

The Concordant Disruption of B7/CD28 Immune Regulators Predicts the Prognosis of Oral Carcinomas
 ,
,
Abstract
:1. Introduction
2. Results
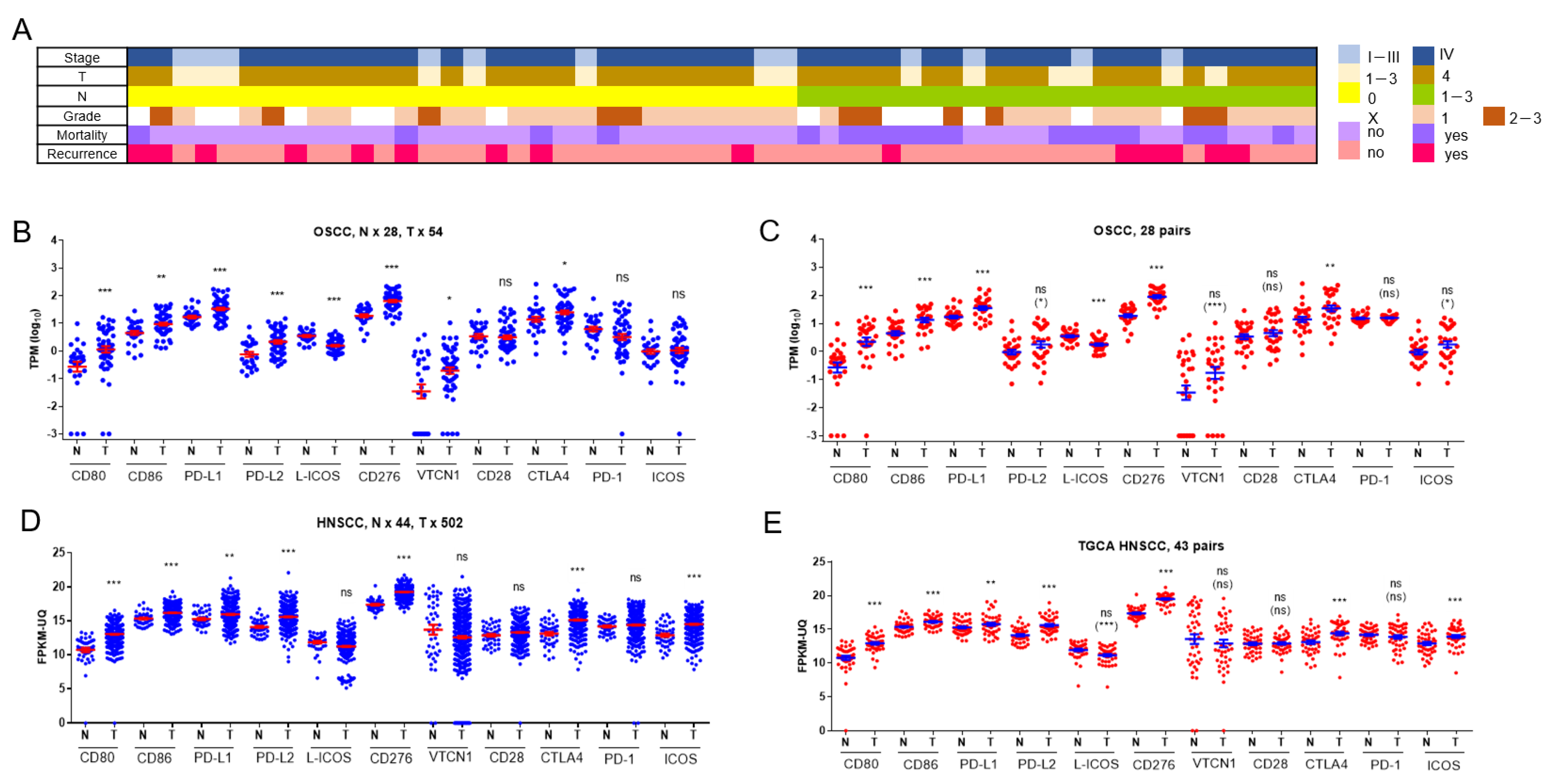
2.1. Aberrances in the Expression of B7/CD28 Family Members in OSCC
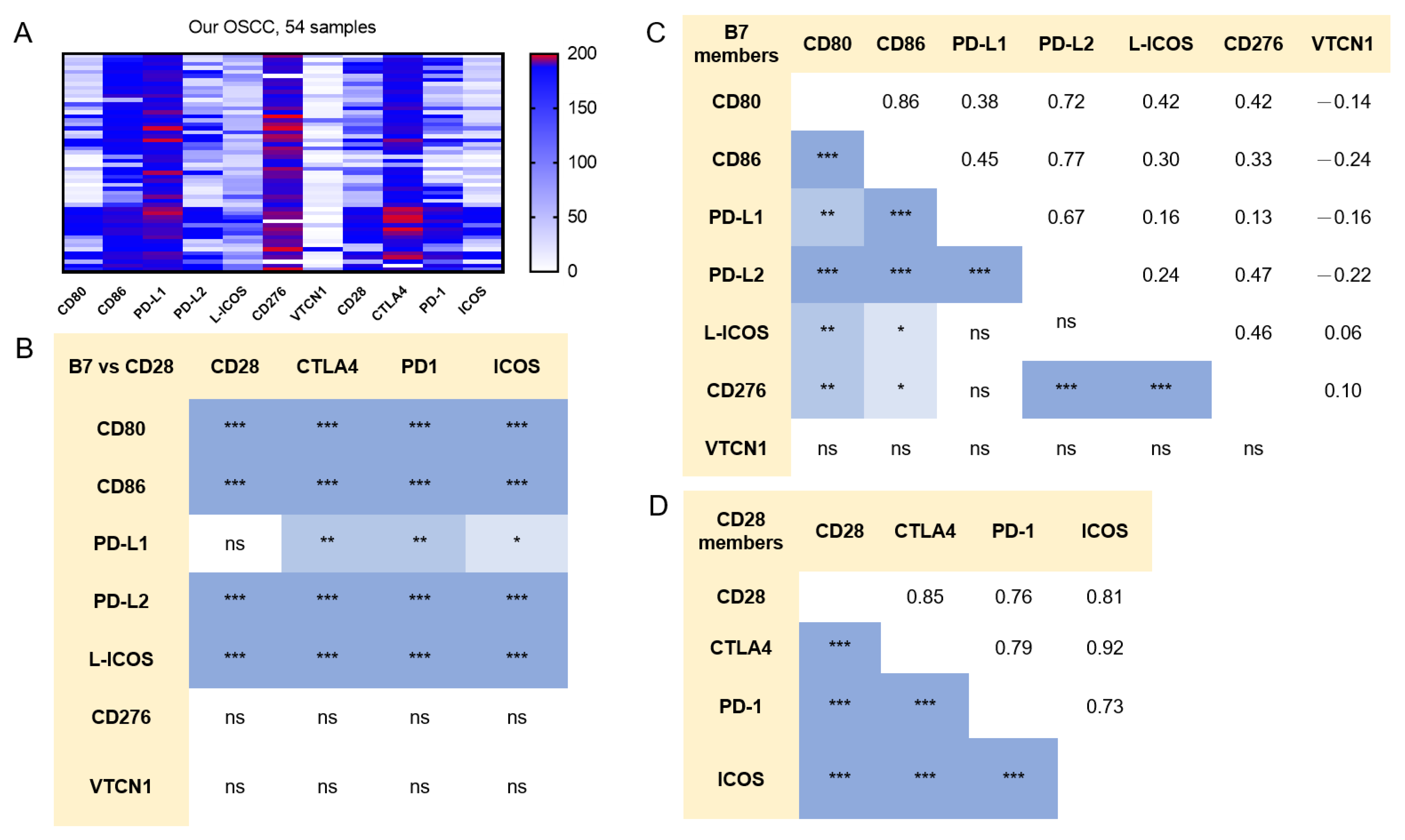
2.2. Concordance in the Expression of B7/CD28 Family Members in OSCC
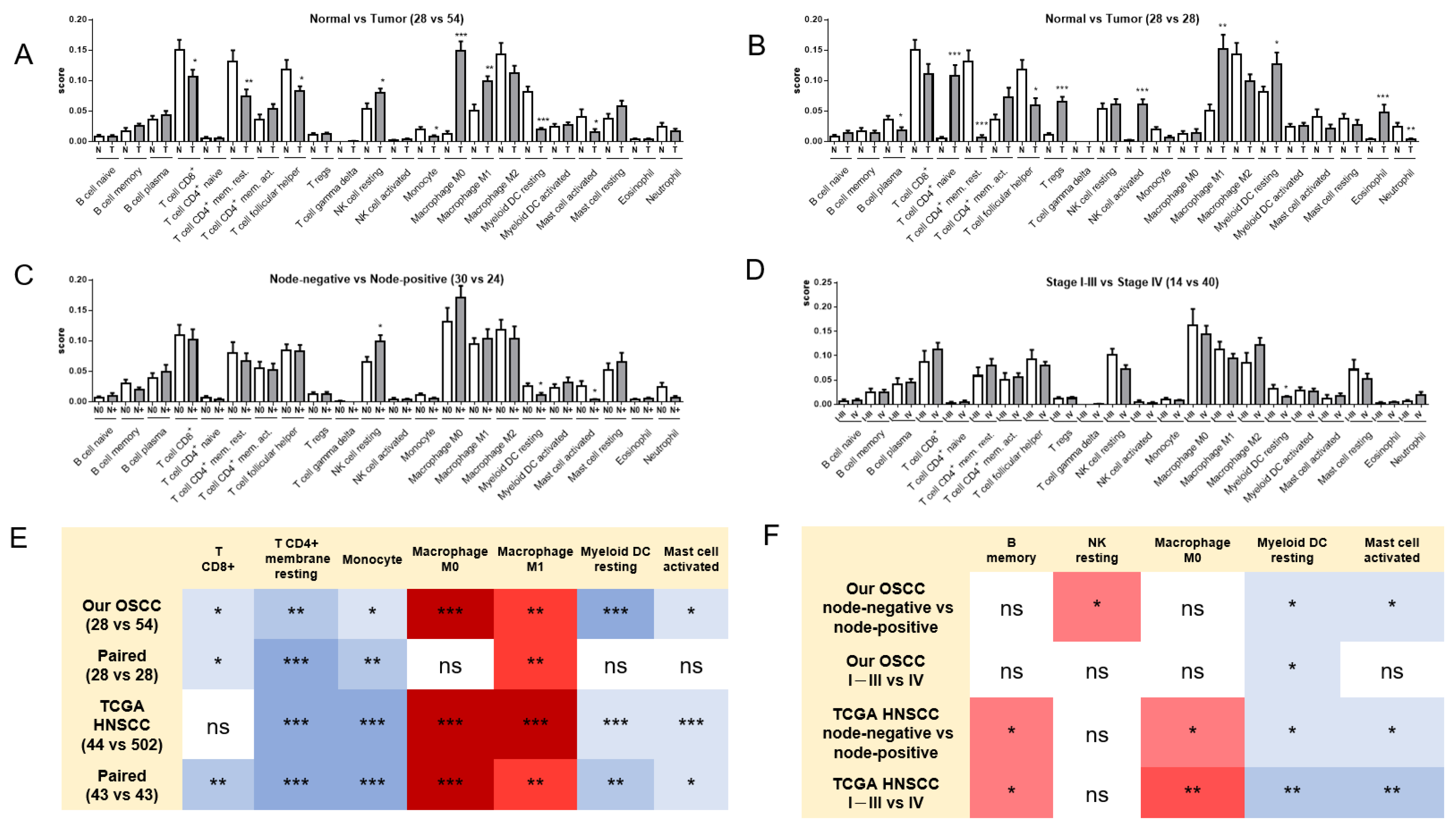
2.3. Alterations in the Immune Cell Population in OSCC
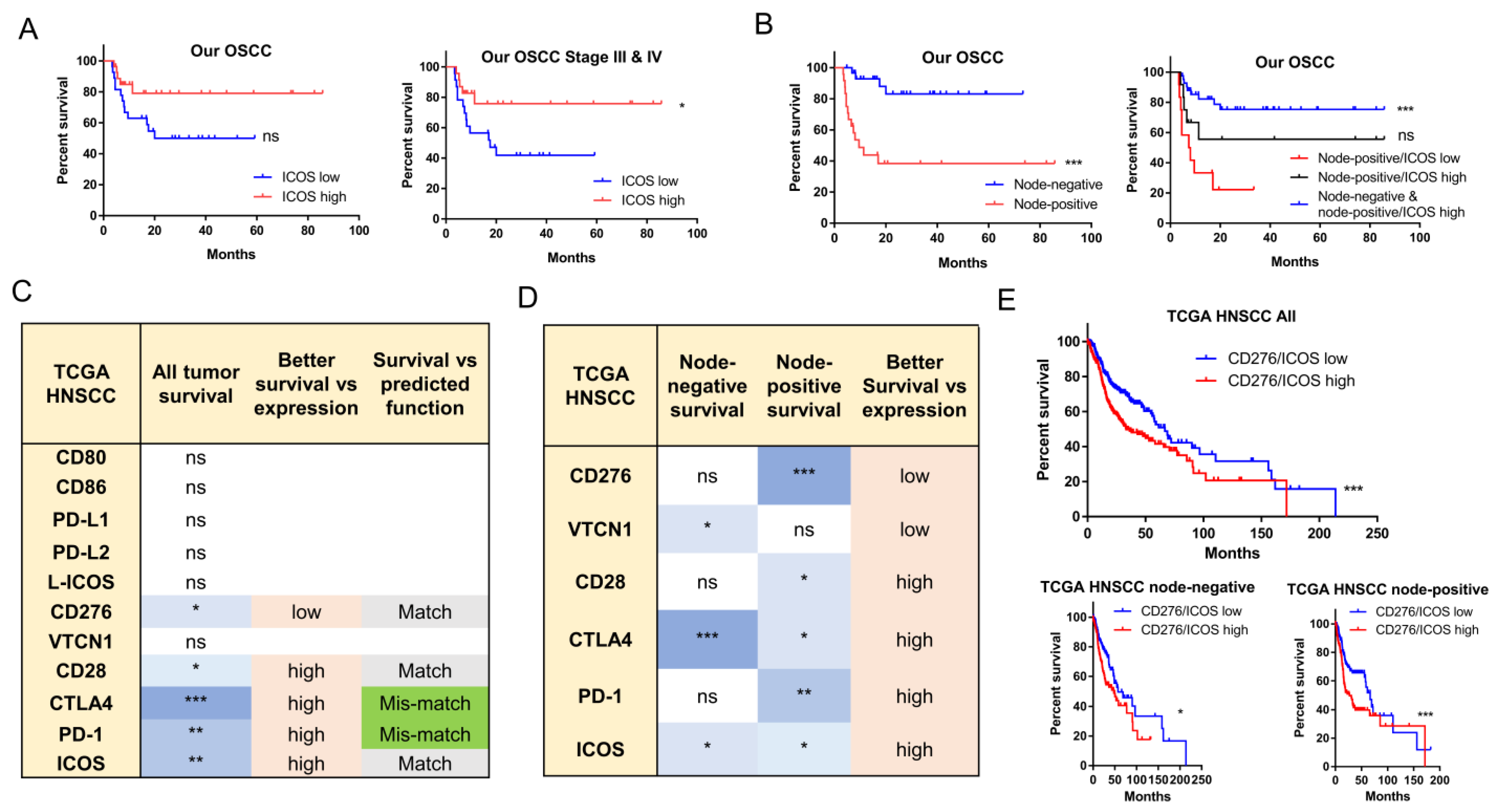
2.4. The Prognostic Implications of ICOS and CD276
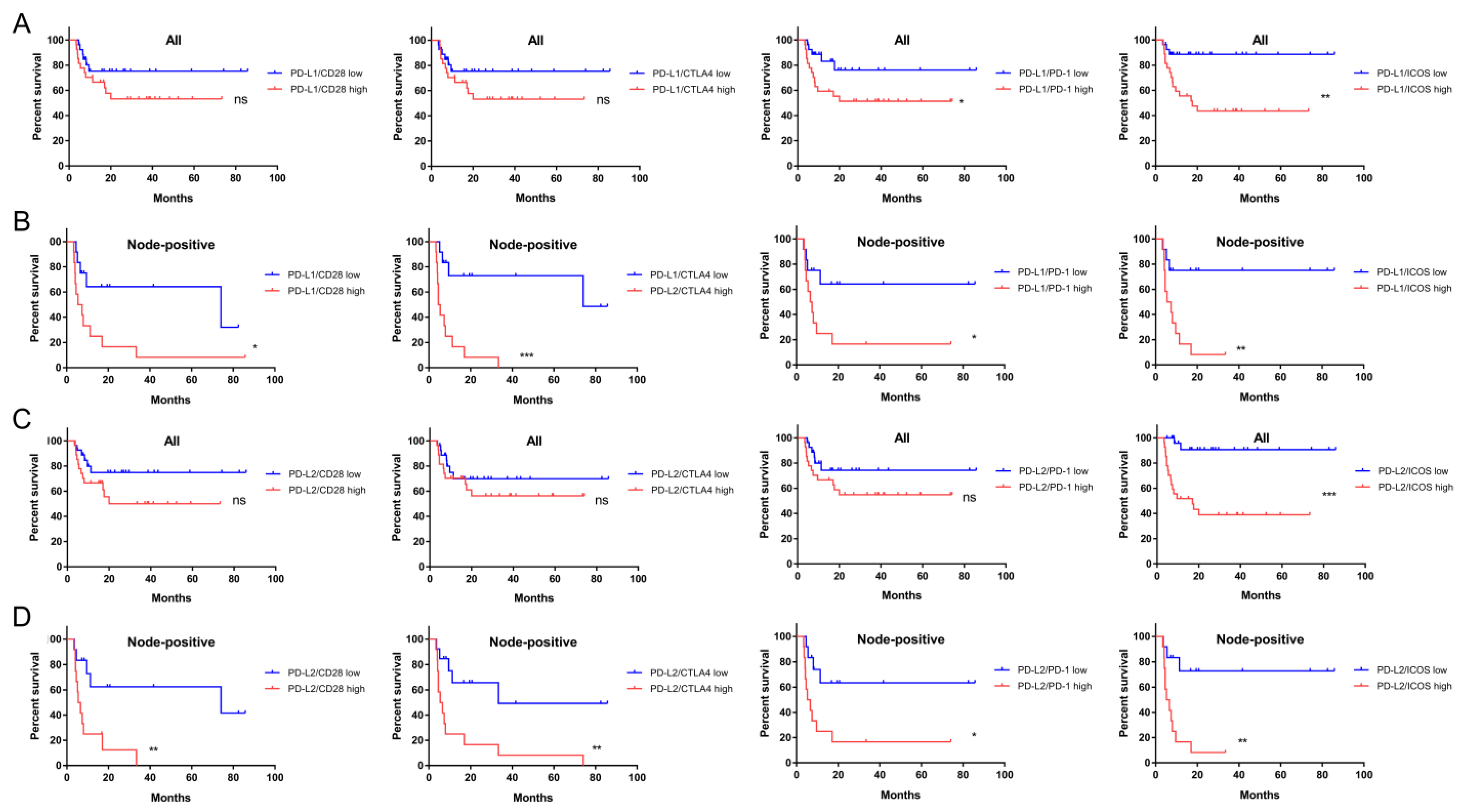
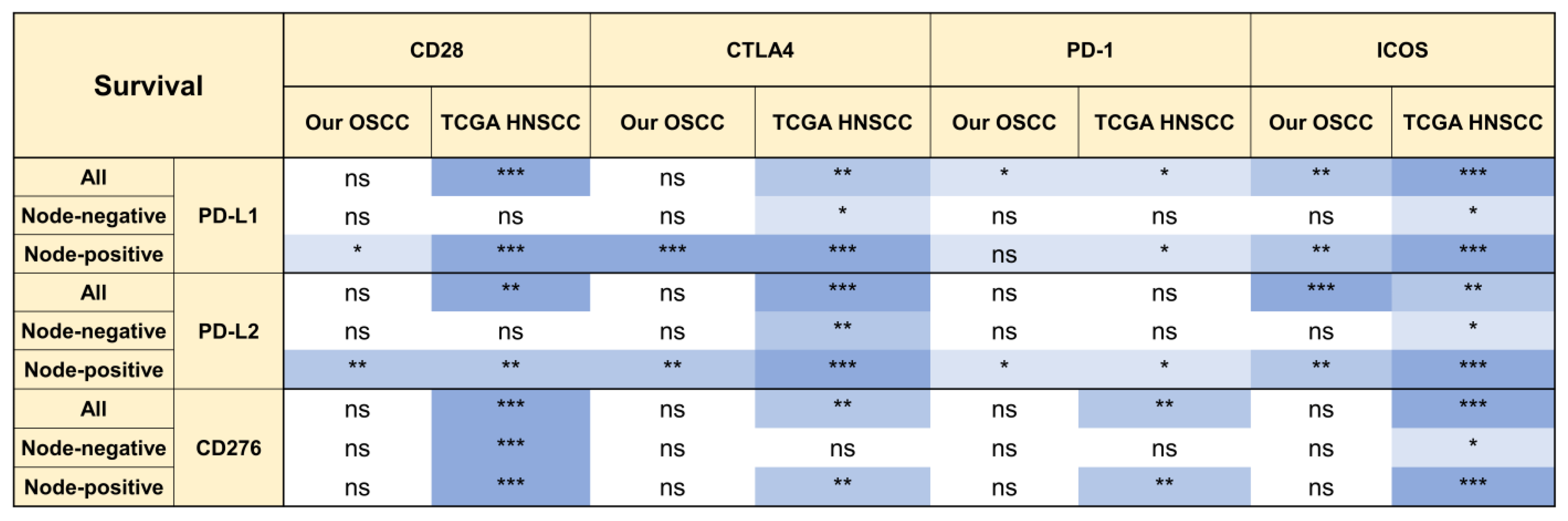
2.5. The Prognostic Implications of PD-L1, PD-L2, and CD276 in Relation to CD28 Members
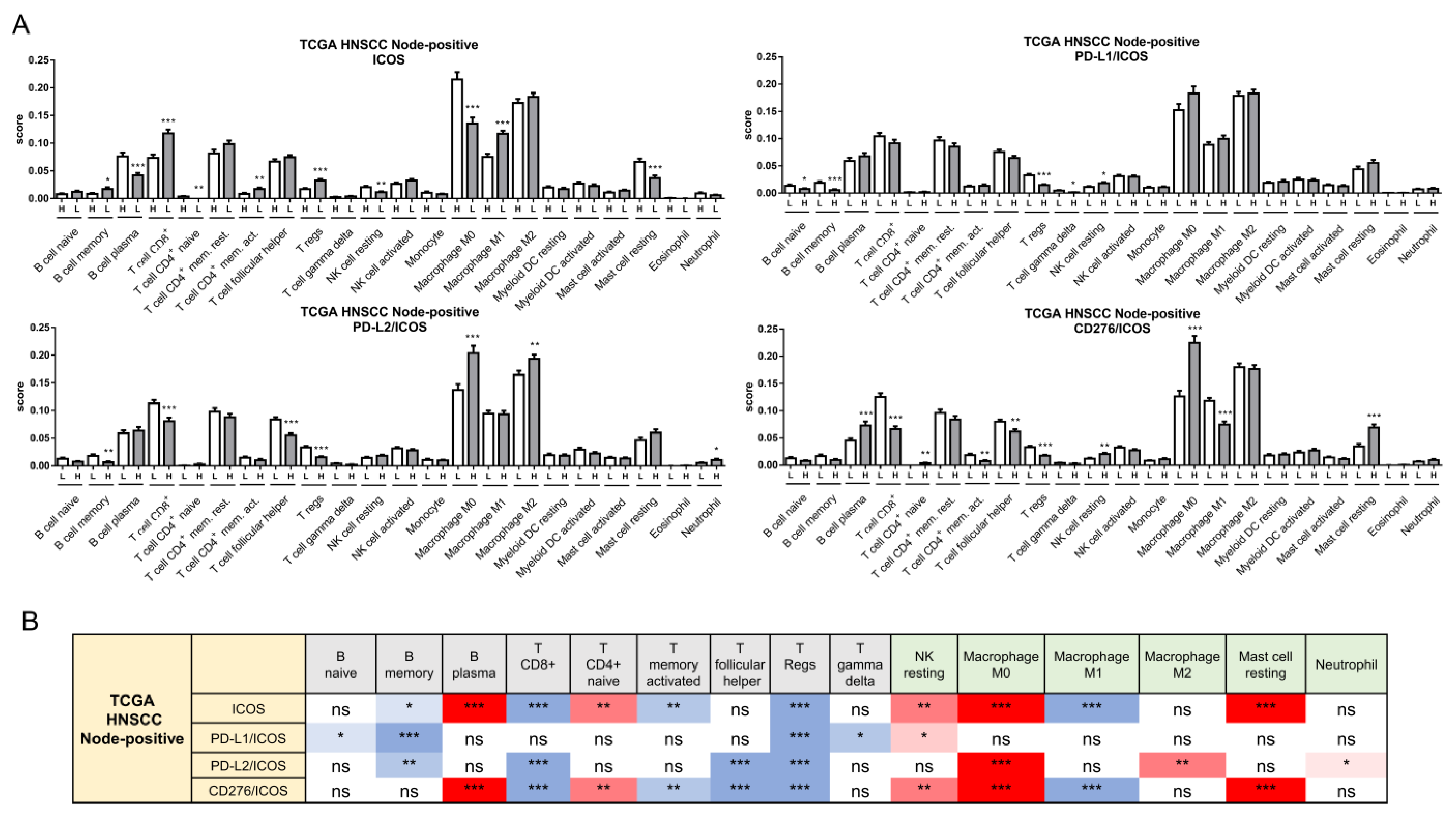
2.6. ICOS Expression and Ratios of PD-L1, PD-L2, and CD276 against ICOS Expression Were Independent Survival Predictors of Node-Positive HNSCC
2.7. Alteration of the Immune Cell Population Associated with Worse Survival in Node-Positive Patients
3. Discussion
4. Materials and Methods
4.1. Samples
4.2. RNA Extraction, RNA Sequencing, and Raw Data Filtering
4.3. Differential Expression Profiles
4.4. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yeh, C.C.; Kao, H.K.; Huang, Y.; Tsai, T.Y.; Young, C.K.; Hung, S.Y.; Lu, C.Y.; Chang, K.P. Discovering the Clinical and Prognostic Role of Pan-Immune-Inflammation Values on Oral Cavity Squamous Cell Carcinoma. Cancers 2023, 15, 322. [Google Scholar] [CrossRef]
- Zhu, C.; Wu, Q.; Yang, N.; Zheng, Z.; Zhou, F.; Zhou, Y. Immune Infiltration Characteristics and a Gene Prognostic Signature Associated with the Immune Infiltration in Head and Neck Squamous Cell Carcinoma. Front. Genet. 2022, 13, 848841. [Google Scholar] [CrossRef]
