
LC-ESI-MS/MS Analysis of Sulfolipids and Galactolipids in Green and Red Lettuce (Lactuca sativa L.) as Influenced by Sulfur Nutrition


Abstract
:1. Introduction
2. Results
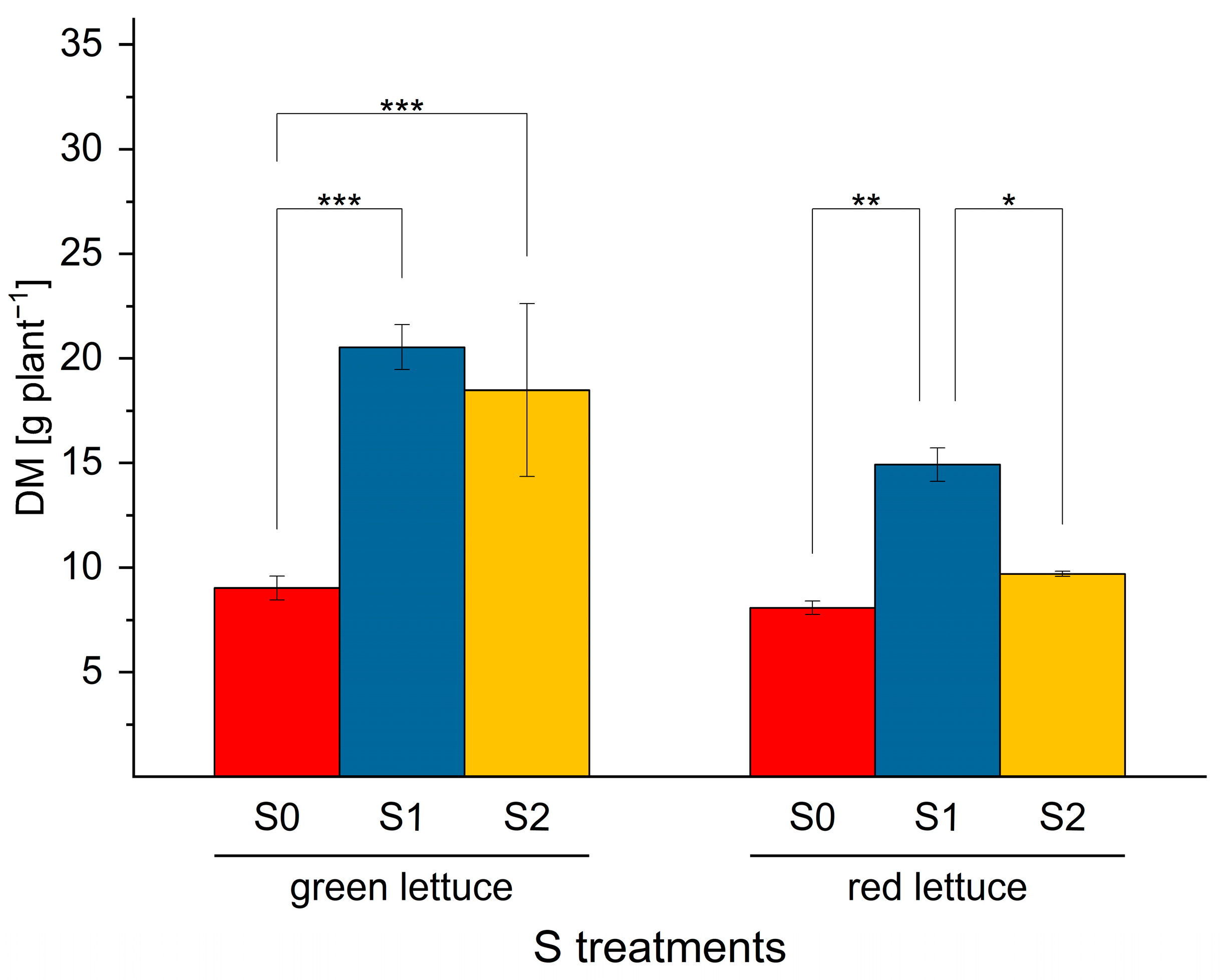
2.1. The Effect of S Nutrition on Dry Matter Accumulation in Green and Red Lettuce
2.2. Method Development for the Determination of Galactolipids
2.3. Method Validation for the Determination of Galactolipids
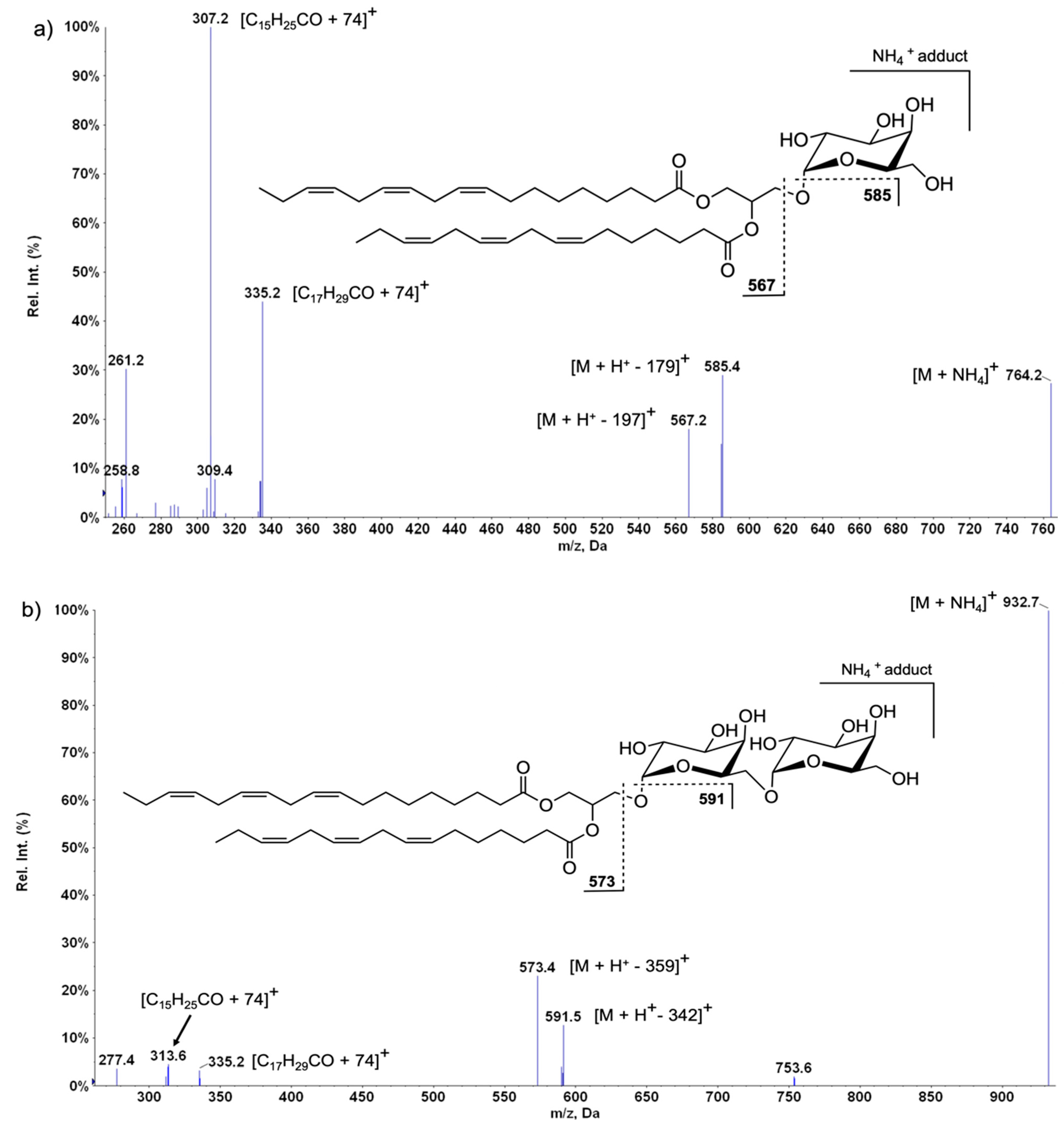
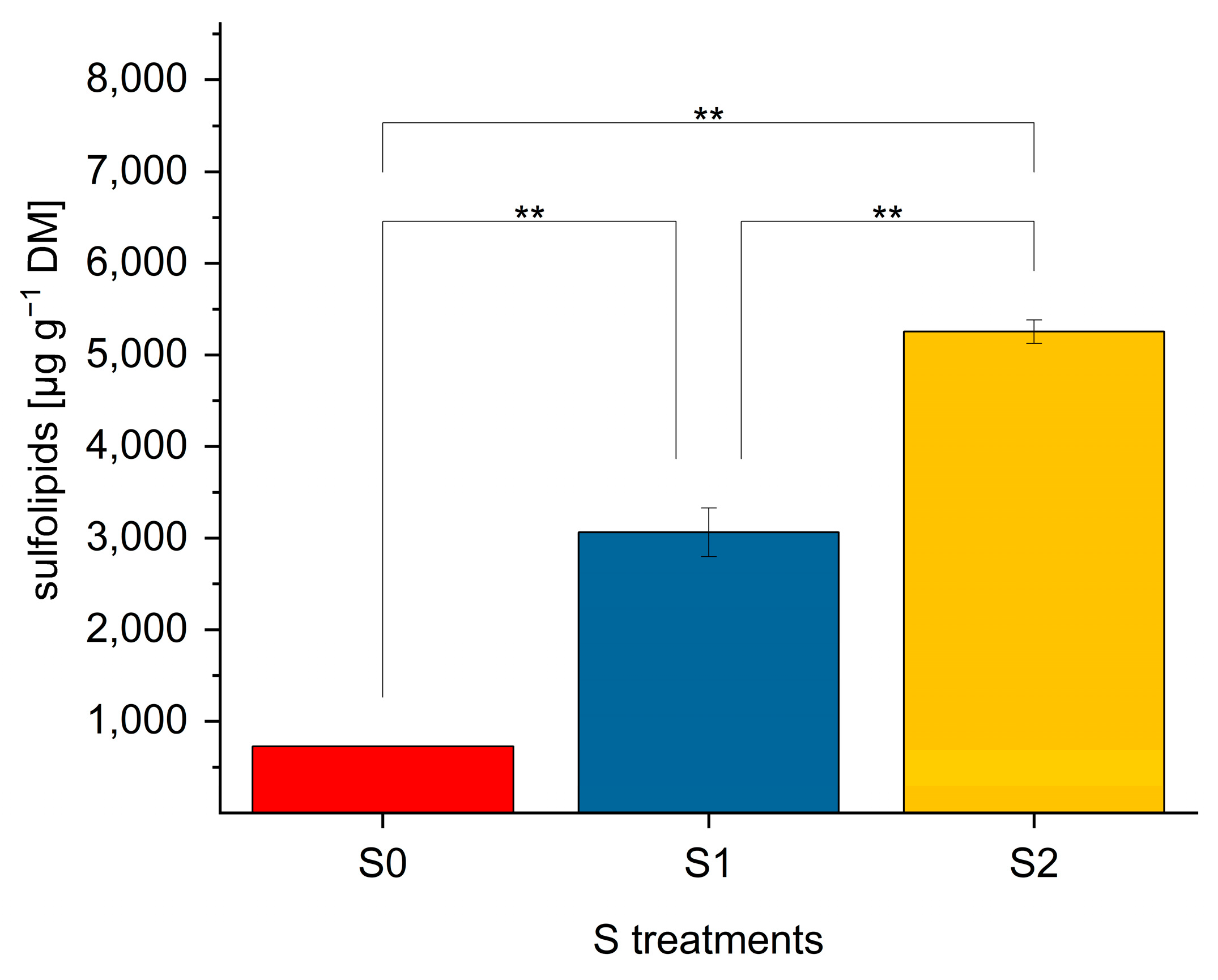
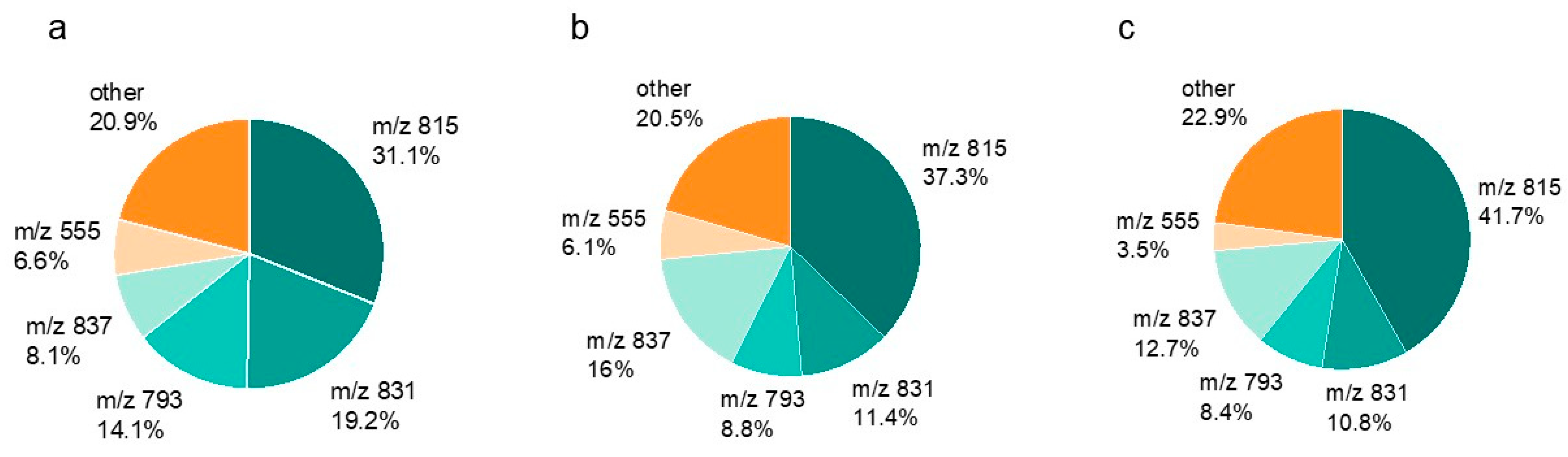
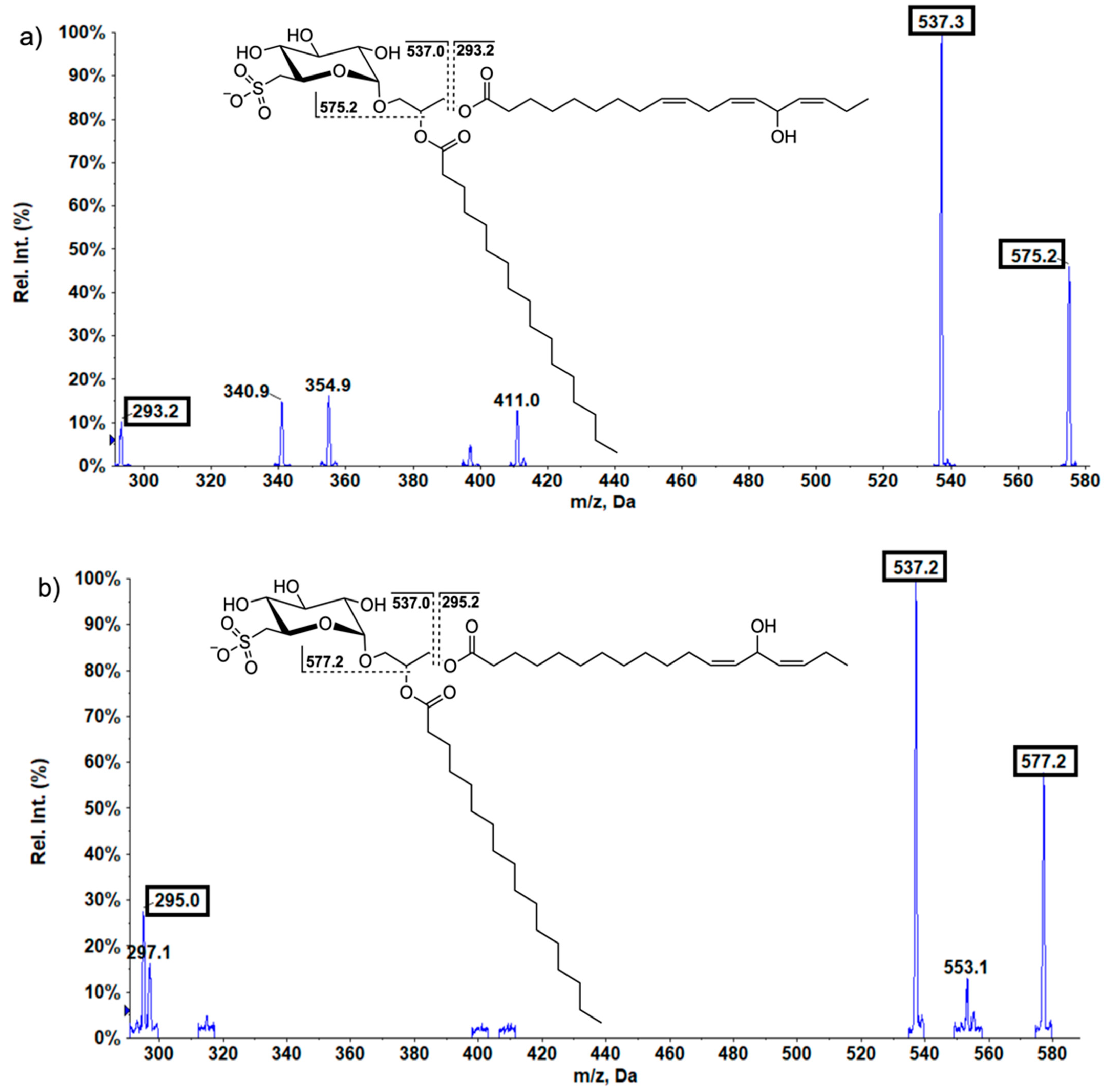
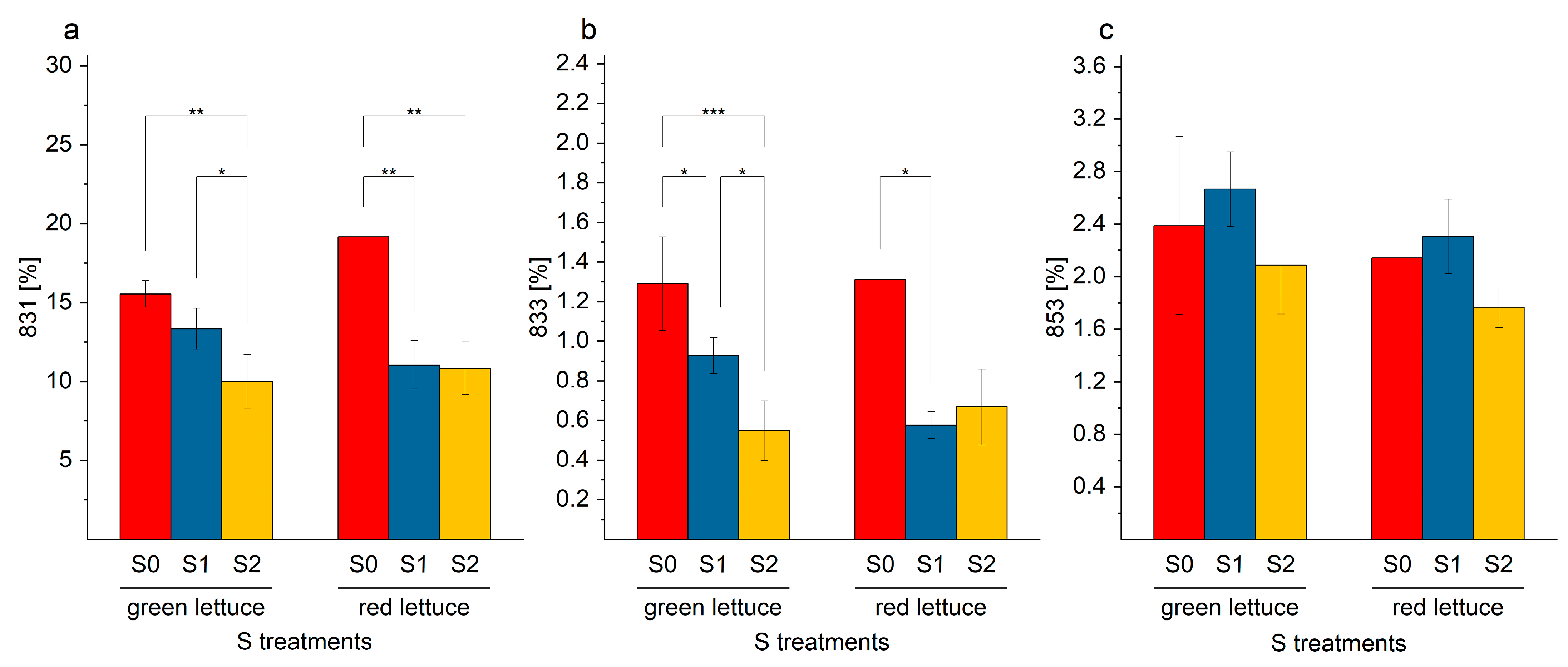
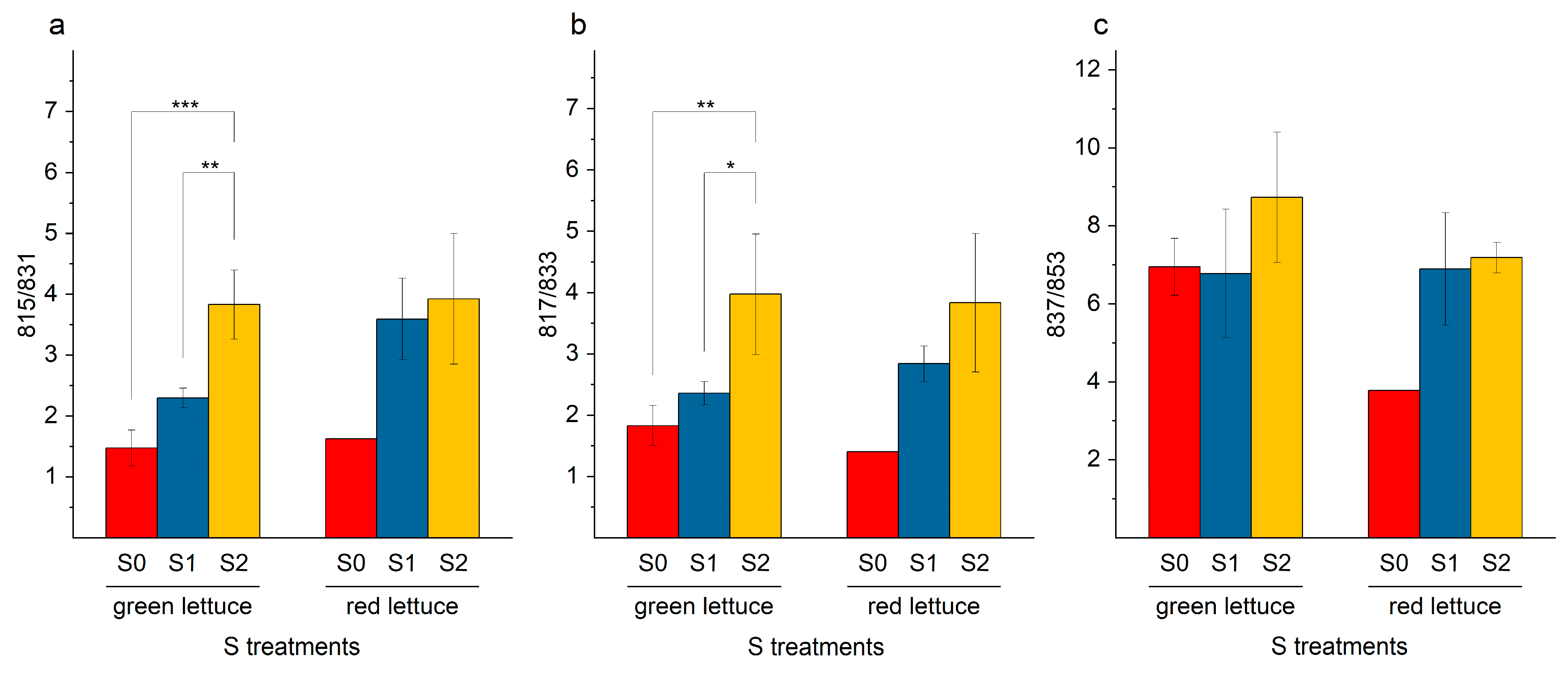
2.4. SQDG in Lettuce Plants
Oxidized SQDG in Lettuce Samples
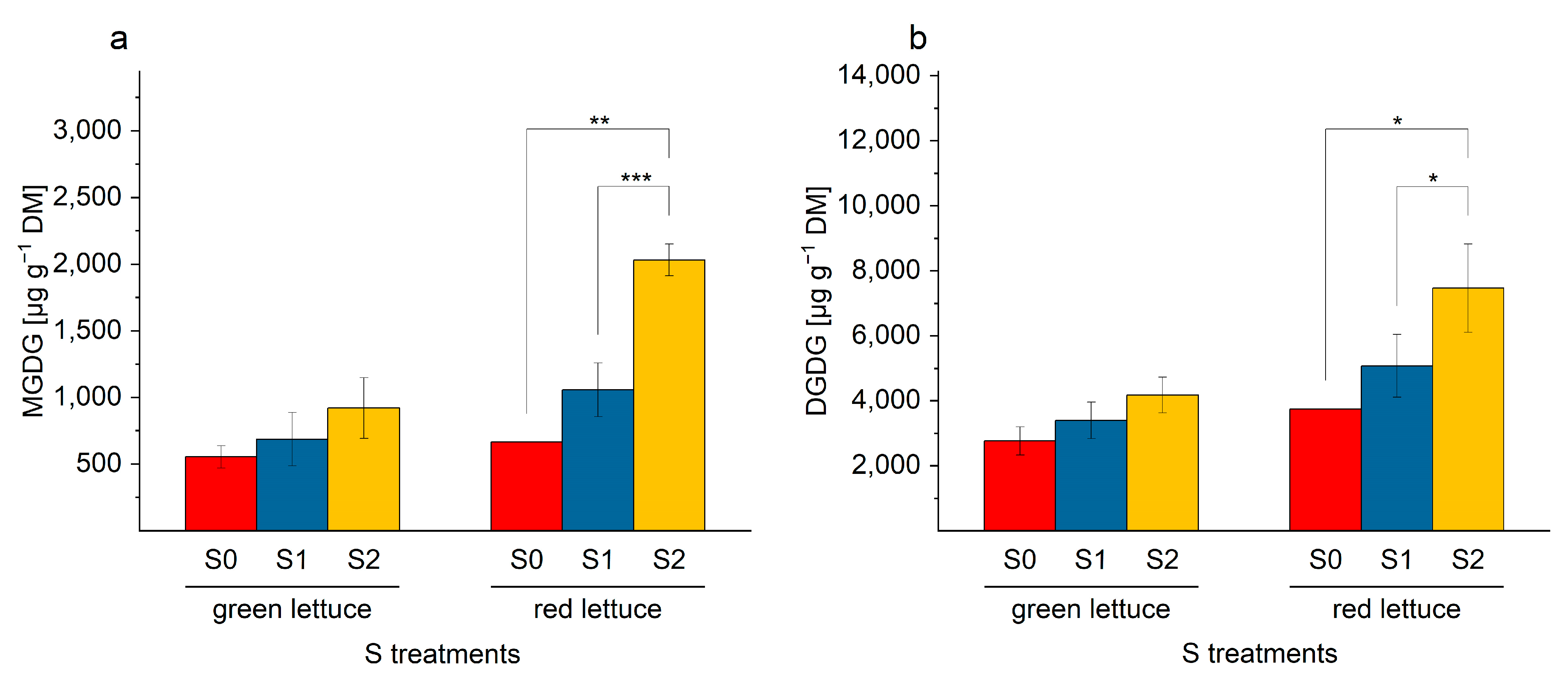
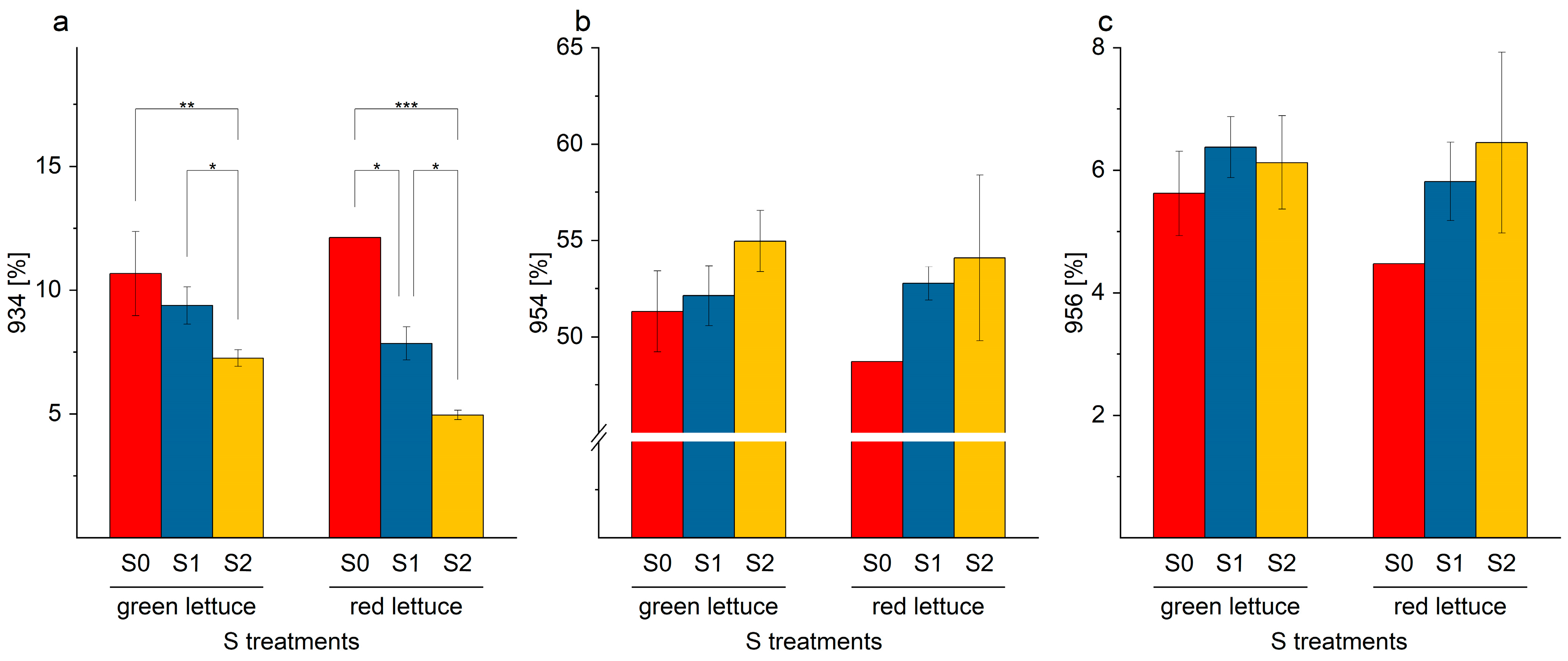
2.5. Galactolipids in Lettuce Samples
2.6. Determination of the Antioxidant Capacity in Lettuce Samples
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Chemicals and Materials
4.3. Preparation of Internal SQDG Standard
4.4. Lipid Extraction Procedure
4.5. Separation and Purification of the Lipids by Solid Phase Extraction
4.6. Determination of Sulfolipids Using LC-ESI-MS/MS
4.7. Quantification of Galactolipids (MGDG, DGDG) by LC-ESI-MS/MS
4.8. Method Validation
4.9. Determination of Antioxidant Capacity
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aires, A. Hydroponic production systems: Impact on nutritional status and bioactive compounds of fresh vegetables. In Vegetables: Importance of Quality Vegetables to Human Health; Asaduzzaman, M., Toshiki, A., Eds.; IntechOpen: London, UK, 2018; pp. 55–66. [Google Scholar]
