
Hypothermic Oxygenated Machine Perfusion Promotes Mitophagy Flux against Hypoxia-Ischemic Injury in Rat DCD Liver

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. HOPE Alleviated DCD Liver Injury in Rats
2.2. HOPE Enhanced Mitochondrial Function and Attenuated Oxidative Stress in DCD Liver
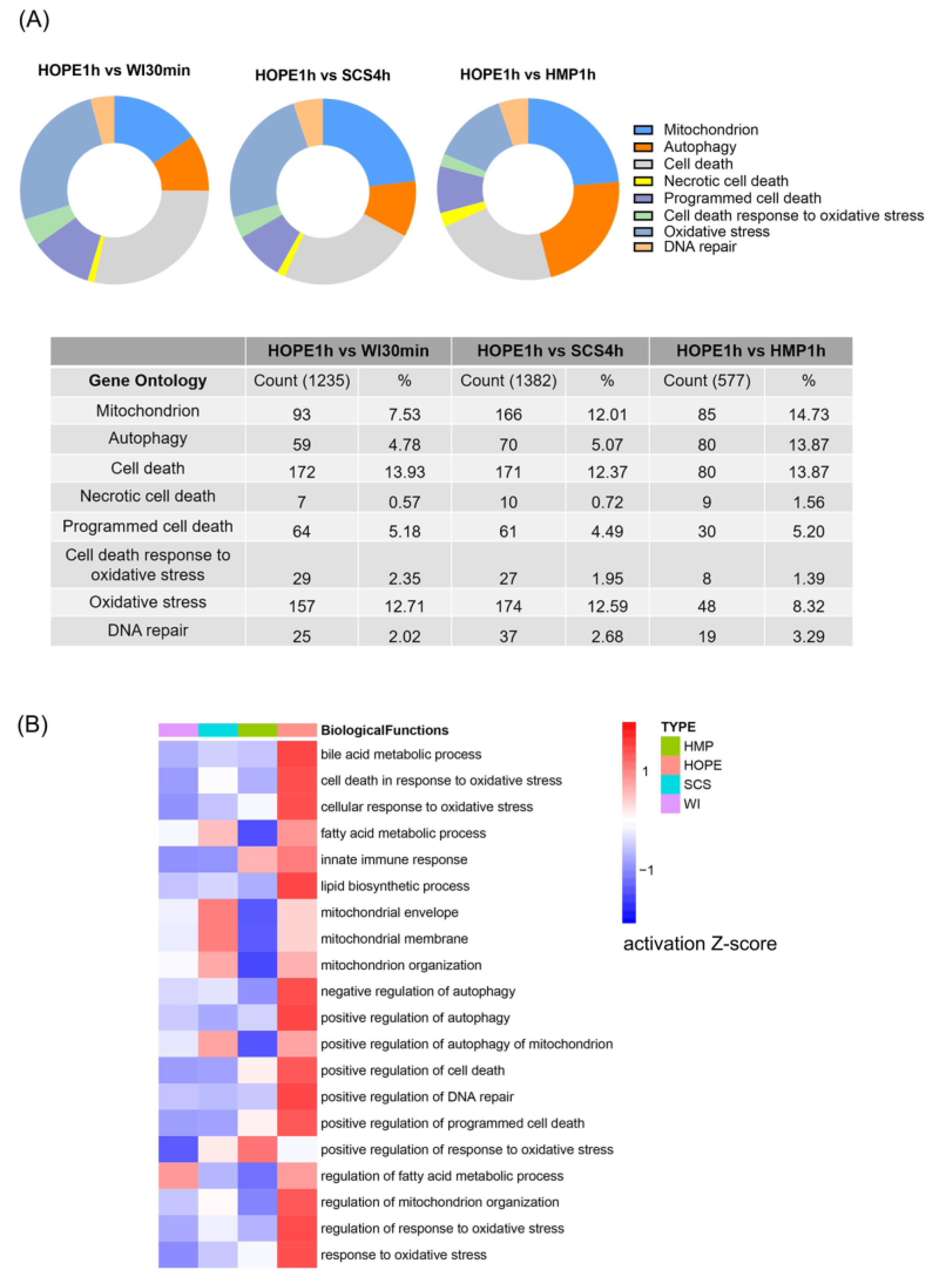
2.3. HOPE Regulated Transcriptomic Profiles in DCD Liver
2.4. HOPE Prevented Hepatocyte Injury and Reduced Apoptosis in Rat Liver