- Weber, M.; Lutz, R.; Olmos, M.; Glajzer, J.; Baran, C.; Nobis, C.P.; Most, T.; Eckstein, M.; Kesting, M.; Ries, J. Beyond PD-L1-Identification of Further Potential Therapeutic Targets in Oral Cancer. Cancers 2022, 14, 1812. [Google Scholar] [CrossRef]
- Xavier, F.C.A.; Silva, J.C.; Rodini, C.O.; Rodrigues, M. Mechanisms of immune evasion by head and neck cancer stem cells. Front. Oral. Health 2022, 3, 957310. [Google Scholar] [CrossRef]
- Li, G.; Quan, Y.; Che, F.; Wang, L. B7-H3 in tumors: Friend or foe for tumor immunity? Cancer Chemother. Pharmacol. 2018, 81, 245–253. [Google Scholar] [CrossRef]
- Sim, F.; Leidner, R.; Bell, R.B. Immunotherapy for Head and Neck Cancer. Oral. Maxillofac. Surg. Clin. N. Am. 2019, 31, 85–100. [Google Scholar] [CrossRef]
- Alegre, M.L.; Frauwirth, K.A.; Thompson, C.B. T-cell regulation by CD28 and CTLA-4. Nat. Rev. Immunol. 2001, 1, 220–228. [Google Scholar] [CrossRef]
- Yu, G.T.; Bu, L.L.; Zhao, Y.Y.; Mao, L.; Deng, W.W.; Wu, T.F.; Zhang, W.F.; Sun, Z.J. CTLA4 blockade reduces immature myeloid cells in head and neck squamous cell carcinoma. Oncoimmunology 2016, 5, e1151594. [Google Scholar] [CrossRef] [Green Version]
- Zenke, S.; Palm, M.M.; Braun, J.; Gavrilov, A.; Meiser, P.; Bottcher, J.P.; Beyersdorf, N.; Ehl, S.; Gerard, A.; Lammermann, T.; et al. Quorum Regulation via Nested Antagonistic Feedback Circuits Mediated by the Receptors CD28 and CTLA-4 Confers Robustness to T Cell Population Dynamics. Immunity 2020, 52, 313–327.e7. [Google Scholar] [CrossRef]
