

Current Understanding of Human Polymorphism in Selenoprotein Genes: A Review of Its Significance as a Risk Biomarker

Abstract
:1. Introduction
2. Glutathione Peroxidases
3. Selenoproteins
4. Thioredoxin Reductases
5. Iodothyronine Deiodinases
6. Selenoproteins and Their Polymorphisms
6.1. Glutathione Peroxidases
6.1.1. rs1050450 Polymorphism
6.1.2. Alanine Repetition Polymorphism
6.1.3. rs1800668 Polymorphism
6.1.4. GPx3 Polymorphism
6.1.5. GPx4 Polymorphism
6.2. Selenoprotein P
6.3. Selenoprotein F
6.4. Selenoprotein S
6.5. Thioredoxin Reductase
6.6. Iodothyronine Deiodinases
6.7. Selenophosphate Synthetase
7. Conclusions
Funding
Conflicts of Interest
References
- Mehdi, Y.; Hornick, J.-L.; Istasse, L.; Dufrasne, I. Selenium in the environment, metabolism and involvement in body functions. Molecules 2013, 18, 3292–3311. [Google Scholar] [CrossRef]
- Santi, C.; Bagnoli, L. Celebrating Two Centuries of Research in Selenium Chemistry: State of the Art and New Prospective. Molecules 2017, 22, 2124. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, K.; Foltz, C.M. Selenium as an integral part of factor 3 against dietary necrotic liver degeneration. J. Am. Chem. Soc. 1957, 79, 3292–3293. [Google Scholar] [CrossRef]
- Schwarz, K. Role of vitamin E, selenium, and related factors in experimental nutritional liver disease. Fed. Proc. 1965, 24, 58–67. [Google Scholar]
- Rocha, J.B.; Piccoli, B.C.; Oliveira, C.S. Biological and chemical interest in selenium: A brief historical account. Arch. Org. Chem. 2017, 24, 457–491. [Google Scholar] [CrossRef]
- Kang, D.; Lee, J.; Wu, C.; Guo, X.; Lee, B.J.; Chun, J.-S.; Kim, J.-H. The role of selenium metabolism and selenoproteins in cartilage homeostasis and arthropathies. Exp. Mol. Med. 2020, 52, 1198–1208. [Google Scholar] [CrossRef]
- Ha, H.Y.; Alfulaij, N.; Berry, M.J.; Seale, L.A. From Selenium Absorption to Selenoprotein Degradation. Biol. Trace Elem. Res. 2019, 192, 26–37. [Google Scholar] [CrossRef] [PubMed]
- Alexander, J. Chapter 29–Selenium. In Handbook on the Toxicology of Metals, 5th ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 729–771. [Google Scholar]
- Shreenath, A.P.; Ameer, M.A.; Dooley, J. Selenium Deficiency. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Belay, A.; Joy, E.J.M.; Chagumaira, C.; Zerfu, D.; Ander, E.L.; Young, S.D.; Bailey, E.H.; Lark, R.M.; Broadley, M.R.; Gashu, D. Selenium Deficiency Is Widespread and Spatially Dependent in Ethiopia. Nutrients 2020, 12, 1565. [Google Scholar] [CrossRef] [PubMed]
- Stoffaneller, R.; Morse, N.L. A review of dietary selenium intake and selenium status in Europe and the Middle East. Nutrients 2015, 7, 1494–1537. [Google Scholar] [CrossRef]
- Alehagen, U.; Johansson, P.; Björnstedt, M.; Rosén, A.; Post, C.; Aaseth, J. Relatively high mortality risk in elderly Swedish subjects with low selenium status. Eur. J. Clin. Nutr. 2016, 70, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.-W.; Shu, X.-O.; Li, H.-L.; Zhang, W.; Gao, J.; Zhao, L.-G.; Zheng, W.; Xiang, Y.-B. Dietary selenium intake and mortality in two population-based cohort studies of 133 957 Chinese men and women. Public Health Nutr. 2016, 19, 2991–2998. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Yao, H.; Wu, Y.; Sun, G.; Yang, W.; Li, Z.; Shang, L. Status and risks of selenium deficiency in a traditional selenium-deficient area in Northeast China. Sci. Total Environ. 2021, 762, 144103. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Wang, P.; Teng, T.; Shi, B.; Shan, A.; Lei, X.G. Effects of Dietary Selenium Deficiency or Excess on Selenoprotein Gene Expression in the Spleen Tissue of Pigs. Animals 2019, 9, 1122. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, X.; Yao, Y.; Zhao, X.; Shi, X.; Cai, Y. Selenium Deficiency Induces Apoptosis and Necroptosis Through ROS/MAPK Signal in Human Uterine Smooth Muscle Cells. Biol. Trace Elem. Res. 2022, 200, 3147–3158. [Google Scholar] [CrossRef] [PubMed]
- Lai, H.; Nie, T.; Zhang, Y.; Chen, Y.; Tao, J.; Lin, T.; Ge, T.; Li, F.; Li, H. Selenium Deficiency-Induced Damage and Altered Expression of Mitochondrial Biogenesis Markers in the Kidneys of Mice. Biol. Trace Elem. Res. 2021, 199, 185–196. [Google Scholar] [CrossRef]
- Zhang, L.; Gao, Y.; Feng, H.; Zou, N.; Wang, K.; Sun, D. Effects of selenium deficiency and low protein intake on the apoptosis through a mitochondria-dependent pathway. J. Trace Elem. Med. Biol. 2019, 56, 21–30. [Google Scholar] [CrossRef]
- Wang, J.; Liu, Z.; He, X.; Lian, S.; Liang, J.; Yu, D.; Sun, D.; Wu, R. Selenium deficiency induces duodenal villi cells apoptosis via oxidative stress-induced mitochondrial apoptosis pathway and inflammatory signaling-induced death receptor pathway. Metallomics 2018, 10, 1390–1400. [Google Scholar] [CrossRef]
- Osaki, Y.; Nishino, I.; Murakami, N.; Matsubayashi, K.; Tsuda, K.; Yokoyama, Y.I.; Morita, M.; Onishi, S.; Goto, Y.I.; Nonaka, I. Mitochondrial abnormalities in selenium-deficient myopathy. Muscle Nerve 1998, 21, 637–639. [Google Scholar] [CrossRef]
- Moghaddam, A.; Heller, R.A.; Sun, Q.; Seelig, J.; Cherkezov, A.; Seibert, L.; Hackler, J.; Seemann, P.; Diegmann, J.; Pilz, M.; et al. Selenium Deficiency Is Associated with Mortality Risk from COVID-19. Nutrients 2020, 12, 2098. [Google Scholar] [CrossRef]
- Skesters, D.; Kustovs, A.; Lece, E.; Moreino, E.; Petrosina, K.; Rainsford, D. Selenium, selenoprotein P, and oxidative stress levels in SARS-CoV-2 patients during illness and recovery. Infammopharmacology 2022, 30, 499–503. [Google Scholar] [CrossRef]
- Taylor, E.W.; Ruzicka, J.A.; Premadasa, L.; Zhao, L. Cellular Selenoprotein mRNA Tethering via Antisense Interactions with Ebola and HIV-1 mRNAs May Impact Host Selenium Biochemistry. Curr. Top. Med. Chem. 2016, 16, 1530–1535. [Google Scholar] [CrossRef] [PubMed]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef]
- Gladyshev, V.N.; Arnér, E.S.; Berry, M.J.; Brigelius-Flohé, R.; Bruford, E.A.; Burk, R.F.; Carlson, B.A.; Castellano, S.; Chavatte, L.; Conrad, M.; et al. Selenoprotein Gene Nomenclature. J. Biol. Chem. 2016, 291, 24036–24040. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.L.; Mendelev, N.; Kumari, S.; Li, P.A. Overexpression of human selenoprotein H in neuronal cells enhances mitochondrial biogenesis and function through activation of protein kinase A, protein kinase B, and cyclic adenosine monophosphate response element-binding protein pathway. Int. J. Biochem. Cell. Biol. 2013, 45, 604–611. [Google Scholar] [CrossRef] [PubMed]
- Roman, M.; Jitaru, P.; Barbante, C. Selenium biochemistry and its role for human health. Metallomics 2014, 6, 25–54. [Google Scholar] [CrossRef]
- El Haddad, M.; Jean, E.; Turki, A.; Hugon, G.; Vernus, B.; Bonnieu, A.; Passerieux, E.; Hamade, A.; Mercier, J.; Laoudj-Chenivesse, D.; et al. Glutathione peroxidase 3, a new retinoid target gene, is crucial for human skeletal muscle precursor cell survival. J. Cell. Sci. Dec. 2012, 125, 6147–6156. [Google Scholar] [CrossRef]
- Chang, C.; Worley, B.L.; Phaëton, R.; Hempel, N. Extracellular Glutathione Peroxidase GPx3 and Its Role in Cancer. Cancers 2020, 12, 2197. [Google Scholar] [CrossRef]
- Forcina, G.C.; Dixon, S.J. GPX4 at the Crossroads of Lipid Homeostasis and Ferroptosis. Proteomics 2019, 19, e1800311. [Google Scholar] [CrossRef]
- Friedmann Angeli, J.P.; Schneider, M.; Proneth, B.; Tyurina, Y.Y.; Tyurin, V.A.; Hammond, V.J.; Herbach, N.; Aichler, M.; Walch, A.; Eggenhofer, E.; et al. Inactivation of the ferroptosis regulator Gpx4 triggers acute renal failure in mice. Nat. Cell Biol. 2014, 16, 1180–1191. [Google Scholar] [CrossRef]
- Ursini, F. Dual Function of the Selenoprotein PHGPx During Sperm Maturation. Science 1999, 285, 1393–1396. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, K.; Zhang, D.; Zhao, Z.; Huang, J.; Zhou, L.; Feng, M.; Shi, J.; Wei, H.; Li, L.; et al. GPx6 is involved in the in vitro induced capacitation and acrosome reaction in porcine sperm. Theriogenology 2020, 156, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.P.; Shah, R.; Copeland, P.R. Regulation of selenocysteine incorporation into the selenium transport protein, selenoprotein P. J. Biol. Chem. 2014, 289, 25317–25326. [Google Scholar] [CrossRef] [PubMed]
