
Glycosylation Analysis of Urinary Peptidome Highlights IGF2 Glycopeptides in Association with CKD
 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Results
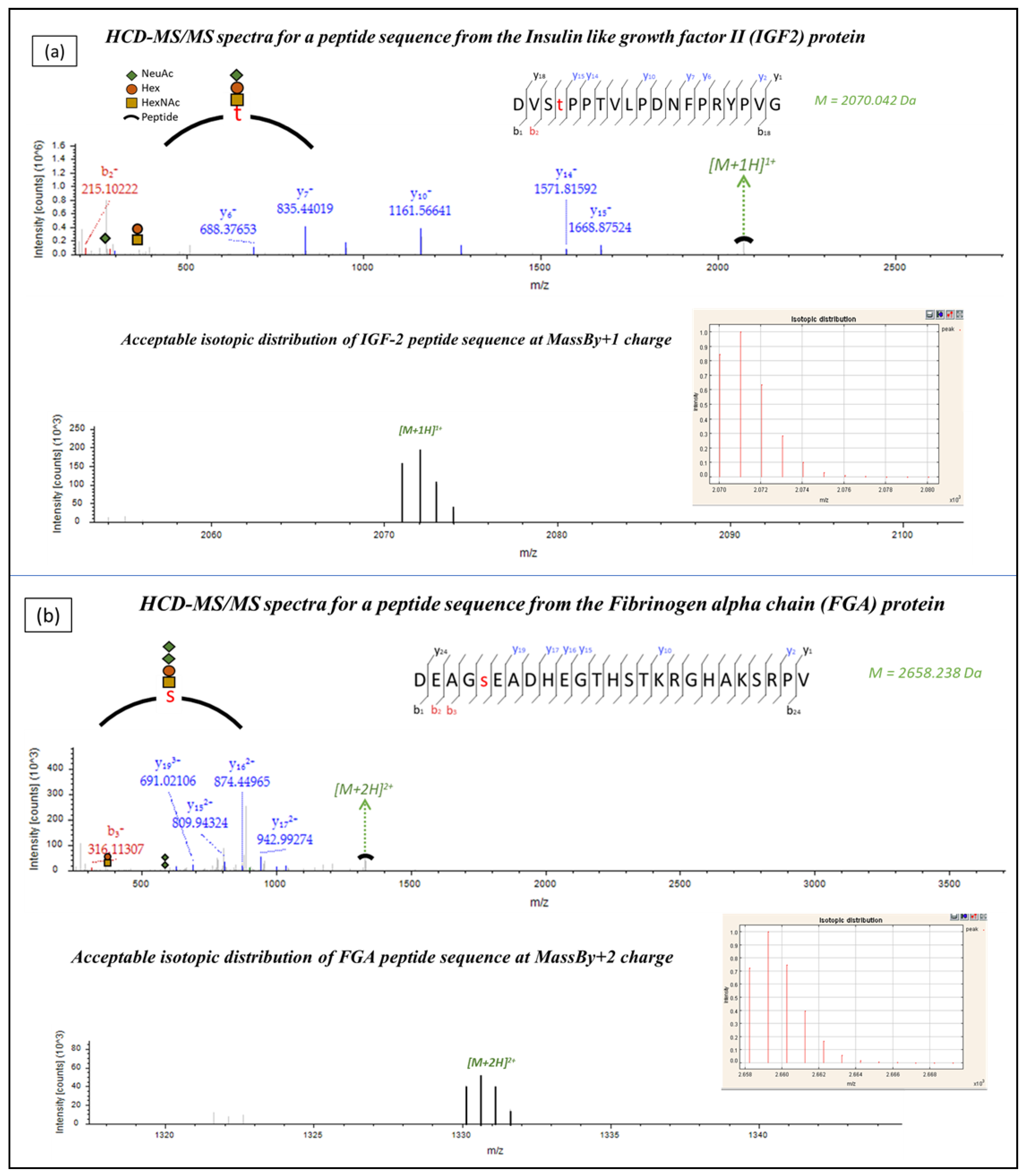
2.1. Identification of Urinary Glycopeptides
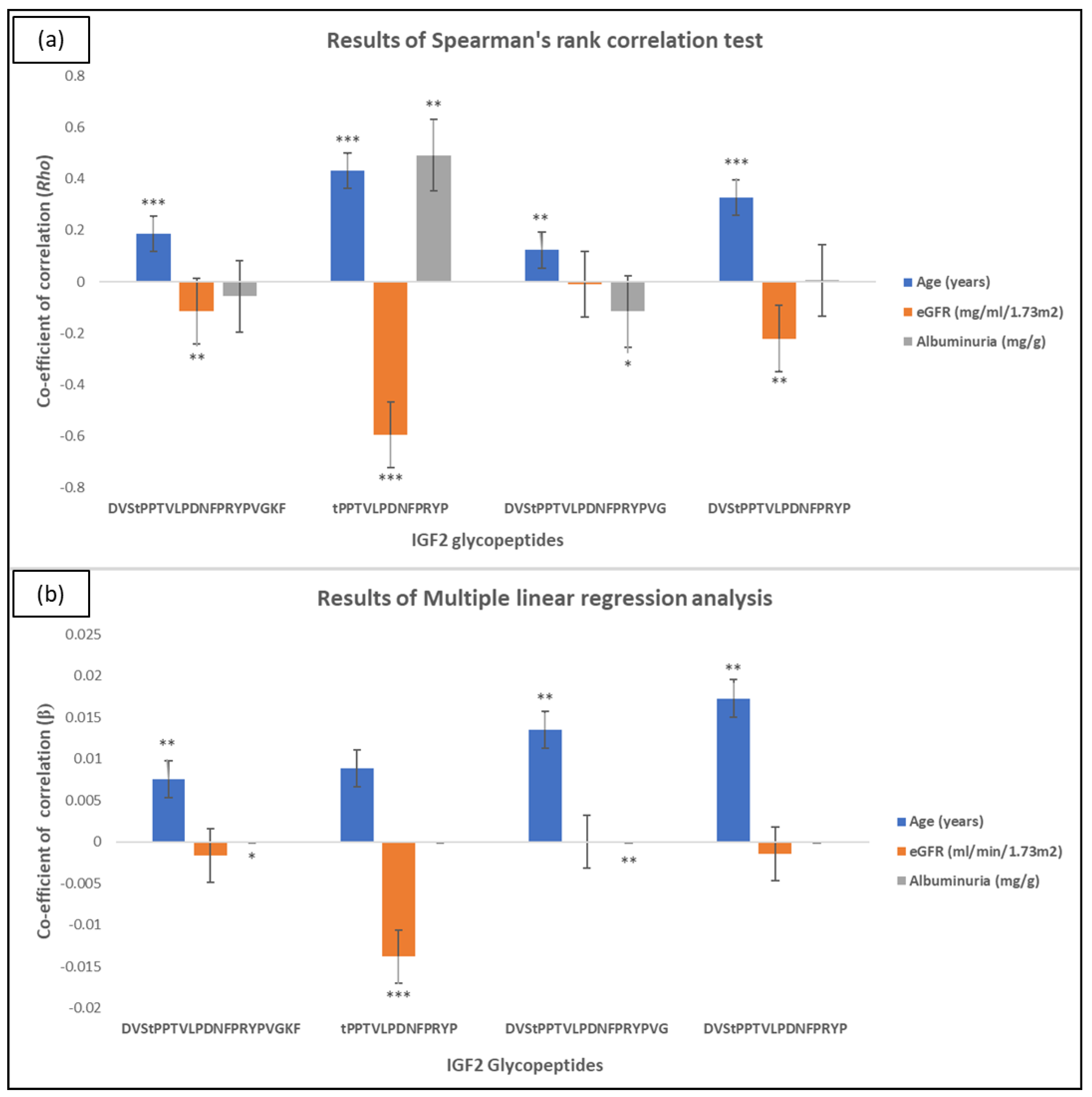
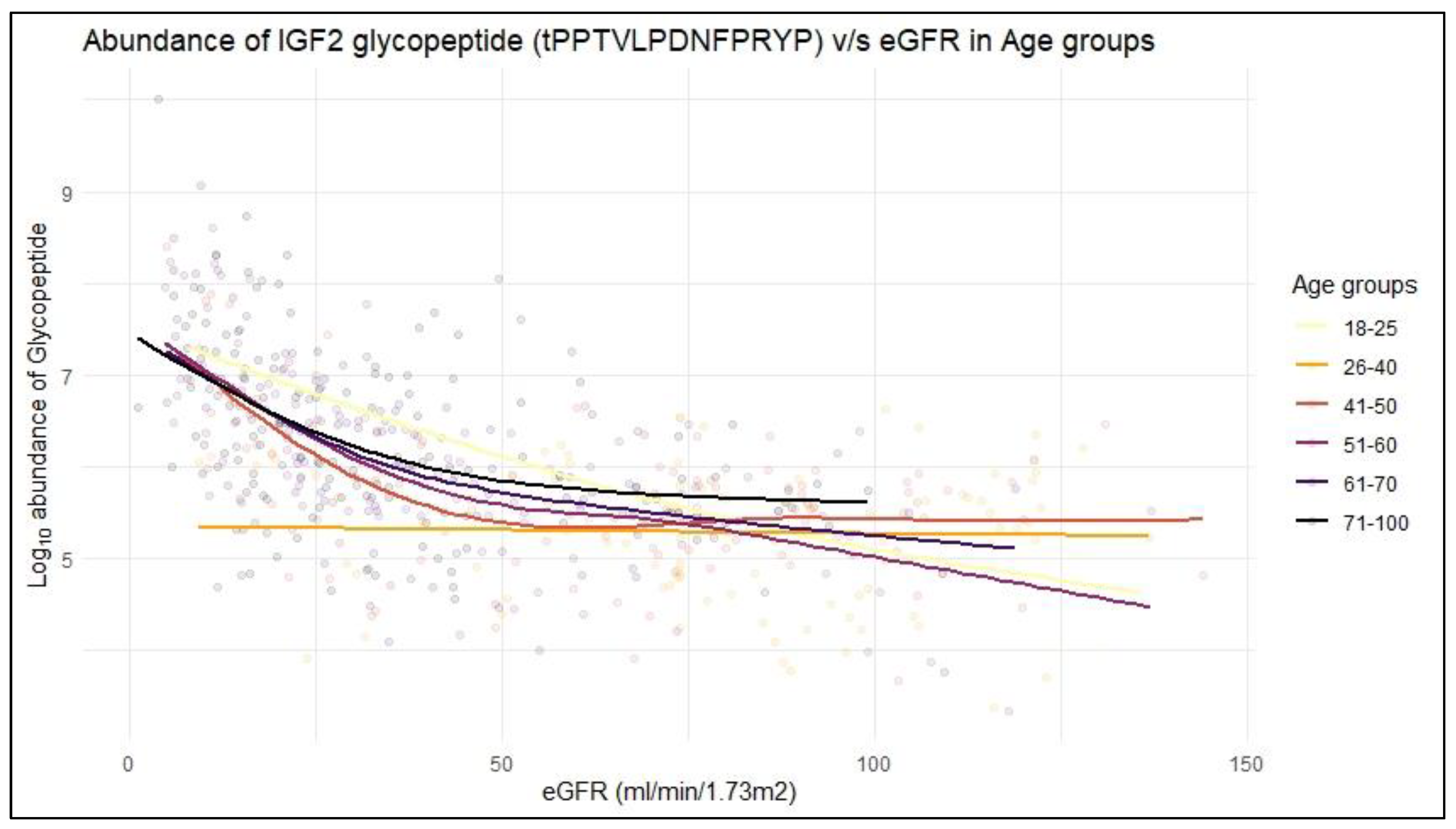
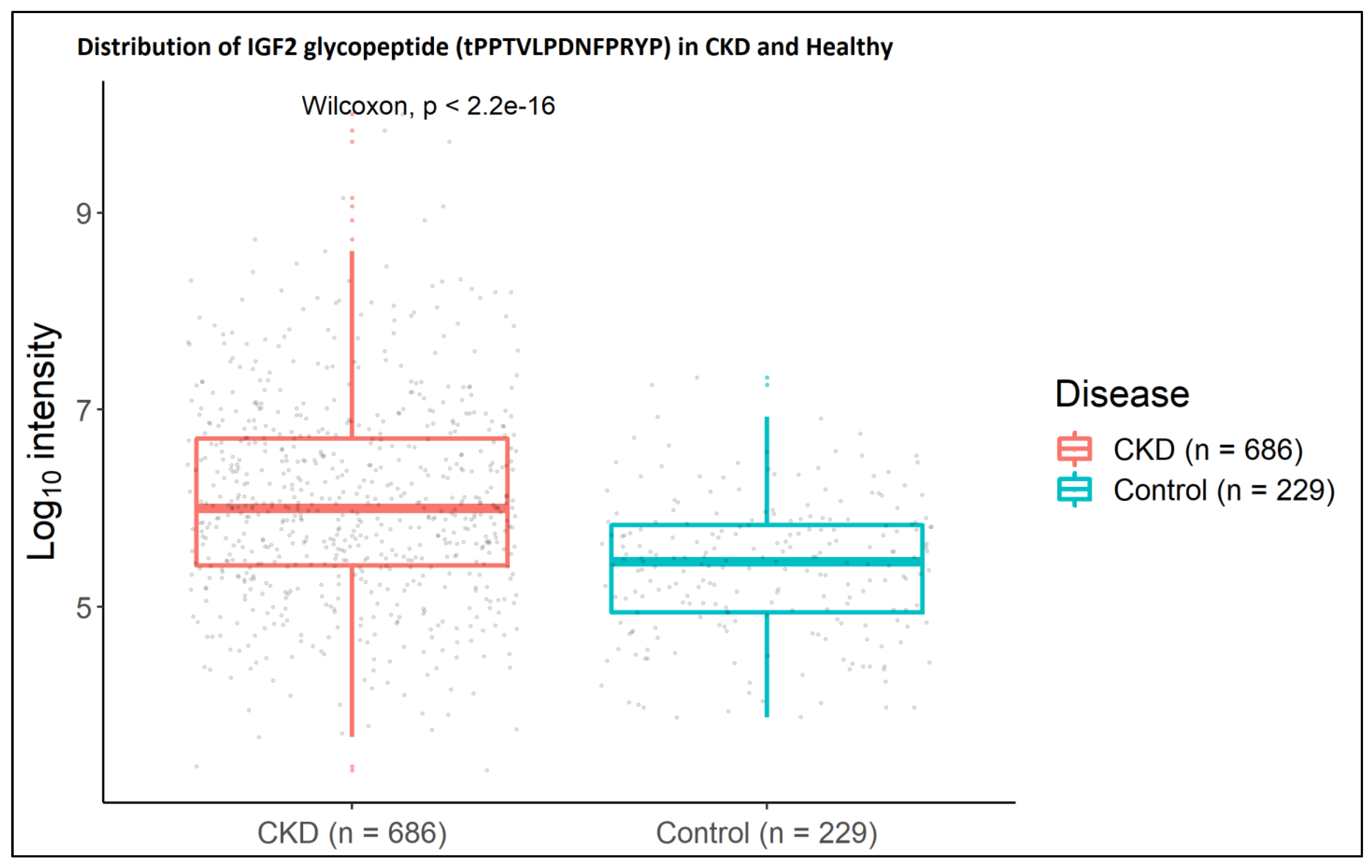
2.2. IGF2 Glycopeptides in Association to CKD
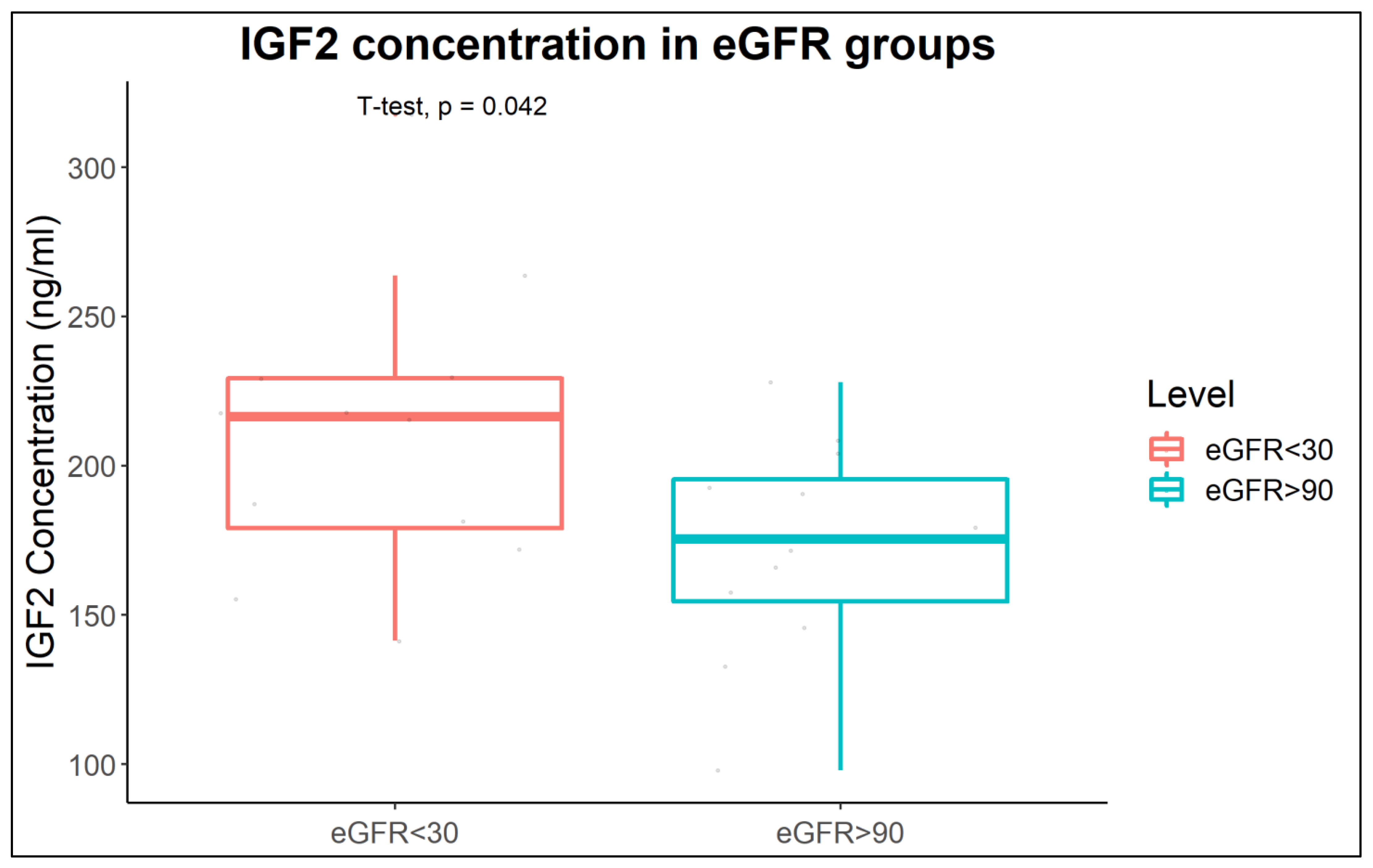
2.3. Increased IGF2 Abundance in Plasma of CKD Patients
2.4. Protease Prediction
3. Discussion
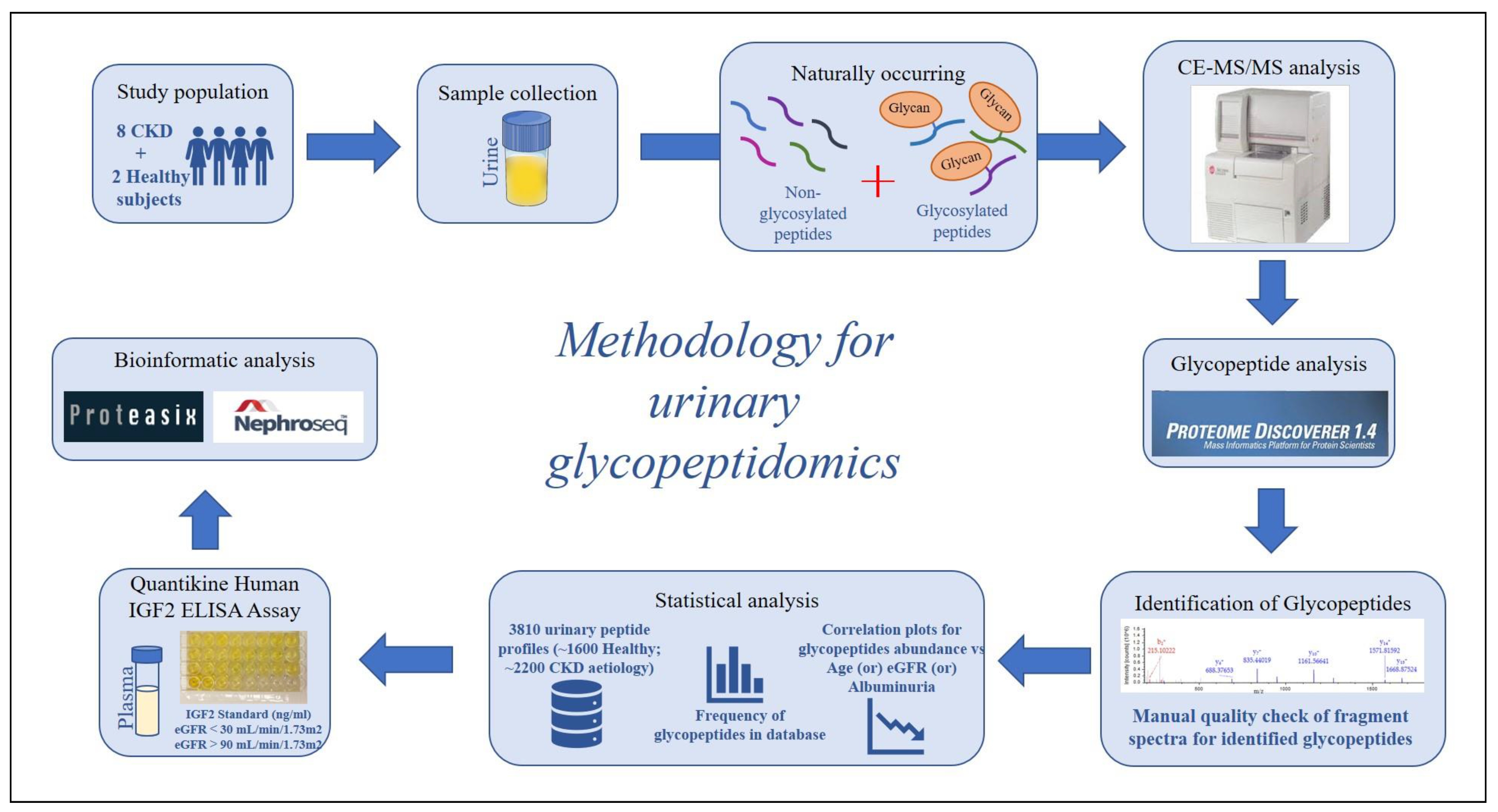
4. Materials and Methods
4.1. Study Population–Urine Samples
4.2. Sample Preparation
4.3. CE-MS/MS Analysis
4.4. Glycopeptide Analysis
4.5. Study Population–Plasma Samples
4.6. ELISA Assay
4.7. Statistical Analysis
4.8. Proteasix Analysis
4.9. NephroSeq Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ortiz, A.; Covic, A.; Fliser, D.; Fouque, D.; Goldsmith, D.; Kanbay, M.; Mallamaci, F.; Massy, Z.A.; Rossignol, P.; Vanholder, R.; et al. Epidemiology, contributors to, and clinical trials of mortality risk in chronic kidney failure. Lancet 2014, 383, 1831–1843. [Google Scholar] [CrossRef] [PubMed]
- Bellasi, A.; Di Lullo, L.; Di Iorio, B. Chronic Kidney Disease: The Silent Epidemy. J. Clin. Med. 2019, 8, 1795. [Google Scholar] [CrossRef] [Green Version]
- Levin, A.; Stevens, P.E. Summary of KDIGO 2012 CKD Guideline: Behind the scenes, need for guidance, and a framework for moving forward. Kidney Int. 2014, 85, 49–61. [Google Scholar] [CrossRef] [Green Version]
- Matsushita, K.; Coresh, J.; Sang, Y.; Chalmers, J.; Fox, C.; Guallar, E.; Jafar, T.; Jassal, S.K.; Landman, G.W.; Muntner, P.; et al. Estimated glomerular filtration rate and albuminuria for prediction of cardiovascular outcomes: A collaborative meta-analysis of individual participant data. Lancet Diabetes Endocrinol. 2015, 3, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanholder, R.; Fouque, D.; Glorieux, G.; Heine, G.H.; Kanbay, M.; Mallamaci, F.; Massy, Z.A.; Ortiz, A.; Rossignol, P.; Wiecek, A.; et al. Clinical management of the uraemic syndrome in chronic kidney disease. Lancet Diabetes Endocrinol. 2016, 4, 360–373. [Google Scholar] [CrossRef]
- Good, D.M.; Thongboonkerd, V.; Novak, J.; Bascands, J.L.; Schanstra, J.P.; Coon, J.J.; Dominiczak, A.; Mischak, H. Body fluid proteomics for biomarker discovery: Lessons from the past hold the key to success in the future. J. Proteome Res. 2007, 6, 4549–4555. [Google Scholar] [CrossRef] [PubMed]
- Pieper, R.; Gatlin, C.L.; McGrath, A.M.; Makusky, A.J.; Mondal, M.; Seonarain, M.; Field, E.; Schatz, C.R.; Estock, M.A.; Ahmed, N.; et al. Characterization of the human urinary proteome: A method for high-resolution display of urinary proteins on two-dimensional electrophoresis gels with a yield of nearly 1400 distinct protein spots. Proteomics 2004, 4, 1159–1174. [Google Scholar] [CrossRef]
