossRef] [Green Version]
- Weekley, C.M.; Jeong, G.; Tierney, M.E.; Hossain, F.; Maw, A.M.; Shanu, A.; Harris, H.H.; Witting, P.K. Selenite-mediated production of superoxide radical anions in A549 cancer cells is accompanied by a selective increase in SOD1 concentration, enhanced apoptosis and Se–Cu bonding. JBIC J. Biol. Inorg. Chem. 2014, 19, 813–828. [Google Scholar] [CrossRef]
- Vinceti, M.; Filippini, T.; Del Giovane, C.; Dennert, G.; Zwahlen, M.; Brinkman, M.; Zeegers, M.P.; Horneber, M.; D’Amico, R.; Crespi, C.M. Selenium for preventing cancer. Cochrane Database Syst. Rev. 2018, 1, Cd005195. [Google Scholar] [CrossRef]
- An, P.; Wan, S.; Luo, Y.; Luo, J.; Zhang, X.; Zhou, S.; Xu, T.; He, J.; Mechanick, J.I.; Wu, W.C.; et al. Micronutrient Supplementation to Reduce Cardiovascular Risk. J. Am. Coll. Cardiol. 2022, 80, 2269–2285. [Google Scholar] [CrossRef]
- Qiao, L.; Zhang, X.; Pi, S.; Chang, J.; Dou, X.; Yan, S.; Song, X.; Chen, Y.; Zeng, X.; Zhu, L.; et al. Dietary supplementation with biogenic selenium nanoparticles alleviate oxidative stress-induced intestinal barrier dysfunction. NPJ Sci. Food 2022, 6, 30. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Wu, H.; Jiang, K.; Xu, Y.; Miao, Z.; Wang, H.; Ma, Y. Zero-Valence Selenium-Enriched Prussian Blue Nanozymes Reconstruct Intestinal Barrier against Inflammatory Bowel Disease via Inhibiting Ferroptosis and T Cells Differentiation. Adv. Healthc. Mater. 2023, e2203160. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Kou, F.; Wang, J.; Quan, Z.; Zhao, S.; Wang, Y.; Hu, X.; Sun, H.; Cao, L. Pretreatment with millet-derived selenylated soluble dietary fiber ameliorates dextran sulfate sodium-induced colitis in mice by regulating inflammation and maintaining gut microbiota balance. Front. Nutr. 2022, 9, 928601. [Google Scholar] [CrossRef] [PubMed]
- Amerikanou, C.; Karavoltsos, S.; Gioxari, A.; Tagkouli, D.; Sakellari, A.; Papada, E.; Kalogeropoulos, N.; Forbes, A.; Kaliora, A.C. Clinical and inflammatory biomarkers of inflammatory bowel diseases are linked to plasma trace elements and toxic metals; new insights into an old concept. Front. Nutr. 2022, 9, 997356. [Google Scholar] [CrossRef]
- Chen, J.; Ruan, X.; Yuan, S.; Deng, M.; Zhang, H.; Sun, J.; Yu, L.; Satsangi, J.; Larsson, S.C.; Therdoratou, E.; et al. Antioxidants, minerals and vitamins in relation to Crohn’s disease and ulcerative colitis: A Mendelian randomization study. Aliment. Pharmacol. Ther. 2023, 57, 399–408. [Google Scholar] [CrossRef]
- Alizadeh, M.; Safaeiyan, A.; Ostadrahimi, A.; Estakhri, R.; Daneghian, S.; Ghaffari, A.; Gargari, B.P. Effect of L-arginine and selenium added to a hypocaloric diet enriched with legumes on cardiovascular disease risk factors in women with central obesity: A randomized, double-blind, placebo-controlled trial. Ann. Nutr. Metab. 2012, 60, 157–168. [Google Scholar] [CrossRef]
- Mahmoodianfard, S.; Vafa, M.; Golgiri, F.; Khoshniat, M.; Gohari, M.; Solati, Z.; Djalali, M. Effects of Zinc and Selenium Supplementation on Thyroid Function in Overweight and Obese Hypothyroid Female Patients: A Randomized Double-Blind Controlled Trial. J. Am. Coll. Nutr. 2015, 34, 391–399. [Google Scholar] [CrossRef]
- Weening, E.H.; Al-Mubarak, A.A.; Dokter, M.M.; Dickstein, K.; Lang, C.C.; Ng, L.L.; Metra, M.; van Veldhuisen, D.J.; Touw, D.J.; de Boer, R.A.; et al. Sexual dimorphism in selenium deficiency is associated with metabolic syndrome and prevalence of heart disease. Cardiovasc. Diabetol. 2023, 22, 8. [Google Scholar] [CrossRef]
- Thompson, H.J.; Sedlacek, S.M.; Fitzgerald, V.K.; Wolfe, P.; McGinley, J.N. A Randomized, Double-Blind, Placebo-Controlled Investigation of Selenium Supplementation in Women at Elevated Risk for Breast Cancer: Lessons for Re-Emergent Interest in Selenium and Cancer. Biomedicines 2023, 11, 49. [Google Scholar] [CrossRef]