- Motsenbocker, M.A.; Tappel, A. A selenocysteine-containing selenium-transport protein in rat plasma. Biochim. Et Biophys. Acta (BBA)–General. Subj. 1982, 719, 147–153. [Google Scholar] [CrossRef]
- Tsutsumi, R.; Saito, Y. Selenoprotein P; P for Plasma, Prognosis, Prophylaxis, and More. Biol. Pharm. Bull. 2020, 43, 366–374. [Google Scholar] [CrossRef] [PubMed]
- Caito, S.W.; Milatovic, D.; Hill, K.E.; Aschner, M.; Burk, R.F.; Valentine, W.M. Progression of neurodegeneration and morphologic changes in the brains of juvenile mice with selenoprotein P deleted. Brain Res. 2011, 1398, 1–12. [Google Scholar] [CrossRef]
- Gladyshev, V.N.; Jeang, K.-T.; Wootton, J.C.; Hatfield, D.L. A new human selenium-containing protein. Purification, characterization, and cDNA sequence. J. Biol. Chem. 1998, 273, 8910–8915. [Google Scholar] [CrossRef]
- Gao, Y.; Hannan, N.R.; Wanyonyi, S.; Konstantopolous, N.; Pagnon, J.; Feng, H.C.; Jowett, J.B.; Kim, K.-H.; Walder, K.; Collier, G.R. Activation of the selenoprotein SEPS1 gene expression by pro-inflammatory cytokines in HepG2 cells. Cytokine 2006, 33, 246–251. [Google Scholar] [CrossRef]
- Alkan, Z.; Duong, F.L.; Hawkes, W.C. Selenoprotein W controls epidermal growth factor receptor surface expression, activation and degradation via receptor ubiquitination. Biochim. Biophys. Acta 2015, 1853, 1087–1095. [Google Scholar] [CrossRef]
- Mariotti, M.; Ridge, P.G.; Zhang, Y.; Lobanov, A.V.; Pringle, T.H.; Guigo, R.; Hatfield, D.L.; Gladyshev, V.N. Composition and evolution of the vertebrate and mammalian selenoproteomes. PLoS ONE 2012, 7, e33066. [Google Scholar] [CrossRef]
- Zhang, X.; Xiong, W.; Chen, L.L.; Huang, J.Q.; Lei, X.G. Selenoprotein V protects against endoplasmic reticulum stress and oxidative injury induced by pro-oxidants. Free. Radic. Biol. Med. 2020, 160, 670–679. [Google Scholar] [CrossRef]
- Chen, L.L.; Huang, J.Q.; Wu, Y.Y.; Chen, L.B.; Li, S.P.; Zhang, X.; Wu, S.; Ren, F.Z.; Lei, X.G. Loss of Selenov predisposes mice to extra fat accumulation and attenuated energy expenditure. Redox Biol. 2021, 45, 102048. [Google Scholar] [CrossRef]
- Sun, W.; Wang, X.; Zou, X.; Song, R.; Du, X.; Hu, J.; Xiong, Y. Selenoprotein P gene r25191g/a polymorphism and quantification of selenoprotein P mRNA level in patients with Kashin-Beck disease. Br. J. Nutr. 2010, 104, 1283–1287. [Google Scholar] [CrossRef]
- Carducci, F.; Ardiccioni, C.; Fiorini, R.; Vignini, A.; Di Paolo, A.; Alia, S.; Barucca, M.; Biscotti, M.A. The ALA5/ALA6/ALA7 repeat polymorphisms of the glutathione peroxidase-1 (GPx1) gene and autism spectrum disorder. Autism Res. 2022, 15, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Ming, X.; Johnson, W.G.; Stenroos, E.S.; Mars, A.; Lambert, G.H.; Buyske, S. Genetic variant of glutathione peroxidase 1 in autism. Brain Dev. 2010, 32, 105–109. [Google Scholar] [CrossRef] [PubMed]
- Jefferies, S.; Kote-Jarai, Z.; Goldgar, D.; Houlston, R.; Frazer-Williams, M.J.; A’Hern, R.; Eeles, R.; Henk, J.; Gore, M.; Rhys-Evans, P.; et al. MPT Collaborators, Association between polymorphisms of the GPX1 gene and second primary tumours after index squamous cell cancer of the head and neck. Oral. Oncol. 2005, 41, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Kote-Jarai, Z.; Durocher, F.; Edwards, S.; Hamoudi, R.; Jackson, A.R.; Ardern-Jones, A.; Murkin, A.; Dearnaley, D.P.; Kirby, R.; Houlston, R.; et al. CRC/BPG UK Familial Prostrate Cancer Collaborators, Association between the GCG polymorphism of the selenium dependent GPX1 gene and the risk of young onset prostate cancer. Prostate Cancer Prostatic Dis. 2002, 5, 189–192. [Google Scholar] [CrossRef]
- Sabet, E.E.; Salehi, Z.; Khodayari, S.; Zarafshan, S.S.; Zahiri, Z. Polymorphisms of glutathione peroxidase 1 (GPX1 Pro198Leu) and catalase (CAT C-262T) in women with spontaneous abortion. Syst. Biol. Reprod. Med. 2014, 60, 304–307. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.V.; Cardoso, B.R.; Zavarize, B.; Almondes, K.; Bordon, I.; Hare, D.J.; Teixeira Favaro, D.I.; Cozzolino, S.M.F. GPX1 Pro198Leu polymorphism and GSTM1 deletion do not affect selenium and mercury status in mildly exposed Amazonian women in an urban population. Sci. Total. Environ. 2016, 571, 801–808. [Google Scholar] [CrossRef]
- Cardoso, B.R.; Ong, T.P.; Jacob-Filho, W.; Jaluul, O.; Freitas, M.I.; Cominetti, C.; Cozzolino, S.M. Glutathione peroxidase 1 Pro198Leu polymorphism in Brazilian Alzheimer’s disease patients: Relations to the enzyme activity and to selenium status. J. Nutr. Nutr. 2012, 5, 72–80. [Google Scholar] [CrossRef]
- Rocha, T.J.; Silva Alves, M.; Guisso, C.C.; Andrade, F.M.; Camozzato, A.; Oliveira, A.A.; Fiegenbaum, M. Association of GPX1 and GPX4 polymorphisms with episodic memory and Alzheimer’s disease. Neurosci. Lett. 2018, 666, 32–37. [Google Scholar] [CrossRef]
- Ichimura, Y.; Habuchi, T.; Tsuchiya, N.; Wang, L.; Oyama, C.; Sato, K.; Nishiyama, H.; Ogawa, O.; Kato, T. Increased risk of bladder cancer associated with a glutathione peroxidase 1 codon 198 variant. J. Urol. 2004, 172, 728–732. [Google Scholar] [CrossRef]
- Hadami, K.; El Hassani, R.A.; Ameur, A.; Dakka, N.; Abbar, M.; Al Bouzidi, A.; Attaleb, M.; El Mzibri, M. Association between GPX1 Pro189Leu polymorphism and the occurrence of bladder cancer in Morocco. Cell. Mol. Biol. 2016, 62, 38–43. [Google Scholar] [CrossRef]
- Paz-y-Miño, C.; Muñoz, M.J.; López-Cortés, A.; Cabrera, A.; Palacios, A.; Castro, B.; Paz-y-Miño, N.; Sánchez, M.E. Frequency of polymorphisms pro198leu in GPX-1 gene and ile58thr in MnSOD gene in the altitude Ecuadorian population with bladder cancer. Oncol. Res. 2010, 18, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Liang, D.; Grossman, H.B.; Wu, X. Glutathione peroxidase 1 gene polymorphism and risk of recurrence in patients with superficial bladder cancer. Urology 2005, 66, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Nikic, P.; Dragicevic, D.; Jerotic, D.; Savic, S.; Djukic, T.; Stankovic, B.; Kovacevic, L.; Simic, T.; Matic, M. Polymorphisms of Antioxidant Enzymes SOD2 (rs4880) and GPX1 (rs1050450) Are Associated with Bladder Cancer Risk or Its Aggressiveness. Medicina 2023, 59, 131. [Google Scholar] [CrossRef] [PubMed]
- Habyarimana, T.; Bakri, Y.; Mugenzi, P.; Mazarati, J.B.; Attaleb, M.; El Mzibri, M. Association between glutathione peroxidase 1 codon 198 variant and the occurrence of breast cancer in Rwanda. Mol. Genet. Genom. Med. 2018, 6, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Gammon, M.D.; Santella, R.M.; Gaudet, M.M.; Britton, J.A.; Teitelbaum, S.L.; Terry, M.B.; Neugut, A.I.; Ambrosone, C.B. No association between glutathione peroxidase Pro198Leu polymorphism and breast cancer risk. Cancer Epidemiol. Biomark. Prev. 2005, 14, 2459–2461. [Google Scholar] [CrossRef]
- Cox, D.G.; Hankinson, S.E.; Kraft, P.; Hunter, D.J. No association between GPX1 Pro198Leu and breast cancer risk. Cancer Epidemiol. Biomark. Prev. 2004, 13, 1821–1822. [Google Scholar] [CrossRef]
- JKnight, A.; Onay, U.V.; Wells, S.; Li, H.; Shi, E.J.; Andrulis, I.L.; Ozcelik, H. Genetic variants of GPX1 and SOD2 and breast cancer risk at the Ontario site of the Breast Cancer Family Registry. Cancer Epidemiol. Biomark. Prev. 2004, 13, 146–149. [Google Scholar] [CrossRef]
- Lei, C.; Niu, X.; Wei, J.; Zhu, J.; Zhu, Y. Interaction of glutathione peroxidase-1 and selenium in endemic dilated cardiomyopathy. Clin. Chim. Acta 2009, 399, 102–108. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, L.; Sun, D.; Li, Z.; Wang, L.; Liu, P. Genetic polymorphisms of superoxide dismutases, catalase, and glutathione peroxidase in age-related cataract. Mol. Vis. 2011, 17, 2325–2332. [Google Scholar] [PubMed]
- Hansen, R.; Saebø, M.; Skjelbred, C.F.; Nexø, B.A.; Hagen, P.C.; Bock, G.; ILothe, M.B.; Johnson, E.; Aase, S.; Hansteen, I.L.; et al. GPX Pro198Leu and OGG1 Ser326Cys polymorphisms and risk of development of colorectal adenomas and colorectal cancer. Cancer Lett. 2005, 229, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Yeh, H.L.; Kuo, L.T.; Sung, F.C.; Yeh, C.C. Association between Polymorphisms of Antioxidant Gene (MnSOD, CAT, and GPx1) and Risk of Coronary Artery Disease. Biomed. Res. Int. 2018, 26, 5086869. [Google Scholar] [CrossRef] [PubMed]
