
The Interplay of Sulfur and Selenium Enabling Variations in Micronutrient Accumulation in Red Spinach



Abstract
:1. Introduction
2. Results
2.1. Spinach Plants Biomass
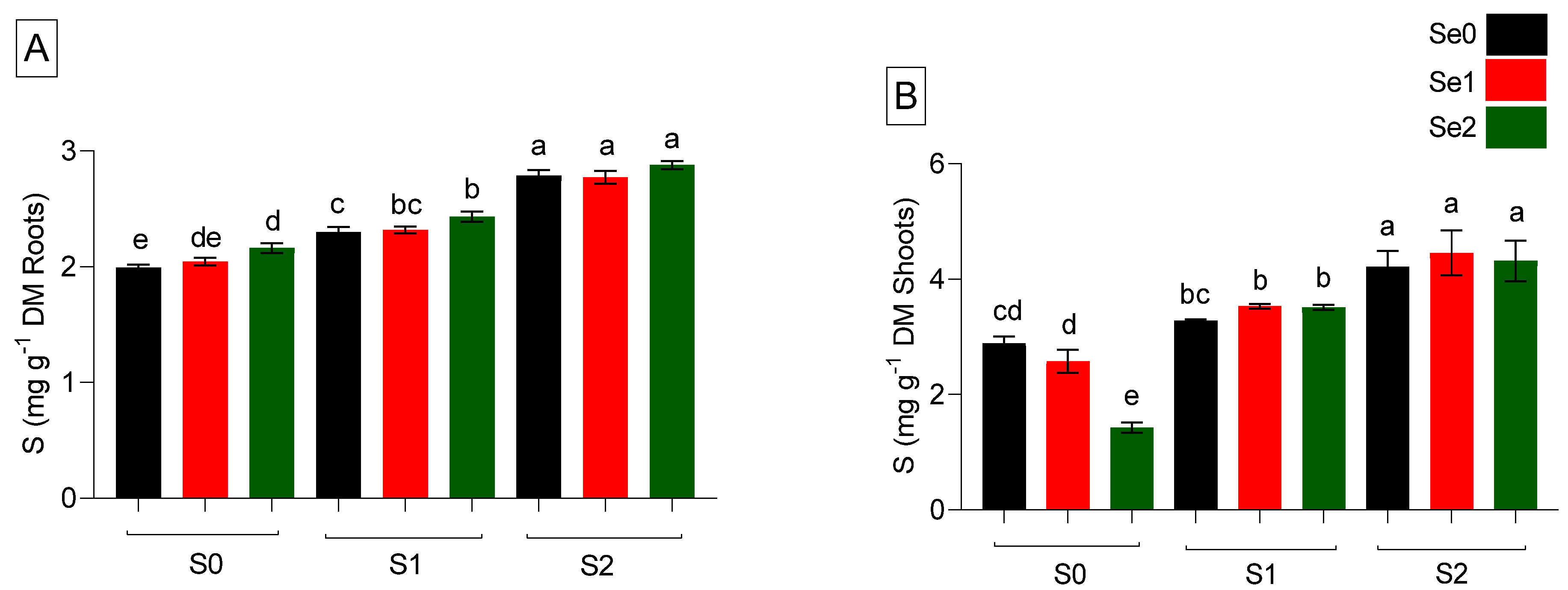
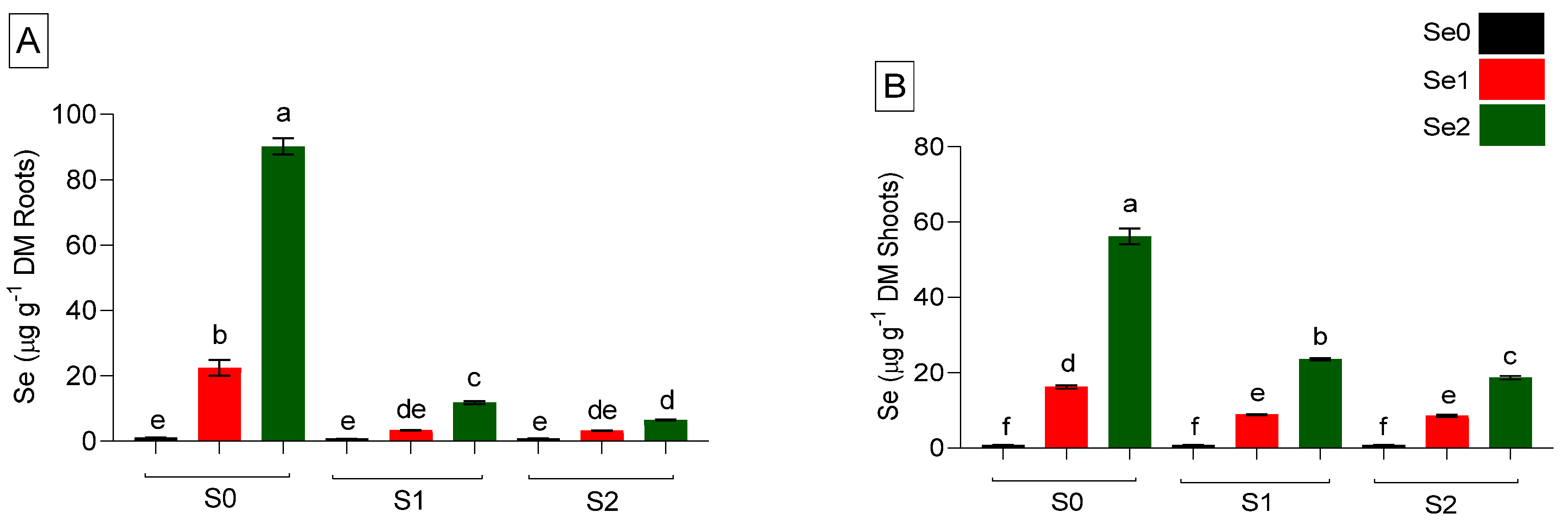
2.2. Sulfur and Selenium Accumulation in Plants
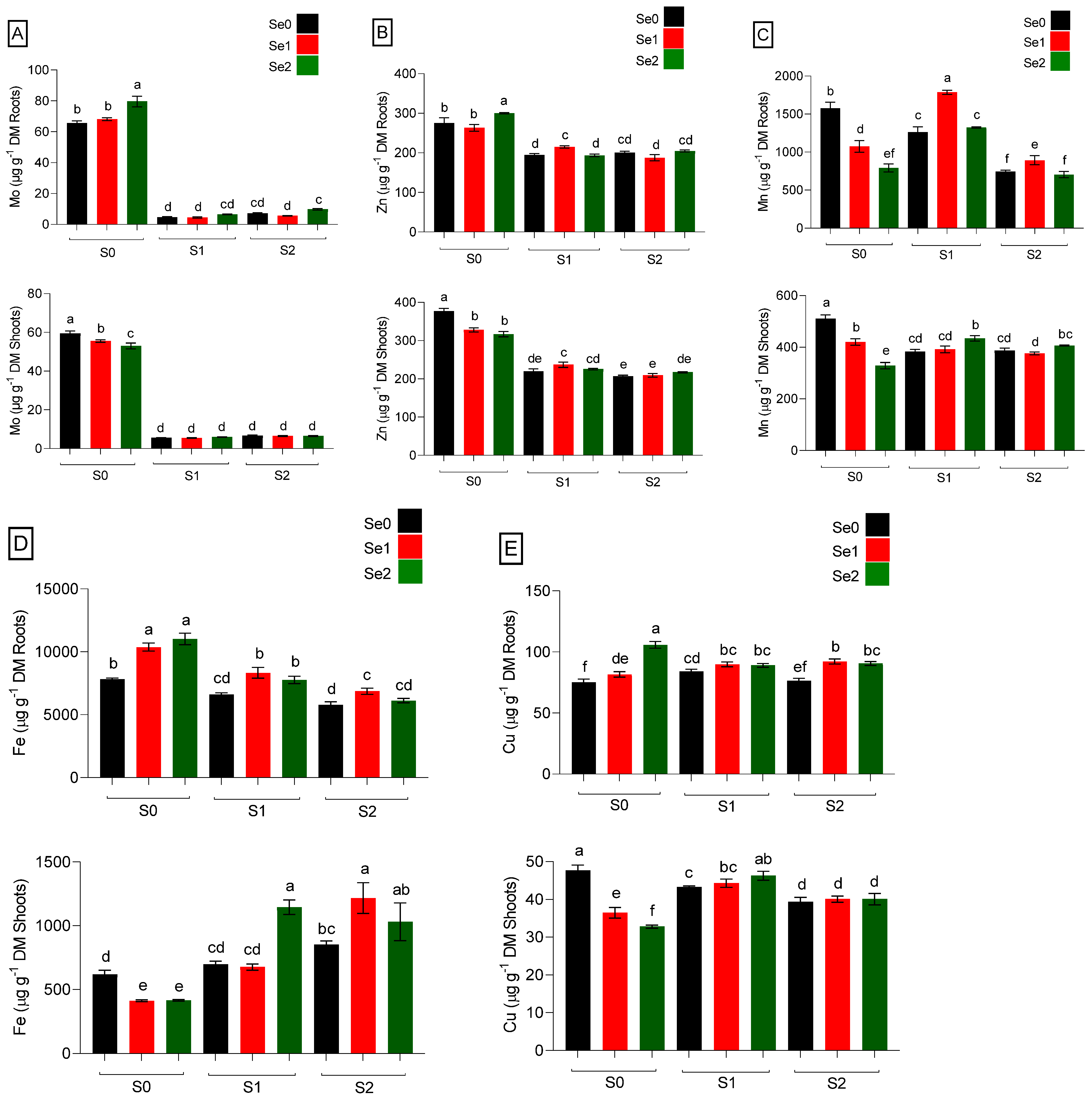
2.3. Micronutrients Accumulation in Spinach
2.4. Quantification of Organic Acids and Water-Soluble Sugars
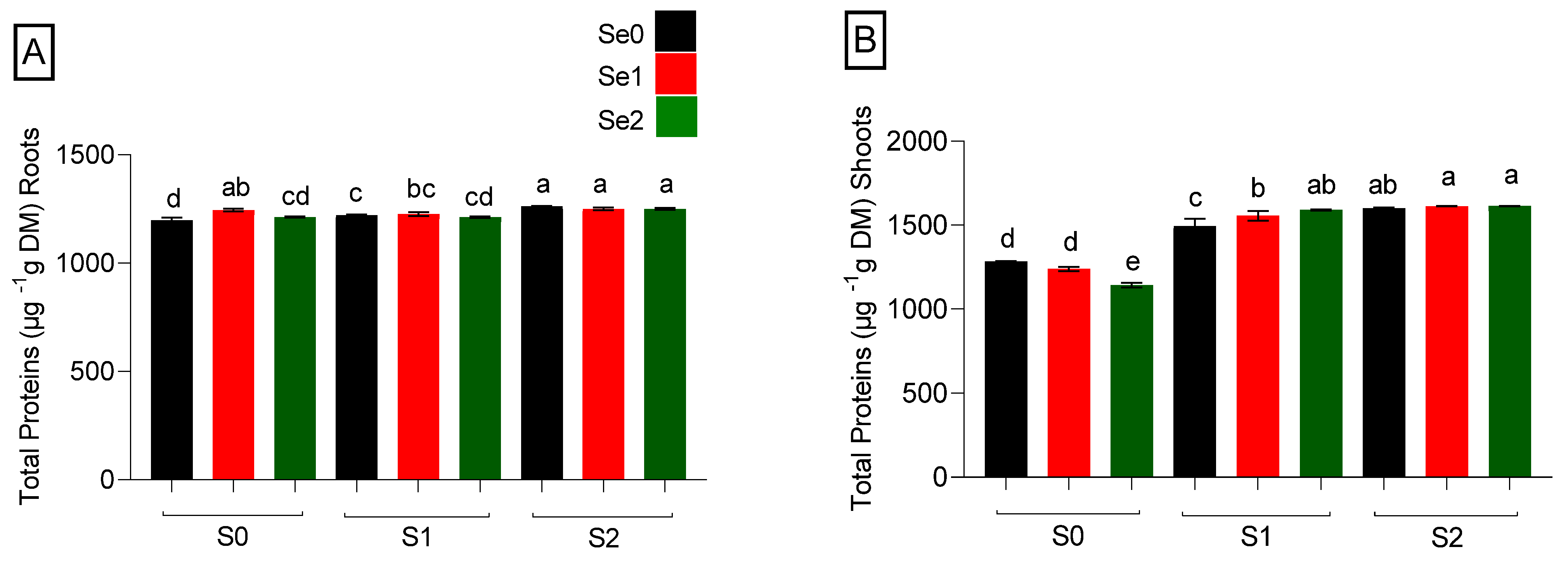
2.5. Quantification of Total Protein
3. Discussion
3.1. Se/S-Interaction
3.2. Role of Se on Micronutrient Accumulation
3.3. Importance of Se for Primary Metabolites and Food Quality
4. Materials and Methods
4.1. Experimental Design and Plant Material
4.2. Treatment Combinations and Chemical Characteristics
4.3. The Null Hypothesis and an Alternative Hypothesis
4.4. Mineral Analyses Using ICP-MS
4.5. Sulfur Determination
4.6. Determination of Organic Acids and Water-Soluble Sugars
4.7. Total Protein Quantification
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kopriva, S.; Talukdar, D.; Takahashi, H.; Hell, R.; Sirko, A.; D’Souza, S.F.; Talukdar, T. Editorial: Frontiers of Sulfur Metabolism in Plant Growth, Development, and Stress Response. Front. Plant Sci. 2016, 6, 1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdalla, M.A.; Mühling, K.H. Plant-derived sulfur containing natural products produced a response to biotic and abiotic stresses: A review of their structural diversity and medicinal importance. J. Appl. Bot. Food Qual. 2019, 92, 204–215. [Google Scholar]
- Abdalla, M.A.; Li, F.; Wenzel-Storjohann, A.; Sulieman, S.; Tasdemir, D.; Mühling, K.H. Comparative metabolite profile, biological activity and overall quality of three lettuce (Lactuca sativa L., Asteraceae) cultivars in response to sulfur nutrition. Pharmaceutics 2021, 13, 713. [Google Scholar] [CrossRef] [PubMed]
- Astolfi, S.; Celletti, S.; Vigani, G.; Mimmo, T.; Cesco, S. Interaction between sulfur and iron in plants. Front. Plant Sci. 2021, 12, 670308. [Google Scholar] [CrossRef]
- Gerendás, J.; Podestát, J.; Stahl, T.; Kübler, K.; Brückner, H.; Mersch-Sundermann, V.; Mühling, K.H. Interactive effects of sulfur and nitrogen supply on the concentration of sinigrin and allyl isothiocyanate in Indian mustard (Brassica juncea L.). J. Agric. Food Chem. 2009, 57, 3837–3844. [Google Scholar] [CrossRef]
- Marschner, H.; Marschner, P. Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Elsevier: London, UK; Academic Press: London, UK, 2012. [Google Scholar]
- Thapa, S.; Bhandari, A.; Ghimire, R.; Xue, Q.; Kidwaro, F.; Ghatrehsamani, S.; Maharjan, B.; Goodwin, M. Managing micronutrients for improving soil fertility, health, and soybean yield. J. Sustain. 2021, 13, 11766. [Google Scholar] [CrossRef]
- Zenda, T.; Liu, S.; Dong, A.; Duan, H. Revisiting Sulphur—The Once Neglected Nutrient: It’s Roles in Plant Growth, Metabolism, Stress Tolerance and Crop Production. Agriculture 2021, 11, 626. [Google Scholar] [CrossRef]
- Gupta, U.C.; Gupta, S.C. Selenium in soils and crops, its deficiencies in livestock and humans: Implications for management. Commun. Soil Sci. Plant Anal. 2000, 31, 1791–1807. [Google Scholar] [CrossRef]