- Arablou, T.; Aryaeian, N.; Djalali, M.; Shahram, F.; Rasouli, L. Association between dietary intake of some antioxidant micronutrients with some inflammatory and antioxidant markers in active Rheumatoid Arthritis patients. Int. J. Vitam. Nutr. Res. 2019, 89, 238–245. [Google Scholar] [CrossRef]
- Hejazi, J.; Mohtadinia, J.; Kolahi, S.; Bakhtiyari, M.; Delpisheh, A. Nutritional Status of Iranian Women with Rheumatoid Arthritis: An Assessment of Dietary Intake and Disease Activity. Women’s Health 2011, 7, 599–605. [Google Scholar] [CrossRef]
- Qu, Z.; Yang, F.; Hong, J.; Wang, W.; Li, S.; Jiang, G.; Yan, S. Causal relationship of serum nutritional factors with osteoarthritis: A Mendelian randomization study. Rheumatology 2020, 60, 2383–2390. [Google Scholar] [CrossRef]
- Guan, T.; Wu, Z.; Xu, C.; Su, G. The association of trace elements with arthritis in US adults: NHANES 2013–2016. J. Trace Elem. Med. Biol. 2023, 76, 127122. [Google Scholar] [CrossRef]
- Cavedon, E.; Manso, J.; Negro, I.; Censi, S.; Serra, R.; Busetto, L.; Vettor, R.; Plebani, M.; Pezzani, R.; Nacamulli, D.; et al. Selenium Supplementation, Body Mass Composition, and Leptin Levels in Patients with Obesity on a Balanced Mildly Hypocaloric Diet: A Pilot Study. Int. J. Endocrinol. 2020, 2020, 7. [Google Scholar] [CrossRef]
- Verma, A.K.; Kumar, A.; Rahal, A.; Kumar, V.; Roy, D. Inorganic versus organic selenium supplementation: A review. Pak. J. Biol. Sci. 2012, 15, 418–425. [Google Scholar]
- Weekley, C.M.; Harris, H.H. Which form is that? The importance of selenium speciation and metabolism in the prevention and treatment of disease. Chem. Soc. Rev. 2013, 42, 8870–8894. [Google Scholar] [CrossRef]
- Weekley, C.M.; Shanu, A.; Aitken, J.B.; Vogt, S.; Witting, P.K.; Harris, H.H. XAS and XFM studies of selenium and copper speciation and distribution in the kidneys of selenite-supplemented rats. Metallomics 2014, 6, 1602–1615. [Google Scholar] [CrossRef]
- Weekley, C.M.; Aitken, J.B.; Witting, P.K.; Harris, H.H. XAS studies of Se speciation in selenite-fed rats. Metallomics 2014, 6, 2193–2203. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, D.; Kishawy, A.T.Y.; Khater, S.I.; Hamed Arisha, A.; Mohammed, H.A.; Abdelaziz, A.S.; Abd El-Rahman, G.I.; Elabbasy, M.T. Effect of Dietary Modulation of Selenium Form and Level on Performance, Tissue Retention, Quality of Frozen Stored Meat and Gene Expression of Antioxidant Status in Ross Broiler Chickens. Animals 2019, 9, 342. [Google Scholar] [CrossRef] [Green Version]
- Mazokopakis, E. Differentiated thyroid cancer and selenium supplements for protection of salivary glands from (131)I treatment. Hell. J. Nucl. Med. 2018, 21, 83–84. [Google Scholar] [CrossRef]
- Sun, C.; Du, Z.; Liu, X.; Yang, Y.; Zhou, S.; Li, C.; Cao, X.; Zhao, Q.; Wong, K.; Chen, W.; et al. Selenium Forms and Dosages Determined Their Biological Actions in Mouse Models of Parkinson’s Disease. Nutrients 2022, 15, 11. [Google Scholar] [CrossRef]
- Sang, L.-X.; Chang, B.; Zhu, J.-F.; Yang, F.-L.; Li, Y.; Jiang, X.-F.; Wang, D.-N.; Lu, C.-L.; Sun, X. Sodium selenite ameliorates dextran sulfate sodium-induced chronic colitis in mice by decreasing Th1, Th17, and γδT and increasing CD4(+)CD25(+) regulatory T-cell responses. World J. Gastroenterol. 2017, 23, 3850–3863. [Google Scholar] [CrossRef] [PubMed]