- Zenke, S.; Sica, M.P.; Steinberg, F.; Braun, J.; Zink, A.; Gavrilov, A.; Hilger, A.; Arra, A.; Brunner-Weinzierl, M.; Elling, R.; et al. Differential trafficking of ligands trogocytosed via CD28 versus CTLA4 promotes collective cellular control of co-stimulation. Nat. Commun. 2022, 13, 6459. [Google Scholar] [CrossRef]
- Ghiotto, M.; Gauthier, L.; Serriari, N.; Pastor, S.; Truneh, A.; Nunes, J.A.; Olive, D. PD-L1 and PD-L2 differ in their molecular mechanisms of interaction with PD-1. Int. Immunol. 2010, 22, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Youngnak, P.; Kozono, Y.; Kozono, H.; Iwai, H.; Otsuki, N.; Jin, H.; Omura, K.; Yagita, H.; Pardoll, D.M.; Chen, L.; et al. Differential binding properties of B7-H1 and B7-DC to programmed death-1. Biochem. Biophys. Res. Commun. 2003, 307, 672–677. [Google Scholar] [CrossRef]
- Song, D.; Tian, J.; Han, X.; Li, X. A model of seven immune checkpoint-related genes predicting overall survival for head and neck squamous cell carcinoma. Eur. Arch. Otorhinolaryngol. 2021, 278, 3467–3477. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.; Jiang, X.; Mo, B.; Wang, C.; Tang, M.; Rong, Y.; Zhang, G.; Hu, M.; Cai, H. Comprehensive analysis of the role of ICOS (CD278) in pan-cancer prognosis and immunotherapy. BMC Cancer 2023, 23, 194. [Google Scholar] [CrossRef]
- Ciprut, S.; Berberich, A.; Knoll, M.; Pusch, S.; Hoffmann, D.; Furkel, J.; Ward Gahlawat, A.; Kahlert-Konzelamnn, L.; Sahm, F.; Warnken, U.; et al. AAMP is a binding partner of costimulatory human B7-H3. Neurooncol. Adv. 2022, 4, vdac098. [Google Scholar] [CrossRef] [PubMed]