- Fakhrolmobasheri, M.; Mazaheri-Tehrani, S.; Kieliszek, M.; Zeinalian, M.; Abbasi, M.; Karimi, F.; Mozafari, A.M. COVID-19 and Selenium Deficiency: A Systematic Review. Biol. Trace Elem. Res. 2022, 200, 3945–3956. [Google Scholar] [CrossRef]
- Barchielli, G.; Capperucci, A.; Tanini, D. The role of selenium in pathologies: An updated review. Antioxidants 2022, 11, 251. [Google Scholar] [CrossRef]
- Neysanian, M.; Iranbakhsh, A.; Ahmadvand, R.; Ardebili, Z.O.; Ebadi, M. Comparative efficacy of selenate and selenium nanoparticles for improving growth, productivity, fruit quality, and postharvest longevity through modifying nutrition, metabolism, and gene expression in tomato; potential benefits and risk assessment. PLoS ONE 2021, 16, e0250192. [Google Scholar] [CrossRef]
- Turakainen, M.; Hartikainen, H.; Ekholm, P.; Seppänen, M.M. Distribution of selenium in different biochemical fractions and raw darkening degree of potato (Solanum tuberosum L.) tubers supplemented with selenate. J. Agric. Food Chem. 2006, 54, 8617–8622. [Google Scholar] [CrossRef] [PubMed]
- Bañuelos, G.S.; Stushnoff, C.; Walse, S.S.; Zuber, T.; Yang, S.I.; Pickering, I.J.; Freeman, J.L. Biofortified, selenium enriched, fruit and cladode from three Opuntia Cactus pear cultivars grown on agricultural drainage sediment for use in nutraceutical foods. Food Chem. 2012, 135, 9–16. [Google Scholar] [CrossRef]
- Filek, M.; Zembala, M.; Kornaś, A.; Walas, S.; Mrowiec, H.; Hartikainen, H. The uptake and translocation of macro and micro elements in rape and wheat seedlings as affected by selenium supply level. Plant Soil 2010, 336, 303–312. [Google Scholar] [CrossRef]
- Wu, L.; Huang, Z. Selenium assimilation and nutrient element uptake in white clover and Tall Fescue under the influence of sulphate concentration and selenium tolerance of the plants. J. Exp. Bot. 1992, 43, 549–555. [Google Scholar] [CrossRef]
- Lima, L.W.; Checchio, M.V.; Reis, A.R.D.; Alves, R.D.C.; Tezzoto, T.; Gratão, P.L. Selenium restricts cadmium uptake and improve micronutrients and proline concentration in tomato fruits. Biocatal. Agric. Biotechnol. 2019, 18, 101057. [Google Scholar] [CrossRef]
- Xie, F.; Chen, C.; Chen, J.; Yuan, Y.; Hua, Q.; Zhang, Z.; Zhao, J.; Hu, G.; Chen, J.; Qin, Y. Metabolic Profiling of Sugars and Organic Acids, and Expression Analyses of Metabolism-Associated Genes in Two Yellow-Peel Pitaya Species. Plants 2022, 11, 694. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, Y.; Liu, J.; Chen, Y.; Zhang, X. Exploring the effects of selenium treatment on the nutritional quality of tomato fruit. Food Chem. 2018, 252, 9–15. [Google Scholar] [CrossRef]
- Rashid, M.; Yousaf, Z.; Din, A.; Munawar, M.; Aftab, A.; Riaz, N.; Younas, A.; Alaraidh, I.A.; Okla, M.K.; AbdElgawad, H. Assessment of mineral nutrient efficiency in genetically diverse spinach accessions by biochemical and functional marker strategies. Front. Plant Sci. 2022, 13, 889604. [Google Scholar] [CrossRef]
