

Semilicoisoflavone B Induces Apoptosis of Oral Cancer Cells by Inducing ROS Production and Downregulating MAPK and Ras/Raf/MEK Signaling
 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
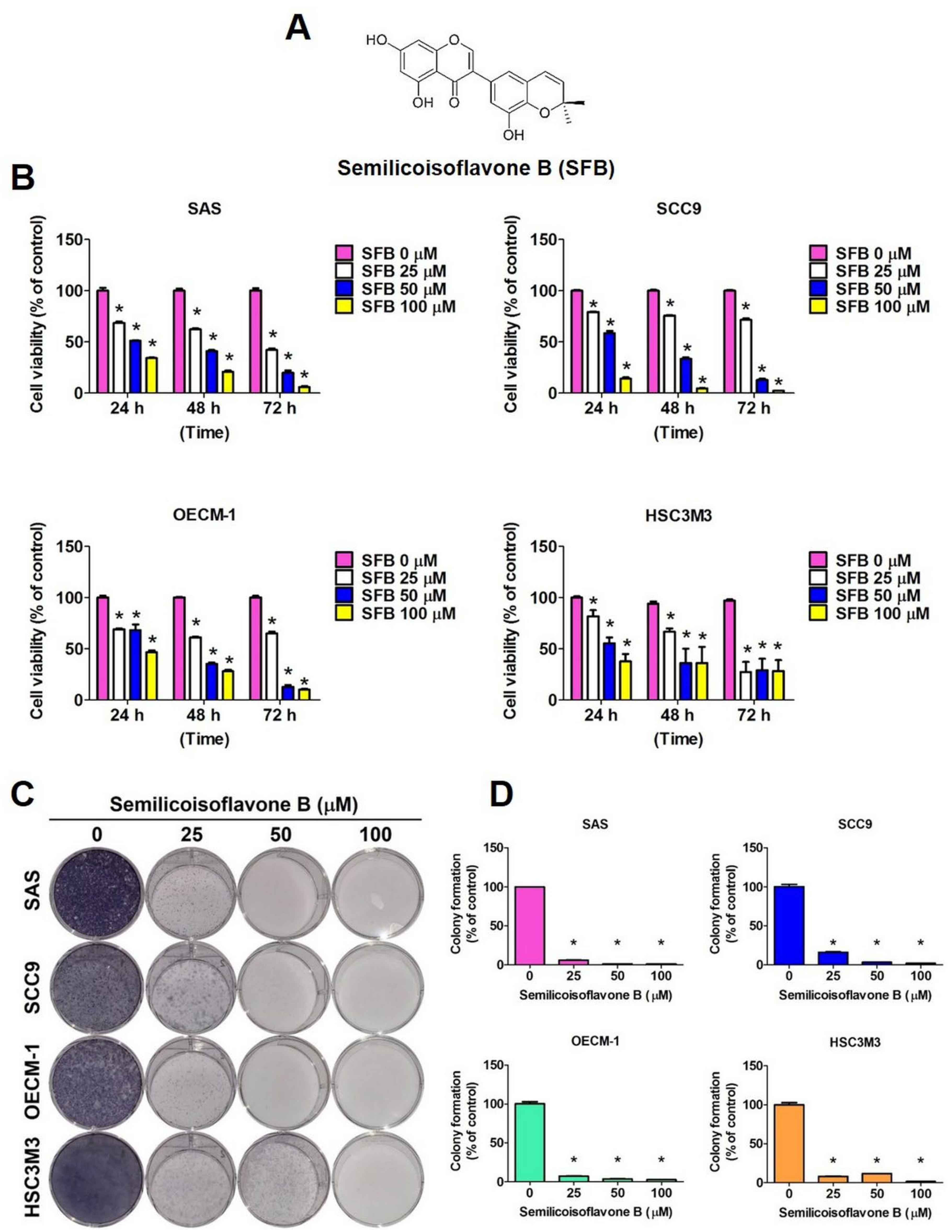
2.1. Cytotoxic Effect of SFB in Oral Cancer Cells
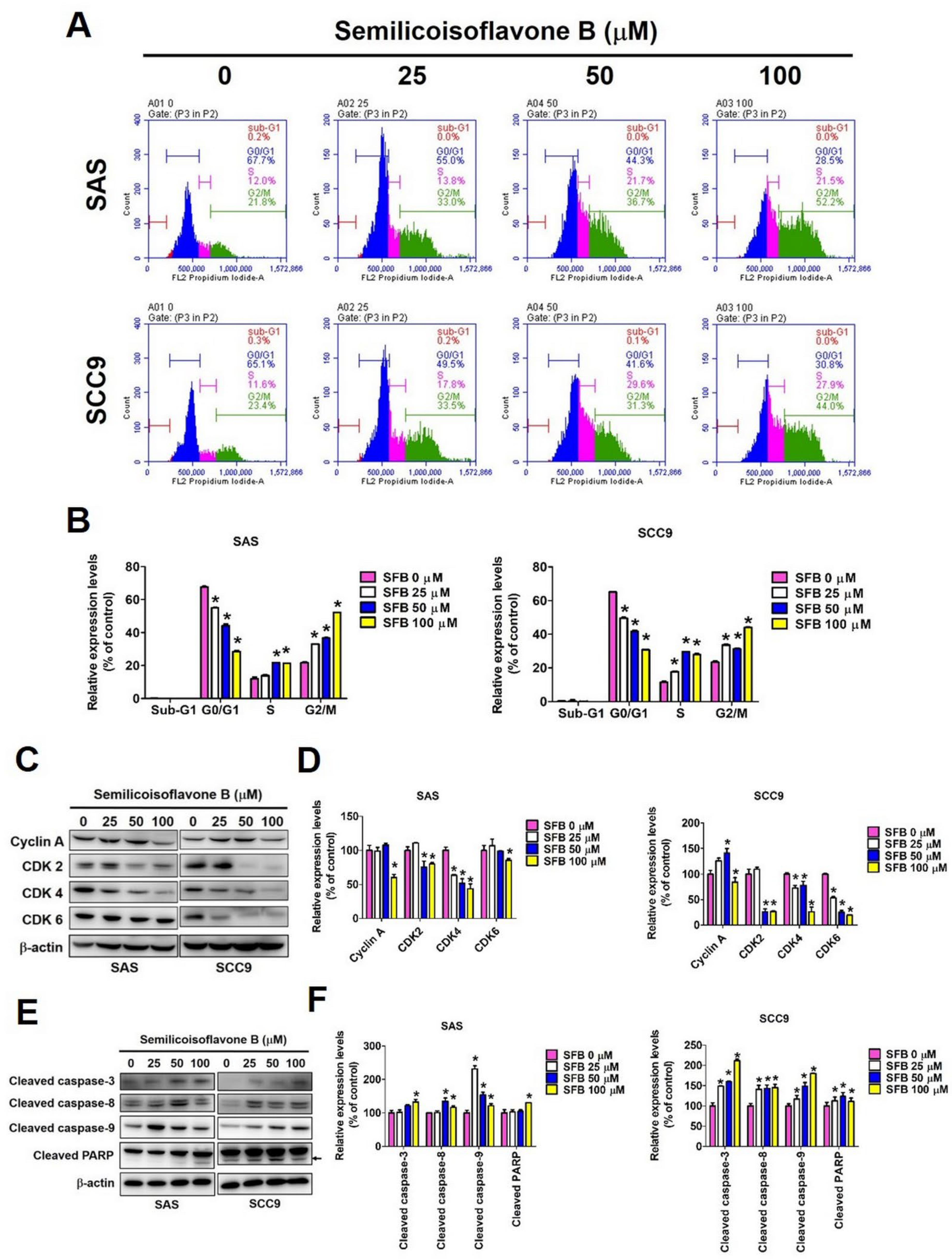
2.2. Effect of SFB on Cell Cycle Arrest and Apoptosis in Oral Cancer Cells
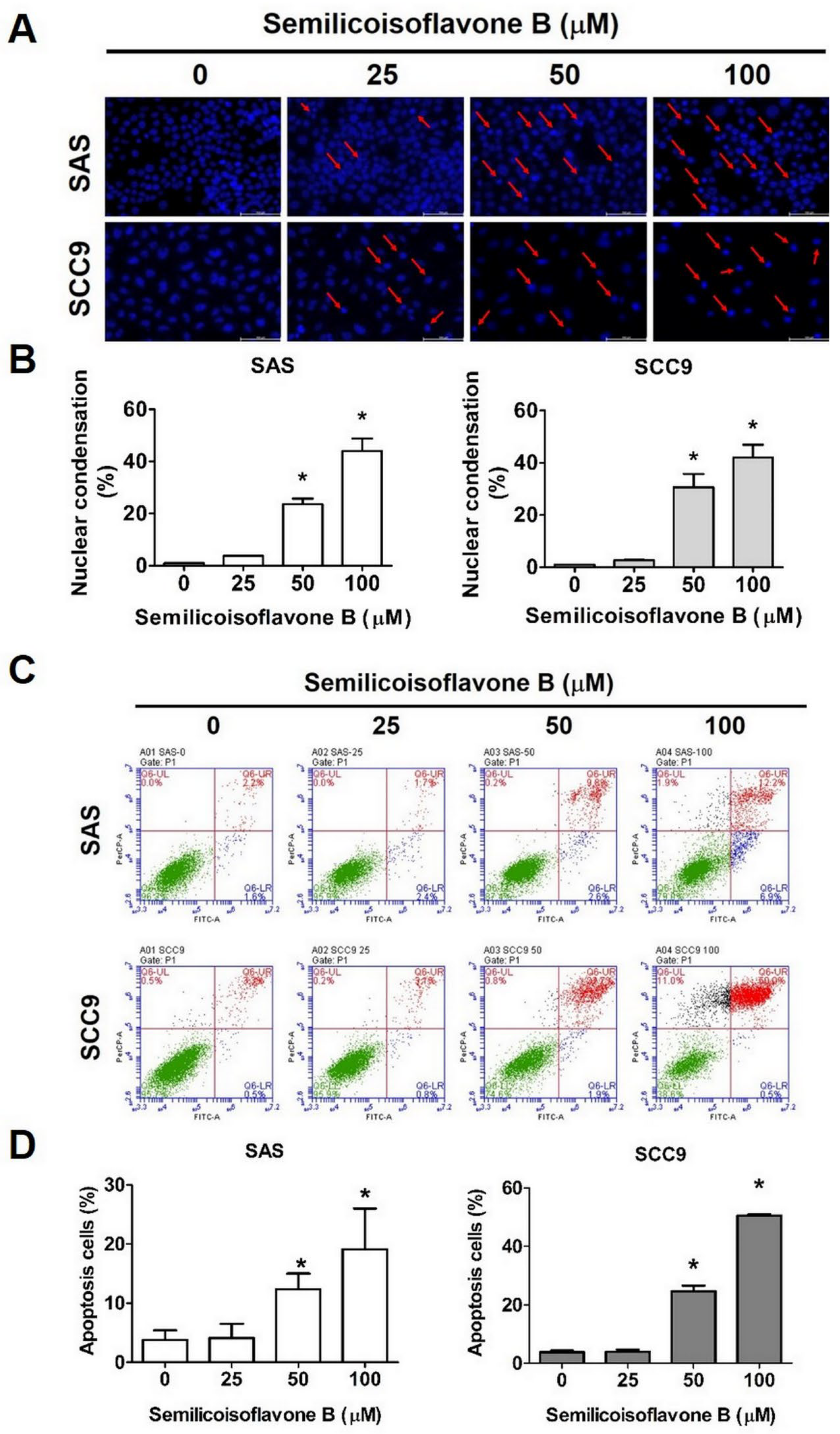
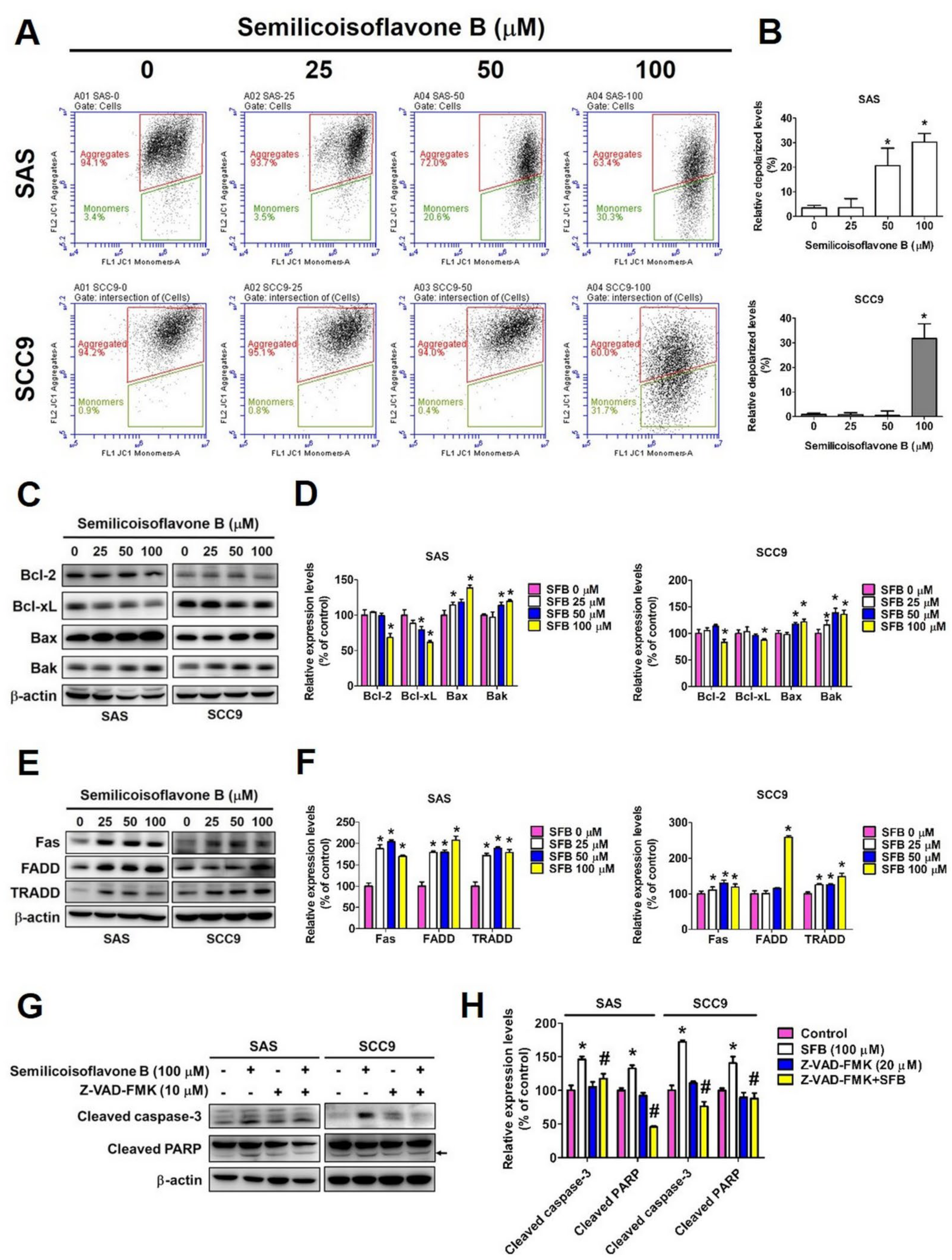
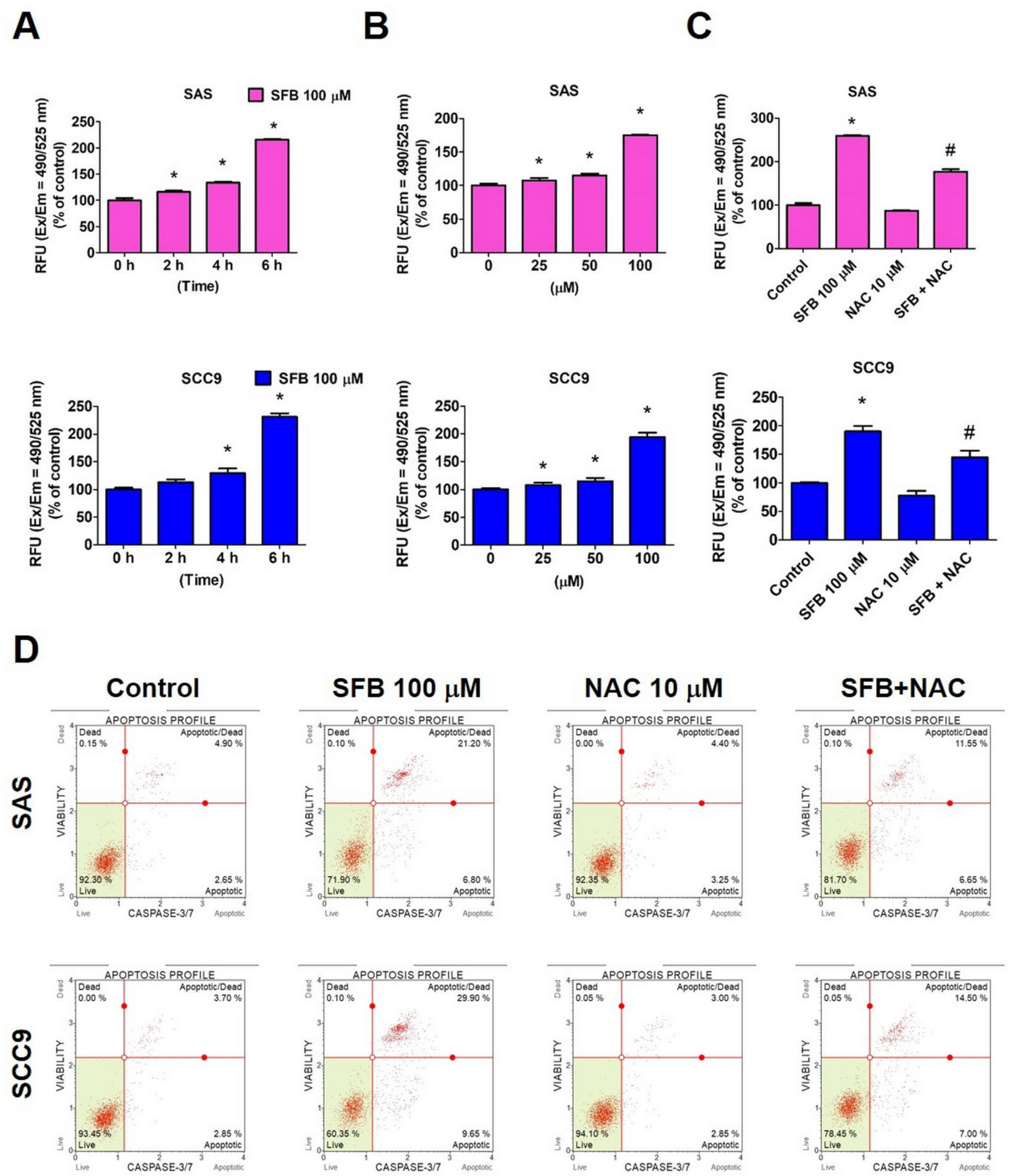
2.3. Effect of SFB on Free Radical Production and Apoptosis in Oral Cancer Cells
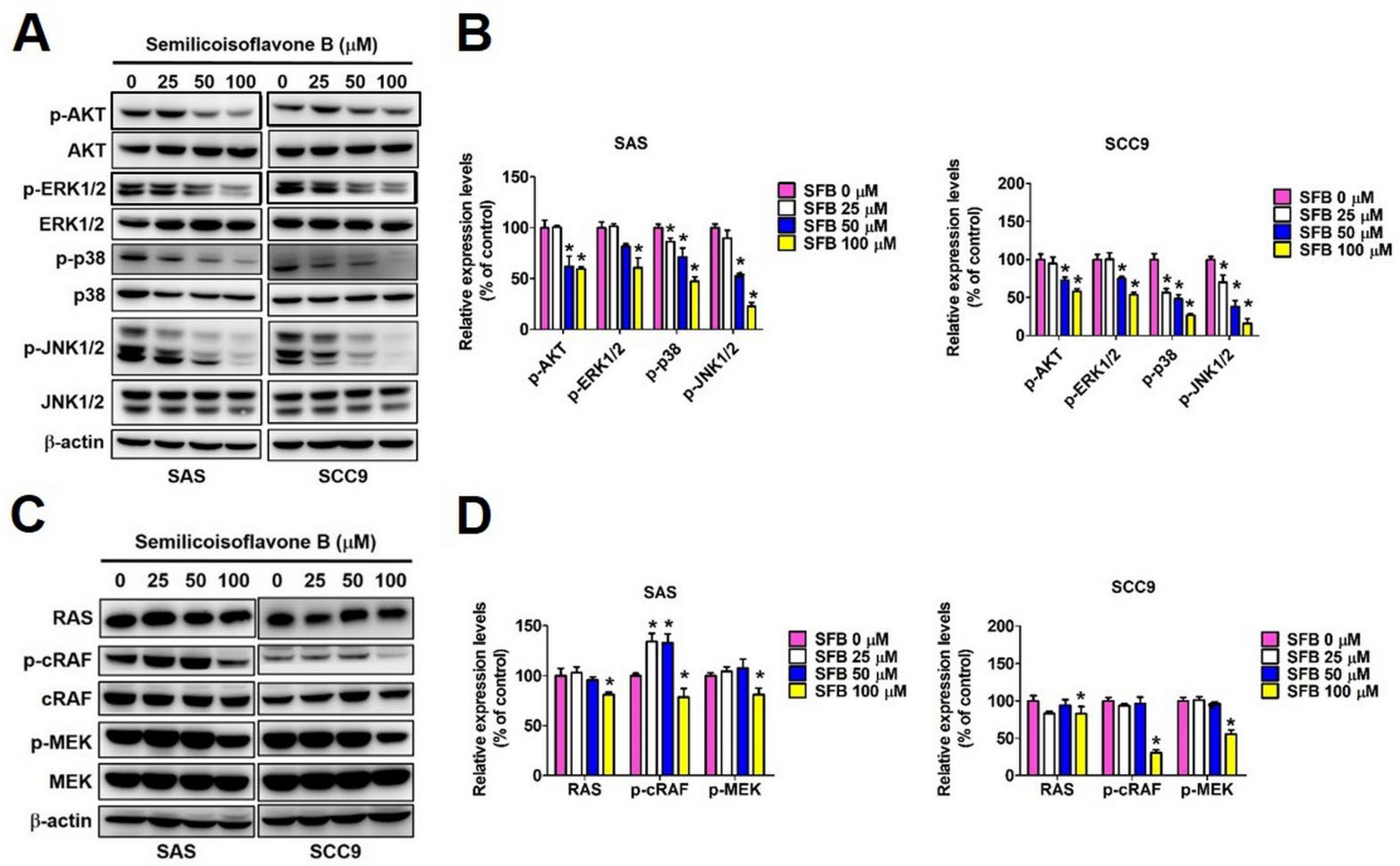
2.4. Effect of SFB on Cellular Signaling Pathways in Oral Cancer Cells
2.5. Effect of SFB on Specific Apoptosis-Related Protein Expression
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Chemical Treatments
4.3. Cell Viability (MTT Assay)
4.4. Colony Formation Assay
4.5. Cell Cycle Analysis
4.6. Western Blot Assay
4.7. DAPI Staining
4.8. Annexin V/PI Double Staining Assay
4.9. Mitochondrial Membrane Potential Measurement
4.10. Reactive Oxygen Species (ROS) Measurement
4.11. Caspase-3/7 Detection and Analysis
4.12. Gene Transfection
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alsaeedi, S.M.; Aggarwal, S. The Holistic Review on Occurrence, Biology, Diagnosis, and Treatment of Oral Squamous Cell Carcinoma. Cureus 2022, 14, e30226. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.; Venegas, B. Histological and molecular aspects of oral squamous cell carcinoma (Review). Oncol. Lett. 2014, 8, 7–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almangush, A.; Leivo, I.; Makitie, A.A. Biomarkers for Immunotherapy of Oral Squamous Cell Carcinoma: Current Status and Challenges. Front. Oncol. 2021, 11, 616629. [Google Scholar] [CrossRef] [PubMed]