- Abdalla, M.A.; Li, F.; Wenzel-Storjohann, A.; Sulieman, S.; Tasdemir, D.; Mühling, K.H. Comparative Metabolite Profile, Biological Activity and Overall Quality of Three Lettuce (Lactuca sativa L., Asteraceae) Cultivars in Response to Sulfur Nutrition. Pharmaceutics 2021, 13, 713. [Google Scholar] [CrossRef] [PubMed]
- Kopriva, S.; Malagoli, M.; Takahashi, H. Sulfur nutrition: Impacts on plant development, metabolism, and stress responses. J. Exp. Bot. 2019, 70, 4069–4073. [Google Scholar] [CrossRef]
- Etienne, P.; Sorin, E.; Maillard, A.; Gallardo, K.; Arkoun, M.; Guerrand, J.; Cruz, F.; Yvin, J.-C.; Ourry, A. Assessment of Sulfur Deficiency under Field Conditions by Single Measurements of Sulfur, Chloride and Phosphorus in Mature Leaves. Plants 2018, 7, 37. [Google Scholar] [CrossRef] [PubMed]
- McGrath, S.P.; Zhao, F.J.; Withers, P.J. Development of sulphur deficiency in crops and its treatment. Proc. Fertil. Soc. 1996, 379, 47. [Google Scholar]
- Jobe, T.O.; Zenzen, I.; Rahimzadeh Karvansara, P.; Kopriva, S. Integration of sulfate assimilation with carbon and nitrogen metabolism in transition from C3 to C4 photosynthesis. J. Exp. Bot. 2019, 70, 4211–4221. [Google Scholar] [CrossRef]
- Anjum, N.A.; Gill, R.; Kaushik, M.; Hasanuzzaman, M.; Pereira, E.; Ahmad, I.; Tuteja, N.; Gill, S.S. ATP-sulfurylase, sulfur-compounds, and plant stress tolerance. Front. Plant Sci. 2015, 6, 210. [Google Scholar] [CrossRef]
- Prioretti, L.; Gontero, B.; Hell, R.; Giordano, M. Diversity and regulation of ATP sulfurylase in photosynthetic organisms. Front. Plant Sci. 2014, 5597, 1–12. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Nazar, R.; Iqbal, N.; Masood, A.; Syeed, S.; Khan, N.A. Understanding the significance of sulphur in improving salinity tolerance in plants. Environ. Exp. Bot. 2011, 70, 80–87. [Google Scholar] [CrossRef]