2.5. HOPE Restored Mitophagy and Promoted Mitophagy Flux after Hypoxia-Ischemic Injury in DCD Liver
2.6. Mitophagy Was Essential for the Protective Effect of HOPE
2.7. HOPE Ameliorated Ischemic Reperfusion Injury in DCD Liver
3. Discussion
4. Materials and Methods
4.1. Ethical Statement
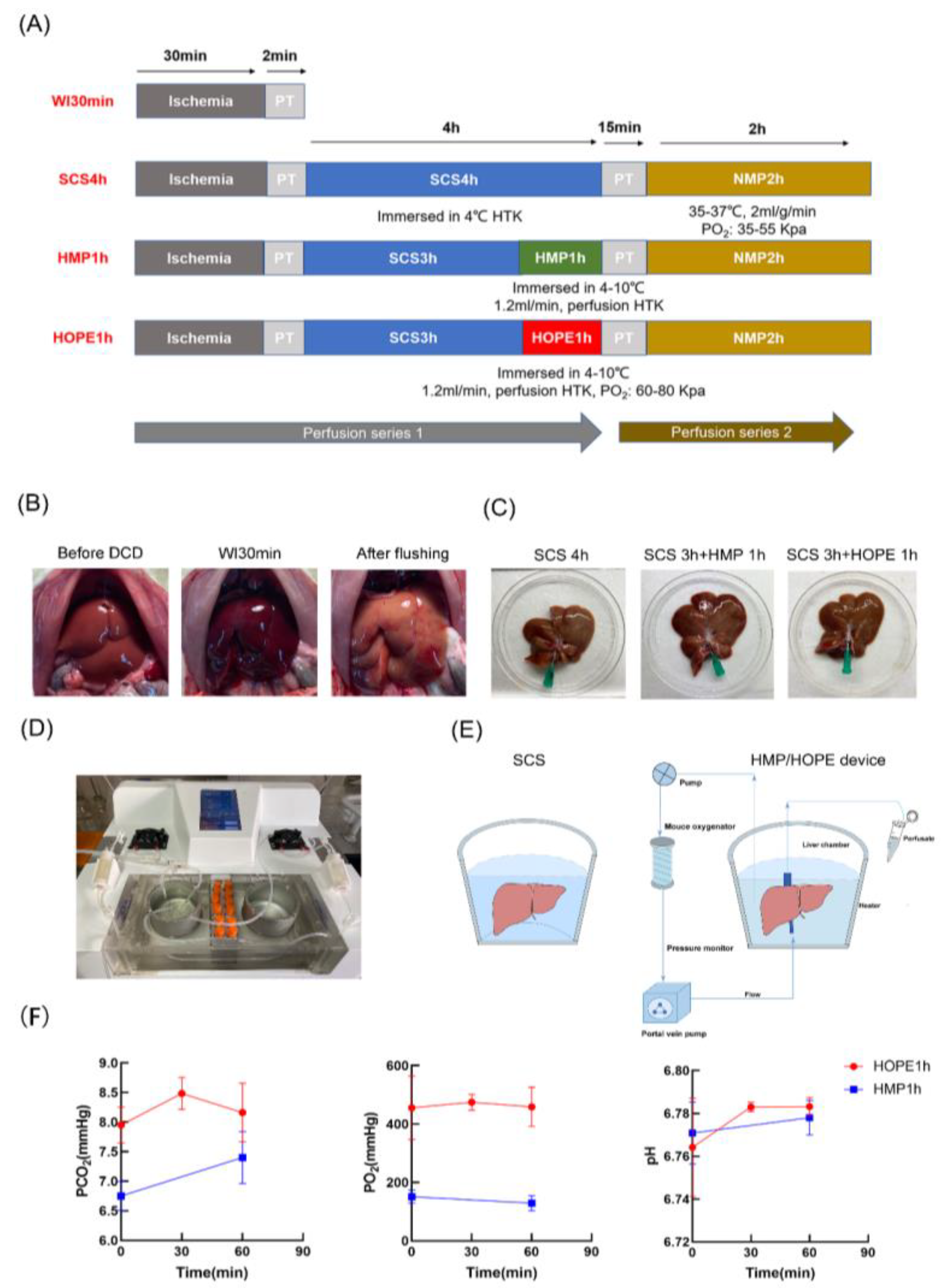
4.2. Experimental Design
- (1)
- Warm ischemic group: livers were exposed to warm ischemia for 30 min without any other intervention (WI 30 min).