- Husbeck, B.; Nonn, L.; Peehl, D.M.; Knox, S.J. Tumor-selective killing by selenite in patient-matched pairs of normal and malignant prostate cells. Prostate 2006, 66, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Casaril, A.M.; Domingues, M.; Lourenço, D.D.A.; Vieira, B.; Begnini, K.; Corcini, C.D.; França, R.T.; Varela Junior, A.S.; Seixas, F.K.; Collares, T.; et al. 3-[(4-chlorophenyl)selanyl]-1-methyl-1H-indole ameliorates long-lasting depression- and anxiogenic-like behaviors and cognitive impairment in post-septic mice: Involvement of neuroimmune and oxidative hallmarks. Chem.-Biol. Interact. 2020, 331, 109278. [Google Scholar] [CrossRef] [PubMed]
- Casaril, A.M.; Domingues, M.; Bampi, S.R.; de Andrade Lourenço, D.; Padilha, N.B.; Lenardão, E.J.; Sonego, M.; Seixas, F.K.; Collares, T.; Nogueira, C.W.; et al. The selenium-containing compound 3-((4-chlorophenyl)selanyl)-1-methyl-1H-indole reverses depressive-like behavior induced by acute restraint stress in mice: Modulation of oxido-nitrosative stress and inflammatory pathway. Psychopharmacology 2019, 236, 2867–2880. [Google Scholar] [CrossRef]
- Casaril, A.M.; Domingues, M.; Bampi, S.R.; Lourenço, D.D.A.; Smaniotto, T.Â.; Segatto, N.; Vieira, B.; Seixas, F.K.; Collares, T.; Lenardão, E.J.; et al. The antioxidant and immunomodulatory compound 3-[(4-chlorophenyl)selanyl]-1-methyl-1H-indole attenuates depression-like behavior and cognitive impairment developed in a mouse model of breast tumor. Brain Behav. Immun. 2020, 84, 229–241. [Google Scholar] [CrossRef]
- Casaril, A.M.; Lourenço, D.D.A.; Domingues, M.; Smaniotto, T.Â.; Birmann, P.T.; Vieira, B.; Sonego, M.S.; Seixas, F.K.; Collares, T.; Lenardão, E.J.; et al. Anhedonic- and anxiogenic-like behaviors and neurochemical alterations are abolished by a single administration of a selenium-containing compound in chronically stressed mice. Compr. Psychoneuroendocrinol. 2021, 6, 100054. [Google Scholar] [CrossRef]
- Abdelnour, S.A.; Alagawany, M.; Hashem, N.M.; Farag, M.R.; Alghamdi, E.S.; Hassan, F.U.; Bilal, R.M.; Elnesr, S.S.; Dawood, M.A.O.; Nagadi, S.A.; et al. Nanominerals: Fabrication Methods, Benefits and Hazards, and Their Applications in Ruminants with Special Reference to Selenium and Zinc Nanoparticles. Animals 2021, 11, 1916. [Google Scholar] [CrossRef]
- Ghaniem, S.; Nassef, E.; Zaineldin, A.I.; Bakr, A.; Hegazi, S. A Comparison of the Beneficial Effects of Inorganic, Organic, and Elemental Nano-selenium on Nile Tilapia: Growth, Immunity, Oxidative Status, Gut Morphology, and Immune Gene Expression. Biol. Trace Elem. Res. 2022, 200, 5226–5241. [Google Scholar] [CrossRef]
- Rana, T. Nano-selenium on reproduction and immunocompetence: An emerging progress and prospect in the productivity of poultry research. Trop. Anim. Health Prod. 2021, 53, 324. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, J.; Yu, H. Elemental selenium at nano size possesses lower toxicity without compromising the fundamental effect on selenoenzymes: Comparison with selenomethionine in mice. Free. Radic. Biol. Med. 2007, 42, 1524–1533. [Google Scholar] [CrossRef]
- Zhang, J.S.; Gao, X.Y.; Zhang, L.D.; Bao, Y.P. Biological effects of a nano red elemental selenium. Biofactors 2001, 15, 27–38. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, H.; Yan, X.; Zhang, L. Comparison of short-term toxicity between Nano-Se and selenite in mice. Life Sci. 2005, 76, 1099–1109. [Google Scholar] [CrossRef]
- Bień, D.; Michalczuk, M.; Szkopek, D.; Kinsner, M.; Konieczka, P. Changes in lipids metabolism indices as a result of different form of selenium supplementation in chickens. Sci. Rep. 2022, 12, 13817. [Google Scholar] [CrossRef]
- Kumar, N.; Krishnani, K.K.; Singh, N.P. Comparative study of selenium and selenium nanoparticles with reference to acute toxicity, biochemical attributes, and histopathological response in fish. Environ. Sci. Pollut. Res. 2018, 25, 8914–8927. [Google Scholar] [CrossRef]