- Rivera, C.; Oliveira, A.K.; Costa, R.A.P.; De Rossi, T.; Leme, A.F.P. Prognostic biomarkers in oral squamous cell carcinoma: A systematic review. Oral Oncol. 2017, 72, 38–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gluckman, J.L.; Pavelic, Z.P.; Welkoborsky, H.J.; Mann, W.; Stambrook, P.; Gleich, L.; Wilson, K.; Righi, P.; Portugal, L.G.; McDonald, J.; et al. Prognostic indicators for squamous cell carcinoma of the oral cavity: A clinicopathologic correlation. Laryngoscope 1997, 107, 1239–1244. [Google Scholar] [CrossRef]
- Ferreira, A.K.; Carvalho, S.H.; Granville-Garcia, A.F.; Sarmento, D.J.; Agripino, G.G.; Abreu, M.H.; Melo, M.C.; Caldas, A.D., Jr.; Godoy, G.P. Survival and prognostic factors in patients with oral squamous cell carcinoma. Med. Oral Patol. Oral Cir. Bucal 2021, 26, e387–e392. [Google Scholar] [CrossRef]
- Zittel, S.; Moratin, J.; Horn, D.; Metzger, K.; Ristow, O.; Engel, M.; Mrosek, J.; Freier, K.; Hoffmann, J.; Freudlsperger, C. Clinical outcome and prognostic factors in recurrent oral squamous cell carcinoma after primary surgical treatment: A retrospective study. Clin. Oral Investig. 2022, 26, 2055–2064. [Google Scholar] [CrossRef]
- Shetty, K.S.R.; Kurle, V.; Greeshma, P.; Ganga, V.B.; Murthy, S.P.; Thammaiah, S.K.; Prasad, P.K.; Chavan, P.; Halkud, R.; Krishnappa, R. Salvage Surgery in Recurrent Oral Squamous Cell Carcinoma. Front. Oral Health 2021, 2, 815606. [Google Scholar] [CrossRef]
- Strickler, J.H.; Hanks, B.A.; Khasraw, M. Tumor Mutational Burden as a Predictor of Immunotherapy Response: Is More Always Better? Clin. Cancer Res. 2021, 27, 1236–1241. [Google Scholar] [CrossRef]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in Cancer Treatment: From Preclinical Studies to Clinical Practice. Front. Pharmacol. 2019, 10, 1614. [Google Scholar] [CrossRef] [Green Version]
- George, B.P.; Chandran, R.; Abrahamse, H. Role of Phytochemicals in Cancer Chemoprevention: Insights. Antioxidants 2021, 10, 1455. [Google Scholar] [CrossRef]
- Rizeq, B.; Gupta, I.; Ilesanmi, J.; AlSafran, M.; Rahman, M.M.; Ouhtit, A. The Power of Phytochemicals Combination in Cancer Chemoprevention. J. Cancer 2020, 11, 4521–4533. [Google Scholar] [CrossRef]