- (2)
- Cold storage group: livers were exposed to warm ischemia for 30 min, followed by cold storage in Histidine-Tryptophan-Ketoglutarate (HTK) solution for 4 h (WI 30 min + SCS 4 h).
- (3)
- HMP group: livers were exposed to warm ischemia for 30 min, followed by cold storage in HTK solution for 3 h, as well as 1 h of hypothermic machine perfusion (WI 30 min + SCS 3 h + HMP 1 h).
- (4)
- HOPE group: livers were exposed to warm ischemia for 30 min, followed by cold storage in the HTK solution for 3 h and 1 h of hypothermic oxygenated machine perfusion (WI 30 min + SCS 3 h + HOPE 1 h).
4.3. Liver Procurement
4.4. Cold Storage and MP Platform
4.5. Endpoints
4.5.1. Perfusate Measurements
4.5.2. Liver Samples
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Czigany, Z.; Lurje, I.; Tolba, R.H.; Neumann, U.P.; Tacke, F.; Lurje, G. Machine perfusion for liver transplantation in the era of marginal organs-New kids on the block. Liver Int. 2019, 39, 228–249. [Google Scholar] [CrossRef]
- Williams, W.W.; Markmann, J.F. Warming Up to Cold Perfusion. N. Engl. J. Med. 2021, 384, 1458–1459. [Google Scholar] [CrossRef]
- Ceresa, C.D.L.; Nasralla, D.; Pollok, J.M.; Friend, P.J. Machine perfusion of the liver: Applications in transplantation and beyond. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 199–209. [Google Scholar] [CrossRef]
- Tullius, S.G.; Rabb, H. Improving the Supply and Quality of Deceased-Donor Organs for Transplantation. N. Engl. J. Med. 2018, 378, 1920–1929. [Google Scholar] [CrossRef]
- Jochmans, I.; Brat, A.; Davies, L.; Hofker, H.S.; van de Leemkolk, F.E.M.; Leuvenink, H.G.D.; Knight, S.R.; Pirenne, J.; Ploeg, R.J.; Abramowicz, D.; et al. Oxygenated versus standard cold perfusion preservation in kidney transplantation (COMPARE): A randomised, double-blind, paired, phase 3 trial. Lancet 2020, 396, 1653–1662. [Google Scholar] [CrossRef]
- Van Rijn, R.; Schurink, I.J.; de Vries, Y.; van den Berg, A.P.; Cortes Cerisuelo, M.; Darwish Murad, S.; Erdmann, J.I.; Gilbo, N.; de Haas, R.J.; Heaton, N.; et al. Hypothermic Machine Perfusion in Liver Transplantation—A Randomized Trial. N. Engl. J. Med. 2021, 384, 1391–1401. [Google Scholar] [CrossRef]
- Xu, J.; Buchwald, J.E.; Martins, P.N. Review of Current Machine Perfusion Therapeutics for Organ Preservation. Transplantation 2020, 104, 1792–1803. [Google Scholar] [CrossRef]
- Kim, J.; Zimmerman, M.A.; Shin, W.Y.; Boettcher, B.T.; Lee, J.S.; Park, J.I.; Ali, M.; Yang, M.; Mishra, J.; Hagen, C.E.; et al. Effects of Subnormothermic Regulated Hepatic Reperfusion on Mitochondrial and Transcriptomic Profiles in a Porcine Model. Ann. Surg. 2022, 277, e366–e375. [Google Scholar] [CrossRef]
- Lascaris, B.; de Meijer, V.E.; Porte, R.J. Normothermic liver machine perfusion as a dynamic platform for regenerative purposes: What does the future have in store for us? J. Hepatol. 2022, 77, 825–836. [Google Scholar] [CrossRef]
- Jing, L.; Yao, L.; Zhao, M.; Peng, L.P.; Liu, M. Organ preservation: From the past to the future. Acta Pharmacol. Sin. 2018, 39, 845–857. [Google Scholar] [CrossRef]