- Li, H.; Zhang, J.; Wang, T.; Luo, W.; Zhou, Q.; Jiang, G. Elemental selenium particles at nano-size (Nano-Se) are more toxic to Medaka (Oryzias latipes) as a consequence of hyper-accumulation of selenium: A comparison with sodium selenite. Aquat. Toxicol. 2008, 89, 251–256. [Google Scholar] [CrossRef]
- Bhattacharjee, A.; Basu, A.; Bhattacharya, S. Selenium nanoparticles are less toxic than inorganic and organic selenium to mice in vivo. Nucleus 2019, 62, 259–268. [Google Scholar] [CrossRef]
- Tugarova, A.V.; Kamnev, A.A. Proteins in microbial synthesis of selenium nanoparticles. Talanta 2017, 174, 539–547. [Google Scholar] [CrossRef]
- Nazıroğlu, M.; Muhamad, S.; Pecze, L. Nanoparticles as potential clinical therapeutic agents in Alzheimer’s disease: Focus on selenium nanoparticles. Expert Rev. Clin. Pharmacol. 2017, 10, 773–782. [Google Scholar] [CrossRef]
- Amin, K.A.; Hashem, K.S.; Alshehri, F.S.; Awad, S.T.; Hassan, M.S. Antioxidant and Hepatoprotective Efficiency of Selenium Nanoparticles Against Acetaminophen-Induced Hepatic Damage. Biol. Trace Elem. Res. 2017, 175, 136–145. [Google Scholar] [CrossRef]
- Huang, X.; Chen, X.; Chen, Q.; Yu, Q.; Sun, D.; Liu, J. Investigation of functional selenium nanoparticles as potent antimicrobial agents against superbugs. Acta Biomater. 2016, 30, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Huo, D.; Zhu, C.; Shen, S.; Wang, W.; Li, H.; Zhu, Z.; Xia, Y. Combination cancer treatment through photothermally controlled release of selenous acid from gold nanocages. Biomaterials 2018, 178, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Qu, W.; Yang, J.; Sun, Z.; Zhang, R.; Zhou, F.; Zhang, K.; Xia, Y.; Huang, K.; Miao, D. Effect of Selenium Nanoparticles on Anti-Oxidative Level, Egg Production and Quality and Blood Parameter of Laying Hens Exposed to Deoxynivalenol. J. Anim. Res. Nutr. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Meng, T.; Liu, Y.-L.; Xie, C.-Y.; Zhang, B.; Huang, Y.-Q.; Zhang, Y.-W.; Yao, Y.; Huang, R.; Wu, X. Effects of Different Selenium Sources on Laying Performance, Egg Selenium Concentration, and Antioxidant Capacity in Laying Hens. Biol. Trace Elem. Res. 2019, 189, 548–555. [Google Scholar] [CrossRef]
- Alian, H.A.; Samy, H.M.; Ibrahim, M.T.; Mahmoud, M.M.A. Nanoselenium effect on growth performance, carcass traits, antioxidant activity, and immune status of broilers. Environ. Sci. Pollut. Res. 2020, 27, 38607–38616. [Google Scholar] [CrossRef]
- Li, Y.; Fan, M.; Qiu, Q.; Wang, Y.; Shen, X.; Zhao, K. Nano-selenium and Macleaya cordata Extracts Improved Immune Function and Reduced Oxidative Damage of Sows and IUGR Piglets After Heat Stress of Sows in Late Gestation. Biol. Trace Elem. Res. 2022, 200, 5081–5090. [Google Scholar] [CrossRef]
- Shokraneh, M.; Sadeghi, A.A.; Mousavi, S.N.; Esmaeilkhanian, S.; Chamani, M. Effects of in ovo injection of nano-selenium and nano-zinc oxide and high eggshell temperature during late incubation on antioxidant activity, thyroid and glucocorticoid hormones and some blood metabolites in broiler hatchlings. Acta Sci. Anim. Sci. 2020, 42. [Google Scholar] [CrossRef] [Green Version]
- Khalil, W.A.; El-Harairy, M.A.; Zeidan, A.E.B.; Hassan, M.A.E. Impact of selenium nano-particles in semen extender on bull sperm quality after cryopreservation. Theriogenology 2019, 126, 121–127. [Google Scholar] [CrossRef]
- El-Naby, A.-s.A.-H.H.; Ibrahim, S.; Hozyen, H.F.; Sosa, A.S.A.; Mahmoud, K.G.M.; Farghali, A.A. Impact of nano-selenium on nuclear maturation and genes expression profile of buffalo oocytes matured in vitro. Mol. Biol. Rep. 2020, 47, 8593–8603. [Google Scholar] [CrossRef]