- Thongboonkerd, V.; McLeish, K.R.; Arthur, J.M.; Klein, J.B. Proteomic analysis of normal human urinary proteins isolated by acetone precipitation or ultracentrifugation. Kidney Int. 2002, 62, 1461–1469. [Google Scholar] [CrossRef]
- Pisitkun, T.; Shen, R.F.; Knepper, M.A. Identification and proteomic profiling of exosomes in human urine. Proc. Natl. Acad. Sci. USA 2004, 101, 13368–13373. [Google Scholar] [CrossRef] [Green Version]
- Schaub, S.; Rush, D.; Wilkins, J.; Gibson, I.W.; Weiler, T.; Sangster, K.; Nicolle, L.; Karpinski, M.; Jeffery, J.; Nickerson, P. Proteomic-based detection of urine proteins associated with acute renal allograft rejection. J. Am. Soc. Nephrol. 2004, 15, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Magalhaes, P.; Pejchinovski, M.; Markoska, K.; Banasik, M.; Klinger, M.; Svec-Billa, D.; Rychlik, I.; Rroji, M.; Restivo, A.; Capasso, G.; et al. Association of kidney fibrosis with urinary peptides: A path towards non-invasive liquid biopsies? Sci. Rep. 2017, 7, 16915. [Google Scholar] [CrossRef] [Green Version]
- An, H.J.; Froehlich, J.W.; Lebrilla, C.B. Determination of glycosylation sites and site-specific heterogeneity in glycoproteins. Curr. Opin. Chem. Biol. 2009, 13, 421–426. [Google Scholar] [CrossRef] [Green Version]
- Lee, L.Y.; Moh, E.S.; Parker, B.L.; Bern, M.; Packer, N.H.; Thaysen-Andersen, M. Toward Automated N-Glycopeptide Identification in Glycoproteomics. J. Proteome Res. 2016, 15, 3904–3915. [Google Scholar] [CrossRef] [PubMed]
- Bollineni, R.C.; Koehler, C.J.; Gislefoss, R.E.; Anonsen, J.H.; Thiede, B. Large-scale intact glycopeptide identification by Mascot database search. Sci. Rep. 2018, 8, 2117. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Qu, Y.; Zhang, Z.; Wang, Z.; Prytkova, I.; Wu, S. Intact glycopeptide characterization using mass spectrometry. Expert Rev. Proteom. 2016, 13, 513–522. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.H.; Kolarich, D.; Packer, N.H. Mucin-type O-glycosylation--putting the pieces together. FEBS J. 2010, 277, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, F.; Sun, W.; Wu, S.; Wang, X.; Zhang, L.; Zheng, D.; Wang, J.; Gao, Y. Concanavalin A-captured glycoproteins in healthy human urine. Mol. Cell. Proteom. 2006, 5, 560–562. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.; Feng, S.; Shedden, K.; Xie, X.; Liu, Y.; Rosser, C.J.; Lubman, D.M.; Goodison, S. Urinary glycoprotein biomarker discovery for bladder cancer detection using LC/MS-MS and label-free quantification. Clin. Cancer Res. 2011, 17, 3349–3359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachi, J.; Kumar, C.; Zhang, Y.; Olsen, J.V.; Mann, M. The human urinary proteome contains more than 1500 proteins, including a large proportion of membrane proteins. Genome Biol. 2006, 7, R80. [Google Scholar] [CrossRef] [Green Version]