- Schlegel, A.; Muller, X.; Dutkowski, P. Machine perfusion strategies in liver transplantation. Hepatobiliary Surg. Nutr. 2019, 8, 490–501. [Google Scholar] [CrossRef]
- De Goeij, F.H.C.; Schlegel, A.; Muiesan, P.; Guarrera, J.V.; Dutkowski, P. Hypothermic Oxygenated Machine Perfusion Protects From Cholangiopathy in Donation after Circulatory Death Liver Transplantation. Hepatology 2021, 74, 3525–3528. [Google Scholar] [CrossRef]
- Bonaccorsi-Riani, E.; Bruggenwirth, I.M.A.; Buchwald, J.E.; Iesari, S.; Martins, P.N. Machine Perfusion: Cold versus Warm, versus Neither. Update on Clinical Trials. Semin. Liver Dis. 2020, 40, 264–281. [Google Scholar] [CrossRef]
- Muth, V.; Gassner, J.; Moosburner, S.; Lurje, G.; Michelotto, J.; Strobl, F.; Knaub, K.; Engelmann, C.; Tacke, F.; Selzner, M.; et al. Ex vivo liver machine perfusion: Comprehensive review of common animal models. Tissue Eng. Part B Rev. 2023, 29, 10–27. [Google Scholar] [CrossRef]
- Wyss, R.K.; Carmona, N.M.; Arnold, M.; Segiser, A.; Mueller, M.; Dutkowski, P.; Carrel, T.P.; Longnus, S.L. Hypothermic, oxygenated perfusion (HOPE) provides cardioprotection via succinate oxidation prior to normothermic perfusion in a rat model of donation after circulatory death (DCD). Am. J. Transplant. 2021, 21, 1003–1011. [Google Scholar] [CrossRef]
- Ishii, D.; Matsuno, N.; Gochi, M.; Iwata, H.; Shonaka, T.; Nishikawa, Y.; Obara, H.; Yokoo, H.; Furukawa, H. Beneficial effects of end-ischemic oxygenated machine perfusion preservation for split-liver transplantation in recovering graft function and reducing ischemia-reperfusion injury. Sci. Rep. 2021, 11, 22608. [Google Scholar] [CrossRef]
- Schlegel, A.; Muller, X.; Dutkowski, P. Hypothermic Machine Preservation of the Liver: State of the Art. Curr. Transplant. Rep. 2018, 5, 93–102. [Google Scholar] [CrossRef]
- Schlegel, A.; Porte, R.; Dutkowski, P. Protective mechanisms and current clinical evidence of hypothermic oxygenated machine perfusion (HOPE) in preventing post-transplant cholangiopathy J. Hepatol. 2022, 76, 1330–1347. [Google Scholar] [CrossRef]
- Clarke, G.; Mergental, H.; Hann, A.; Perera, M.; Afford, S.C.; Mirza, D.F. How Machine Perfusion Ameliorates Hepatic Ischaemia Reperfusion Injury. Int. J. Mol. Sci. 2021, 22, 7523. [Google Scholar] [CrossRef]
- Zhou, J.; Li, X.Y.; Liu, Y.J.; Feng, J.; Wu, Y.; Shen, H.M.; Lu, G.D. Full-coverage regulations of autophagy by ROS: From induction to maturation. Autophagy 2022, 18, 1240–1255. [Google Scholar] [CrossRef]
- Zeng, X.; Wang, S.; Li, S.; Yang, Y.; Fang, Z.; Huang, H.; Wang, Y.; Fan, X.; Ye, Q. Hypothermic oxygenated machine perfusion alleviates liver injury in donation after circulatory death through activating autophagy in mice. Artif. Organs 2019, 43, E320–E332. [Google Scholar] [CrossRef]
- Parzych, K.R.; Klionsky, D.J. An overview of autophagy: Morphology, mechanism, and regulation. Antioxid. Redox Signal. 2014, 20, 460–473. [Google Scholar] [CrossRef]