- Leitzmann, C. Characteristics and Health Benefits of Phytochemicals. Forsch. Komplementmed. 2016, 23, 69–74. [Google Scholar] [CrossRef]
- Asl, M.N.; Hosseinzadeh, H. Review of pharmacological effects of Glycyrrhiza sp. and its bioactive compounds. Phytother. Res. 2008, 22, 709–724. [Google Scholar] [CrossRef]
- Mukhia, S.; Mandal, P.; Singh, D.K.; Singh, D. Comparison of pharmacological properties and phytochemical constituents of in vitro propagated and naturally occurring liverwort Lunularia cruciata. BMC Complement. Altern. Med. 2019, 19, 181. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.H.; Li, T.; Tong, Y.G.; Chen, X.J.; Chen, X.P.; Wang, Y.T.; Lu, J.J. A Systematic Review of the Anticancer Properties of Compounds Isolated from Licorice (Gancao). Planta Med. 2015, 81, 1670–1687. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Yang, L.; Hou, J.; Tian, S.; Liu, Y. Molecular mechanisms underlying the anticancer activities of licorice flavonoids. J. Ethnopharmacol. 2021, 267, 113635. [Google Scholar] [CrossRef]
- Zhang, X.R.; Wang, S.Y.; Sun, W.; Wei, C. Isoliquiritigenin inhibits proliferation and metastasis of MKN28 gastric cancer cells by suppressing the PI3K/AKT/mTOR signaling pathway. Mol. Med. Rep. 2018, 18, 3429–3436. [Google Scholar] [CrossRef] [Green Version]
- Kwon, S.J.; Park, S.Y.; Kwon, G.T.; Lee, K.W.; Kang, Y.H.; Choi, M.S.; Yun, J.W.; Jeon, J.H.; Jun, J.G.; Park, J.H. Licochalcone E present in licorice suppresses lung metastasis in the 4T1 mammary orthotopic cancer model. Cancer Prev. Res. 2013, 6, 603–613. [Google Scholar] [CrossRef] [Green Version]
- Si, L.; Yan, X.; Hao, W.; Ma, X.; Ren, H.; Ren, B.; Li, D.; Dong, Z.; Zheng, Q. Licochalcone D induces apoptosis and inhibits migration and invasion in human melanoma A375 cells. Oncol. Rep. 2018, 39, 2160–2170. [Google Scholar] [CrossRef] [Green Version]