- Zhang, X.; Du, X.; Li, Y.Z.; Nie, C.N.; Wang, C.M.; Bian, J.L.; Luo, F. Are organic acids really related to the sour taste difference between Chinese black tea and green tea? Food Sci Nutr. 2022, 10, 2071–2081. [Google Scholar] [CrossRef]
- Zhu, Y.G.; Huang, Y.; Hu, Y.; Liu, Y.; Christie, P. Interaction between selenium and iodine uptake by spinach (Spinacia oleracea L.) in solution culture. Plant Soil 2004, 261, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Xue, T.; Hartikainen, H.; Piironen, V. Antioxidative and growth-promoting effects of selenium on senescing lettuce. Plant Soil 2001, 237, 55–61. [Google Scholar] [CrossRef]
- Turakainen, M.; Hartikainen, H.; Seppanen, M.M. Effects of selenium treatments on potato (Solanum tuberosum L.) growth and concentrations of soluble sugars and starch. J. Agric. Food Chem. 2004, 52, 5378–5382. [Google Scholar] [CrossRef]
- Smits, E.A.H.P.; Quinn, C.F.; Tapken, W.; Malagoli, M.; Schiavon, M. Physiological functions of beneficial elements. Curr. Opin. Plant Biol. 2009, 12, 267–274. [Google Scholar] [CrossRef]
- Narayan, O.P.; Kumar, P.; Yadav, B.; Dua, M.; Johri, A.K. Sulfur nutrition and its role in plant growth and development. Plant Signal. Behav. 2022, 2030082. [Google Scholar] [CrossRef]
- Saffaryazdi, A.; Lahouti, M.; Ganjeali, A.; Bayat, H. Impact of selenium supplementation on growth and selenium accumulation on spinach (Spinacia oleracea L.) plants. Not. Sci. Biol. 2012, 4, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Schiavon, M.; Pittarello, M.; Pilon-Smits, E.A.H.; Wirtz, M.; Hell, R.; Malagoli, M. Selenate and molybdate alter sulfate transport and assimilation in Brassica juncea L. Czern.: Implications for phytoremediation. Environ. Exp. Bot. 2012, 75, 41–51. [Google Scholar] [CrossRef]
- Cao, M.J.; Wang, Z.; Wirtz, M.; Hell, R.; Oliver, D.J.; Xiang, C.B. SULTR 3;1 is a chloroplast-localized sulfate transporter in Arabidopsis thaliana. Plant J. 2013, 73, 607–616. [Google Scholar] [CrossRef]
- Shinmachi, F.; Buchner, P.; Stroud, J.L.; Parmar, S.; Zhao, F.J.; McGrath, S.P.; Hawkesford, M.J. Influence of sulfur deficiency on the expression of specific sulfate transporters and the distribution of sulfur, selenium, and molybdenum in wheat. Plant Physiol. 2010, 153, 327–336. [Google Scholar] [CrossRef] [Green Version]
- White, P.J.; Bowen, H.C.; Parmaguru, P.; Fritz, M.; Spracklen, W.P.; Spiby, R.E.; Meacham, M.C.; Mead, A.; Harriman, M.; Trueman, L.J.; et al. Interactions between selenium and sulphur nutrition in Arabidopsis thaliana. J. Exp. Bot. 2004, 55, 1927–1937. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, H.; Takahashi, A.W.; Smith, F.W.; Kalff, M.B.; Hawkesford, M.J.; Saito, K. The roles of three functional sulphate transporters involved in uptake and translocation of sulphate in Arabidopsis thaliana. Plant J. 2000, 23, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Ramos, S.J.; Rutzke, M.A.; Hayes, R.J.; Faquin, V.; Guilherme, L.R.G.; Li, L. Selenium accumulation in lettuce germplasm. Planta 2011, 233, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Abdalla, M.A.; Wick, J.E.; Famuyide, I.M.; McGaw, L.J.; Mühling, K.H. Selenium enrichment of green and red lettuce and the induction of radical scavenging potential. Horticulturae 2021, 7, 488. [Google Scholar] [CrossRef]