- Cantley, L.G.; Colangelo, C.M.; Stone, K.L.; Chung, L.; Belcher, J.; Abbott, T.; Cantley, J.L.; Williams, K.R.; Parikh, C.R. Development of a Targeted Urine Proteome Assay for kidney diseases. Proteom. Clin. Appl. 2016, 10, 58–74. [Google Scholar] [CrossRef] [Green Version]
- Coon, J.J.; Zurbig, P.; Dakna, M.; Dominiczak, A.F.; Decramer, S.; Fliser, D.; Frommberger, M.; Golovko, I.; Good, D.M.; Herget-Rosenthal, S.; et al. CE-MS analysis of the human urinary proteome for biomarker discovery and disease diagnostics. Proteom. Clin. Appl. 2008, 2, 964. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, J.; Halim, A.; Grahn, A.; Larson, G. Targeting the glycoproteome. Glycoconj. J. 2013, 30, 119–136. [Google Scholar] [CrossRef] [Green Version]
- Halim, A.; Nilsson, J.; Ruetschi, U.; Hesse, C.; Larson, G. Human urinary glycoproteomics; attachment site specific analysis of N- and O-linked glycosylations by CID and ECD. Mol. Cell. Proteom. 2012, 11, M111.013649. [Google Scholar] [CrossRef] [Green Version]
- Vivekanandan-Giri, A.; Slocum, J.L.; Buller, C.L.; Basrur, V.; Ju, W.; Pop-Busui, R.; Lubman, D.M.; Kretzler, M.; Pennathur, S. Urine glycoprotein profile reveals novel markers for chronic kidney disease. Int. J. Proteom. 2011, 2011, 214715. [Google Scholar] [CrossRef] [Green Version]
- Kolch, W.; Neususs, C.; Pelzing, M.; Mischak, H. Capillary electrophoresis-mass spectrometry as a powerful tool in clinical diagnosis and biomarker discovery. Mass Spectrom. Rev. 2005, 24, 959–977. [Google Scholar] [CrossRef] [PubMed]
- Fliser, D.; Novak, J.; Thongboonkerd, V.; Argiles, A.; Jankowski, V.; Girolami, M.A.; Jankowski, J.; Mischak, H. Advances in urinary proteome analysis and biomarker discovery. J. Am. Soc. Nephrol. 2007, 18, 1057–1071. [Google Scholar] [CrossRef] [Green Version]
- Schiffer, E.; Mischak, H.; Novak, J. High resolution proteome/peptidome analysis of body fluids by capillary electrophoresis coupled with MS. Proteomics 2006, 6, 5615–5627. [Google Scholar] [CrossRef] [PubMed]
- Good, D.M.; Zurbig, P.; Argiles, A.; Bauer, H.W.; Behrens, G.; Coon, J.J.; Dakna, M.; Decramer, S.; Delles, C.; Dominiczak, A.F.; et al. Naturally occurring human urinary peptides for use in diagnosis of chronic kidney disease. Mol. Cell. Proteom. 2010, 9, 2424–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zurbig, P.; Renfrow, M.B.; Schiffer, E.; Novak, J.; Walden, M.; Wittke, S.; Just, I.; Pelzing, M.; Neususs, C.; Theodorescu, D.; et al. Biomarker discovery by CE-MS enables sequence analysis via MS/MS with platform-independent separation. Electrophoresis 2006, 27, 2111–2125. [Google Scholar] [CrossRef] [PubMed]
- Siwy, J.; Mullen, W.; Golovko, I.; Franke, J.; Zurbig, P. Human urinary peptide database for multiple disease biomarker discovery. Proteom. Clin. Appl. 2011, 5, 367–374. [Google Scholar] [CrossRef]
- Latosinska, A.; Siwy, J.; Mischak, H.; Frantzi, M. Peptidomics and proteomics based on CE-MS as a robust tool in clinical application: The past, the present, and the future. Electrophoresis 2019, 40, 2294–2308. [Google Scholar] [CrossRef]