- Batiha, G.E.-S.; Beshbishy, A.M.; El-Mleeh, A.; Abdel-Daim, M.M.; Devkota, H.P. Traditional Uses, Bioactive Chemical Constituents, and Pharmacological and Toxicological Activities of Glycyrrhiza glabra L. (Fabaceae). Biomolecules 2020, 10, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, M.Y.; Chun, Y.S.; Zhao, D.; Ryu, S.Y.; Yang, H.O. Glycyrrhiza uralensis and Semilicoisoflavone B Reduce Abeta Secretion by Increasing PPARgamma Expression and Inhibiting STAT3 Phosphorylation to Inhibit BACE1 Expression. Mol. Nutr. Food Res. 2018, 62, e1700633. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.S.; Kim, S.H.; Jung, S.H.; Kim, J.K.; Pan, C.H.; Lim, S.S. Aldose reductase inhibitory compounds from Glycyrrhiza uralensis. Biol. Pharm. Bull. 2010, 33, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gu, M.Y.; Chun, Y.S.; Yong, R.S.; Yang, H.O. Licoflavonol Reduces Abeta Secretion by Increasing BACE1 Phosphorylation to Facilitate BACE1 Degradation. Mol. Nutr. Food Res. 2019, 63, e1800474. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Kuang, Y.; Li, K.; Wang, S.; Song, W.; Qiao, X.; Sabir, G.; Ye, M. Screening for bioactive natural products from a 67-compound library of Glycyrrhiza inflata. Bioorg. Med. Chem. 2017, 25, 3706–3713. [Google Scholar] [CrossRef]
- Majtnerova, P.; Capek, J.; Petira, F.; Handl, J.; Rousar, T. Quantitative spectrofluorometric assay detecting nuclear condensation and fragmentation in intact cells. Sci. Rep. 2021, 11, 11921. [Google Scholar] [CrossRef]
- McIlwain, D.R.; Berger, T.; Mak, T.W. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008656. [Google Scholar] [CrossRef] [Green Version]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Hayashi, H.; Hosono, N.; Kondo, M.; Hiraoka, N.; Ikeshiro, Y. Phylogenetic relationship of Glycyrrhiza plants based on rbcL sequences. Biol. Pharm. Bull. 1998, 21, 782–783. [Google Scholar] [CrossRef] [Green Version]
- Cevik, D.; Kan, Y.; Kirmizibekmez, H. Mechanisms of action of cytotoxic phenolic compounds from Glycyrrhiza iconica roots. Phytomedicine 2019, 58, 152872. [Google Scholar] [CrossRef]