- Borgmann, M.; Oetting, A.; Meyer, F.; Mockelmann, N.; Droste, C.; von Bargen, C.M.; Moller-Koop, C.; Witt, M.; Borgmann, K.; Rothkamm, K.; et al. The prognostic impact of B7-H3 and B7-H4 in head and neck squamous cell carcinoma. J. Cancer Res. Clin. Oncol. 2022, 1–11. [Google Scholar] [CrossRef]
- Chen, J.T.; Chen, C.H.; Ku, K.L.; Hsiao, M.; Chiang, C.P.; Hsu, T.L.; Chen, M.H.; Wong, C.H. Glycoprotein B7-H3 overexpression and aberrant glycosylation in oral cancer and immune response. Proc. Natl. Acad. Sci. USA 2015, 112, 13057–13062. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Li, Y.; Jia, L.; Kim, J.K.; Li, J.; Deng, P.; Zhang, W.; Krebsbach, P.H.; Wang, C.Y. CD276 expression enables squamous cell carcinoma stem cells to evade immune surveillance. Cell Stem Cell 2021, 28, 1597–1613.e7. [Google Scholar] [CrossRef] [PubMed]
- Katayama, A.; Takahara, M.; Kishibe, K.; Nagato, T.; Kunibe, I.; Katada, A.; Hayashi, T.; Harabuchi, Y. Expression of B7-H3 in hypopharyngeal squamous cell carcinoma as a predictive indicator for tumor metastasis and prognosis. Int. J. Oncol. 2011, 38, 1219–1226. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Xu, Y.; Gao, J.; Zhang, H.; Sun, Y.; Qiu, X.; Huang, Q.; Kong, L.; Lu, J.J. Multi-Omics Data Analyses Identify B7-H3 as a Novel Prognostic Biomarker and Predict Response to Immune Checkpoint Blockade in Head and Neck Squamous Cell Carcinoma. Front. Immunol. 2021, 12, 757047. [Google Scholar] [CrossRef]