- Dawit Moges, F.; Hamdi, H.; Al-Barty, A.; Zaid, A.A.; Sundaray, M.; Parashar, S.K.S.; Gubale, A.G.; Das, B. Effects of selenium nanoparticle on the growth performance and nutritional quality in Nile Tilapia, Oreochromis niloticus. PLoS ONE 2022, 17, e0268348. [Google Scholar] [CrossRef]
- Loeschner, K.; Hadrup, N.; Hansen, M.; Pereira, S.A.; Gammelgaard, B.; Møller, L.H.; Mortensen, A.; Lam, H.R.; Larsen, E.H. Absorption, distribution, metabolism and excretion of selenium following oral administration of elemental selenium nanoparticles or selenite in rats. Metallomics 2014, 6, 330–337. [Google Scholar] [CrossRef] [Green Version]
- Gangadoo, S.; Dinev, I.; Willson, N.-L.; Moore, R.J.; Chapman, J.; Stanley, D. Nanoparticles of selenium as high bioavailable and non-toxic supplement alternatives for broiler chickens. Environ. Sci. Pollut. Res. 2020, 27, 16159–16166. [Google Scholar] [CrossRef]
- Hu, C.H.; Li, Y.L.; Xiong, L.; Zhang, H.M.; Song, J.; Xia, M.S. Comparative effects of nano elemental selenium and sodium selenite on selenium retention in broiler chickens. Anim. Feed. Sci. Technol. 2012, 177, 204–210. [Google Scholar] [CrossRef]
- Rashad, M.M.; Galal, M.K.; Abou-El-Sherbini, K.S.; El-Behairy, A.M.; Gouda, E.M.; Moussa, S.Z. Nano-sized selenium attenuates the developmental testicular toxicity induced by di-n-butyl phthalate in pre-pubertal male rats. Biomed. Pharmacother. 2018, 107, 1754–1762. [Google Scholar] [CrossRef]
- Sharma, R.; Biedenharn, K.R.; Fedor, J.M.; Agarwal, A. Lifestyle factors and reproductive health: Taking control of your fertility. Reprod. Biol. Endocrinol. 2013, 11, 66. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; You, Y.; Yu, T.; Li, J. Insights into Modifiable Risk Factors of Infertility: A Mendelian Randomization Study. Nutrients 2022, 14, 4042. [Google Scholar] [CrossRef]
- Mohammed Ali, I.A.; AL-Ahmed, H.I.; Ben Ahmed, A. Evaluation of Green Synthesis (Withania somnifera) of Selenium Nanoparticles to Reduce Sperm DNA Fragmentation Diabetic Mice Induced with Streptozotocin. Appl. Sci. 2023, 13, 728. [Google Scholar] [CrossRef]
- Sánchez-Gutiérrez, M.; García-Montalvo, E.A.; Izquierdo-Vega, J.A.; Del Razo, L.M. Effect of dietary selenium deficiency on the in vitro fertilizing ability of mice spermatozoa. Cell Biol. Toxicol. 2008, 24, 321–329. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Liang, C.; Shen, L.; Zhang, Z.; Jiang, T.; Li, D.; Zou, W.; Wang, J.; Zong, K.; Liang, D.; et al. The association between essential trace element (copper, zinc, selenium, and cobalt) status and the risk of early embryonic arrest among women undergoing assisted reproductive techniques. Front. Endocrinol. 2022, 13, 906849. [Google Scholar] [CrossRef]
- Li, D.; Jiang, T.; Wang, X.; Yin, T.; Shen, L.; Zhang, Z.; Zou, W.; Liu, Y.; Zong, K.; Liang, D.; et al. Serum Essential Trace Element Status in Women and the Risk of Endometrial Diseases: A Case-Control Study: Serum Essential Trace Element Status in Women and the Risk of Endometrial Diseases: A Case-Control Study. Biol. Trace Elem. Res. 2022, 201, 2151–2161. [Google Scholar] [CrossRef]
- Abdel-Halim, B.R.; Helmy, N.A. Effect of nano-selenium and nano-zinc particles during in vitro maturation on the developmental competence of bovine oocytes. Anim. Prod. Sci. 2018, 58, 2021–2028. [Google Scholar] [CrossRef]
- Eini, F.; Kutenaei, M.A.; Shirzeyli, M.H.; Dastjerdi, Z.S.; Omidi, M.; Novin, M.G. Normal seminal plasma could preserve human spermatozoa against cryopreservation damages in Oligozoospermic patients. BMC Mol. Cell Biol. 2021, 22, 50. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ren, J.; Zhang, W.; Wang, B.; Ma, Y.; Su, L.; Dai, Y.; Liu, G. Glutathione and selenium nanoparticles have a synergistic protective effect during cryopreservation of bull semen. Front. Vet. Sci. 2023, 10, 1093274. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Singh, A.K. Impact of environmental factors on human semen quality and male fertility: A narrative review. Environ. Sci. Eur. 2022, 34, 6. [Google Scholar] [CrossRef]