- Kovesdy, C.P. Epidemiology of chronic kidney disease: An update 2022. Kidney Int. Suppl. 2022, 12, 7–11. [Google Scholar] [CrossRef]
- Toghi Eshghi, S.; Yang, W.; Hu, Y.; Shah, P.; Sun, S.; Li, X.; Zhang, H. Classification of Tandem Mass Spectra for Identification of N- and O-linked Glycopeptides. Sci. Rep. 2016, 6, 37189. [Google Scholar] [CrossRef]
- Wada, Y.; Azadi, P.; Costello, C.E.; Dell, A.; Dwek, R.A.; Geyer, H.; Geyer, R.; Kakehi, K.; Karlsson, N.G.; Kato, K.; et al. Comparison of the methods for profiling glycoprotein glycans--HUPO Human Disease Glycomics/Proteome Initiative multi-institutional study. Glycobiology 2007, 17, 411–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belczacka, I.; Pejchinovski, M.; Krochmal, M.; Magalhaes, P.; Frantzi, M.; Mullen, W.; Vlahou, A.; Mischak, H.; Jankowski, V. Urinary Glycopeptide Analysis for the Investigation of Novel Biomarkers. Proteom. Clin. Appl. 2019, 13, e1800111. [Google Scholar] [CrossRef]
- Darula, Z.; Pap, A.; Medzihradszky, K.F. Extended Sialylated O-Glycan Repertoire of Human Urinary Glycoproteins Discovered and Characterized Using Electron-Transfer/Higher-Energy Collision Dissociation. J. Proteome Res. 2019, 18, 280–291. [Google Scholar] [CrossRef] [Green Version]
- Campos, D.; Freitas, D.; Gomes, J.; Magalhaes, A.; Steentoft, C.; Gomes, C.; Vester-Christensen, M.B.; Ferreira, J.A.; Afonso, L.P.; Santos, L.L.; et al. Probing the O-glycoproteome of gastric cancer cell lines for biomarker discovery. Mol. Cell. Proteom. 2015, 14, 1616–1629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajiri, M.; Yoshida, S.; Wada, Y. Differential analysis of site-specific glycans on plasma and cellular fibronectins: Application of a hydrophilic affinity method for glycopeptide enrichment. Glycobiology 2005, 15, 1332–1340. [Google Scholar] [CrossRef] [PubMed]
- Duguay, S.J.; Jin, Y.; Stein, J.; Duguay, A.N.; Gardner, P.; Steiner, D.F. Post-translational processing of the insulin-like growth factor-2 precursor. Analysis of O-glycosylation and endoproteolysis. J. Biol. Chem. 1998, 273, 18443–18451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daughaday, W.H.; Trivedi, B.; Baxter, R.C. Serum “big insulin-like growth factor II” from patients with tumor hypoglycemia lacks normal E-domain O-linked glycosylation, a possible determinant of normal propeptide processing. Proc. Natl. Acad. Sci. USA 1993, 90, 5823–5827. [Google Scholar] [CrossRef] [Green Version]
- Qiu, Q.; Basak, A.; Mbikay, M.; Tsang, B.K.; Gruslin, A. Role of pro-IGF-II processing by proprotein convertase 4 in human placental development. Proc. Natl. Acad. Sci. USA 2005, 102, 11047–11052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulle, N.; Gicquel, C.; Logie, A.; Christol, R.; Feige, J.J.; Le Bouc, Y. Fibroblast growth factor-2 inhibits the maturation of pro-insulin-like growth factor-II (Pro-IGF-II) and the expression of insulin-like growth factor binding protein-2 (IGFBP-2) in the human adrenocortical tumor cell line NCI-H295R. Endocrinology 2000, 141, 3127–3136. [Google Scholar] [CrossRef] [PubMed]