- Liu, X.; Yang, Y.; Deng, X.; Li, M.; Zhang, W.; Zhao, Z. Effects of sulfur and sulfate on selenium uptake and quality of seeds in rapeseed (Brassica napus L.) treated with selenite and selenate. Environ. Exp. Bot. 2017, 135, 13–20. [Google Scholar] [CrossRef]
- Tian, M.; Hui, M.; Thannhauser, T.W.; Pan, S.; Li, L. Selenium-induced toxicity is counteracted by sulfur in broccoli (Brassica oleracea L. var. italica). Front Plant Sci. 2017, 8, 1425. [Google Scholar] [CrossRef] [Green Version]
- Kipp, A.P.; Strohm, D.; Brigelius-Flohé, R.; Schomburg, L.; Bechthold, A.; Leschik-Bonnet, E.; Heseker, H. German Nutrition Society (DGE) revised reference values for selenium intake. J. Trace Elem. Med. Biol. 2015, 32, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Ramachanderan, R.; Schaefer, B. Lipoic acid. ChemTexts 2019, 5, 18. [Google Scholar] [CrossRef]
- Kabata-Pendias, A.; Pendias, H. Trace Elements in Soils and Plants, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2001. [Google Scholar]
- Fargašová, A.; Pastierová, J.; Svetková, K. Effect of Se-metal pair combinations (Cd, Zn, Cu, Pb) on photosynthetic pigments production and metal accumulation in Sinapis alba L. seedlings. Plant Soil Environ. 2006, 52, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Nahar, K.; García-Caparrós, P.; Parvin, K.; Zulfiqar, F.; Ahmed, N.; Fujita, M. Selenium supplementation and crop plant tolerance to metal/metalloid toxicity. Front. Plant Sci. 2022, 12, 792770. [Google Scholar] [CrossRef]
- Arvy, M.P.; Thiersault, M.; Doireau, P. Relationships between selenium, micronutrients, carbohydrates, and alkaloid accumulation in Catharanthus roseus cells. J. Plant Nutr. 1995, 18, 1535–1546. [Google Scholar] [CrossRef]
- Baxter, I.; Muthukumar, B.; Park, H.C.; Buchner, P.; Lahner, B.; Danku, J.; Zhao, K.; Lee, J.; Hawkesford, M.J.; Guerinot, M.L.; et al. Variation in molybdenum content across broadly distributed populations of Arabidopsis thaliana is controlled by a mitochondrial molybdenum transporter (MOT1). PLoS Genet. 2008, 4, e1000004. [Google Scholar] [CrossRef] [Green Version]
- Alhendawi, R.A.; Kirkby, E.A.; Pilbeam, D.J. Evidence that sulfur deficiency enhances molybdenum transport in xylem sap of tomato plants. J. Plant Nutr. 2005, 28, 1347–1353. [Google Scholar] [CrossRef]
- Zhang, M.; Hu, C.; Zhao, X.; Tan, Q.; Sun, X.; Li, N. Impact of molybdenum on Chinese cabbage response to selenium in solution culture. Soil Sci. Plant Nutr. 2012, 58, 595–603. [Google Scholar] [CrossRef] [Green Version]
- Abdalla, M.A.; Lentz, C.; Mühling, K.H. Selenium and sulfur crosstalk is associated with changes in primary metabolism in lettuce plants grown under Se and S enrichment. Plants 2022, 11, 927. [Google Scholar] [CrossRef] [PubMed]
- Coolong, T.W.; Kopsell, D.A.; Kopsell, D.E.; Randle, W.M. Nitrogen and sulfur influence nutrient usage and accumulation in onion. J. Plant Nutr. 2005, 27, 1667–1686. [Google Scholar] [CrossRef]
- Singh, D.P.; Gulpadiya, V.K.; Chauhan, R.S.; Singh, S.P. Effect of sulphur on productivity, economics and nutrient uptake in spinach. Ann. Plant Soil Res. 2015, 17, 29–32. [Google Scholar]