- He, F.; Chen, Z.; Deng, W.; Zhan, T.; Huang, X.; Zheng, Y.; Yang, H. Development and validation of a novel ferroptosis-related gene signature for predicting prognosis and immune microenvironment in head and neck squamous cell carcinoma. Int. Immunopharmacol. 2021, 98, 107789. [Google Scholar] [CrossRef]
- Wu, L.; Deng, W.W.; Yu, G.T.; Mao, L.; Bu, L.L.; Ma, S.R.; Liu, B.; Zhang, W.F.; Sun, Z.J. B7-H4 expression indicates poor prognosis of oral squamous cell carcinoma. Cancer Immunol. Immunother. 2016, 65, 1035–1045. [Google Scholar] [CrossRef]
- Deng, W.W.; Wu, L.; Sun, Z.J. Co-inhibitory immune checkpoints in head and neck squamous cell carcinoma. Oral. Dis. 2018, 24, 120–123. [Google Scholar] [CrossRef]
- Yu, C.; Li, Q.; Zhang, Y.; Wen, Z.F.; Dong, H.; Mou, Y. Current status and perspective of tumor immunotherapy for head and neck squamous cell carcinoma. Front. Cell Dev. Biol. 2022, 10, 941750. [Google Scholar] [CrossRef] [PubMed]
- Masarwy, R.; Kampel, L.; Horowitz, G.; Gutfeld, O.; Muhanna, N. Neoadjuvant PD-1/PD-L1 Inhibitors for Resectable Head and Neck Cancer: A Systematic Review and Meta-analysis. JAMA Otolaryngol. Head Neck Surg. 2021, 147, 871–878. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Li, X.; Huang, L.; Xu, Z.; Lin, J. Prognostic value of PD-1, PD-L1 and PD-L2 deserves attention in head and neck cancer. Front. Immunol. 2022, 13, 988416. [Google Scholar] [CrossRef] [PubMed]
- Qiao, Y.; Liu, C.; Zhang, X.; Zhou, Q.; Li, Y.; Xu, Y.; Gao, Z.; Xu, Y.; Kong, L.; Yang, A.; et al. PD-L2 based immune signature confers poor prognosis in HNSCC. Oncoimmunology 2021, 10, 1947569. [Google Scholar] [CrossRef]