- Hasan, A.; Zhang, M.; Shang, Z.P.; Yi, Y.; Kuang, Y.; Yu, R.; Fan, J.J.; Huang, Y.X.; Nijat, D.; Qiao, X.; et al. Bioactive prenylated phenolic compounds from the aerial parts of Glycyrrhiza uralensis. Phytochemistry 2022, 201, 113284. [Google Scholar] [CrossRef]
- Xiao, Y.; Han, F.; Lee, I.S. Biotransformation of the Phenolic Constituents from Licorice and Cytotoxicity Evaluation of Their Metabolites. Int. J. Mol. Sci. 2021, 22, 109. [Google Scholar] [CrossRef]
- Shults, E.E.; Shakirov, M.M.; Pokrovsky, M.A.; Petrova, T.N.; Pokrovsky, A.G.; Gorovoy, P.G. Phenolic compounds from Glycyrrhiza pallidiflora Maxim. and their cytotoxic activity. Nat. Prod. Res. 2017, 31, 445–452. [Google Scholar] [CrossRef]
- Liu, X.; Xing, Y.; Li, M.; Zhang, Z.; Wang, J.; Ri, M.; Jin, C.; Xu, G.; Piao, L.; Jin, H.; et al. Licochalcone A inhibits proliferation and promotes apoptosis of colon cancer cell by targeting programmed cell death-ligand 1 via the NF-kappaB and Ras/Raf/MEK pathways. J. Ethnopharmacol. 2021, 273, 113989. [Google Scholar] [CrossRef]
- Pastorino, G.; Cornara, L.; Soares, S.; Rodrigues, F.; Oliveira, M. Liquorice (Glycyrrhiza glabra): A phytochemical and pharmacological review. Phytother. Res. 2018, 32, 2323–2339. [Google Scholar] [CrossRef]
- Farooqui, A.; Khan, F.; Khan, I.; Ansari, I.A. Glycyrrhizin induces reactive oxygen species-dependent apoptosis and cell cycle arrest at G(0)/G(1) in HPV18(+) human cervical cancer HeLa cell line. Biomed. Pharmacother. 2018, 97, 752–764. [Google Scholar] [CrossRef]
- Weidner, C.; Rousseau, M.; Micikas, R.J.; Fischer, C.; Plauth, A.; Wowro, S.J.; Siems, K.; Hetterling, G.; Kliem, M.; Schroeder, F.C.; et al. Amorfrutin C Induces Apoptosis and Inhibits Proliferation in Colon Cancer Cells through Targeting Mitochondria. J. Nat. Prod. 2016, 79, 2–12. [Google Scholar] [CrossRef]
- Guo, W.; Liu, B.; Yin, Y.; Kan, X.; Gong, Q.; Li, Y.; Cao, Y.; Wang, J.; Xu, D.; Ma, H.; et al. Licochalcone A Protects the Blood-Milk Barrier Integrity and Relieves the Inflammatory Response in LPS-Induced Mastitis. Front. Immunol. 2019, 10, 287. [Google Scholar] [CrossRef] [Green Version]