- Tirmenstein, M.A.; Mangipudy, R. Aflatoxin. In Encyclopedia of Toxicology, 3rd ed.; Wexler, P., Ed.; Academic Press: Oxford, UK, 2014; pp. 104–106. [Google Scholar] [CrossRef]
- Asadpour, R.; Aliyoldashi, M.H.; Saberivand, A.; Hamidian, G.; Hejazi, M. Ameliorative effect of selenium nanoparticles on the structure and function of testis and in vitro embryo development in Aflatoxin B1-exposed male mice. Andrologia 2020, 52, e13824. [Google Scholar] [CrossRef]
- Zhang, X.; Gan, X.; E, Q.; Zhang, Q.; Ye, Y.; Cai, Y.; Han, A.; Tian, M.; Wang, Y.; Wang, C.; et al. Ameliorative effects of nano-selenium against NiSO(4)-induced apoptosis in rat testes. Toxicol. Mech. Methods 2019, 29, 467–477. [Google Scholar] [CrossRef]
- Eisa, C.S.; Mohammed, S.S.; El-Aziz, H.I.A.; Farghaly, L.M.; Hosny, S. The Effect of Selenium Nanoparticles versus Royal Jelly against Cisplatin-Induced Testicular Toxicity in Adult Male Albino Rats: A Light and Transmission Electron Microscopic Study. J. Microsc. Ultrastruct. 2022, 10, 180–196. [Google Scholar] [CrossRef]
- Rattan, S.; Zhou, C.; Chiang, C.; Mahalingam, S.; Brehm, E.; Flaws, J.A. Exposure to endocrine disruptors during adulthood: Consequences for female fertility. J. Endocrinol. 2017, 233, R109–R129. [Google Scholar] [CrossRef] [Green Version]
- Rodprasert, W.; Toppari, J.; Virtanen, H.E. Endocrine Disrupting Chemicals and Reproductive Health in Boys and Men. Front. Endocrinol. 2021, 12, 706532. [Google Scholar] [CrossRef]
- Khalaf, A.A.; Ahmed, W.; Moselhy, W.A.; Abdel-Halim, B.R.; Ibrahim, M.A. Protective effects of selenium and nano-selenium on bisphenol-induced reproductive toxicity in male rats. Hum. Exp. Toxicol. 2019, 38, 398–408. [Google Scholar] [CrossRef]
- Malyugina, S.; Skalickova, S.; Skladanka, J.; Slama, P.; Horky, P. Biogenic Selenium Nanoparticles in Animal Nutrition: A Review. Agriculture 2021, 11, 1244. [Google Scholar] [CrossRef]
- Tidwell, J.H.; Allan, G.L. Fish as food: Aquaculture’s contribution. Ecological and economic impacts and contributions of fish farming and capture fisheries. EMBO Rep. 2001, 2, 958–963. [Google Scholar] [CrossRef] [Green Version]
- Boyd, C.E.; McNevin, A.A.; Davis, R.P. The contribution of fisheries and aquaculture to the global protein supply. Food Secur. 2022, 14, 805–827. [Google Scholar] [CrossRef]
- Ibrahim, D.; Arisha, A.H.; Khater, S.I.; Gad, W.M.; Hassan, Z.; Abou-Khadra, S.H.; Mohamed, D.I.; Ahmed Ismail, T.; Gad, S.A.; Eid, S.A.M.; et al. Impact of Omega-3 Fatty Acids Nano-Formulation on Growth, Antioxidant Potential, Fillet Quality, Immunity, Autophagy-Related Genes and Aeromonas hydrophila Resistance in Nile Tilapia (Oreochromis niloticus). Antioxidants 2022, 11, 1523. [Google Scholar] [CrossRef]
- Kasaikina, M.V.; Kravtsova, M.A.; Lee, B.C.; Seravalli, J.; Peterson, D.A.; Walter, J.; Legge, R.; Benson, A.K.; Hatfield, D.L.; Gladyshev, V.N. Dietary selenium affects host selenoproteome expression by influencing the gut microbiota. FASEB J. 2011, 25, 2492–2499. [Google Scholar] [CrossRef] [Green Version]
- Werder, E.J.; Engel, L.S.; Curry, M.D.; Sandler, D.P. Selenium modifies associations between multiple metals and neurologic symptoms in Gulf states residents. Environ. Epidemiol. 2020, 4, e115. [Google Scholar] [CrossRef]