- Mazid, M.; Khan, Z.H.; Quddusi, S.; Khan, T.A.; Mohammad, F. Significance of sulphur nutrition against metal induced oxida-tive stress in plants. J. Stress Physiol. Biochem. 2011, 7, 165–184. [Google Scholar]
- Bystrická, J.; Kavalcová, P.; Vollmannová, A.; Tomáš, J.; Orsák, M. The Role of Sulphur on the Content of Total Polyphenols and Antioxidant Activity in Onion (Allium cerpa L.). Potravinarstvo 2014, 8, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, S.; Zuchi, S. Adequate sulfur supply protects barley plants from adverse effects of salinity stress by increasing thiol contents. Acta Physiol. Plant. 2013, 35, 175–181. [Google Scholar] [CrossRef]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef]
- Pospíšil, P.; Yamamoto, Y. Damage to photosystem II by lipid peroxidation products. Biochim. Biophys. Acta-Gen. Subj. 2017, 1861, 457–466. [Google Scholar] [CrossRef]
- Berlett, B.S.; Stadtman, E.R. Protein oxidation in aging, disease, and oxidative stress. J. Biol. Chem. 1997, 272, 20313–20316. [Google Scholar] [CrossRef] [PubMed]
- Czarnocka, W.; Karpiński, S. Friend or foe? Reactive oxygen species production, scavenging and signaling in plant response to environmental stresses. Free. Radic. Biol. Med. 2018, 122, 4–20. [Google Scholar] [CrossRef]
- Edreva, A. Generation and scavenging of reactive oxygen species in chloroplasts: A submolecular approach. Agric. Ecosyst. Environ. 2005, 106, 119–133. [Google Scholar] [CrossRef]
- Foyer, C.H.; Lelandais, M.; Kunert, K.J. Photooxidative stress in plants. Physiol. Plant. 1994, 92, 696–717. [Google Scholar] [CrossRef]