- Chang, G.H.; Yang, P.R.; Cheng, Y.C.; Hsu, K.H.; Wu, C.Y.; Yang, Y.H.; Lin, Y.S.; Hsu, C.M.; Tsai, M.S.; Tsai, Y.T.; et al. Nasal irrigation with licorice extract (Glycyrrhiza glabra) in treating nasal polyps by reducing fibroblast differentiation and extracellular matrix production in TGF-beta1-stimulated nasal polyp-derived fibroblasts by inhibiting the MAPK/ERK-1/2 pathway—An in vitro and in clinic study. BMC Complement. Med. Ther. 2022, 22, 313. [Google Scholar] [CrossRef]
- Wang, J.; Wei, B.; Thakur, K.; Wang, C.Y.; Li, K.X.; Wei, Z.J. Transcriptome Analysis Reveals the Anti-cancerous Mechanism of Licochalcone A on Human Hepatoma Cell HepG2. Front. Nutr. 2021, 8, 807574. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, Z.; Ma, X.; Yang, S.; Hu, X.; Tao, J.; Hou, Y.; Bai, G. Glycyrrhetinic acid binds to the conserved P-loop region and interferes with the interaction of RAS-effector proteins. Acta Pharm. Sin. B 2019, 9, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liang, X.; Zhou, X.; Wu, Z.; Yuan, L.; Wang, Y.; Li, Y. Selection of Reference Genes for qRT-PCR Analysis in Medicinal Plant Glycyrrhiza under Abiotic Stresses and Hormonal Treatments. Plants 2020, 9, 1441. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Nie, Y.; Hou, Y.; Ma, X.; Ding, G.; Gao, J.; Jiang, M.; Bai, G. Chemomics-Integrated Proteomics Analysis of Jie-Geng-Tang to Ameliorate Lipopolysaccharide-Induced Acute Lung Injury in Mice. Evid. Based Complement. Altern. Med. 2016, 2016, 7379146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, H.; Suri, P.; Gupta, J.C.; Talwar, G.P.; Dubey, S. Survivin: A unique target for tumor therapy. Cancer Cell Int. 2016, 16, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, Y.S.; Thakur, K.; Hussain, S.S.; Zhang, J.G.; Xiao, G.R.; Wei, Z.J. Licochalcone A from licorice root, an inhibitor of human hepatoma cell growth via induction of cell apoptosis and cell cycle arrest. Food Chem. Toxicol. 