- Lamark, T.; Johansen, T. Mechanisms of Selective Autophagy. Annu. Rev. Cell Dev. Biol. 2021, 37, 143–169. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, Q.; Wang, X.; Chen, X.; Chen, Y.; Du, J.; Chen, L. The Role of Mitochondria in Liver Ischemia-Reperfusion Injury: From Aspects of Mitochondrial Oxidative Stress, Mitochondrial Fission, Mitochondrial Membrane Permeable Transport Pore Formation, Mitophagy, and Mitochondria-Related Protective Measures. Oxidative Med. Cell. Longev. 2021, 2021, 6670579. [Google Scholar] [CrossRef]
- Goikoetxea-Usandizaga, N.; Serrano-Macia, M.; Delgado, T.C.; Simon, J.; Fernandez Ramos, D.; Barriales, D.; Cornide, M.E.; Jimenez, M.; Perez-Redondo, M.; Lachiondo-Ortega, S.; et al. Mitochondrial bioenergetics boost macrophage activation, promoting liver regeneration in metabolically compromised animals. Hepatology 2022, 75, 550–566. [Google Scholar] [CrossRef]
- Onishi, M.; Yamano, K.; Sato, M.; Matsuda, N.; Okamoto, K. Molecular mechanisms and physiological functions of mitophagy. EMBO J. 2021, 40, e104705. [Google Scholar] [CrossRef]
- Youle, R.J.; Narendra, D.P. Mechanisms of mitophagy. Nat. Rev. Mol. Cell Biol. 2011, 12, 9–14. [Google Scholar] [CrossRef]
- Harper, J.W.; Ordureau, A.; Heo, J.M. Building and decoding ubiquitin chains for mitophagy. Nat. Rev. Mol. Cell Biol. 2018, 19, 93–108. [Google Scholar] [CrossRef]
- Santidrian, A.F.; Matsuno-Yagi, A.; Ritland, M.; Seo, B.B.; LeBoeuf, S.E.; Gay, L.J.; Yagi, T.; Habermann, B.F. Felding-Habermann, Mitochondrial complex I activity and NAD+/NADH balance regulate breast cancer progression. J. Clin. Investig. 2013, 123, 1068–1081. [Google Scholar] [CrossRef]
- Muller, X.; Schlegel, A.; Kron, P.; Eshmuminov, D.; Wurdinger, M.; Meierhofer, D.; Clavien, P.A.; Dutkowski, P. Novel Real-time Prediction of Liver Graft Function During Hypothermic Oxygenated Machine Perfusion Before Liver Transplantation. Ann. Surg. 2019, 270, 783–790. [Google Scholar] [CrossRef]
- Schlegel, A.; Muller, X.; Mueller, M.; Stepanova, A.; Kron, P.; de Rougemont, O.; Muiesan, P.; Clavien, P.A.; Galkin, A.; Meierhofer, D.; et al. Hypothermic oxygenated perfusion protects from mitochondrial injury before liver transplantation. EBioMedicine 2020, 60, 103014. [Google Scholar] [CrossRef]
- Kron, P.; Schlegel, A.; de Rougemont, O.; Oberkofler, C.E.; Clavien, P.A.; Dutkowski, P. Short, Cool, and Well Oxygenated—HOPE for Kidney Transplantation in a Rodent Model. Ann. Surg. 2016, 264, 815–822. [Google Scholar] [CrossRef]
- Schlegel, A.; de Rougemont, O.; Graf, R.; Clavien, P.A.; Dutkowski, P. Protective mechanisms of end-ischemic cold machine perfusion in DCD liver grafts. J. Hepatol. 2013, 58, 278–286. [Google Scholar] [CrossRef]
- Schlegel, A.; Graf, R.; Clavien, P.A.; Dutkowski, P. Hypothermic oxygenated perfusion (HOPE) protects from biliary injury in a rodent model of DCD liver transplantation. J. Hepatol. 2013, 59, 984–991. [Google Scholar] [CrossRef]
- Sha, Z.; Yang, Y.; Liu, R.; Bao, H.; Song, S.; Dong, J.; Guo, M.; Zhao, Y.; Liu, H.; Ding, G. Hepatic Ischemia-reperfusion Injury in Mice was Alleviated by Rac1 Inhibition—More Than Just ROS-inhibition. J. Clin. Transl. Hepatol. 2022, 10, 42–52. [Google Scholar] [CrossRef]