- Makus, D.J.; Zibilske, L. Spinach and mustard greens response to soil texture, sulfur addition and lithium level. Subtrop. Plant Sci. J. 2009, 60, 69–77. [Google Scholar]
- Ali, J.; Singh, S.; Singh, S. Response of fababean to boron, zinc and sulphur application in Alluvial Soil. J. Indian Soc. Soil Sci. 2013, 61, 202–206. [Google Scholar]
- Longchamp, M.; Angeli, N.; Castrec-Rouelle, M. Effects on the accumulation of calcium, magnesium, iron, manganese, copper and zinc of adding the two inorganic forms of selenium to solution cultures of Zea mays. Plant Physiol. Biochem. 2016, 98, 128–137. [Google Scholar] [CrossRef]
- Narvaez-Ortiz, W.A.; Martínez-Hernández, M.; Fuentes-Lara, L.O.; Benavides-Mendoza, A.; Valenzuela-García, J.R.; Gonzalez-Fuentes, J.A. Effect of selenium application on mineral macro-and micronutrients and antioxidant status in strawberries. J. Appl. Bot. Food Qual. 2018, 91, 321–331. [Google Scholar]
- Feng, R.; Wei, C.; Tu, S.; Wu, F. Effects of Se on the uptake of essential elements in Pteris vittata L. Plant Soil 2009, 325, 123–132. [Google Scholar] [CrossRef]
- Drahoňovský, J.; Száková, J.; Mestek, O.; Tremlová, J.; Kaňa, A.; Najmanová, J.; Tlustoš, P. Selenium uptake, transformation and inter-element interactions by selected wildlife plant species after foliar selenate application. Environ. Exp. Bot. 2016, 125, 12–19. [Google Scholar] [CrossRef]
- De Souza, G.A.; De Carvalho, J.G.; Rutzke, M.; Albrecht, J.C.; Guilherme, L.R.G.; Li, L. Evaluation of germplasm effect on Fe, Zn and Se content in wheat seedlings. Plant Sci. 2013, 210, 206–213. [Google Scholar] [CrossRef] [PubMed]
- Tobiaz, A.; Walas, S.; Filek, M.; Mrowiec, H.; Samsel, K.; Sieprawska, A.; Hatikainen, H. Effect of selenium on distribution of macro and micro elements to different tissues during wheat ontogeny. Biol. Plant. 2014, 58, 370–374. [Google Scholar] [CrossRef]
- Kleiber, T.; Krzesiński, W.; Przygocka-Cyna, K.; Spiżewski, T. Alleviation effect of selenium on manganese stress of plants. Ecol. Chem. Eng. S 2018, 25, 143–152. [Google Scholar] [CrossRef] [Green Version]
- Pazurkiewicz-Kocot, K.; Galas, W.; Kita, A. The effect of selenium on the accumulation of some metals in Zea mays L. plants treated with indole-3-acetic acid. Cell. Mol. Biol. Lett. 2003, 8, 97–103. [Google Scholar]
- Zuchi, S.; Cesco, S.; Astolfi, S. High S supply improves Fe accumulation in durum wheat plants grown under Fe limitation. Environ. Exp. Bot. 2012, 77, 25–32. [Google Scholar] [CrossRef]
- Reich, M.; Shahbaz, M.; Prajapati, D.H.; Parmar, S.; Hawkesford, M.J.; De Kok, L.J. Interactions of sulfate with other nutrients as revealed by H2S fumigation of Chinese cabbage. Front. Plant Sci. 2016, 7, 541. [Google Scholar] [CrossRef] [Green Version]
- Ciaffi, M.; Paolacci, A.R.; Celletti, S.; Catarcione, G.; Kopriva, S.; Astolfi, S. Transcriptional and physiological changes in the S assimilation pathway due to single or combined S and Fe deprivation in durum wheat (Triticum durum L.) seedlings. J. Exp. Bot. 2013, 64, 1663–1675. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, K.J.; Budzyński, W.S.; Kijewski, Ł.; Klasa, A. Concentrations of copper, zinc and manganese in the roots, straw and oil cake of white mustard (Sinapis alba L.) and Indian mustard (Brassica juncea (L.) Czern. et Coss.) depending on sulphur fertilization. Plant Soil Environ. 2014, 60, 364–371. [Google Scholar] [CrossRef] [Green Version]