- Wickremasinghe, D.; Peiris, H.; Chandrasena, L.G.; Senaratne, V.; Perera, R. Case control feasibility study assessing the association between severity of coronary artery disease with Glutathione Peroxidase-1 (GPX-1) and GPX-1 polymorphism (Pro198Leu). BMC Cardiovasc. Disord. 2016, 16, 111. [Google Scholar] [CrossRef]
- Souiden, Y.; Mallouli, H.; Meskhi, S.; Chaabouni, Y.; Rebai, A.; Chéour, F.; Mahdouani, K. MnSOD and GPx1 polymorphism relationship with coronary heart disease risk and severity. Biol. Res. 2016, 49, 22. [Google Scholar] [CrossRef]
- Buraczynska, M.; Buraczynska, K.; Dragan, M.; Ksiazek, A. Pro198Leu Polymorphism in the Glutathione Peroxidase 1 Gene Contributes to Diabetic Peripheral Neuropathy in Type 2 Diabetes Patients. Neuromolecular Med. 2017, 19, 147–153. [Google Scholar] [CrossRef]
- Kasznicki, J.; Sliwinska, A.; Kosmalski, M.; Merecz, A.; Majsterek, I.; Drzewoski, J. Genetic polymorphisms (Pro197Leu of Gpx1, +35A/C of SOD1, -262C/T of CAT), the level of antioxidant proteins (GPx1, SOD1, CAT) and the risk of distal symmetric polyneuropathy in Polish patients with type 2 diabetes mellitus. Adv. Med. Sci. 2016, 61, 123–129. [Google Scholar] [CrossRef]
- Mushtaq, S.; Yousaf, M.J.; Rashid, A.; Khan, S.A.; Raza, I.; Majeed, A. Association of Pro198Leu polymorphism in glutathione peroxidase one (GPX1) gene with peripheral neuropathy in type 2 diabetic patients. J. Pak. Med. Assoc. 2020, 70, 1182–1186. [Google Scholar] [CrossRef]
- Janowska, M.; Potocka, N.; Paszek, S.; Skrzypa, M.; Żulewicz, K.; Kluz, M.; Januszek, S.; Baszuk, P.; Gronwald, J.; Lubiński, J.; et al. An Assessment of GPX1 (rs1050450), DIO2 (rs225014) and SEPP1 (rs7579) Gene Polymorphisms in Women with Endometrial Cancer. Genes 2022, 13, 188. [Google Scholar] [CrossRef]
- Akbas, A.; Inanir, A.; Benli, I.; Onder, Y.; Aydogan, L. Evaluation of some antioxidant enzyme activities (SOD and GPX) and their polymorphisms (MnSOD2 Ala9Val, GPX1 Pro198Leu) in fibromyalgia. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 1199–1203. [Google Scholar]
- Malinowska, K.; Kowalski, M.; Szaflik, J.; Szaflik, J.P.; Majsterek, I. The role of Cat -262C/T, GPX1 Pro198Leu and Sod1+35A/C gene polymorphisms in a development of primary open-angle glaucoma in a Polish population. Pol. J. Pathol. 2016, 67, 404–410. [Google Scholar] [CrossRef]
- Xiong, Y.M.; Mo, X.Y.; Zou, X.Z.; Song, R.X.; Sun, W.Y.; Lu, W.; Chen, Q.; Yu, Y.X.; Zang, W.J. Association study between polymorphisms in selenoprotein genes and susceptibility to Kashin-Beck disease. Osteoarthr. Cartil. 2010, 18, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Yari, D.; Saravani, R.; Saravani, S.; Ebrahimian, K.; Galavi, H.R. Genetic Polymorphisms of Catalase and Glutathione Peroxidase-1 in Keratoconus. Iran. J. Public. Health 2018, 47, 1567–1574. [Google Scholar]
- Ilhan, A.; Altun, S.; Durukan, I.; Yolcu, U.; Erdem, U. The association between genetic polymorphism of glutathione peroxidase 1 (rs1050450) and keratoconus in a Turkish population. Arq. Bras. Oftalmol. 2019, 82, 501–506. [Google Scholar] [CrossRef]
- Soerensen, M.; Christensen, K.; Stevnsner, T.; Christiansen, L. The Mn-superoxide dismutase single nucleotide polymorphism rs4880 and the glutathione peroxidase 1 single nucleotide polymorphism rs1050450 are associated with aging and longevity in the oldest old. Mech. Ageing Dev. 2009, 130, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.H.; Lee, K.Y.; Choe, K.H.; Hong, Y.C.; Noh, S.I.; Eom, S.Y.; Ko, Y.J.; Zhang, Y.W.; Yim, D.H.; Kang, J.W.; et al. Effects of oxidative DNA damage and genetic polymorphism of the glutathione peroxidase 1 (GPX1) and 8-oxoguanine glycosylase 1 (hOGG1) on lung cancer. J. Prev. Med. Public. Health 2006, 39, 130–134. [Google Scholar]
- Ratnasinghe, D.; Tangrea, J.A.; Andersen, M.R.; Barrett, M.J.; Virtamo, J.; Taylor, P.R.; Albanes, D. Glutathione peroxidase codon 198 polymorphism variant increases lung cancer risk. Cancer Res. 2000, 60, 6381–6383. [Google Scholar] [PubMed]
- Aghakhani, R.; Nasiri, M.; Irani, D. Glutathione Peroxidase 1 gene polymorphism in nephrolithiasis patients from south of Iran. Iran. J. Kidney Dis. 2017, 11, 29–35. [Google Scholar]
- Ściskalska, M.; Milnerowicz, H. Association of genetic variants in the GPX1 and GPX4 genes with the activities of glutathione-dependent enzymes, their interaction with smoking and the risk of acute pancreatitis. Biomed. Pharmacother. 2022, 146, 112591. [Google Scholar] [CrossRef]
- Cengiz, M.; Bayoglu, B.; Alansal, N.O.; Cengiz, S.; Dirican, A.; Kocabasoglu, N. Pro198Leu polymorphism in the oxidative stress gene, glutathione peroxidase-1, is associated with a gender-specific risk for panic disorder. Int. J. Psychiatry Clin. Pract. 2015, 19, 201–207. [Google Scholar] [CrossRef]
- Lubiński, J.; Jaworowska, E.; Derkacz, R.; Marciniak, W.; Białkowska, K.; Baszuk, P.; Scott, R.J.; Lubiński, J.A. Survival of Laryngeal Cancer Patients Depending on Zinc Serum Level and Oxidative Stress Genotypes. Biomolecules 2021, 11, 865. [Google Scholar] [CrossRef]
- Steinbrecher, A.; Méplan, C.; Hesketh, J.; Schomburg, L.; Endermann, T.; Jansen, E.; Akesson, B.; Rohrmann, S.; Linseisen, J. Effects of selenium status and polymorphisms in selenoprotein genes on prostate cancer risk in a prospective study of European men. Cancer Epidemiol. Biomark. Prev. 2010, 19, 2958–2968. [Google Scholar] [CrossRef]
- Kucukgergin, C.; Gokpinar, M.; Sanli, O.; Tefik, T.; Oktar, T.; Seckin, S. Association between genetic variants in glutathione peroxidase 1 (GPx1) gene, GPx activity and the risk of prostate cancer. Minerva Urol. Nefrol. 2011, 63, 183–190. [Google Scholar]
- Erdem, O.; Eken, A.; Akay, C.; Arsova-Sarafinovska, Z.; Matevska, N.; Suturkova, L.; Erten, K.; Özgök, Y.; Dimovski, A.; Sayal, A.; et al. Association of GPX1 polymorphism, GPX activity and prostate cancer risk. Hum. Exp. Toxicol. 2012, 31, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Blein, S.; Berndt, S.; Joshi, A.D.; Campa, D.; Ziegler, R.G.; Riboli, E.; Cox, D.G. NCI Breast and Prostate Cancer Cohort Consortium. Factors associated with oxidative stress and cancer risk in the Breast and Prostate Cancer Cohort Consortium. Free. Radic. Res. 2014, 48, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Arsova-Sarafinovska, Z.; Matevska, N.; Eken, A.; Petrovski, D.; Banev, S.; Dzikova, S.; Georgiev, V.; Sikole, A.; Erdem, O.; Sayal, A.; et al. Dimovski Glutathione peroxidase 1 (GPX1) genetic polymorphism, erythrocyte GPX activity, and prostate cancer risk. Int. Urol. Nephrol. 2009, 41, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Teimoori, B.; Moradi-Shahrebabak, M.; Razavi, M.; Rezaei, M.; Harati-Sadegh, M.; Salimi, S. The effect of GPx-1 rs1050450 and MnSOD rs4880 polymorphisms on PE susceptibility: A case- control study. Mol. Biol. Rep. 2019, 46, 6099–6104. [Google Scholar] [CrossRef]
- Shuvalova, Y.A.; Kaminnyi, A.I.; Meshkov, A.N.; Shirokov, R.O.; Samko, A.N. Association between polymorphisms of eNOS and GPx-1 genes, activity of free-radical processes and in-stent restenosis. Mol. Cell. Biochem. 2012, 370, 241–249. [Google Scholar] [CrossRef]
- Huang, Y.S.; Chang, T.E.; Perng, C.L.; Huang, Y.H. Genetic variations of three important antioxidative enzymes SOD2, CAT, and GPX1 in nonalcoholic steatohepatitis. J. Chin. Med. Assoc. 2021, 84, 14–18. [Google Scholar] [CrossRef]
- Salimi, S.; Harati-Sadegh, M.; Eskandari, M.; Heidari, Z. The effects of the genetic polymorphisms of antioxidant enzymes on susceptibility to papillary thyroid carcinoma. IUBMB Life 2020, 72, 1045–1053. [Google Scholar] [CrossRef]
- Shao, X.; Yan, C.; Sun, D.; Fu, C.; Tian, C.; Duan, L.; Zhu, G. Association Between Glutathione Peroxidase-1 (GPx-1) Polymorphisms and Schizophrenia in the Chinese Han Population. Neuropsychiatr. Dis. Treat. 2020, 16, 2297–2305. [Google Scholar] [CrossRef]
- Bhatti, P.; Stewart, P.A.; Hutchinson, A.; Rothman, N.; Linet, M.S.; Inskip, P.D.; Rajaraman, P. Lead exposure, polymorphisms in genes related to oxidative stress, and risk of adult brain tumors. Cancer Epidemiol. Biomark. Prev. 2009, 18, 1841–1848. [Google Scholar] [CrossRef] [PubMed]