- Yuhai, G.U.; Zhen, Z. Significance of the changes occurring in the levels of interleukins, SOD and MDA in rat pulmonary tissue following exposure to different altitudes and exposure times. Exp. Ther. Med. 2015, 10, 915–920. [Google Scholar] [CrossRef]
- Hendriks, K.D.W.; Bruggenwirth, I.M.A.; Maassen, H.; Gerding, A.; Bakker, B.; Porte, R.J.; Henning, R.H.; Leuvenink, H.G.D. Renal temperature reduction progressively favors mitochondrial ROS production over respiration in hypothermic kidney preservation. J. Transl. Med. 2019, 17, 265. [Google Scholar] [CrossRef]
- Hoyer, D.P.; Gallinat, A.; Swoboda, S.; Wohlschlaeger, J.; Rauen, U.; Paul, A.; Minor, T. Influence of oxygen concentration during hypothermic machine perfusion on porcine kidneys from donation after circulatory death. Transplantation 2014, 98, 944–950. [Google Scholar] [CrossRef]
- Ma, X.; McKeen, T.; Zhang, J.; Ding, W.X. Role and Mechanisms of Mitophagy in Liver Diseases. Cells 2020, 9, 837. [Google Scholar] [CrossRef]
- Pankiv, S.; Clausen, T.H.; Lamark, T.; Brech, A.; Bruun, J.A.; Outzen, H.; Øvervatn, A.; Bjørkøy, G.; Johansen, T. p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J. Biol. Chem. 2007, 282, 24131–24145. [Google Scholar] [CrossRef]
- Liu, W.J.; Ye, L.; Huang, W.F.; Guo, L.J.; Xu, Z.G.; Wu, H.L.; Yang, C.; Liu, H.F. p62 links the autophagy pathway and the ubiqutin-proteasome system upon ubiquitinated protein degradation. Cell. Mol. Biol. Lett. 2016, 21, 29. [Google Scholar] [CrossRef]
- He, N.; Jia, J.J.; Xie, H.Y.; Li, J.H.; He, Y.; Yin, S.Y.; Liang, R.P.; Jiang, L.; Liu, J.F.; Xu, K.D.; et al. Partial Inhibition of HO-1 Attenuates HMP-Induced Hepatic Regeneration against Liver Injury in Rats. Oxidative Med. Cell. Longev. 2018, 2018, 9108483. [Google Scholar] [CrossRef]
- Yang, L.; Cao, H.; Sun, D.; Lin, L.; Zheng, W.P.; Shen, Z.Y.; Song, H.L. Normothermic Machine Perfusion Combined with Bone Marrow Mesenchymal Stem Cells Improves the Oxidative Stress Response and Mitochondrial Function in Rat Donation After Circulatory Death Livers. Stem Cells Dev. 2020, 29, 835–852. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, J.; Hu, Y.; Qiao, Y.; Li, H.; Huang, J.; Xu, K.; Jiang, L.; Wu, H.; Hu, X.; Jia, J.; et al. Hypothermic Oxygenated Machine Perfusion Promotes Mitophagy Flux against Hypoxia-Ischemic Injury in Rat DCD Liver. Int. J. Mol. Sci. 2023, 24, 5403. https://doi.org/10.3390/ijms24065403
Luo J, Hu Y, Qiao Y, Li H, Huang J, Xu K, Jiang L, Wu H, Hu X, Jia J, et al. Hypothermic Oxygenated Machine Perfusion Promotes Mitophagy Flux against Hypoxia-Ischemic Injury in Rat DCD Liver. International Journal of Molecular Sciences. 2023; 24(6):5403. https://doi.org/10.3390/ijms24065403
Chicago/Turabian StyleLuo, Jia, Yiqing Hu, Yinbiao Qiao, Haoyu Li, Jiacheng Huang, Kangdi Xu, Li Jiang, Hao Wu, Xiaoyi Hu, Junjun Jia, and et al. 2023. "Hypothermic Oxygenated Machine Perfusion Promotes Mitophagy Flux against Hypoxia-Ischemic Injury in Rat DCD Liver" International Journal of Molecular Sciences 24, no. 6: 5403. https://doi.org/10.3390/ijms24065403