- Chao, W.; D’Amore, P.A. IGF2: Epigenetic regulation and role in development and disease. Cytokine Growth Factor Rev. 2008, 19, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Allard, J.B.; Duan, C. IGF-Binding Proteins: Why Do They Exist and Why Are There So Many? Front. Endocrinol. 2018, 9, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khosravi, M.J.; Diamandi, A.; Mistry, J.; Krishna, R.G.; Khare, A. Acid-labile subunit of human insulin-like growth factor-binding protein complex: Measurement, molecular, and clinical evaluation. J. Clin. Endocrinol. Metab. 1997, 82, 3944–3951. [Google Scholar] [CrossRef]
- Frystyk, J.; Skjaerbaek, C.; Dinesen, B.; Orskov, H. Free insulin-like growth factors (IGF-I and IGF-II) in human serum. FEBS Lett. 1994, 348, 185–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holly, J.M.P.; Biernacka, K.; Perks, C.M. The Neglected Insulin: IGF-II, a Metabolic Regulator with Implications for Diabetes, Obesity, and Cancer. Cells 2019, 8, 1207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotterill, A.M.; Holly, J.M.; Davies, S.C.; Coulson, V.J.; Price, P.A.; Wass, J.A. The insulin-like growth factors and their binding proteins in a case of non-islet-cell tumour-associated hypoglycaemia. J. Endocrinol. 1991, 131, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Itawaki, A.; Morita, M.; Miyake, H.; Yamamoto, M.; Sonoyama, H.; Tanaka, S.; Notsu, M.; Yamauchi, M.; Fujii, Y.; et al. A case of insulin-like growth factor 2-producing gastrointestinal stromal tumor with severe hypoglycemia. BMC Endocr. Disord. 2020, 20, 60. [Google Scholar] [CrossRef]
- Ozkan, Y.; Timurkan, E.S.; Aydin, S.; Sahin, I.; Timurkan, M.; Citil, C.; Kalayci, M.; Yilmaz, M.; Aksoy, A.; Catak, Z. Acylated and desacylated ghrelin, preptin, leptin, and nesfatin-1 Peptide changes related to the body mass index. Int. J. Endocrinol. 2013, 2013, 236085. [Google Scholar] [CrossRef]
- Yang, G.; Li, L.; Chen, W.; Liu, H.; Boden, G.; Li, K. Circulating preptin levels in normal, impaired glucose tolerance, and type 2 diabetic subjects. Ann. Med. 2009, 41, 52–56. [Google Scholar] [CrossRef]
- Sireesha, M.; Sambasivan, V.; Kumar, V.K.; Radha, S.; Raj, A.Y.; Qurratulain, H. Relevance of insulin-like growth factor 2 in the etiopathophysiology of diabetic nephropathy: Possible roles of phosphatase and tensin homolog on chromosome 10 and secreted protein acidic and rich in cysteine as regulators of repair. J. Diabetes 2009, 1, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y. The insulin-like growth factor system in chronic kidney disease: Pathophysiology and therapeutic opportunities. Kidney Res. Clin. Pr. 2012, 31, 26–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, L.L.; Du, R.; Liu, J.S.; Jin, J.Y.; Wang, C.Y.; Dong, Y.; He, W.X.; Yan, R.Q.; Xiang, R. Loss of RTN3 phenocopies chronic kidney disease and results in activation of the IGF2-JAK2 pathway in proximal tubular epithelial cells. Exp. Mol. Med. 2022, 54, 653–661. [Google Scholar] [CrossRef]