2018, 120, 407–417. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Wang, Y.; Xie, S.; Zhou, Y.; Ren, X.; Li, X.; Cai, Y. Liquiritigenin induces mitochondria-mediated apoptosis via cytochrome c release and caspases activation in HeLa Cells. Phytother. Res. 2011, 25, 277–283. [Google Scholar] [CrossRef]
- Kim, K.H.; Yoon, G.; Cho, J.J.; Cho, J.H.; Cho, Y.S.; Chae, J.I.; Shim, J.H. Licochalcone A induces apoptosis in malignant pleural mesothelioma through downregulation of Sp1 and subsequent activation of mitochondria-related apoptotic pathway. Int. J. Oncol. 2015, 46, 1385–1392. [Google Scholar] [CrossRef] [Green Version]
- Oh, H.N.; Seo, J.H.; Lee, M.H.; Kim, C.; Kim, E.; Yoon, G.; Cho, S.S.; Cho, Y.S.; Choi, H.W.; Shim, J.H.; et al. Licochalcone C induced apoptosis in human oral squamous cell carcinoma cells by regulation of the JAK2/STAT3 signaling pathway. J. Cell Biochem. 2018, 119, 10118–10130. [Google Scholar] [CrossRef]
- Cho, J.J.; Chae, J.I.; Yoon, G.; Kim, K.H.; Cho, J.H.; Cho, S.S.; Cho, Y.S.; Shim, J.H. Licochalcone A, a natural chalconoid isolated from Glycyrrhiza inflata root, induces apoptosis via Sp1 and Sp1 regulatory proteins in oral squamous cell carcinoma. Int. J. Oncol. 2014, 45, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.B.; Hsieh, M.J.; Mahalakshmi, B.; Chuang, Y.C.; Lin, C.C.; Lo, Y.S.; Ho, H.Y.; Lin, J.T. 7-Epitaxol Induces Apoptosis and Autophagy in Head and Neck Squamous Cell Carcinoma through Inhibition of the ERK Pathway. Cells 2021, 10, 2633. [Google Scholar] [CrossRef]
- Liu, Y.T.; Ho, H.Y.; Lin, C.C.; Chuang, Y.C.; Lo, Y.S.; Hsieh, M.J.; Chen, M.K. Platyphyllenone Induces Autophagy and Apoptosis by Modulating the AKT and JNK Mitogen-Activated Protein Kinase Pathways in Oral Cancer Cells. Int. J. Mol. Sci. 2021, 22, 4211. [Google Scholar] [CrossRef]
- Ho, H.Y.; Lin, C.C.; Chuang, Y.C.; Lo, Y.S.; Hsieh, M.J.; Chen, M.K. Apoptotic effects of dehydrocrenatidine via JNK and ERK pathway regulation in oral squamous cell carcinoma. Biomed. Pharmacother. 2021, 137, 111362. [Google Scholar] [CrossRef]