- Slooten, L.; Van Montagu, M.; Inzé, D. Manipulation of oxidative stress tolerance in transgenic plants. Transgenic Plant Res. 1998, 1, 241–262. [Google Scholar]
- Niyogi, K.K. Photoprotection revisited: Genetic and molecular approaches. Annu. Rev. Plant Biol. 1999, 50, 333–359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Weng, J.; Pan, J.; Tu, T.; Yao, S.; Xu, C. Study on the photo-generation of superoxide radicals in Photosystem II with EPR spin trapping techniques. Photosynth. Res. 2003, 75, 41–48. [Google Scholar] [CrossRef]
- Lunde, C.; Zygadlo, A.; Simonsen, H.T.; Nielsen, P.L.; Blennow, A.; Haldrup, A. Sulfur starvation in rice: The effect on photosynthesis, carbohydrate metabolism, and oxidative stress protective pathways. Physiol. Plant. 2008, 134, 508–521. [Google Scholar] [CrossRef]
- Körber, T.T.; Frantz, N.; Sitz, T.; Abdalla, M.A.; Mühling, K.H.; Rohn, S. Alterations of Content and Composition of Individual Sulfolipids, and Change of Fatty Acids Profile of Galactolipids in Lettuce Plants (Lactuca sativa L.) Grown under Sulfur Nutrition. Plants 2022, 11, 1342. [Google Scholar] [CrossRef]
- Lambers, H.; Cawthray, G.R.; Giavalisco, P.; Kuo, J.; Laliberté, E.; Pearse, S.J.; Turner, B.L. Proteaceae from severely phosphorus-impoverished soils extensively replace phospholipids with galactolipids and sulfolipids during leaf development to achieve a high photosynthetic phosphorus-use-efficiency. New Phytol. 2012, 196, 1098–1108. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K.; Sato, N.; Tsuzuki, M. Utilization of a chloroplast membrane sulfolipid as a major internal sulfur source for protein synthesis in the early phase of sulfur starvation in Chlamydomonas reinhardtii. FEBS Lett. 2007, 581, 4519–4522. [Google Scholar] [CrossRef]