- Schanstra, J.P.; Zurbig, P.; Alkhalaf, A.; Argiles, A.; Bakker, S.J.; Beige, J.; Bilo, H.J.; Chatzikyrkou, C.; Dakna, M.; Dawson, J.; et al. Diagnosis and Prediction of CKD Progression by Assessment of Urinary Peptides. J. Am. Soc. Nephrol. JASN 2015, 26, 1999–2010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catanese, L.; Siwy, J.; Mavrogeorgis, E.; Amann, K.; Mischak, H.; Beige, J.; Rupprecht, H. A Novel Urinary Proteomics Classifier for Non-Invasive Evaluation of Interstitial Fibrosis and Tubular Atrophy in Chronic Kidney Disease. Proteomes 2021, 9, 32. [Google Scholar] [CrossRef]
- Pontillo, C.; Zhang, Z.Y.; Schanstra, J.P.; Jacobs, L.; Zurbig, P.; Thijs, L.; Ramirez-Torres, A.; Heerspink, H.J.L.; Lindhardt, M.; Klein, R.; et al. Prediction of Chronic Kidney Disease Stage 3 by CKD273, a Urinary Proteomic Biomarker. Kidney Int. Rep. 2017, 2, 1066–1075. [Google Scholar] [CrossRef]
- Rudnicki, M.; Siwy, J.; Wendt, R.; Lipphardt, M.; Koziolek, M.J.; Maixnerova, D.; Peters, B.; Kerschbaum, J.; Leierer, J.; Neprasova, M.; et al. Urine proteomics for prediction of disease progression in patients with IgA nephropathy. Nephrol. Dial. Transplant. 2021, 37, 42–52. [Google Scholar] [CrossRef]
- Siwy, J.; Zurbig, P.; Argiles, A.; Beige, J.; Haubitz, M.; Jankowski, J.; Julian, B.A.; Linde, P.G.; Marx, D.; Mischak, H.; et al. Noninvasive diagnosis of chronic kidney diseases using urinary proteome analysis. Nephrol. Dial. Transplant. 2017, 32, 2079–2089. [Google Scholar] [CrossRef]
- He, T.; Mischak, M.; Clark, A.L.; Campbell, R.T.; Delles, C.; Diez, J.; Filippatos, G.; Mebazaa, A.; McMurray, J.J.V.; Gonzalez, A.; et al. Urinary peptides in heart failure: A link to molecular pathophysiology. Eur. J. Heart Fail. 2021, 23, 1875–1887. [Google Scholar] [CrossRef]
- Mavrogeorgis, E.; Mischak, H.; Latosinska, A.; Vlahou, A.; Schanstra, J.P.; Siwy, J.; Jankowski, V.; Beige, J.; Jankowski, J. Collagen-Derived Peptides in CKD: A Link to Fibrosis. Toxins 2021, 14, 10. [Google Scholar] [CrossRef] [PubMed]
- Mischak, H.; Kolch, W.; Aivaliotis, M.; Bouyssie, D.; Court, M.; Dihazi, H.; Dihazi, G.H.; Franke, J.; Garin, J.; Gonzalez de Peredo, A.; et al. Comprehensive human urine standards for comparability and standardization in clinical proteome analysis. Proteom. Clin. Appl. 2010, 4, 464–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haubitz, M.; Good, D.M.; Woywodt, A.; Haller, H.; Rupprecht, H.; Theodorescu, D.; Dakna, M.; Coon, J.J.; Mischak, H. Identification and validation of urinary biomarkers for differential diagnosis and evaluation of therapeutic intervention in anti-neutrophil cytoplasmic antibody-associated vasculitis. Mol. Cell. Proteom. 2009, 8, 2296–2307. [Google Scholar] [CrossRef] [Green Version]