- Hsieh, M.C.; Lo, Y.S.; Chuang, Y.C.; Lin, C.C.; Ho, H.Y.; Hsieh, M.J.; Lin, J.T. Dehydrocrenatidine extracted from Picrasma quassioides induces the apoptosis of nasopharyngeal carcinoma cells through the JNK and ERK signaling pathways. Oncol. Rep. 2021, 46, 166. [Google Scholar] [CrossRef]
- Jin, W.; Lu, S.; Wang, X.; Shu, Y.; Shi, H. Raddeanin A suppresses lung cancer cell proliferation via induction of apoptosis and increased production of ROS. Cell. Mol. Biol. 2020, 66, 174–179. [Google Scholar] [CrossRef]
- Tunc, D.; Dere, E.; Karakas, D.; Cevatemre, B.; Yilmaz, V.T.; Ulukaya, E. Cytotoxic and apoptotic effects of the combination of palladium (II) 5,5-diethylbarbiturate complex with bis(2-pyridylmethyl)amine and curcumin on non small lung cancer cell lines. Bioorg. Med. Chem. 2017, 25, 1717–1723. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, M.-J.; Ho, H.-Y.; Lo, Y.-S.; Lin, C.-C.; Chuang, Y.-C.; Abomughaid, M.M.; Hsieh, M.-C.; Chen, M.-K. Semilicoisoflavone B Induces Apoptosis of Oral Cancer Cells by Inducing ROS Production and Downregulating MAPK and Ras/Raf/MEK Signaling. Int. J. Mol. Sci. 2023, 24, 4505. https://doi.org/10.3390/ijms24054505
Hsieh M-J, Ho H-Y, Lo Y-S, Lin C-C, Chuang Y-C, Abomughaid MM, Hsieh M-C, Chen M-K. Semilicoisoflavone B Induces Apoptosis of Oral Cancer Cells by Inducing ROS Production and Downregulating MAPK and Ras/Raf/MEK Signaling. International Journal of Molecular Sciences. 2023; 24(5):4505. https://doi.org/10.3390/ijms24054505
Chicago/Turabian StyleHsieh, Ming-Ju, Hsin-Yu Ho, Yu-Sheng Lo, Chia-Chieh Lin, Yi-Ching Chuang, Mosleh Mohammad Abomughaid, Ming-Chang Hsieh, and Mu-Kuan Chen. 2023. "Semilicoisoflavone B Induces Apoptosis of Oral Cancer Cells by Inducing ROS Production and Downregulating MAPK and Ras/Raf/MEK Signaling" International Journal of Molecular Sciences 24, no. 5: 4505. https://doi.org/10.3390/ijms24054505