- Morgan, A.R.; Han, D.Y.; Lam, W.J.; Fraser, A.G.; Ferguson, L.R. Association analysis of 3p21 with Crohn’s disease in a New Zealand population. Hum. Immunol. 2010, 71, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Song, S.; Zhang, J.; Li, X.; Gao, H. Effects of GSTA1 and GPX3 Polymorphisms on the Risk of Schizophrenia in Chinese Han Population. Neuropsychiatr. Dis. Treat. 2020, 16, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Baez-Duarte, B.G.; Mendoza-Carrera, F.; García-Zapién, A.; Flores-Martínez, S.E.; Sánchez-Corona, J.; Zamora-Ginez, I.; Torres-Rasgado, E.; León-Chávez, B.A.; Pérez-Fuentes, R. Multidisciplinary Research Group on Diabetes of the Instituto Mexicano del Seguro Social, Glutathione peroxidase 3 serum levels and GPX3 gene polymorphisms in subjects with metabolic syndrome. Arch. Med. Res. 2014, 45, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Chien, C.-Y.; Huang, T.-Y.; Tai, S.-Y.; Chang, N.-C.; Wang, H.-M.; Wang, L.-F.; Ho, K.-Y. Glutathione peroxidase 3 gene polymorphisms and the risk of sudden sensorineural hearing loss. Kaohsiung J. Med. Sci. 2017, 33, 359–364. [Google Scholar] [CrossRef] [PubMed]
- Bumbasirevic, U.; Bojanic, N.; Pljesa-Ercegovac, M.; Zivkovic, M.; Djukic, T.; Zekovic, M.; Milojevic, B.; Kajmakovic, B.; Janicic, A.; Simic, T.; et al. The Polymorphisms of Genes Encoding Catalytic Antioxidant Proteins Modulate the Susceptibility and Progression of Testicular Germ Cell Tumor. Cancers 2022, 14, 1068. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Lin, Y.; Li, J.; Liu, M.; Wang, J.; Li, X.; Liu, J.; Jia, X.; Jing, Z.; Huang, Z.; et al. Evaluation of Glutathione Peroxidase 4 role in Preeclampsia. Sci. Rep. 2016, 6, 33300. [Google Scholar] [CrossRef]
- Silander, K.; Alanne, M.; Kristiansson, K.; Saarela, O.; Ripatti, S.; Auro, K.; Karvanen, J.; Kulathinal, S.; Niemelä, M.; Ellonen, P.; et al. Gender differences in genetic risk profiles for cardiovascular disease. PLoS ONE 2008, 3, e3615. [Google Scholar] [CrossRef]
- Crosley, L.K.; Bashir, S.; Nicol, F.; Arthur, J.R.; Hesketh, J.E.; Sneddon, A.A. The single-nucleotide polymorphism (GPX4c718t) in the glutathione peroxidase 4 gene influences endothelial cell function: Interaction with selenium and fatty acids. Mol. Nutr. Food Res. 2013, 57, 2185–2194. [Google Scholar] [CrossRef]
- Polonikov, A.V.; Vialykh, E.K.; Churnosov, M.I.; Illig, T.; Freidin, M.B.; Vasil’eva, O.V.; Bushueva, O.Y.; Ryzhaeva, V.N.; Bulgakova, I.V.; Solodilova, M.A. The C718T polymorphism in the 3’-untranslated region of glutathione peroxidase-4 gene is a predictor of cerebral stroke in patients with essential hypertension. Hypertens. Res. 2012, 35, 507–512. [Google Scholar] [CrossRef]
- Huang, Y.Y.; Wu, C.H.; Liu, C.H.; Yang, S.F.; Wang, P.H.; Lin, L.Y.; Lee, T.H.; Lee, M.S. Association between the Genetic Variants of Glutathione Peroxidase 4 and Severity of Endometriosis. Int. J. Environ. Res. Public Health 2020, 17, 5089. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Zhao, H.; Wang, J.; Zhang, R.; Liu, J.; Zhao, X.; Li, C.; Jia, X.; Li, X.; Lin, Y.; et al. Haplotype Analysis of Candidate Genes Involved in Inflammation and Oxidative Stress and the Susceptibility to Preeclampsia. J. Immunol. Res. 2020, 26, 4683798. [Google Scholar] [CrossRef] [PubMed]
- Admoni, S.N.; Santos-Bezerra, D.P.; Perez, R.V.; Patente, T.A.; Monteiro, M.B.; Cavaleiro, A.M.; Parisi, M.C.; Moura Neto, A.; Pavin, E.J.; Queiroz, M.S.; et al. Glutathione peroxidase 4 functional variant rs713041 modulates the risk for cardiovascular autonomic neuropathy in individuals with type 1 diabetes. Diab Vasc. Dis. Res. 2019, 16, 297–299. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Yuan, J.; Yao, Q.; Yan, N.; Song, R.; Jiang, W.; Li, D.; Shi, L.; Zhang, J.A. A case-control study of selenoprotein genes polymorphisms and autoimmune thyroid diseases in a Chinese population. BMC Med. Genet. 2017, 18, 54. [Google Scholar] [CrossRef]
- Philibert, R.A.; Beach, S.R.H.; Gunter, T.D.; Todorov, A.A.; Brody, G.H.; Vijayendran, M.; Cutrona, C. The relationship of deiodinase 1 genotype and thyroid function to lifetime history of major depression in three independent populations. Am. J. Med. Genet. Part. B Neuropsychiatr. Genet. 2011, 156, 593–599. [Google Scholar] [CrossRef] [PubMed]
- Comarella, A.P.; Vilagellin, D.; Bufalo, N.E.; Euflauzino, J.F.; Teixeira, E.S.; Miklos, A.B.P.P.; Santos, R.B.; Romaldini, J.H.; Ward, L.S. The polymorphic inheritance of DIO2 rs225014 may predict body weight variation after Graves’ disease treatment. Arch. Endocrinol. Metab. 2021, 64, 787–795. [Google Scholar] [CrossRef]
- Taroza, S.; Rastenytė, D.; Podlipskytė, A.; Patamsytė, V.; Mickuvienė, N. Deiodinases, organic anion transporter polypeptide polymorphisms and ischemic stroke outcomes. J. Neurol. Sci. 2019, 29, 116457. [Google Scholar] [CrossRef]
- Beltrão, F.E.; Beltrão, D.C.A.; Carvalhal, G.; Beltrão, F.E.L.; Braga Filho, J.S.; Oliveira, J.B.; Jesus, J.D.S.; Machado, G.J.R.; Santos Silva, H.; Teixeira, H.M.P.; et al. Heterozygote Advantage of the Type II Deiodinase Thr92Ala Polymorphism on Intrahospital Mortality of COVID-19. J. Clin. Endocrinol. Metab. 2022, 107, e2488–e2501. [Google Scholar] [CrossRef]
- Benenati, N.; Bufano, A.; Cantara, S.; Ricci, C.; Marzocchi, C.; Ciuoli, C.; Sannino, I.; Tirone, A.; Voglino, C.; Vuolo, G.; et al. Type 2 deiodinase p.Thr92Ala polymorphism does not affect the severity of obesity and weight loss after bariatric surgery. Sci. Rep. 2022, 12, 10643. [Google Scholar] [CrossRef]
- Schwengber, W.K.; Silveira, V.B.; Hetzel, G.M.; Robaina, A.; Ceolin, L.; Camelier, M.T.; Goemann, I.; Dalla Corte, R.R.; Scheffel, R.S.; RMello, G.B.; et al. Type 2 Deiodinase Thr92Ala polymorphism is not associated with cognitive impairment in older adults: A cross-sectional study. Metabolites 2022, 12, 375. [Google Scholar] [CrossRef]
- Peters, U.; Chatterjee, N.; Hayes, R.B.; Schoen, R.E.; Wang, Y.; Chanock, S.J.; Foster, C.B. Variation in the selenoenzyme genes and risk of advanced distal colorectal adenoma. Cancer Epidemiol. Biomark. Prev. 2008, 17, 1144–1154. [Google Scholar] [CrossRef]
- Wu, R.; Zhang, R.; Xiong, Y.; Sun, W.; Li, Y.; Yang, X.; Liu, J.; Jiang, Y.; Guo, H.; Mo, X.; et al. The study on polymorphisms of Sep15 and TrxR2 and the expression of AP-1 signaling pathway in Kashin-Beck disease. Bone 2019, 120, 239–245. [Google Scholar] [CrossRef]
- Kariž, S.; Mankoč, S.; Petrovič, D. Association of thioredoxin reductase 2 (TXNRD2) gene polymorphisms with myocardial infarction in Slovene patients with type 2 diabetes mellitus. Diabetes Res. Clin. Pract. 2015, 108, 323–328. [Google Scholar] [CrossRef]
- Lu, W.; Mo, X.Y.; Xiong, Y.M. TrxR2 gene polymorphisms may not be associated with the susceptibility to Kashin-Beck disease. Nan Fang. Yi Ke Da Xue Xue Bao 2010, 30, 2246–2468. [Google Scholar]
- Lu, W.; Mo, X.-Y.; Xiong, Y.-M. The Study on Polymorphism of TrxR and Nrf2/HO-1 Signaling Pathway in Kaschin-Beck Disease. Biol. Trace Elem. Res. 2019, 190, 303–308. [Google Scholar]
- Sutherland, A.; Kim, D.H.; Relton, C.; Ahn, Y.O.; Hesketh, J. Polymorphisms in the selenoprotein S and 15-kDa selenoprotein genes are associated with altered susceptibility to colorectal cancer. Genes. Nutr. 2010, 5, 215–223. [Google Scholar] [CrossRef]
- Benelli, J.L.; Medeiros, R.M.; Matte, M.C.; Melo, M.G.; Almeida, S.E.M.; Fiegenbaum, M. Role of SEP15 gene polymorphisms in the time of progression to AIDS. Genet. Test. Mol. Biomark. 2016, 20, 383–387. [Google Scholar] [CrossRef]
- Shibata, T.; Arisawa, T.; Tahara, T.; Ohkubo, M.; Yoshioka, D.; Maruyama, N.; Fujita, H.; Kamiya, Y.; Nakamura, M.; Nagasaka, M.; et al. Selenoprotein S (SEPS1) gene -105G>A promoter polymorphism influences the susceptibility to gastric cancer in the Japanese population. BMC Gastroenterol. 2009, 9, 2. [Google Scholar] [CrossRef]
- Wang, Y.; Yang, X.; Zheng, Y.; Wu, Z.H.; Zhang, X.A.; Li, Q.P.; He, X.Y.; Wang, C.Z.; Feng, Z.C. The SEPS1 G-105A polymorphism is associated with risk of spontaneous preterm birth in a Chinese population. PLoS ONE 2013, 8, e65657. [Google Scholar] [CrossRef]
- Santos, L.R.; Durães, C.; Mendes, A.; Prazeres, H.; Alvelos, M.I.; Moreira, C.S.; Canedo, P.; Esteves, C.; Neves, C.; Carvalho, D.; et al. A polymorphism in the promoter region of the selenoprotein S gene (SEPS1) contributes to Hashimoto’s thyroiditis susceptibility. J. Clin. Endocrinol. Metab. 2014, 99, E719–E723. [Google Scholar] [CrossRef]