- Aggarwal, C.; Prawira, A.; Antonia, S.; Rahma, O.; Tolcher, A.; Cohen, R.B.; Lou, Y.; Hauke, R.; Vogelzang, N.D.P.Z.; Kalebasty, A.R.; et al. Dual checkpoint targeting of B7-H3 and PD-1 with enoblituzumab and pembrolizumab in advanced solid tumors: Interim results from a multicenter phase I/II trial. J. Immunother. Cancer. 2022, 10, e004424. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, L.; Liu, S. The Immune Infiltration in HNSCC and Its Clinical Value: A Comprehensive Study Based on the TCGA and GEO Databases. Comput. Math. Methods Med. 2021, 2021, 1163250. [Google Scholar] [CrossRef] [PubMed]
- Duhen, R.; Fesneau, O.; Samson, K.A.; Frye, A.K.; Beymer, M.; Rajamanickam, V.; Ross, D.; Tran, E.; Bernard, B.; Weinberg, A.D.; et al. PD-1 and ICOS coexpression identifies tumor-reactive CD4+ T cells in human solid tumors. J. Clin. Investig. 2022, 132, e156821. [Google Scholar] [CrossRef]
- Kurten CH, L.; Kulkarni, A.; Cillo, A.R.; Santos, P.M.; Roble, A.K.; Onkar, S.; Reeder, C.; Lang, S.; Chen, X.; Duvvuri, U.; et al. Investigating immune and non-immune cell interactions in head and neck tumors by single-cell RNA sequencing. Nat. Commun. 2021, 12, 7338. [Google Scholar] [CrossRef] [PubMed]
- Chang, K.W.; Hung, W.W.; Chou, C.H.; Tu, H.F.; Chang, S.R.; Liu, Y.C.; Liu, C.J.; Lin, S.C. LncRNA MIR31HG Drives Oncogenicity by Inhibiting the Limb-Bud and Heart Development Gene (LBH) during Oral Carcinoma. Int. J. Mol. Sci. 2021, 22, 8383. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Ye, Z.; Stanton, R. Misuse of RPKM or TPM normalization when comparing across samples and sequencing protocols. RNA 2020, 26, 