- Badgar, K.; Prokisch, J. Testing Toxicity and Antidote Effect of Selenium Nanoparticles with Paramecium caudatum. Open J. Anim. Sci. 2021, 11, 532–542. [Google Scholar] [CrossRef]
- Gangadoo, S.; Dinev, I.; Chapman, J.; Hughes, R.J.; Van, T.T.H.; Moore, R.J.; Stanley, D. Selenium nanoparticles in poultry feed modify gut microbiota and increase abundance of Faecalibacterium prausnitzii. Appl. Microbiol. Biotechnol. 2018, 102, 1455–1466. [Google Scholar] [CrossRef]
- Alfonso, S.; Gesto, M.; Sadoul, B. Temperature increase and its effects on fish stress physiology in the context of global warming. J. Fish Biol. 2021, 98, 1496–1508. [Google Scholar] [CrossRef]
- Li, L.; Liu, Z.; Quan, J.; Lu, J.; Zhao, G.; Sun, J. Metabonomics analysis reveals the protective effect of nano-selenium against heat stress of rainbow trout (Oncorhynchus mykiss). J. Proteom. 2022, 259, 104545. [Google Scholar] [CrossRef]
- Li, L.; Liu, Z.; Quan, J.; Sun, J.; Lu, J.; Zhao, G. Comprehensive proteomic analysis to elucidate the anti-heat stress effects of nano-selenium in rainbow trout (Oncorhynchus mykiss). Ecotoxicol. Environ. Saf. 2022, 241, 113736. [Google Scholar] [CrossRef] [PubMed]
- El-Saadony, M.T.; Saad, A.M.; Najjar, A.A.; Alzahrani, S.O.; Alkhatib, F.M.; Shafi, M.E.; Selem, E.; Desoky, E.-S.M.; Fouda, S.E.E.; El-Tahan, A.M.; et al. The use of biological selenium nanoparticles to suppress Triticum aestivum L. crown and root rot diseases induced by Fusarium species and improve yield under drought and heat stress. Saudi J. Biol. Sci. 2021, 28, 4461–4471. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Wu, Y.; Zhang, J.; An, Q.; Zhou, C.; Li, D.; Pan, C. Nano-selenium enhances the antioxidant capacity, organic acids and cucurbitacin B in melon (Cucumis melo L.) plants. Ecotoxicol. Environ. Saf. 2022, 241, 113777. [Google Scholar] [CrossRef] [PubMed]
- Dahlen, C.R.; Reynolds, L.P.; Caton, J.S. Selenium supplementation and pregnancy outcomes. Front. Nutr. 2022, 9, 1011850. [Google Scholar] [CrossRef]
- El-Azab, M.F.; Al-Karmalawy, A.A.; Antar, S.A.; Hanna, P.A.; Tawfik, K.M.; Hazem, R.M. A novel role of Nano selenium and sildenafil on streptozotocin-induced diabetic nephropathy in rats by modulation of inflammatory, oxidative, and apoptotic pathways. Life Sci. 2022, 303, 120691. [Google Scholar] [CrossRef]
- Jin, Y.; Cai, L.; Yang, Q.; Luo, Z.; Liang, L.; Liang, Y.; Wu, B.; Ding, L.; Zhang, D.; Xu, X.; et al. Anti-leukemia activities of selenium nanoparticles embedded in nanotube consisted of triple-helix β-d-glucan. Carbohydr. Polym. 2020, 240, 116329. [Google Scholar] [CrossRef]
- Abozaid, O.A.R.; Sallam, M.W.; El-Sonbaty, S.; Aziza, S.; Emad, B.; Ahmed, E.S.A. Resveratrol-Selenium Nanoparticles Alleviate Neuroinflammation and Neurotoxicity in a Rat Model of Alzheimer’s Disease by Regulating Sirt1/miRNA-134/GSK3β Expression. Biol. Trace Elem. Res. 2022, 200, 5104–5114. [Google Scholar] [CrossRef]
- Rastegar Moghaddam, S.H.; Hosseini, M.; Sabzi, F.; Hojjati Fard, F.; Marefati, N.; Beheshti, F.; Darroudi, M.; Ebrahimzadeh Bideskan, A.; Anaeigoudari, A. Cardiovascular protective effect of nano selenium in hypothyroid rats: Protection against oxidative stress and cardiac fibrosis. Clin. Exp. Hypertens. 2022, 44, 268–279. [Google Scholar] [CrossRef]
- Zheng, Z.; Liu, L.; Zhou, K.; Ding, L.; Zeng, J.; Zhang, W. Anti-Oxidant and Anti-Endothelial Dysfunctional Properties of Nano-Selenium in vitro and in vivo of Hyperhomocysteinemic Rats. Int. J. Nanomed. 2020, 15, 4501–4521. [Google Scholar] [CrossRef]
- Bai, K.; Hong, B.; He, J.; Hong, Z.; Tan, R. Preparation and antioxidant properties of selenium nanoparticles-loaded chitosan microspheres. Int. J. Nanomed. 2017, 12, 4527–4539. [Google Scholar] [CrossRef] [Green Version]