- Kistler, A.D.; Mischak, H.; Poster, D.; Dakna, M.; Wuthrich, R.P.; Serra, A.L. Identification of a unique urinary biomarker profile in patients with autosomal dominant polycystic kidney disease. Kidney Int. 2009, 76, 89–96. [Google Scholar] [CrossRef] [Green Version]
- Klein, J.; Papadopoulos, T.; Mischak, H.; Mullen, W. Comparison of CE-MS/MS and LC-MS/MS sequencing demonstrates significant complementarity in natural peptide identification in human urine. Electrophoresis 2014, 35, 1060–1064. [Google Scholar] [CrossRef]
- Cao, Q.; Yu, Q.; Liu, Y.; Chen, Z.; Li, L. Signature-Ion-Triggered Mass Spectrometry Approach Enabled Discovery of N- and O-Linked Glycosylated Neuropeptides in the Crustacean Nervous System. J. Proteome Res. 2020, 19, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Madsen, T.D.; Hansen, L.H.; Hintze, J.; Ye, Z.; Jebari, S.; Andersen, D.B.; Joshi, H.J.; Ju, T.; Goetze, J.P.; Martin, C.; et al. An atlas of O-linked glycosylation on peptide hormones reveals diverse biological roles. Nat. Commun. 2020, 11, 4033. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhao, W.; Mao, Y.; Chen, Y.; Zheng, S.; Cao, W.; Zhu, J.; Hu, L.; Gong, M.; Cheng, J.; et al. O-Glycosylation Landscapes of SARS-CoV-2 Spike Proteins. Front. Chem. 2021, 9, 689521. [Google Scholar] [CrossRef]
- Klein, J.; Eales, J.; Zurbig, P.; Vlahou, A.; Mischak, H.; Stevens, R. Proteasix: A tool for automated and large-scale prediction of proteases involved in naturally occurring peptide generation. Proteomics 2013, 13, 1077–1082. [Google Scholar] [CrossRef] [PubMed]
- Petra, E.; He, T.; Lygirou, V.; Latosinska, A.; Mischak, H.; Vlahou, A.; Jankowski, J. Urine peptidome analysis in cardiorenal syndrome reflects molecular processes. Sci. Rep. 2021, 11, 16219. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glycopeptide Sequence | Mass | CE-Migration Time | Protein Name | Protein Accession ID | Gene Symbol | Glycan Composition | Glycan Position | Amino Acid | Frequency | Abundance | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [Da] | [mins] | (UniProt) | Start | Stop | (%) | Range | Mean | |||||
| DVStPPTVLPDNFPRYPVG | 2726.28 | 42.55 | Insulin-like growth factor II | P01344 | IGF2 | Hex(1)HexNAc(1)NeuAc(1) | T4 | 93 | 111 | 52.86 | 17.1–49,508.6 | 6115.74 |
| tPPTVLPDNFPRYP | 2269.05 | 39.25 | Insulin-like growth factor II | P01344 | IGF2 | Hex(1)HexNAc(1)NeuAc(1) | T1 | 96 | 109 | 24.02 | 28.4–22,103.9 | 666.5 |
| DVStPPTVLPDNFPRYP | 2861.28 | 48.80 | Insulin-like growth factor II | P01344 | IGF2 | Hex(1)HexNAc(1)NeuAc(2) | T4 | 93 | 109 | 23.62 | 21.4–27,681.8 | 2067.24 |
| DVStPPTVLPDNFPRYPVGKF | 3292.53 | 39.27 | Insulin-like growth factor II | P01344 | IGF2 | Hex(1)HexNAc(1)NeuAc(2) | T4 | 93 | 113 | 89.74 | 19.0–85,843 | 9884.46 |