- Bybordi, A.; Tabatabaei, S.J.; Ahmadev, A. Effects of salinity on fatty acid composition of canola (Brassica napus L.). J. Food Agric. Environ. 2010, 8, 113–115. [Google Scholar]
- Monge, E.; Perez, C.; Pequerul, A.; Madero, P.; Val, J. Effect of iron chlorosis on mineral nutrition and lipid composition of thylakoid biomembrane in Prunus persica (L.) Bastch, in Optimization of Plant Nutrition. In Proceedings of the Refereed Papers from the Eighth International Colloquium for the Optimization of Plant Nutrition, Lisbon, Portugal, 31 August–8 September 1999; Fragoso, M.A.C., Van Beusichem, M.L., Houwers, A., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 477–482. [Google Scholar]
- Cuppettz, S.L.; McCluskey, M.M.; Paparozzi, E.T.; Parkhursp, A. Nitrogen and sulfur effects on leaf lettuce quality. J. Food Qual. 1999, 22, 363–373. [Google Scholar] [CrossRef]
- Chung, J.S.; Kim, H.C.; Yun, S.M.; Kim, H.J.; Kim, C.S.; Lee, J.J. Metabolite analysis of lettuce in response to sulfur nutrition. Horticulturae 2022, 8, 734. [Google Scholar] [CrossRef]
- Murphy, R.C. Glyceryl Esters. In Tandem Mass Spectrometry of Lipids; Royal Society of Chemistry: Cambridge, UK, 2014; pp. 219–224. [Google Scholar]
- Zhang, M.; Deng, X.; Yin, L.; Qi, L.; Wang, X.; Wang, S.; Li, H. Regulation of galactolipid biosynthesis by overexpression of the rice MGD gene contributes to enhanced aluminum tolerance in tobacco. Front. Plant Sci. 2016, 7, 337. [Google Scholar] [CrossRef]
- FDA. In Guidance for Industry: Bioanalytical Method Validation; US Department of Health and Human Services Food and Drug Administration Center for Drug Evaluation and Research (CDER), Center for Veterinary Medicine (CVM): South Laurel, Maryland, 2018.