903–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, B.; Zhao, M.; Wu, J.; Wu, H.; Zhao, Y.; Meng, F.; Tian, Y.; Wang, S.; Shi, J.; Zhang, H. A Novel RNA-Seq-Based Model for Preoperative Prediction of Lymph Node Metastasis in Oral Squamous Cell Carcinoma. Biomed. Res. Int. 2020, 2020, 4252580. [Google Scholar] [CrossRef] [PubMed]
- Rau, A.; Flister, M.; Rui, H.; Auer, P.L. Exploring drivers of gene expression in the Cancer Genome Atlas. Bioinformatics 2019, 35, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Newman, A.M.; Liu, C.L.; Green, M.R.; Gentles, A.J.; Feng, W.; Xu, Y.; Hoang, C.D.; Diehn, M.; Alizadeh, A.A. Robust enumeration of cell subsets from tissue expression profiles. Nat. Methods 2015, 12, 453–457. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TCGA HNSCC Node-Positive | Logistic Regression (Univariate) | Logistic Regression (Multivariate) | Cox Proportional Hazards Regression | |||||
|---|---|---|---|---|---|---|---|---|
| OR | 95% CI | p | OR | 95% CI | p | HR | 95% CI | |
| CD276 | 1.33 | 1.02 to 1.74 | 0.024 * | 1.39 | 1.06 to 1.84 | ns | 1.30 | 1.06 to 1.60 |
| ICOS | 0.79 | 0.67 to 0.93 | 0.018 * | 0.77 | 0.65 to 0.91 | 0.0018 ** | 0.85 | 0.76 to 0.95 |
| PD-L1/CD28 | 11.61 | 2.14 to 67 | 0.003 ** | 0.18 | 0.01 to 5.61 | ns | 0.28 | 0.03 to 2.99 |
| PD-L1/CTLA4 | 173 | 16.87 to 2194 | <0.0001 *** | 116 | 0.33 to 83,302 | ns | 2.30 | 0.05 to 100 |
| PD-L1/PD-1 | 69 | 8.21 to 678 | 0.0002 *** | 11.45 | 0.32 to 424 | ns | 10.73 | 1.01 to 111 |
| PD-L1/ICOS | 76 | 9.02 to 739 | 0.0001 *** | 1.05 | 0.00 to 594 | <0.0001 *** | 3.33 | 0.04 to 226 |
| PD-L2/CD28 | 7.78 | 1.45 to 45.51 | 0.012 * | 0.16 | 0.00 to 5.21 | ns | 0.20 | 0.02 to 2.00 |