- National Institute of Health (NIH), Office of Dietary Supplements, U.S. Department of Health and Human Services. Available online: https://ods.od.nih.gov/HealthInformation/nutrientrecommendations.aspx#dri (accessed on 9 August 2023).
- Lidon, F.C.; Oliveira, K.; Ribeiro, M.M.; Pelica, J.; Pataco, I.; Ramalho, J.C.; Leitão, A.E.; Almeida, A.S.; Campos, P.S.; Ribeiro-Barros, A.I.; et al. Selenium biofortification of rice grains and implications on macronutrients quality. J. Cereal Sci. 2018, 81, 22–29. [Google Scholar] [CrossRef]
- Yin, N.; Mu, L.; Liang, Y.L.; Hao, W.L.; Yin, H.F.; Zhu, S.M.; An, X.J. Effects of foliar selenium fertilizer on fruit yield, quality and selenium content of three varieties of Vitis vinifera. J. Appl. Ecol. 2020, 31, 953–958. [Google Scholar]
- Batista-Silva, W.; Nascimento, V.L.; Medeiros, D.B.; Nunes-Nesi, A.; Ribeiro, D.M.; Zsögön, A.; Araújo, W.L. Modifications in organic acid profiles during fruit development and ripening: Correlation or causation? Front. Plant Sci. 2018, 871, 1689. [Google Scholar] [CrossRef] [PubMed]
- Ferrarese, M.; Sourestani, M.M.; Quattrini, E.; Schiavi, M.; Ferrante, A. Biofortification of spinach plants applying selenium in the nutrient solution of floating system. J. Fruit Ornam. Plant Res. 2012, 76, 127–136. [Google Scholar] [CrossRef]
- Tan, C.; Zhang, L.; Duan, X.; Chai, X.; Huang, R.; Kang, Y.; Yang, X. Effects of exogenous sucrose and selenium on plant growth, quality, and sugar metabolism of pea sprouts. J. Sci. Food Agric. 2022, 102, 2855–2863. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Liu, Z.; Han, Y.; Sun, Y. Exogenous dopamine promotes photosynthesis and carbohydrate metabolism of downy mildew-infected cucumber. Sci. Hortic. 2022, 295, 110842. [Google Scholar] [CrossRef]
- Hu, W.; Su, Y.; Zhou, J.; Zhu, H.; Guo, J.; Huo, H.; Gong, H. Foliar application of silicon and selenium improves the growth, yield and quality characteristics of cucumber in field conditions. Sci. Hortic. 2022, 294, 110776. [Google Scholar] [CrossRef]
- Samanta, S.; Singh, A.; Banerjee, A.; Roychoudhury, A. Exogenous supplementation of melatonin alters representative organic acids and enzymes of respiratory cycle as well as sugar metabolism during arsenic stress in two contrasting indica rice cultivars. J. Biotechnol. 2020, 324, 220–232. [Google Scholar] [CrossRef]
- Lv, J.; Dong, Y.; Dong, K.; Zhao, Q.; Yang, Z.; Chen, L. Intercropping with wheat suppressed Fusarium wilt in faba bean and modulated the composition of root exudates. Plant Soil 2020, 448, 153–164. [Google Scholar] [CrossRef]
- Lv, J.; Xiao, J.; Guo, Z.; Dong, K.; Dong, Y. Nitrogen supply and intercropping control of Fusarium wilt in faba bean depend on organic acids exuded from the roots. Sci. Rep. 2021, 11, 9589. [Google Scholar] [CrossRef]