- Triantaphylides, C.; Krischke, M.; Hoeberichts, F.A.; Ksas, B.; Gresser, G.; Havaux, M.; Mueller, M.J. Singlet oxygen is the major reactive oxygen species involved in photooxidative damage to plants. Plant Physiol. 2008, 148, 960–968. [Google Scholar] [CrossRef]
- Fischer, J.; Treblin, M.; Sitz, T.; Rohn, S. Development of a targeted HPLC-ESI-QqQ-MS/MS method for the quantification of sulfolipids from a cyanobacterium, selected leafy vegetables, and a microalgae species. Anal. Bioanal. Chem. 2021, 413, 1941–1954. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Giordano, M.; Raven, J.A. Nitrogen and Sulfur assimilation in plants and algae. Aquat. Bot. 2014, 118, 45–61. [Google Scholar] [CrossRef]
- Samanta, S.; Singh, A.; Roychoudhury, A. Involvement of sulfur in the regulation of abiotic stress tolerance in plants. Protective chemical agents in the amelioration of plant abiotic stress. Biochem. Mol. Perspect. 2020, 22, 437–466. [Google Scholar]
- Senizza, B.; Zhang, L.; Miras-Moreno, B.; Righetti, L.; Zengin, G.; Ak, G.; Bruni, R.; Lucini, L.; Sifola, M.I.; El-Nakhel, C. The strength of the nutrient solution modulates the functional profile of hydroponically grown lettuce in a genotype-dependent manner. Foods 2020, 9, 1156. [Google Scholar] [CrossRef]
- Liu, X.; Ma, D.; Zhang, Z.; Wang, S.; Du, S.; Deng, X.; Yin, L. Plant lipid remodeling in response to abiotic stresses. Environ. Exp. Bot. 2019, 165, 174–184. [Google Scholar] [CrossRef]
- Fujii, S.; Kobayashi, K.; Nakamura, Y.; Wada, H. Inducible knockdown of Monogalactosyldiacylglycerol Synthase1reveals roles of galactolipids in organelle differentiation in Arabidopsis cotyledons. Plant Physiol. 2014, 166, 1436–1449. [Google Scholar] [CrossRef]
- Kobayashi, K. Role of membrane glycerolipids in photosynthesis, thylakoid biogenesis and chloroplast development. J. Plant Res. 2016, 129, 565–580. [Google Scholar] [CrossRef] [PubMed]
- Stroebel, D.; Choquet, Y.; Popot, J.L.; Picot, D. An atypical haem in the cytochrome b6f complex. Nature 2003, 426, 413–418. [Google Scholar] [CrossRef] [PubMed]
- Standfuss, J.; Terwisscha van Scheltinga, A.C.; Lamborghini, M.; Kühlbrandt, W. Mechanisms of photoprotection and nonphotochemical quenching in pea light-harvesting complex at 2.5 Å resolution. EMBO J. 2005, 24, 919–928. [Google Scholar] [CrossRef]
- Guskov, A.; Kern, J.; Gabdulkhakov, A.; Broser, M.; Zouni, A.; Saenger, W. Cyanobacterial photosystem II at 2.9-Å resolution and the role of quinones, lipids, channels and chloride. Nat. Struct. Mol. Biol. 2009, 16, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yan, H.; Wang, K.; Kuang, T.; Zhang, J.; Gui, L.; Chang, W. Crystal structure of spinach major light-harvesting complex at 2.72 Å resolution. Nature 2004, 428, 287–292. [Google Scholar] [CrossRef]
- Sato, N.; Tsuzuki, M.; Matsuda, Y.; Ehara, T.; Osafune, T.; Kawaguchi, A. Isolation and characterization of mutants affected in lipid metabolism of Chlamydomonas reinhardtii. Eur. J. Biochem. 1995, 230, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Minoda, A.; Sonoike, K.; Okada, K.; Sato, N.; Tsuzuki, M. Decrease in the efficiency of the electron donation to tyrosine Z of photosystem II in an SQDG-deficient mutant of Chlamydomonas. FEBS Lett. 2003, 553, 109–112. [Google Scholar] [CrossRef]
- Sugimoto, K.; Tsuzuki, M.; Sato, N. Regulation of synthesis and degradation of a sulfolipid under sulfur-starved conditions and its physiological significance in Chlamydomonas reinhardtii. New Phytol. 2010, 185, 676–686. [Google Scholar] [CrossRef] [PubMed]
- Gaude, N.; Bréhélin, C.; Tischendorf, G.; Kessler, F.; Dörmann, P. Nitrogen deficiency in Arabidopsis affects galactolipid composition and gene expression and results in accumulation of fatty acid phytyl esters. Plant J. 2007, 49, 729–739. [Google Scholar] [CrossRef]
- Shimojima, M.; Ohta, H. Critical regulation of galactolipid synthesis controls membrane differentiation and remodeling in distinct plant organs and following environmental changes. Prog. Lipid Res. 2011, 50, 258–266. [Google Scholar] [CrossRef]
- Garab, G.; Lohner, K.; Laggner, P.; Farkas, T. Self-regulation of the lipid content of membranes by non-bilayer lipids: A hypothesis. Trends Plant Sci. 2000, 5, 489–494. [Google Scholar] [CrossRef]
- Wang, S.; Uddin, M.I.; Tanaka, K.; Yin, L.; Shi, Z.; Qi, Y. Maintenance of chloroplast structure and function by overexpression of the rice Monogalactosyldiacylglycerol Synthase1 gene leads to enhanced salt tolerance in tobacco. Plant Physiol. 2014, 165, 1144–1155. [Google Scholar] [CrossRef]
- Zhang, M.; Barg, R.; Yin, M. Modulated fatty acid desaturation via overexpression of two distinct omega-3 desaturases differentially alters tolerance to various abiotic stresses in transgenic tobacco cells and plants. Plant J. 2005, 44, 361–371. [Google Scholar] [CrossRef]
- Sui, N.; Li, M.; Li, K.; Song, J.; Wang, B.S. Increase in unsaturated fatty acids in membrane lipids of Suaeda salsa L. enhances protection of photosystem II under high salinity. Photosynthetica 2010, 48, 623–629. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, X.Y.; Li, F.; Luo, Y.; Wang, W. Overaccumulation of glycine betaine enhances tolerance of the photosynthetic apparatus to drought and heat stress in wheat. Photosynthetica 2010, 48, 30–41. [Google Scholar] [CrossRef]