- Jia, X.; Liu, Q.; Zou, S.; Xu, X.; Zhang, L. Construction of selenium nanoparticles/β-glucan composites for enhancement of the antitumor activity. Carbohydr. Polym. 2015, 117, 434–442. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, X.H.; Hu, X.D.; Liu, P.D.; Zhang, H.Q. The effects of combined selenium nanoparticles and radiation therapy on breast cancer cells in vitro. Artif. Cells Nanomed. Biotechnol. 2018, 46, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.; Zeng, L.; Jiang, W.; Fu, Y.; Zheng, W.; Chen, T. Rational design of cancer-targeted selenium nanoparticles to antagonize multidrug resistance in cancer cells. Nanomedicine 2015, 11, 947–958. [Google Scholar] [CrossRef]
- Ali, H.F.H.; El-Sayed, N.M.; Khodeer, D.M.; Ahmed, A.A.M.; Hanna, P.A.; Moustafa, Y.M.A. Nano selenium ameliorates oxidative stress and inflammatory response associated with cypermethrin-induced neurotoxicity in rats. Ecotoxicol. Environ. Saf. 2020, 195, 110479. [Google Scholar] [CrossRef]
- El-Ghazaly, M.A.; Fadel, N.; Rashed, E.; El-Batal, A.; Kenawy, S.A. Anti-inflammatory effect of selenium nanoparticles on the inflammation induced in irradiated rats. Can. J. Physiol. Pharmacol. 2017, 95, 101–110. [Google Scholar] [CrossRef]
- Kieliszek, M.; Błażejak, S. Current Knowledge on the Importance of Selenium in Food for Living Organisms: A Review. Molecules 2016, 21, 609. [Google Scholar] [CrossRef] [Green Version]
- Dhawan, G.; Singh, I.; Dhawan, U.; Kumar, P. Synthesis and Characterization of Nanoselenium: A Step-by-Step Guide for Undergraduate Students. J. Chem. Educ. 2021, 98, 2982–2989. [Google Scholar] [CrossRef]
- Bronze-Uhle, E.S.; Costa, B.C.; Ximenes, V.F.; Lisboa-Filho, P.N. Synthetic nanoparticles of bovine serum albumin with entrapped salicylic acid. Nanotechnol. Sci. Appl. 2017, 10, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Gumustas, M.; Sengel-Turk, C.T.; Gumustas, A.; Ozkan, S.A.; Uslu, B. Chapter 5—Effect of Polymer-Based Nanoparticles on the Assay of Antimicrobial Drug Delivery Systems. In Multifunctional Systems for Combined Delivery, Biosensing and Diagnostics; Grumezescu, A.M., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 67–108. [Google Scholar] [CrossRef]
- Zhang, S.-Y.; Zhang, J.; Wang, H.-Y.; Chen, H.-Y. Synthesis of selenium nanoparticles in the presence of polysaccharides. Mater. Lett. 2004, 58, 2590–2594. [Google Scholar] [CrossRef]
- Shakibaie, M.; Forootanfar, H.; Golkari, Y.; Mohammadi-Khorsand, T.; Shakibaie, M.R. Anti-biofilm activity of biogenic selenium nanoparticles and selenium dioxide against clinical isolates of Staphylococcus aureus, Pseudomonas aeruginosa, and Proteus mirabilis. J. Trace Elem. Med. Biol. 2015, 29, 235–241. [Google Scholar] [CrossRef]
- Wang, G.; Guo, Y.; Yang, G.; Yang, L.; Ma, X.; Wang, K.; Zhu, L.; Sun, J.; Wang, X.; Zhang, H. Mitochondria-Mediated Protein Regulation Mechanism of Polymorphs-Dependent Inhibition of Nanoselenium on Cancer Cells. Sci. Rep. 2016, 6, 31427. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Au, A.; Mojadadi, A.; Shao, J.-Y.; Ahmad, G.; Witting, P.K. Physiological Benefits of Novel Selenium Delivery via Nanoparticles. Int. J. Mol. Sci. 2023, 24, 6068. https://doi.org/10.3390/ijms24076068
Au A, Mojadadi A, Shao J-Y, Ahmad G, Witting PK. Physiological Benefits of Novel Selenium Delivery via Nanoparticles. International Journal of Molecular Sciences. 2023; 24(7):6068. https://doi.org/10.3390/ijms24076068
Chicago/Turabian StyleAu, Alice, Albaraa Mojadadi, Jia-Ying Shao, Gulfam Ahmad, and Paul K. Witting. 2023. "Physiological Benefits of Novel Selenium Delivery via Nanoparticles" International Journal of Molecular Sciences 24, no. 7: 6068. https://doi.org/10.3390/ijms24076068