- Kang, L.; Wu, Y.; Zhang, J.; An, Q.; Zhou, C.; Li, D.; Pan, C. Nano-selenium enhances the antioxidant capacity, organic acids and cucurbitacin B in melon (Cucumis melo L.) plants. Ecotoxicol. Environ. Saf. 2022, 241, 113777. [Google Scholar] [CrossRef] [PubMed]
- Kopsell, D.A.; Randle, W.M. Selenate concentration affects selenium and sulfur uptake and accumulation by ‘Granex33’ onions. J. Amer. Soc. Hort. Sci. 1997, 122, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Honsel, A.; Kojima, M.; Haas, R.; Frank, W.; Sakakibara, H.; Herschbach, C.; Rennenberg, H. Sulphur limitation and early sulphur deficiency responses in poplar: Significance of gene expression, metabolites, and plant hormones. J. Exp. Bot. 2012, 63, 1873–1893. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lei, N.; Xiong, Y.; Liu, Y.; Tong, L.; Wang, F.; Fan, B.; Maesen, P.; Blecker, C. Influence of selenium biofortification of soybeans on speciation and transformation during seed germination and sprouts quality. Foods 2022, 11, 1200. [Google Scholar] [CrossRef] [PubMed]
- Jezek, M.; Geilfus, C.M.; Bayer, A.; Mühling, K.H. Photosynthetic capacity, nutrient status, and growth of maize (Zea mays L.) upon MgSO4 leaf-application. Front. Plant Sci. 2015, 5, 781. [Google Scholar] [CrossRef] [Green Version]
- Zörb, C.; Mühling, K.H.; Hasler, M.; Gödde, V.; Niehaus, K.; Becker, D.; Geilfus, C.M. Metabolomics responses in grain, ear, and straw of winter wheat under increasing sulfur treatment. J. Plant. Nutr. Soil Sci. 2013, 176, 964–970. [Google Scholar] [CrossRef]
- Cataldi, T.R.; Margiotta, G.; Iasi, L.; Di-Chio, B.; Xiloyannis, C.; Bufo, S.A. Determination of sugar compounds in olive plant extracts by anion-exchange chromatography with pulsed amperometric detection. Anal. Chem. 2000, 72, 3902–3907. [Google Scholar] [CrossRef]
- Damerval, C.; De Vienne, D.; Zivy, M.; Thiellement, H. Technical improvements in two-dimensional electrophoresis increase the level of genetic variation detected in wheat-seedling proteins. Electrophoresis 1986, 7, 52–54. [Google Scholar] [CrossRef]
- Zörb, C.; Schmitt, S.; Neeb, A.; Karl, S.; Linder, M.; Schubert, S. The biochemical reaction of maize (Zea mays L.) to salt stress is characterized by a mitigation of symptoms and not by a specific adaptation. Plant Sci. 2004, 167, 91–100. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Roots | Shoots | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant Biomass (g Plant−1) | Organic Acids (mg g−1 DM) | Water-Soluble Sugars (mg g−1 DM) | Plant Biomass (g) | Organic Acids (mg g−1 DM) | Water-Soluble Sugars (mg g−1 DM) | ||||||||||
| DM | Malate | Oxalate | Citrate | Glucose | Fructose | Sucrose | DM | Malate | Oxalate | Citrate | Glucose | Fructose | Sucrose | ||
| S0 | Se0 | 0.85 ± 0.02 de | 1.8 ± 0.17 de | 24.9 ± 0.42 bc | 1.8 ± 0.08 ef | 1.7 ± 0.37 d | 0.7 ± 0.08 d | 8.1 ± 0.13 ab | 2.78 ± 0.05 f | 7.3 ± 0.41 d | 115.9 ± 2.69 d | 2.9 ± 0.16 c | 7.6 ± 0.60 d | 2.6 ± 0.11 d | 7.2 ± 0.45 cde |
| Se1 | 0.95 ± 0.01 d | 1.5 ± 0.11 ef | 24.0 ± 0.58 bc | 2.9 ± 0.31 a | 2.2 ± 0.18 d | 1.3 ± 0.10 d | 8.6 ± 0.60 ab | 3.64 ± 0.04 e | 13.7 ± 1.80 b | 116.1 ± 0.92 d | 6.4 ± 0.03 b | 23.0 ± 1.17 a | 5.3 ± 0.27 a | 10.9 ± 0.97 b | |