| PD-L2/CTLA4 | 114 | 10.88 to 1547 | <0.0001 *** | 110 | 0.27 to 86,428 | ns | 1.49 | 0.04 to 63 |
| PD-L2/PD-1 | 23.33 | 3.41 to 177 | 0.001 *** | 2.26 | 0.06 to 76 | ns | 6.04 | 0.62 to 57 |
| PD-L2/ICOS | 80 | 8.05 to 961 | 0.0004 *** | 3.40 | 0.00 to 2670 | <0.0001 *** | 13.00 | 0.12 to 996 |
| CD276/CD28 | 5.44 | 1.45 to 21.66 | 0.007 ** | 0.51 | 0.03 to 8.11 | ns | 0.56 | 0.08 to 3.81 |
| CD276/CTLA4 | 41.50 | 6.70 to 304 | <0.0001 *** | 298 | 1.88 to 95,050 | ns | 3.56 | 0.22 to 58 |
| CD276/PD-1 | 8.98 | 2.15 to 40.14 | 0.003 ** | 0.77 | 0.04 to 12.82 | ns | 3.39 | 0.54 to 21.00 |
| CD276/ICOS | 17.87 | 3.63 to 103 | 0.001 *** | 0.33 | 0.00 to 52 | 0.0001 *** | 1.24 | 0.04 to 28.76 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, S.-R.; Chou, C.-H.; Liu, C.-J.; Lin, Y.-C.; Tu, H.-F.; Chang, K.-W.; Lin, S.-C. The Concordant Disruption of B7/CD28 Immune Regulators Predicts the Prognosis of Oral Carcinomas. Int. J. Mol. Sci. 2023, 24, 5931. https://doi.org/10.3390/ijms24065931
Chang S-R, Chou C-H, Liu C-J, Lin Y-C, Tu H-F, Chang K-W, Lin S-C. The Concordant Disruption of B7/CD28 Immune Regulators Predicts the Prognosis of Oral Carcinomas. International Journal of Molecular Sciences. 2023; 24(6):5931. https://doi.org/10.3390/ijms24065931
Chicago/Turabian StyleChang, Shi-Rou, Chung-Hsien Chou, Chung-Ji Liu, Yu-Cheng Lin, Hsi-Feng Tu, Kuo-Wei Chang, and Shu-Chun Lin. 2023. "The Concordant Disruption of B7/CD28 Immune Regulators Predicts the Prognosis of Oral Carcinomas" International Journal of Molecular Sciences 24, no. 6: 5931. https://doi.org/10.3390/ijms24065931