- Martínez, A.; Santiago, J.L.; Varadé, J.; Márquez, A.; Lamas, J.R.; Mendoza, J.L.; de la Calle, H.; Díaz-Rubio, M.; de la Concha, E.G.; Fernández-Gutiérrez, B.; et al. Polymorphisms in the selenoprotein S gene: Lack of association with autoimmune inflammatory diseases. BMC Genom. 2008, 9, 329. [Google Scholar] [CrossRef]
- Zhou, L.; Luo, C.; Yin, J.; Zhu, Y.; Li, P.; Chen, S.; Sun, T.; Xie, M.; Shan, Z.; Cao, B.; et al. Diverse Associations of Plasma Selenium Concentrations and SELENOP Gene Polymorphism with Metabolic Syndrome and Its Components. Oxid. Med. Cell. Longev. 2020, 22, 5343014. [Google Scholar] [CrossRef]
- Mohammaddoust, S.; Salehi, Z.; Saeidi Saedi, H. SEPP1 and SEP15 gene polymorphisms and susceptibility to breast cancer. Br. J. Biomed. Sci. 2018, 75, 36–39. [Google Scholar] [CrossRef]
- Penney, K.L.; Li, H.; Mucci, L.A.; Loda, M.; Sesso, H.D.; Stampfer, M.J.; Ma, J. Selenoprotein P genetic variants and mrna expression, circulating selenium, and prostate cancer risk and survival. Prostate 2013, 73, 700–705. [Google Scholar] [CrossRef]
- Xie, W.; Yang, M.; Chan, J.; Sun, T.; Mucci, L.A.; Penney, K.L.; Kantoff, P.W. Association of genetic variations of selenoprotein genes, plasma selenium levels, and prostate cancer aggressiveness at diagnosis. Prostate 2016, 76, 691–699. [Google Scholar] [CrossRef]
- Akbaba, G.; Akbaba, E.; Sahin, C.; Kara, M. The relationship between gestational diabetes mellitus and selenoprotein-P plasma 1 (SEPP1) gene polymorphisms. Gynecol. Endocrinol. 2018, 34, 849–852. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigó, R.; Gladyshev, V.N. Characterization of Mammalian Selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef]
- Esworthy, R.S.; Ho, Y.-S.; Chu, F.-F. The Gpx1Gene encodes mitochondrial glutathione peroxidase in the mouse liver. Arch. Biochem. Biophys. 1997, 340, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Antunes, F.; Han, D.; Cadenas, E. Relative contributions of heart mitochondria glutathione peroxidase and catalase to H2O2 detoxification in in vivo conditions. Free Radic. Biol. Med. 2002, 33, 1260–1267. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Cohen, H.J. Selenium-dependent glutathione peroxidase protein and activity: Immunological investigations on cellular and plasma enzymes. Blood 1986, 68, 640–645. [Google Scholar] [CrossRef]
- Whitin, J.C.; Bhamre, S.; Tham, D.M.; Cohen, H.J. Extracellular glutathione peroxidase is secreted basolaterally by human renal proximal tubule cells. Am. J. Physiol. Ren. Physiol. 2002, 283, F20–F28. [Google Scholar] [CrossRef] [PubMed]
- Cozza, G.; Rossetto, M.; Bosello-Travain, V.; Maiorino, M.; Roveri, A.; Toppo, S.; Zaccarin, M.; Zennaro, L.; Ursini, F. Glutathione peroxidase 4-catalyzed reduction of lipid hydroperoxides in membranes: The polar head of membrane phospholipids binds the enzyme and addresses the fatty acid hydroperoxide group toward the redox center. Free Radic. Biol. Med. 2017, 112, 1–11. [Google Scholar] [CrossRef]
- Miao, Y.; Chen, Y.; Xue, F.; Liu, K.; Zhu, B.; Gao, J.; Yin, J.; Zhang, C.; Li, G. Contribution of ferroptosis and GPX4’s dual functions to osteoarthritis progression. EBioMedicine 2022, 76, 103847. [Google Scholar] [CrossRef]
- Imai, H.; Suzuki, K.; Ishizaka, K.; Ichinose, S.; Oshima, H.; Okayasu, I.; Emoto, K.; Umeda, M.; Nakagawa, Y. Failure of the expression of phospholipid hydroperoxide glutathione peroxidase in the spermatozoa of human infertile males. Biol. Reprod. 2001, 64, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Schomburg, L. Selenoprotein P–Selenium transport protein, enzyme and biomarker of selenium status. Free Radic. Biol. Med. 2022, 191, 150–163. [Google Scholar] [CrossRef] [PubMed]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. The Sep15 protein family: Roles in disulfide bond formation and quality control in the endoplasmic reticulum. IUBMB Life 2007, 59, 1–5. [Google Scholar] [CrossRef]
- Ekoue, D.N.; Ansong, E.; Liu, L.; Macias, V.; Deaton, R.; Lacher, C.; Picklo, M.; Nonn, L.; Gann, P.H.; Kajdacsy-Balla, A.; et al. Correlations of SELENOF and SELENOP genotypes with serum selenium levels and prostate cancer. Prostate 2018, 78, 279–288. [Google Scholar] [CrossRef]
- Turanov, A.A.; Shchedrina, V.A.; Everley, R.A.; Lobanov, A.V.; Yim, S.H.; Marino, S.M.; Gygi, S.P.; Hatfield, D.L.; Gladyshev, V.N. Selenoprotein S is involved in maintenance and transport of multiprotein complexes. Biochem. J. 2014, 462, 555–565. [Google Scholar] [CrossRef]
- Taylor, D.; Dalton, C.; Hall, A.; Woodroofe, M.; Gardiner, P. Recent developments in selenium research. Br. J. Biomed. Sci. 2009, 66, 107–116. [Google Scholar] [CrossRef]
- Vendeland, S.C.; Beilstein, M.A.; Yeh, J.Y.; Ream, W.; Whanger, P.D. Rat skeletal muscle selenoprotein W: cDNA clone and mRNA modulation by dietary selenium. Proc. Natl. Acad. Sci. USA 1995, 92, 8749–8753. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Lee, T.-J.; Sebastian, A.; McGuigan, J.; Liao, C.; Koo, I.; Patterson, A.D.; Rossi, R.M.; Hall, M.A.; Albert, I.; et al. Loss of selenoprotein W in murine macrophages alters the hierarchy of selenoprotein expression, redox tone, and mitochondrial functions during inflammation. Redox Biol. 2023, 59, 102571. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, K.; Kim, J.M.; Kim, M.Y.; Kim, J.-R.; Lee, H.-W.; Chung, Y.W.; Shin, H.-I.; Kim, T.; Park, E.-S.; et al. Selenoprotein W ensures physiological bone remodeling by preventing hyperactivity of osteoclasts. Nat. Commun. 2021, 12, 2258. [Google Scholar] [CrossRef]
- Liu, A.; Li, F.; Xu, P.; Chen, Y.; Liang, X.; Zheng, S.; Meng, H.; Zhu, Y.; Mo, J.; Gong, C.; et al. Gpx4, Selenov, and Txnrd3 Are Three Most Testis-Abundant Selenogenes Resistant to Dietary Selenium Concentrations and Actively Expressed During Reproductive Ages in Rats. Biol. Trace Elem. Res. Jan. 2022, 201, 250–259. [Google Scholar] [CrossRef]
- Sengupta, A.; Carlson, B.A.; Labunskyy, V.M.; Gladyshev, V.N.; Hatfield, D.L. Selenoprotein T deficiency alters cell adhesion and elevates selenoprotein W expression in murine fibroblast cells. Biochem. Cell. Biol. 2009, 87, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Horibata, Y.; Hirabayashi, Y. Identification and characterization of human ethanolaminephosphotransferase1. J. Lipid Res. 2007, 48, 503–508. [Google Scholar] [CrossRef]
- Horibata, Y.; Elpeleg, O.; Eran, A.; Hirabayashi, Y.; Savitzki, D.; Tal, G.; Mandel, H.; Sugimoto, H. EPT1 (selenoprotein I) is critical for the neural development and maintenance of plasmalogen in humans. J. Lipid Res. 2018, 59, 1015–1026. [Google Scholar] [CrossRef]
- Avery, J.C.; Yamazaki, Y.; Hoffmann, F.W.; Folgelgren, B.; Hoffmann, P.R. Selenoprotein I is essential for murine embryogenesis. Arch. Biochem. Biophys. 2020, 689, 108444. [Google Scholar] [CrossRef]
- Papp, L.V.; Lu, J.; Holmgren, A.; Khanna, K.K. From selenium to selenoproteins: Synthesis, identity, and their role in human health. Antioxid. Redox Signal. 2007, 9, 775–806. [Google Scholar] [CrossRef]
- Fredericks, G.J.; Hoffmann, P.R. Selenoprotein K and protein palmitoylation. Antioxid. Redox Signal. 2015, 23, 854–862. [Google Scholar] [CrossRef]
- Reeves, M.A.; Bellinger, F.P.; Berry, M.J. The neuroprotective functions of selenoprotein M and its role in cytosolic calcium regulation. Antioxid. Redox Signal. 2010, 12, 809–818. [Google Scholar] [CrossRef]
- Dai, J.; Liu, H.; Zhou, J.; Huang, K. Selenoprotein R Protects Human Lens Epithelial Cells against D-Galactose-Induced Apoptosis by Regulating Oxidative Stress and Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2016, 17, 231. [Google Scholar] [CrossRef]
- Tarrago, L.; Kaya, A.; Kim, H.-Y.; Manta, B.; Lee, B.-C.; Gladyshev, V.N. The selenoprotein methionine sulfoxide reductase B1 (MSRB1). Free Radic. Biol. Med. 2022, 191, 228–240. [Google Scholar] [CrossRef]
- Li, H.; He, Q.; Meng, F.; Feng, X.; Chen, J.; Li, L.; Liu, J. Methionine sulfoxide reductase B1 regulates proliferation and invasion by affecting mitogen-activated protein kinase pathway and epithelial-mesenchymal transition in u2os cells. Biochem. Biophys. Res. Commun. 2018, 496, 806–813. [Google Scholar] [CrossRef]
- Han, S.J.; Lee, B.C.; Yim, S.H.; Gladyshev, V.N.; Lee, S.R. Characterization of mammalian selenoprotein O: A redox-active mitochondrial protein. PLoS ONE 2014, 9, e95518. [Google Scholar] [CrossRef] [PubMed]