| Se2 | 0.36 ± 0.01 f | 1.2 ± 0.12 f | 25.7 ± 0.49 b | 3.0 ± 0.17 a | 1.3 ± 0.10 d | 0.7 ± 0.08 d | 9.4 ± 0.93 a | 3.03 ± 0.05 f | 11.7 ± 1.09 b | 115.1 ± 1.15 d | 11.6 ± 0.54 a | 13.2 ± 0.67 b | 2.9 ± 0.23 cd | 75.9 ± 2.80 a | |
| S1 | Se0 | 1.51 ± 0.03 b | 2.6 ± 0.06 c | 31.5 ± 0.87 a | 2.2 ± 0.21 bcd | 1.9 ± 0.14 d | 1.2 ± 0.06 d | 7.3 ± 0.26 bc | 8.42 ± 0.13 d | 7.8 ± 1.62 cd | 154.7 ± 3.34 a | 1.1 ± 0.30 d | 6.3 ± 1.16 def | 2.9 ± 0.34 cd | 6.7 ± 0.56 de |
| Se1 | 1.38 ± 0.01 b | 2.6 ± 0.13 c | 29.8 ± 1.14 a | 1.5 ± 0.04 f | 1.7 ± 0.24 d | 0.8 ± 0.05 d | 3.1 ± 0.35 e | 9.37 ± 0.06 c | 2.4 ± 0.02 e | 131.5 ± 2.10 c | 1.0 ± 0.18 d | 7.5 ± 0.54 de | 3.6 ± 0.13 b | 8.2 ± 0.45 bcd | |
| Se2 | 0.67 ± 0.02 e | 5.0 ± 0.05 a | 25.7 ± 0.44 b | 2.6 ± 0.10 ab | 4.8 ± 0.41 c | 2.3 ± 0.19 c | 6.0 ± 0.54 cd | 9.93 ± 0.16 b | 13.3 ± 1.58 bc | 128.6 ± 1.02 c | 2.7 ± 0.18 c | 10.0 ± 0.47 c | 5.6 ± 0.21 a | 11.0 ± 0.54 b | |
| S2 | Se0 | 1.81 ± 0.02 a | 2.0 ± 0.24 d | 23.3 ± 0.41 c | 2.1 ± 0.13 cde | 3.6 ± 0.50 c | 2.2 ± 0.27 c | 5.9 ± 0.16 cd | 10.37 ± 0.10 a | 18.1 ± 0.48 a | 134.5 ± 1.83 c | 3.0 ± 0.31 c | 5.4 ± 0.19 f | 3.3 ± 0.05 bc | 7.4 ± 0.22 cde |
| Se1 | 1.17 ± 0.18 c | 4.4 ± 0.21 b | 24.5 ± 0.72 bc | 2.4 ± 0.24 bc | 8.0 ± 0.71 b | 6.0 ± 0.46 b | 7.7 ± 0.26 b | 10.39 ± 0.17 a | 21.0 ± 0.88 a | 135.0 ± 3.40 c | 3.2 ± 0.32 c | 6.3 ± 0.28 def | 3.3 ± 0.09 bc | 10.0 ± 0.68 bc | |
| Se2 | 0.45 ± 0.01 f | 2.7 ± 0.10 c | 23.2 ± 0.55 c | 1.9 ± 0.12 def | 11.8 ± 0.68 a | 10.4 ± 0.58 a | 4.9 ± 1.05 d | 9.72 ± 0.08 b | 11.1 ± 1.60 bc | 142.4 ± 2.86 b | 3.1 ± 0.23 c | 5.5 ± 0.22 ef | 3.4 ± 0.20 bc | 4.8 ± 0.34 e | |
| Micronutrients | Male | Female |
|---|---|---|
| Fe | 8 mg day−1 | 18 mg day−1 |
| Zn | 11 mg day−1 | 8 mg day−1 |
| Mn | 2.3 mg day−1 | 1.8 mg day−1 |
| Cu | 900 µg day−1 | 900 µg day−1 |
| Mo | 45 µg day−1 | 45 µg day−1 |
| Micronutrients | Se1S1 | Se2S1 | Se1S2 | Se2S2 |
|---|---|---|---|---|
| Fe | 6 mg day−1 | 10.3 mg day−1 | 10.9 mg day−1 | 9.2 mg day−1 |
| Zn | 2.1 mg day−1 | 2 mg day−1 | 1.8 mg day−1 | 1.9 mg day−1 |
| Mn | 3.5 mg day−1 | 3.9 mg day−1 | 3.3 mg day−1 | 3.6 mg day−1 |
| Cu | 397.8 µg day−1 | 415.8 µg day−1 | 360.9 µg day−1 | 360 µg day−1 |
| Mo | 48.6 µg day−1 | 53.1 µg day−1 | 58.5 µg day−1 | 58.5 µg day−1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, K.; Nisa, F.K.; Abdalla, M.A.; Mühling, K.H. The Interplay of Sulfur and Selenium Enabling Variations in Micronutrient Accumulation in Red Spinach. Int. J. Mol. Sci. 2023, 24, 12766. https://doi.org/10.3390/ijms241612766
Saeed K, Nisa FK, Abdalla MA, Mühling KH. The Interplay of Sulfur and Selenium Enabling Variations in Micronutrient Accumulation in Red Spinach. International Journal of Molecular Sciences. 2023; 24(16):12766. https://doi.org/10.3390/ijms241612766
Chicago/Turabian StyleSaeed, Kashif, Fatiha Kalam Nisa, Muna Ali Abdalla, and Karl Hermann Mühling. 2023. "The Interplay of Sulfur and Selenium Enabling Variations in Micronutrient Accumulation in Red Spinach" International Journal of Molecular Sciences 24, no. 16: 12766. https://doi.org/10.3390/ijms241612766