- Dakhma, W.S.; Zarrouk, M.; Cherif, A. Effects of drought-stress on lipids in rape leaves. Phytochemistry 1995, 40, 1383–1386. [Google Scholar] [CrossRef]
- Barnes, A.C.; Benning, C.; Roston, R.L. Chloroplast membrane remodeling during freezing stress is accompanied by cytoplasmic acidification activating Sensitive to freezing. Plant Physiol. 2016, 171, 2140–2149. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wang, S.; Qi, L.; Yin, L.; Deng, X. Galactolipid remodeling is involved in drought-induced leaf senescence in maize. Environ. Exp. Bot. 2018, 150, 57–68. [Google Scholar] [CrossRef]
- Ozolina, N.V.; Gurina, V.V.; Nesterkina, I.S.; Nurminsky, V.N. Variations in the content of tonoplast lipids under abiotic stress. Planta 2020, 251, 1–15. [Google Scholar] [CrossRef]
- Webb, M.S.; Green, B.R. Biochemical and biophysical properties of thylakoid acyl lipids. Biochim. Biophys. Acta (BBA)-Bioenerg. 1991, 1060, 133–158. [Google Scholar] [CrossRef]
- Mahajan, S.; Tuteja, N. Cold, salinity and drought stresses: An overview. Arch. Biochem. Biophys. 2005, 444, 139–158. [Google Scholar] [CrossRef]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed]
- Anjum, N.A.; Sofo, A.; Scopa, A. Lipids and proteins—Major targets of oxidative modifications in abiotic stressed plants. Environ. Sci. Pollut. Res. 2015, 22, 4099–4121. [Google Scholar] [CrossRef]
- De Souza, L.M.; Sassaki, G.L.; Romanos, M.T.V.; Barreto-Bergter, E. Structural Characterization and Anti-HSV-1 and HSV-2 Activity of Glycolipids from the Marine Algae Osmundaria obtusiloba Isolated from Southeastern Brazilian Coast. Mar. Drugs 2012, 10, 918–931. [Google Scholar] [CrossRef]
- Tiwari, A.M.; Rác, P.; Pospíšil, P. Formation of superoxide anion and carbon-centered radicals by photosystem II under high light and heat stress—EPR spin-trapping study. J. Bioenerg. Biomembr. 2013, 45, 551–559. [Google Scholar] [CrossRef]
- Dixon, R.A.; Paiva, N.L. Stress-induced phenylpropanoid metabolism. Plant Cell 1995, 7, 1085. [Google Scholar] [CrossRef] [PubMed]
- Gershenzon, J. Changes in the levels of plant secondary metabolites under water and nutrient stress. In Phytochemical Adaptations to Stress; Timmermann, B.N., Steelink, C., Loewus, F.A., Eds.; Springer: New York, NY, USA, 1984; pp. 273–320. [Google Scholar]
- Rice-Evans, C.; Miller, N.; Paganga, G. Antioxidant properties of phenolic compounds. Trends Plant Sci. 1997, 2, 152–159. [Google Scholar] [CrossRef]
- Gharibi, S.; Riahi, L.; Yangui, I.; Messaoud, C.; Rzigui, T.; Zouhair, N. Effect of drought stress on total phenolic, lipid peroxidation, and antioxidant activity of Achillea species. Appl. Biochem. Biotechnol. 2016, 178, 796–809. [Google Scholar] [CrossRef]
- Sitz, T.; Domey, H.; Fischer, J.; Rohn, S. An Alternative Approach for the Synthesis of Sulfoquinovosyldiacylglycerol. Molecules 2021, 26, 4275. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Cheng, K.; Wong, C.; Fan, K.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyte [M+ NH4+]+ | RT (min) | MRM (m/z) | DT | DP (V) | EP (V) | CE (V) | FA |
|---|---|---|---|---|---|---|---|
| 742 | 25.9 | 742.5 → 307.5 | 200 | 86 | 10 | 37 | 16:0/16:3 |
| 742.5 → 313.5 | 200 | 86 | 10 | 37 | |||
| 742.5 → 563.5 | 200 | 86 | 10 | 17 | |||
| 764 | 16.1 | 764.5 → 335.5 | 200 | 56 | 10 | 43 | 16:3/18:3 |
| 764.5 → 567.5 | 200 | 56 | 10 | 27 | |||
| 764.5 → 585.5 | 200 | 56 | 10 | 25 | |||
| 766 | 19.7 | 766.5 → 335.5 | 200 | 76 | 10 | 35 | 16:2/18:3 |
| 766.5 → 569.5 | 200 | 76 | 10 | 33 | |||
| 766.5 → 587.5 | 200 | 76 | 10 | 23 | |||
| 768 | 25.5 | 768.5 → 309.5 | 200 | 76 | 10 | 47 | 16:1/18:3 |
| 768.5 → 571.5 | 200 | 76 | 10 | 27 | |||
| 768.5 → 589.5 | 200 | 76 | 10 | 37 | |||
| 770 | 36.9 | 770.5 → 261.5 | 200 | 71 | 10 | 51 | 16:0/18:3 |
| 770.5 → 313.5 | 200 | 71 | 10 | 49 | |||
| 770.5 → 573.5 | 200 | 71 | 10 | 52 | |||
| 792 | 20.8 | 792.5 → 261.5 | 200 | 51 | 10 | 45 | 18:3/18:3 |
| 792.5 → 335.5 | 200 | 51 | 10 | 45 | |||
| 792.5 → 613.5 | 200 | 51 | 10 | 27 | |||
| 794 | 26.8 | 794.5 → 337.5 | 200 | 81 | 10 | 43 | 18:2/18:3 |
| 794.5 → 335.5 | 200 | 81 | 10 | 47 | |||
| 794.5 → 597.5 | 200 | 81 | 10 | 25 | |||
| 796 | 30.2 | 796.5 → 337.5 | 200 | 56 | 10 | 55 | 18:2/18:2 |
| 796.5 → 537.5 | 200 | 56 | 10 | 37 | |||
| 796.5 → 613.5 | 200 | 56 | 10 | 52 |
| Analyte [M+ NH4+]+ | RT (min) | MRM (m/z) | DT | DP (V) | EP (V) | CE (V) | FA |
|---|---|---|---|---|---|---|---|
| 904 | 17.6 | 904.6 → 233.0 | 200 | 121 | 10 | 43 | 16:3/16:0 |
| 904.6 → 563.4 | 200 | 121 | 10 | 35 | |||
| 904.6 → 545.6 | 200 | 121 | 10 | 37 | |||
| 926 | 12.2 | 926.6 → 233.0 | 200 | 126 | 10 | 41 | 16:3/18:3 |
| 926.6 → 261.2 | 200 | 126 | 10 | 57 | |||
| 926.6 → 585.0 | 200 | 126 | 10 | 31 | |||
| 928 | 14.3 | 928.6 → 587.6 | 200 | 141 | 10 | 29 | 16:3/18:2 |
| 928.6 → 233.2 | 200 | 141 | 10 | 37 | |||
| 928.6 → 263.2 | 200 | 141 | 10 | 57 | |||
| 932 | 23.8 | 932.6 → 573.4 | 200 | 121 | 10 | 31 | 16:0/18:3 |
| 932.6 → 591.2 | 200 | 121 | 10 | 27 | |||
| 932.6 → 313.0 | 200 | 121 | 10 | 41 | |||
| 934 | 31.7 | 934.6 → 263.1 | 200 | 116 | 10 | 43 | 16:1/18:1 |
| 934.6 → 575.2 | 200 | 116 | 10 | 41 | |||
| 934.6 → 593.6 | 200 | 116 | 10 | 25 | |||
| 904 | 17.6 | 904.6 → 233.0 | 200 | 121 | 10 | 43 | 16:3/16:0 |
| 904.6 → 563.4 | 200 | 121 | 10 | 35 | |||
| 904.6 → 545.6 | 200 | 121 | 10 | 37 | |||