- Sreelatha, A.; Yee, S.S.; Lopez, V.A.; Park, B.C.; Kinch, L.N.; Pilch, S.; Servage, K.A.; Zhang, J.; Jiou, J.; Karasiewicz-Urbańska, M.; et al. Protein AMPylation by an Evolutionarily Conserved Pseudokinase. Cell 2018, 175, 809–821.e19. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Fei, Y.; Han, Y.; Lu, S. Selenoprotein O deficiencies suppress chondrogenic differentiation of ATDC5 cells. Cell. Biol. Int. 2016, 40, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Zhang, J.; Peng, S.; Liu, R.; Li, X.; Hou, Y.; Fang, J. Thioredoxin reductase inhibitors: A patent review. Expert. Opin. Ther. Pat. 2017, 27, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Rundlof, A.K.; Arner, E.S. Regulation of the mammalian selenoprotein thioredoxin reductase 1 in relation to cellular phenotype, growth, and signaling events. Antioxid. Redox Signal. 2004, 6, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Gladyshev, V.N. Alternative Splicing Involving the Thioredoxin Reductase Module in Mammals: A Glutaredoxin-Containing Thioredoxin Reductase 1. Biochemistry 2004, 43, 12177–12188. [Google Scholar] [CrossRef] [PubMed]
- Turanov, A.A.; Su, D.; Gladyshev, V.N. Characterization of alternative cytosolic forms and cellular targets of mouse mitochondrial thioredoxin reductase. J. Biol. Chem. 2006, 281, 22953–22963. [Google Scholar] [CrossRef] [PubMed]
- Rigobello, M.P.; Callegaro, M.T.; Barzon, E.; Benetti, M.; Bindoli, A. Purification of mitochondrial thioredoxin reductase and its involvement in the redox regulation of membrane permeability. Free Radic. Biol. Med. 1998, 24, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Jakupoglu, C.; Przemeck, G.K.; Schneider, M.; Moreno, S.G.; Mayr, N.; Hatzopoulos, A.K.; de Angelis, M.H.; Wurst, W.; Bornkamm, G.W.; Brielmeier, M.; et al. Cytoplasmic thioredoxin reductase is essential for embryogenesis but dispensable for cardiac development. Mol. Cell. Biol. 2005, 25, 1980–1988. [Google Scholar] [CrossRef] [PubMed]
- Nonn, L.; Williams, R.R.; Erickson, R.P.; Powis, G. The absence of mitochondrial thioredoxin 2 causes massive apoptosis, exencephaly, and early embryonic lethality in homozygous mice. Mol. Cell. Biol. 2003, 23, 916–922. [Google Scholar] [CrossRef]
- Stancill, J.S.; Broniowska, K.A.; Oleson, B.J.; Naatz, A.; Corbett, J.A. Pancreatic β-cells detoxify H2O2 through the peroxiredoxin/thioredoxin antioxidant system. J. Biol. Chem. 2019, 294, 4843–4853. [Google Scholar] [CrossRef]
- Lopert, P.; Day, B.J.; Patel, M. Thioredoxin Reductase Deficiency Potentiates Oxidative Stress, Mitochondrial Dysfunction and Cell Death in Dopaminergic Cells. PLoS ONE 2012, 7, e50683. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Xu, J.; Fei, Y.; Jiang, C.; Zhu, W.; Han, Y.; Lu, S. TrxR2 deficiencies promote chondrogenic differentiation and induce apoptosis of chondrocytes through mitochondrial reactive oxygen species. Exp. Cell. Res. 2016, 344, 67–75. [Google Scholar] [CrossRef]
- Wang, H.; Dou, Q.; Jeong, K.J.; Choi, J.; Gladyshev, V.N.; Chung, J.-J. Redox regulation by TXNRD3 during epididymal maturation underlies capacitation-associated mitochondrial activity and sperm motility in mice. J. Biol. Chem. 2022, 298, 102077. [Google Scholar] [CrossRef]
- Jakobs, T.C.; Koehler, M.R.; Schmutzler, C.; Glaser, F.; Schmid, M.; Köhrle, J. Structure of the human type I iodothyronine 5’-deiodinase gene and localization to chromosome 1p32–p33. Genomics 1997, 42, 361–363. [Google Scholar] [CrossRef]
- Baqui, M.; Botero, D.; Gereben, B.; Curcio, C.; Harney, J.W.; Salvatore, D.; Sorimachi, K.; Larsen, P.R.; Bianco, A.C. Human type 3 iodothyronine selenodeiodinase is located in the plasma membrane and undergoes rapid internalization to endosomes. J. Biol. Chem. 2003, 278, 1206–1211. [Google Scholar] [CrossRef]
- Labunskyy, V.M.; Hatfield, D.L.; Gladyshev, V.N. Selenoproteins: Molecular pathways and physiological roles. Physiol. Rev. 2014, 94, 739–777. [Google Scholar] [CrossRef] [PubMed]
- Baqui, M.M.A.; Gereben, B.; Harney, J.W.; Larsen, P.R.; Bianco, A.C. Distinct Subcellular Localization of Transiently Expressed Types 1 and 2 Iodothyronine Deiodinases as Determined by Immunofluorescence Confocal Microscopy. Endocrinology 2000, 141, 4309–4312. [Google Scholar] [CrossRef]
- Peeters, R.P.; Visser, T.J. Metabolism of Thyroid Hormone. In Endotext [Internet]; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2017. [Google Scholar]
- Paragliola, R.M.; Corsello, A.; Concolino, P.; Ianni, F.; Papi, G.; Pontecorvi, A.; Corsello, S.M. Iodothyronine deiodinases and reduced sensitivity to thyroid hormones. Front. Biosci. 2020, 25, 201–228. [Google Scholar]
- Forsberg, L.; de Faire, U.; Marklund, S.L.; Andersson, P.M.; Stegmayr, B.; Morgenstern, R. Phenotype determination of a common Pro-Leu polymorphism in human Glutathione Peroxidase 1. Blood Cells Mol. Dis. 2000, 26, 423–426. [Google Scholar] [CrossRef] [PubMed]
- Méplan, C.; Dragsted, L.O.; Ravn-Haren, G.; Tjønneland, A.; Vogel, U.; Hesketh, J. Association between Polymorphisms in Glutathione Peroxidase and Selenoprotein P Genes, Glutathione Peroxidase Activity, HRT Use and Breast Cancer Risk. PLoS ONE 2013, 8, e73316. [Google Scholar] [CrossRef] [PubMed]
- Abdul-Maksoud, R.S.; Fouad, R.A.; Elsayed, T.G.; Ibrahem, R.A.; Badawi, A.E. The impact of catalase and glutathione peroxidase-1 genetic polymorphisms on their enzyme activities among Egyptian patients with keratoconus. J. Gene Med. 2020, 22, e3192. [Google Scholar] [CrossRef] [PubMed]
- Yalin, G.Y.; Akgul, S.; Tanrikulu, S.; Purisa, S.; Gul, N.; Uzum, A.K.; Sarvan, F.O.; Sever, M.S.; Satman, I. Evaluation of Glutathione Peroxidase and KCNJ11 Gene Polymorphisms in Patients with New Onset Diabetes Mellitus After Renal Transplantation. Exp. Clin. Endocrinol. Diabetes 2017, 125, 408–413. [Google Scholar] [CrossRef]
- Hamanishi, T.; Furuta, H.; Kato, H.; Doi, A.; Tamai, M.; Shimomura, H.; Sakagashira, S.; Nishi, M.; Sasaki, H.; Sanke, T.; et al. Functional variants in the glutathione peroxidase-1 (GPx-1) gene are associated with increased intima-media thickness of carotid arteries and risk of macrovascular diseases in Japanese type 2 diabetic patients. Diabetes 2004, 53, 2455–2460. [Google Scholar] [CrossRef]
- Hernández Guerrero, C.; Hernández Chávez, P.; Martínez Castro, N.; Parra Carriedo, A.; García Del Rio, S.; Pérez Lizaur, A. Glutathione peroxidase-1 pro200leu polymorphism (rs1050450) is associated with morbid obesity independently of the presence of prediabetes or diabetes in women from central Mexico. Nutr. Hosp. 2015, 32, 1516–1525. [Google Scholar]
- Jerotic, D.; Ranin, J.; Bukumiric, Z.; Djukic, T.; Coric, V.; Savic-Radojevic, A.; Todorovic, N.; Asanin, M.; Ercegovac, M.; Milosevic, I.; et al. SOD2 rs4880 and GPX1 rs1050450 polymorphisms do not confer risk of COVID-19, but influence inflammation or coagulation parameters in Serbian cohort. Redox Rep. 2022, 27, 85–91. [Google Scholar] [CrossRef]
- Mazjin, M.A.; Salehi, Z.; Mashayekhi, F.; Bahadori, M. Evaluation of GPx1 Pro198Leu Polymorphism in Idiopathic Male Infertility. Mol. Biol. 2016, 50, 89–93. [Google Scholar] [CrossRef]
- Moscow, J.A.; Schmidt, L.; Ingram, D.T.; Gnarra, J.; Johnson, B.; Cowan, K.H. Loss of heterozygosity of the human cytosolic glutathione peroxidase I gene in lung cancer. Carcinogenesis 1994, 15, 2769–2773. [Google Scholar] [CrossRef]
- Zhuo, P.; Goldberg, M.; Herman, L.; Lee, B.S.; Wang, H.; Brown, R.L.; Foster, C.B.; Peters, U.; Diamond, A.M. Molecular consequences of genetic variations in the glutathione peroxidase 1 selenoenzyme. Cancer Res. 2009, 69, 8183–8190. [Google Scholar] [CrossRef]
- Najafi, M.; Ghasemi, H.; Roustazadeh, A.; Alipoor, B. Phenotype and genotype relationship of glutathione peroxidase1 (GPx1) and rs 1800668 variant: The homozygote effect on kinetic parameters. Gene 2012, 505, 19–22. [Google Scholar] [CrossRef] [PubMed]
- Méplan, C.; Crosley, L.K.; Nicol, F.; Horgan, G.W.; Mathers, J.C.; Arthur, J.R.; Hesketh, J.E. Functional effects of a common single-nucleotide polymorphism (GPX4c718t) in the glutathione peroxidase 4 gene: Interaction with sex. Am. J. Clin. Nutr. 2008, 87, 1019–1027. [Google Scholar] [CrossRef]
- Gautrey, H.; Nicol, F.; Sneddon, A.A.; Hall, J.; Hesketh, J. AT/C polymorphism in the GPX4 3′UTR affects the selenoprotein expression pattern and cell viability in transfected Caco-2 cells. Biochim. Biophys. Acta 2011, 1810, 584–591. [Google Scholar] [CrossRef]
- Méplan, C.; Crosley, L.K.; Nicol, F.; Beckett, G.J.; Howie, A.F.; Hill, K.E.; Horgan, G.; Mathers, J.C.; Arthur, J.R.; Hesketh, J.E. Genetic polymorphisms in the human selenoprotein P gene determine the response of selenoprotein markers to selenium supplementation in a gender-specific manner (the SELGEN study). FASEB J. 2007, 21, 3063–3074. [Google Scholar] [CrossRef]
- Maiorino, M.; Bosello, V.; Ursini, F.; Foresta, C.; Garolla, A.; Scapin, M.; Sztajer, H.; Flohe, L. Genetic variations of gpx-4 and male infertility in humans. Biol. Reprod. 2003, 68, 1134–1141. [Google Scholar] [CrossRef]
- Kumaraswamy, E.; Malykh, A.; Korotkov, K.V.; Kozyavkin, S.; Hu, Y.; Kwon, S.Y.; Gladyshev, V.N. Structure-Expression Relationships of the 15-kDa Selenoprotein Gene. J. Biol. Chem. 2000, 275, 35540–35547. [Google Scholar] [CrossRef]
- Hu, Y.J.; Korotkov, K.V.; Mehta, R.; Hatfield, D.L.; Rotimi, C.N.; Luke, A.; Prewitt, T.E.; Cooper, R.S.; Stock, W.; Vokes, E.E.; et al. Distribution and functional consequences of nucleotide polymorphisms in the 3′-untranslated region of the human Sep15 gene. Cancer Res. 2001, 61, 2307–2310. [Google Scholar]
- Watrowski, R.; Castillo-Tong, D.C.; Fabjani, G.; Schuster, E.; Fischer, M.; Zeillinger, R. The 811 C/T polymorphism in the 3′ untranslated region of the selenoprotein 15-kDa (Sep15) gene and breast cancer in Caucasian women. Tumour Biol. 2016, 37, 1009–1015. [Google Scholar] [CrossRef]
- Jablonska, E.; Gromadzinska, J.; Sobala, W.; Reszka, E.; Wasowicz, W. Lung cancer risk associated with selenium status is modified in smoking individuals by Sep15 polymorphism. Eur. J. Nutr. 2008, 47, 47–54. [Google Scholar] [CrossRef]
- Curran, J.E.; Jowett, J.B.; Elliott, K.S.; Gao, Y.; Gluschenko, K.; Wang, J.; Abel Azim, D.M.; Cai, G.; Mahaney, M.C.; Comuzzie, A.G.; et al. Genetic variation in selenoprotein S influences inflammatory response. Nat. Genet. 2005, 37, 1234–1241. [Google Scholar] [CrossRef]
- Fedorova, L.; Fedorov, A. Introns in gene evolution. Genetica 2003, 118, 123–131. [Google Scholar] [CrossRef]
- Sarzo, B.; Abumallouh, R.; Marín, N.; Llop, S.; Beneito, A.; Lopez-Flores, I.; Ferrero, N.; Sakhi, A.K.; Ballester, F.; Lopez-Espinosa, M.J. Association between phenols and thyroid hormones: The role of iodothyronine deiodinase genes. Environ. Pollut. 2022, 311, 119926. [Google Scholar] [CrossRef]
- Peeters, R.P.; van den Beld, A.W.; Attalki, H.; Toor, H.; de Rijke, Y.B.; Kuiper, G.G.; Lamberts, S.W.; Janssen, J.A.; Uitterlinden, A.G.; Visser, T.J. A new polymorphism in the type II deiodinase gene is associated with circulating thyroid hormone parameters. Am. J. Physiol. Endocrinol. Metab. 2005, 289, E75–E81. [Google Scholar] [CrossRef]
- Peltsverger, M.Y.; Butler, P.W.; Alberobello, A.T.; Smith, S.; Guevara, Y.; Dubaz, O.M.; Luzon, J.A.; Linderman, J.; Celi, F.S. The−258A/G (SNP rs12885300) polymorphism of the human type 2 deiodinase gene is associated with a shift in the pattern of secretion of thyroid hormones following a TRH-induced acute rise in TSH. Eur. J. Endocrinol. 2012, 166, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.E.; Kang, Y.M.; Park, B.; Shong, M.; Yi, H.S. Type 2 deiodinase Thr92Ala polymorphism is associated with a reduction in bone mineral density: A community-based Korean genome and epidemiology study. Clin. Endocrinol. 2020, 93, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Copeland, P.R.; Fletcher, J.E.; Carlson, B.A.; Hatfield, D.L.; Driscoll, D.M. A novel RNA binding protein, SBP2, is required for the translation of mammalian selenoprotein mRNAs. EMBO J. 2000, 19, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Tobe, R.; Mihara, H. Delivery of selenium to selenophosphate synthetase for selenoprotein biosynthesis. Biochim. Et Biophys. Acta (BBA)-General. Subj. 2018, 1862, 2433–2440. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.M.; Carlson, B.A.; Irons, R.; Mix, H.; Zhong, N.; Gladyshev, V.N.; Hatfield, D.L. Selenophosphate synthetase 2 is essential for selenoprotein biosynthesis. Biochem. J. 2007, 404, 115–120. [Google Scholar] [CrossRef]
- Zhang, L.; Zhao, Q.; Mao, L.; Li, H.; Zhuang, M.; Wang, J.; Liu, Y.; Qi, M.; Du, X.; Xia, Z.; et al. Bioinformatics Analyses Reveal the Prognostic Value and Biological Roles of SEPHS2 in Various Cancers. Int. J. Gen. Med. 2021, 14, 6059–6076. [Google Scholar] [CrossRef]
- Nunziata, C.; Polo, A.; Sorice, A.; Capone, F.; Accardo, M.; Guerriero, E.; Marino, F.Z.; Orditura, M.; Budillon, A.; Costantini, S. Structural analysis of human SEPHS2 protein, a selenocysteine machinery component, over-expressed in triple negative breast cancer. Sci. Rep. 2019, 9, 16131. [Google Scholar] [CrossRef]


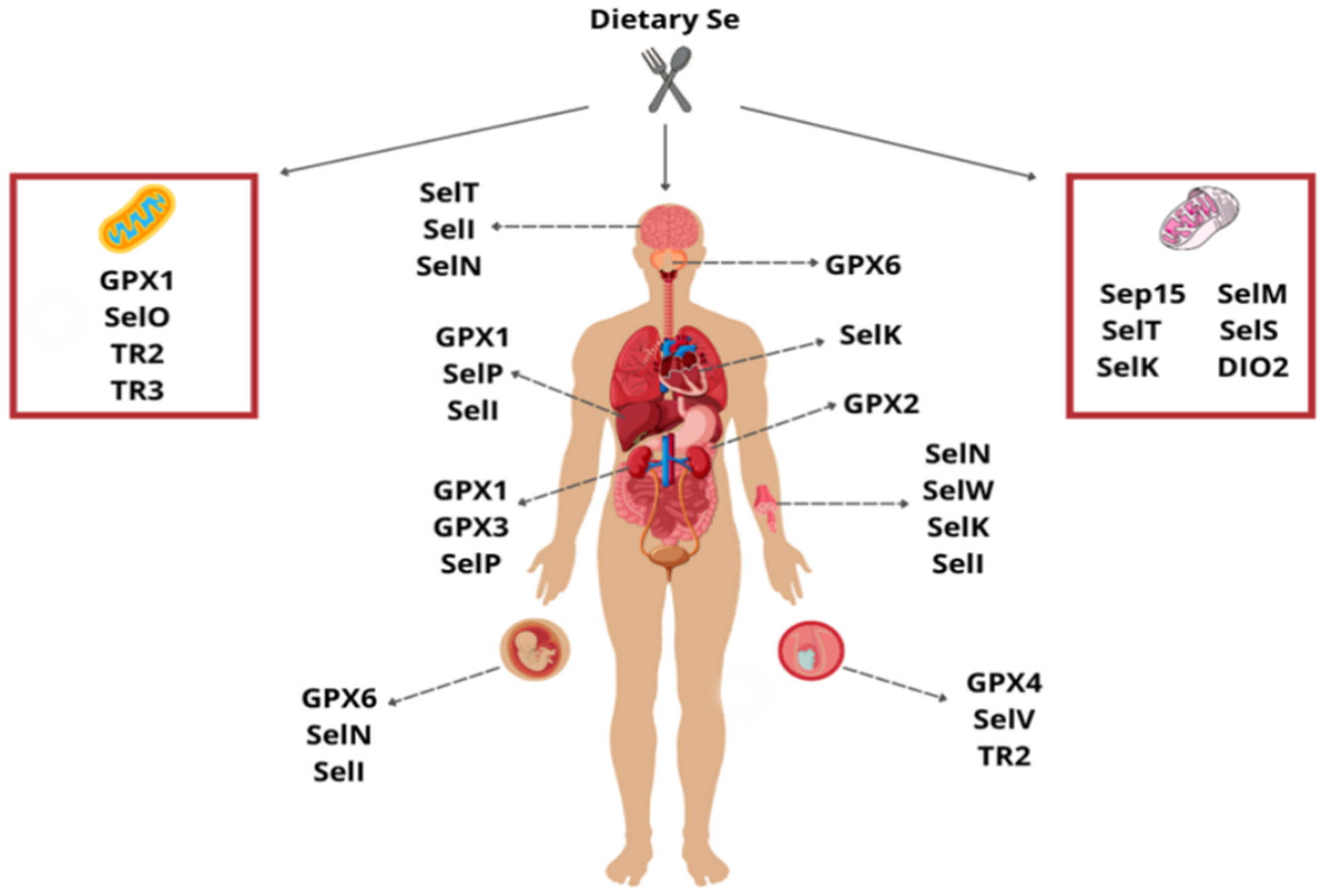{kind=link}
| Selenoprotein Genes | Chromosome | Exons | Function | Ref. |
|---|---|---|---|---|
| SelH | 11q12.1 | 4 | Unknown | Mehta, 2013 [16] |
| GPX6 | 6p22.1 | 5 | Reduction of hydroperoxides | Kryukov, 2003 [17] |
| GPX1 | 3p21.31 | 2 | Reduction of hydroperoxides | Esworthy, 1997 [18] |
| GPX2 | 14q23.3 | 4 | Reduction of hydroperoxides | Roman, 2014 [20] |
| GPX3 | 5q33.1 | 6 | Reduction of hydroperoxides | Takahashi, 1986 [21] |
| GPX4 | 19p13.3 | 8 | Reduction of oxidized phospholipids | Cozza, 2017 [25] |
| SelP | 5p12 | 7 | Sel transportation | Schomburg, 2022 [27] |
| SelF | 1p22.3 | 6 | Protein folding control | Gladyshev, 1998 [28] Labunskyy, 2007 [29] |
| SelS | 15q26.3 | 7 | Production of inflammatory cytokines | Gao, 2006 [30] |
| SelN | 1p36.11 | 13 | Unknown | Taylor, 2009 [31] |
| SelW | 19q13.33 | 6 | Unknown | Vendeland, 1995 [32] |
| SelV | 19q13.2 | 6 | Regulation of O-GlcNAcylation | Mariotti, 2017 [33] |
| SelT | 3q25.1 | 6 | Cell adhesion | Sengupta, 2009 [34] |
| SelI | Motor neuron function | Horibata, 2007 [35] | ||
| SelM | 22q12.2 | 5 | Protection of oxidative stress | Reeves, 2010 [36] |
| SelR | 16p13.3 | 4 | Regulation of methionine by methionine sulfoxide catalysis | Dai, 2016 [37] |
| SelO | 22q13.33 | 9 | Redox homeostasis | Han, 2014 [38] |
| TXNRD1 | 12q23.3 | 18 | Regulatory mechanisms | Su, 2004 [39] |
| TXNRD2 | 22q11.21 | 22 | Regulatory mechanisms | Su, 2004 [39] |
| SelK | 3p21.1 | 5 | Cofactor in protein palmitoylation | Fredericks, 2015 [40] |
| TXNRD3 | 3q21.3 | 16 | Sperm maturation | Wang, 2022 [41] |
| DIO1 | 1p32.3 | 5 | Thyroid hormone regulation by converting T4 to T3 | Jakobs, 1997 [42] |
| DIO2 | 14q31.1 | 7 | Regulation of the HPT axis | Baqui, 2003 [43] |
| DIO3 | 14q32.31 | 1 | Inactivation of thyroid hormone | Baqui, 2003 [43] |
| SEPHS2 | 16p11.2 | 1 | Active selenium donor | Copeland, 2000 [44] |
| Gene | Polymorphism | Country | Disease | Correlation with the Disease | References |
|---|---|---|---|---|---|
| GPX1 | ALA repetition | Italy | Autism | Susceptibility/risk | Carducci, 2022 [45] |
| United States | Autism | Protection | Xue Ming, 2010 [46] | ||
| United Kingdom | Head and neck cancer | Progression | Jefferies, 2005 [47] | ||
| United Kingdom | Prostate cancer | No correlation found | Z Kote-Jarai, 2002 [48] | ||
| Pro198Leu (rs1050450) | Iran | Abortion | No correlation found | Eskafi Sabet, 2014 [49] | |
| Brazil | Alzheimer’s | No correlation found | Rocha, 2016 [50] | ||
| Brazil | Alzheimer’s | Low Se concentration | Cardoso, 2012 [51] | ||
| Brazil | Alzheimer’s | Susceptibility/risk | Da Rocha, 2018 [52] | ||
| Japan | Bladder cancer | Susceptibility/risk | Ichimura, 2004 [53] | ||
| Morocco | Bladder cancer | No correlation found | Hadami, 2016 [54] | ||
| Ecuador | Bladder cancer | Susceptibility/risk | Paz-y-Miño, 2010 [55] | ||
| United States | Bladder cancer | Protection | Zhao H, 2005 [56] | ||
| Serbia | Bladder cancer | Susceptibility/risk | Nikic, 2023 [57] | ||
| Rwanda | Breast cancer | No correlation found | Habyarimana, 2018 [58] | ||
| United States | Breast cancer | No correlation found | Ahn, 2005 [59] | ||
| United States | Breast cancer | No correlation found | Cox, 2004 [60] | ||
| Canada | Breast cancer | No correlation found | Knight, 2004 [61] | ||
| Poland | Breast cancer | Protection | Jablonska, 2015 | ||