| 926 | 12.2 | 926.6 → 233.0 | 200 | 126 | 10 | 41 | 16:3/18:3 |
| 926.6 → 261.2 | 200 | 126 | 10 | 57 | |||
| 926.6 → 585.0 | 200 | 126 | 10 | 31 | |||
| 928 | 14.3 | 928.6 → 587.6 | 200 | 141 | 10 | 29 | 16:3/18:2 |
| 928.6 → 233.2 | 200 | 141 | 10 | 37 | |||
| 928.6 → 263.2 | 200 | 141 | 10 | 57 | |||
| 932 | 23.8 | 932.6 → 573.4 | 200 | 121 | 10 | 31 | 16:0/18:3 |
| 932.6 → 591.2 | 200 | 121 | 10 | 27 | |||
| 932.6 → 313.0 | 200 | 121 | 10 | 41 | |||
| 934 | 31.7 | 934.6 → 263.1 | 200 | 116 | 10 | 43 | 16:1/18:1 |
| 934.6 → 575.2 | 200 | 116 | 10 | 41 | |||
| 934.6 → 593.6 | 200 | 116 | 10 | 25 | |||
| 954 | 15.0 | 954.6 → 333.0 | 200 | 56 | 10 | 39 | 18:3/18:3 |
| 954.6 → 613.2 | 200 | 56 | 10 | 55 | |||
| 954.6 → 596.6 | 200 | 56 | 10 | 35 | |||
| 956 | 18.4 | 956.6 → 615.4 | 200 | 136 | 10 | 27 | 18:2/18:3 |
| 956.6 → 335.2 | 200 | 136 | 10 | 55 | |||
| 956.6 → 595.4 | 200 | 136 | 10 | 37 | |||
| 958 | 23.5 | 958.6 → 261.4 | 200 | 136 | 10 | 46 | 18:1/18:3 |
| 958.6 → 617.2 | 200 | 136 | 10 | 27 | |||
| 958.6 → 337.4 | 200 | 136 | 10 | 52 |
| Lettuce Species | Individual SQDG m/z | SQDG Content at Varied S Treatments [mM] | Enrichment Factor (C1.5mM/c0mM) | ||
|---|---|---|---|---|---|
| S0 [%] 1 | S1 [%] 2 | S2 [%] 3 | |||
| Red multi-leaf | 765 | 0.6 a | 0.2 ± 0.01 b | 0.3 ± 0.00 c | 3.3 |
| 787 | 0.2 a | 0.4 ± 0.03 b | 0.3 ± 0.02 ab | 10.6 | |
| 789 | 0.3 | 0.4 ± 0.02 | 0.4 ± 0.04 | 8.6 | |
| 791 | 0.7 | 0.7 ± 0.06 | 0.8 ± 0.11 | 8.2 | |
| 793 | 14.1 a | 8.1± 1.06 b | 8.5 ± 1.40 b | 4.3 | |
| 801 | 0.2 | 0.2 ±0.01 | 0.2 ± 0.00 | 8.2 | |
| 803 | 0.1 a | 0.1 ± 0.01 b | 0.1 ± 0.01 ac | 7.5 | |
| 813 | 1.5 a | 2.8 ± 0.26 b | 2.9 ± 0.49 b | 14.0 | |
| 815 | 31.1 | 39.1 ± 3.51 | 41.6 ± 5.05 | 9.7 | |
| 817 | 1.8 a | 1.6 ± 0.16 ab | 2.5 ± 0.02 c | 9.6 | |
| 819 | 0.5 a | 0.9 ± 0.11 ab | 1.1 ± 0.14 b | 16.2 | |
| 821 | 0.4 | 0.5 ± 0.07 | 0.6 ± 0.08 | 9.3 | |
| 837 | 8.1 a | 15.6 ± 1.64 b | 12.7 ± 0.43 ab | 11.3 | |
| 839 | 4.4 | 3.7 ± 0.59 | 4.1 ± 0.37 | 6.6 | |
| 841 | 3.0 | 2.8 ± 0.38 | 3.1 ± 0.14 | 7.4 | |
| 843 | 2.9 ab | 2.4 ± 0.20 a | 3.4 ± 0.33 b | 8.4 | |
| 845 | 0.4 ab | 0.5 ± 0.08 a | 0.7 ± 0.10 b | 12.1 | |
| 867 | 0.2 | 0.1 ± 0.02 | 0.1 ± 0.02 | 3.9 | |
| 555 | 6.6 | 6.2 ± 1.25 | 3.5 ± 0.65 | 3.8 | |
| 831 | 19.2 a | 11.1 ± 1.53 b | 10.8 ± 1.67 b | 4.1 | |
| 833 | 1.3 a | 0.6 ± 0.07 b | 0.7 ± 0.19 b | 3.7 | |
| 853 | 2.1 | 2.3 ± 0.28 | 1.8 ± 0.16 | 6.0 | |
| 831 | 15.6 ± 0.83 | 13.3 ± 1.28 | 10.0 ± 1.73 | 4.8 | |
| Green multi-leaf | 833 | 1.3 ± 0.24 | 0.9 ± 0.09 | 0.5 ± 0.15 | 5.0 |
| 853 | 2.4 ± 0.68 | 2.7 ± 0.28 | 2.1 ± 0.37 | 3.0 | |
| Individual MGDG or DGDG m/z | Green Lettuce | Red Lettuce | ||||
|---|---|---|---|---|---|---|
| Galactolipid Content at Varied S Treatments [mM] | Galactolipid Content at Varied S Treatments [mM] | |||||
| S0 1 | S1 2 | S2 3 | S0 4 | S1 5 | S2 6 | |
| MGDG | ||||||
| 764 [%] | 0.11 ± 0.0 | 0.14 ± 0.0 | 0.15 ± 0.0 | 0.13 | 0.16 ± 0.0 | 0.19 ± 0.0 |
| 766 [%] | 0.30 ± 0.0 | 0.28 ± 0.0 | 0.26 ± 0.0 | 0.24 | 0.25 ± 0.0 | 0.30 ± 0.1 |
| 768 [%] | 0.41 ± 0.1 | 0.45 ± 0.1 | 0.39 ±0.0 | 0.38 | 0.39 ± 0.0 | 0.38 ±0.0 |
| 770 [%] | 1.45 ± 0.1 | 1.30 ±0.2 | 1.07 ± 0.1 | 1.41 | 1.25 ± 0.2 | 1.06 ± 0.2 |
| 792 [%] | 95.04 ± 0.4 | 94.77 ± 0.7 | 95.38 + 0.4 | 95.84 | 95.60 ± 0.5 | 95.41 ± 0.7 |
| 794 [%] | 2.14 ± 0.3 | 2.47 ± 0.4 | 2.28 ± 0.2 | 1.60 | 1.94 ± 0.2 | 2.35 ± 0.4 |
| 796 [%] | 0.56 ± 0.1 | 0.60 ± 0.1 | 0.47 ± 0.1 | 0.41 | 0.42 ± 0.1 | 0.32 ± 0.0 |
| DGDG | ||||||
| 926 [%] | 0.07 ± 0.0 | 0.09 ± 0.0 | 0.10 ± 0.0 | 0.07 | 0.12± 0.0 | 0.101± 0.0 |
| 928 [%] | 0.12 ± 0.0 | 0.13 ± 0.0 | 0.16 ± 0.0 | 0.12 | 0.15 ± 0.0 | 0.17 ± 0.0 |
| 932 [%] | 26.50 ± 1.7 | 24.98 ± 2.4 | 24.96 ± 0.6 | 30.25 | 28.43 ± 1.2 | 30.67 ± 2.0 |
| 934 [%] | 10.67 ± 1.7 a | 9.38 ± 0.8 a | 7.3 ± 0.3 b | 12.13 a | 7.86 ± 0.7 b | 4.97 ± 0.2 c |
| 954 [%] | 51.31 ± 2.1 | 52.14 ± 1.6 | 54.97 ± 1.6 | 48.71 | 52.77 ± 0.9 | 54.10 ± 4.3 |
| 956 [%] | 5.62 ± 0.7 | 6.38 ± 0.5 | 6.13 ± 0.8 | 4.48 | 5.82 ± 0.6 | 6.45 ± 1.5 |
| 958 [%] | 5.70 ± 0.6 | 6.90 ± 0.4 | 6.44 ± 0.4 | 4.24 | 4.86 ± 0.14 | 3.53 ± 0.6 |
| Parameter | Green Lettuce | Red Lettuce | ||||
|---|---|---|---|---|---|---|
| S0 1 | S1 2 | S2 3 | S0 4 | S1 5 | S2 6 | |
| TPC [mg GAE g−1 DM] | 1.11 ± 0.3 | 0.75 ± 0.06 | 0.71 ± 0.1 | 2.2 | 2.0 ± 0.4 | 1.5 |
| TEAC [mM TE g−1 DM] | 2.14 ± 0.44 | 1.55 ± 0.2 | 1.37 ± 0.1 | 5.18 a | 3.73 ± 0.5 b | 3.88 a,b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Körber, T.T.; Sitz, T.; Abdalla, M.A.; Mühling, K.H.; Rohn, S. LC-ESI-MS/MS Analysis of Sulfolipids and Galactolipids in Green and Red Lettuce (Lactuca sativa L.) as Influenced by Sulfur Nutrition. Int. J. Mol. Sci. 2023, 24, 3728. https://doi.org/10.3390/ijms24043728
Körber TT, Sitz T, Abdalla MA, Mühling KH, Rohn S. LC-ESI-MS/MS Analysis of Sulfolipids and Galactolipids in Green and Red Lettuce (Lactuca sativa L.) as Influenced by Sulfur Nutrition. International Journal of Molecular Sciences. 2023; 24(4):3728. https://doi.org/10.3390/ijms24043728
Chicago/Turabian StyleKörber, Tania T., Tobias Sitz, Muna A. Abdalla, Karl H. Mühling, and Sascha Rohn. 2023. "LC-ESI-MS/MS Analysis of Sulfolipids and Galactolipids in Green and Red Lettuce (Lactuca sativa L.) as Influenced by Sulfur Nutrition" International Journal of Molecular Sciences 24, no. 4: 3728. https://doi.org/10.3390/ijms24043728