| China | Cardiomyopathy | Susceptibility/risk | Lei C, 2009 [62] | ||
| China | Cataract | No correlation found | Zhang, 2011 [63] | ||
| Norway | Colorectal cancer | No correlation found | Hansen, 2005 [64] | ||
| Taiwan | Coronary disease | No correlation found | Hseng-Long Yeh, 2018 [65] | ||
| Sri Lanka | Coronary disease | Susceptibility/risk | Wickremasinghe, 2016 [66] | ||
| Tunisia | Coronary disease | No correlation found | Souiden, 2016 [67] | ||
| Serbia | COVID-19 | No correlation found | Jerotic, 2022 | ||
| Poland | Diabetes | Susceptibility/risk | Buraczynska, 2017 [68] | ||
| Poland | Diabetes | No correlation found | Kasznicki, 2016 [69] | ||
| Pakistan | Diabetes | No correlation found | Mushtaq, 2020 [70] | ||
| Turkey | Diabetes | Susceptibility/risk | Yalin, 2017 [12] | ||
| Poland | Endometrial cancer | No correlation found | Janowska, 2022 [71] | ||
| Turkey | Fibromyalgia | No correlation found | Akbas, 2014 [72] | ||
| Poland | Glaucoma | Susceptibility/risk | Malinowska, 2016 [73] | ||
| China | Kashin–Beck disease | Susceptibility/risk | Xiong, 2010 [74] | ||
| Iran | Keratoconus | Susceptibility/risk | Yari, 2018 [75] | ||
| Turkey | Keratoconus | Susceptibility/risk | Abdullah Ilhan, 2019 [76] | ||
| Denmark | Longevity | Protection | Soerensen, 2009 [77] | ||
| South Korea | Lung cancer | Susceptibility/risk | Chul-Ho Lee, 2006 [78] | ||
| Finland | Lung cancer | Susceptibility/risk | D Ratnasinghe, 2000 [79] | ||
| Iran | Nephrolithiasis | Susceptibility/risk | Aghakhani, 2017 [80] | ||
| Poland | Pancreatitis | Susceptibility/risk | Ściskalska, 2022 [81] | ||
| Turkey | Panic syndrome | No correlation found | Cengiz, 2015 [82] | ||
| Poland | Pharyngeal cancer | No correlation found | Lubiński, 2021 [83] | ||
| Germany | Prostate cancer | Susceptibility/risk | Astrid Steinbrecher, 2010 [84] | ||
| Turkey | Prostate cancer | Progression | Kucukgergin, 2011 [85] | ||
| Turkey | Prostate cancer | No correlation found | Erdem O, 2012 [86] | ||
| Denmark, France, Great Britain, Germany, Greece, Italy, Netherlands, Spain, and Sweden | Prostate cancer | No correlation found | Blein, 2014 [87] | ||
| Macedonia | Prostate cancer | Protection | Arsova-Sarafinovska, 2009 [88] | ||
| Iran | Pulmonary embolism | Susceptibility/risk | Teimoori, 2019 [89] | ||
| Russia | Restenosis | Susceptibility/risk | Shuvalova, 2012 [90] | ||
| Taiwan | Steate hepatitis | No correlation found | Huang, 2021 [91] | ||
| Iran | Thyroid carcinoma | No correlation found | Salimi, 2020 [92] | ||
| rs1800668 and rs1050450 | China | Schizophrenia | Susceptibility/risk | Xiaojun Shao, 2020 [93] | |
| rs1050450 and rs18006688 | United States | Brain tumor | Susceptibility/risk | Bhatti, 2009 [94] | |
| rs1800668 | New Zealand | Crohn’s disease | Susceptibility/risk | Morgan, 2010 [95] | |
| China | Schizophrenia | Protection | Xiaojun Shao, 2020 [93] | ||
| GPX3 | rs736775 | China | Macrovascular disease | No correlation found | Liu C, 2020 [96] |
| rs8177409 | Mexico | Metabolic syndrome | Susceptibility/risk | Baez-Duarte, 2014 [97] | |
| rs3805435 | Taiwan | Hearing loss | Protection | Chien, 2017 [98] | |
| rs3763013, rs8177412, rs3828599, and rs2070593 | Taiwan | Hearing loss | No correlation found | Chien, 2017 [98] | |
| rs8177412 | Serbia | Testicular tumor | Susceptibility/risk | Bumbasirevic, 2022 [99] | |
| GPX4 | China | Preeclampsia | No correlation found | Peng, 2016 [100] | |
| rs4965814 and rs9874 | Finland | Cardiovascular disease | Susceptibility/risk | Silander, 2008 [101] | |
| rs713041 | Scotland | Vascular disease | Susceptibility/risk | Crosley, 2013 [102] | |
| Russia | Stroke | Susceptibility/risk | Polonikov, 2012 [103] | ||
| Taiwan | Endometriosis | Progression | Huang, 2020 [104] | ||
| Poland | Pancreatitis | Susceptibility/risk | Ściskalska, 2022 [81] | ||
| China | Preeclampsia | Susceptibility/risk | Chen, 2020 [105] | ||
| China | Preeclampsia | Susceptibility/risk | Peng, 2016 [100] | ||
| Brazil | Diabetes | Protection | Admoni, 2019 [106] | ||
| rs713041, rs2074451, and rs3746165 | China | Thyroid disease | No correlation found | Xiao, 2017 [107] | |
| DIO1 | rs11206244 | United States | Depression | Susceptibility/risk | Philibert, 2011 [108] |
| DIO2 | rs12885300 | Brazil | Graves’ disease | Susceptibility/risk | Comarella, 2021 [109] |
| rs225014 and rs225015 | Lithuania | Stroke | No correlation found | Taroza, 2019 [110] | |
| Thr92Ala [rs225014] | Brazil | COVID-19 | Protection | Beltrão, 2022 [111] | |
| Italy | Obesity | No correlation found | Benenati, 2022 [112] | ||
| Brazil | Cognitive impairment | No correlation found | Schwengber, 2022 [113] | ||
| DIO3 | rs945006 | Lithuania | Stroke | Progression | Taroza, 2019 [110] |
| rs12095080, rs11206244, and rs2235544 | Lithuania | Stroke | No correlation found | Taroza, 2019 [110] | |
| TXNRD1 | rs35009941 | United States | Colorectal cancer | Protection | Peters, 2008 [114] |
| TXNRD2 | rs1139793 | China | Kashin–Beck | No correlation found | Wu, 2019 [115] |
| rs4485648 | Slovenia | Diabetes | Susceptibility/risk | Kariž, 2015 [116] | |
| rs5748469 | China | Kashin–Beck | No correlation found | Lu, 2010 [117] | |
| rs5746841 | China | Kashin–Beck | No correlation found | Li, 2019 [118] | |
| SelF | 1125 G/A | Poland | Lung cancer | Modifies the selenium level | Jablonska, 2015 |
| rs5845 and rs5859 | South Korea | Colorectal cancer | Susceptibility/risk | Sutherland, 2010 [119] | |
| rs5859 | China | Kashin–Beck | Susceptibility/risk | Wu, 2019 [115] | |
| rs5859 | Brazil | AIDS | Progression | Benelli, 2016 [120] | |
| SelS | rs34713741 | South Korea | Colorectal cancer | Susceptibility/risk | Sutherland, 2010 [119] |
| -105G --> A | Germany | Inflammatory bowel disease | No correlation found | Seiderer, 2007 | |
| -105G --> A | Japan | Gastric cancer | Susceptibility/risk | Tomoyuki, 2009 [121] | |
| -105G>A | China | Premature birth | Susceptibility/risk | Wang, 2013 [122] | |
| -105G>A | Portugal | Hashimoto’s thyroiditis | Susceptibility/risk | Santos, 2014 [123] | |
| rs11327127, rs28665122, rs4965814, rs12917258, rs4965373, and rs2101171 | Spain | Autoimmune inflammatory disease | No correlation found | Martínez, 2008 [124] | |
| SelP | rs7579 | China | Metabolic syndrome | Susceptibility/risk | Li Zhou, 2020 [125] |
| rs3877899 | United States | Breast cancer | Susceptibility/risk | Mohammaddoust, 2018 [126] | |
| rs11959466 | United States | Prostate cancer | Susceptibility/risk | Penney, 2013 [127] | |
| rs13168440 | United States | Prostate cancer | Protection | Penney, 2013 [127] | |
| rs230820 | United States | Prostate cancer | No correlation found | Xie, 2016 [128] | |
| r25191G/A | China | Kashin–Beck | No correlation found | Sun, 2010 [44] | |
| rs7579 | Germany | Prostate cancer | Susceptibility/risk | Astrid Steinbrecher, 2010 [84] | |
| rs146125471, rs28919926, and rs16872762 | Turkey | Diabetes | No correlation found | Akbaba, 2018 [129] | |
| rs13154178 | Turkey | Diabetes | Susceptibility/risk | Akbaba, 2018 [129] | |
| rs7579 | Poland | Endothelial cancer | No correlation found | Janowska, 2022 [71] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, R.R.; Carvalho, R.V.; Coelho, L.L.; Gonzaga, B.M.d.S.; Bonecini-Almeida, M.d.G.; Garzoni, L.R.; Araujo-Jorge, T.C. Current Understanding of Human Polymorphism in Selenoprotein Genes: A Review of Its Significance as a Risk Biomarker. Int. J. Mol. Sci. 2024, 25, 1402. https://doi.org/10.3390/ijms25031402
Ferreira RR, Carvalho RV, Coelho LL, Gonzaga BMdS, Bonecini-Almeida MdG, Garzoni LR, Araujo-Jorge TC. Current Understanding of Human Polymorphism in Selenoprotein Genes: A Review of Its Significance as a Risk Biomarker. International Journal of Molecular Sciences. 2024; 25(3):1402. https://doi.org/10.3390/ijms25031402
Chicago/Turabian StyleFerreira, Roberto Rodrigues, Regina Vieira Carvalho, Laura Lacerda Coelho, Beatriz Matheus de Souza Gonzaga, Maria da Gloria Bonecini-Almeida, Luciana Ribeiro Garzoni, and Tania C. Araujo-Jorge. 2024. "Current Understanding of Human Polymorphism in Selenoprotein Genes: A Review of Its Significance as a Risk Biomarker" International Journal of Molecular Sciences 25, no. 3: 1402. https://doi.org/10.3390/ijms25031402