| DVStPPTVLPDNFPRYPVGK | 2854.37 | 35.74 | Insulin-like growth factor II | P01344 | IGF2 | Hex(1)HexNAc(1)NeuAc(1) | T4 | 93 | 112 | 0.37 | 40.43–969.34 | 281.18 |
| DVsTPPTVLPDNFPRYPVGKF | 3001.44 | 33.45 | Insulin-like growth factor II | P01344 | IGF2 | Hex(1)HexNAc(1)NeuAc(1) | S3 | 93 | 113 | 2.36 | 14.6–17,332.3 | 1904.29 |
| tPAPLDSVFSSSERAHQVLR | 3143.46 | 30.06 | Vitamin K dependent protein C | P04070 | PROC | Hex(1)HexNAc(1)NeuAc(2) | T1 | 19 | 39 | 1.58 | 10.0–589.32 | 185.93 |
| tPAPLDSVFSSSERAHQ | 2775.20 | 38.31 | Vitamin K dependent protein C | P04070 | PROC | Hex(1)HexNAc(1)NeuAc(2) | T1 | 19 | 39 | 11.05 | 6.56–4972.9 | 370.02 |
| tPAPLDSVFSSSERAHQVLRI | 3256.54 | 30.65 | Vitamin K dependent protein C | P04070 | PROC | Hex(1)HexNAc(1)NeuAc(2) | T1 | 19 | 40 | 37.09 | 16.78–35,415.4 | 2030.91 |
| sALQPSPTHSSSNTQRLPDRVTG | 3091.45 | 29.11 | Inter-alpha-trypsin inhibitor heavy chain H1 | P19827 | ITIH1 | Hex(1)HexNAc(1)NeuAc(1) | S1 | 645 | 668 | 0.05 | 50.69–97.77 | 74.23 |
| LQPsPTHSSSNTQRLPD | 2520.15 | 27.49 | Inter-alpha-trypsin inhibitor heavy chain H1 | P19827 | ITIH1 | Hex(1)HexNAc(1)NeuAc(1) | S4 | 648 | 664 | 0.55 | 12.12–270.39 | 117.54 |
| LQPsPTHSSSNTQRLPDRVTG | 2933.38 | 28.59 | Inter-alpha-trypsin inhibitor heavy chain H1 | P19827 | ITIH1 | Hex(1)HexNAc(1)NeuAc(1) | S4 | 648 | 668 | 0.32 | 44.7–3739.5 | 620.02 |
| DEAGsEADHEGTHSTKRGHAKSRPV | 3605.59 | 21.07 | Fibrinogen alpha chain | P02671 | FGA | Hex(1)HexNAc(1)NeuAc(2) | S5 | 605 | 629 | 20.71 | 18.9–4788.0 | 476.98 |
| DEAGsEADHEGTHSTKRGHAKSRP | 3215.42 | 19.996 | Fibrinogen alpha chain | P02671 | FGA | Hex(1)HexNAc(1)NeuAc(1) | S5 | 605 | 628 | 10.08 | 8.6–2873.3 | 266.59 |
| DGGFDLSDALPDNENKKPtAIP | 3260.45 | 38.15 | CD99 antigen | P14209 | CD99 | Hex(1)HexNAc(1)NeuAc(2) | T19 | 23 | 44 | 25.51 | 13.6–14,136.9 | 986.31 |
| tAVPSTGASKSKR | 1944.95 | 24.88 | Fibronectin | P02751 | FN1 | Hex(1)HexNAc(1)NeuAc(1) | T1 | 19 | 31 | 4.17 | 28.5–2130.5 | 425.66 |
| ATIPRQDEVPQQtVAPQQQ | 2789.31 | 40.96 | Tumor necrosis factor receptor superfamily member 10D | Q9UBN6 | TNFRSF10D | Hex(1)HexNAc(1)NeuAc(1) | T13 | 56 | 74 | 2.02 | 16.1–642.4 | 205.34 |
| Disease Etiology | IGF2 Glycopeptide | |||||||
|---|---|---|---|---|---|---|---|---|
| DVStPPTVLPDNFPRYPVGKF | tPPTVLPDNFPRYP | DVStPPTVLPDNFPRYPVG | DVStPPTVLPDNFPRYP | |||||
| Rho | n | Rho | n | Rho | n | Rho | n | |
| Healthy | −0.10 * | 802 | −0.23 * | 117 | −0.11 | 310 | −0.21 * | 175 |
| ADPKD | −0.19 * | 271 | 0.04 | 58 | −0.05 | 220 | −0.18 * | 120 |
| IgAN | −0.13 * | 452 | −0.51 * | 188 | 0.08 | 237 | −0.21 | 79 |
| CKD | −0.32 * | 577 | −0.58 * | 243 | −0.04 | 415 | −0.26 * | 151 |
| DKD | −0.28 * | 198 | −0.68 * | 64 | −0.01 | 132 | −0.27 | 49 |
| Nephrosclerosis | −0.18 | 118 | −0.70 * | 71 | −0.05 | 68 | −0.66 | 6 |
| FSGS | −0.26 * | 86 | −0.69 * | 32 | 0.01 | 57 | −0.1 | 13 |
| MCD | −0.41 * | 52 | −0.43 | 11 | −0.25 | 29 | −0.3 | 10 |
| MGN | −0.31 * | 72 | −0.37 | 27 | −0.19 | 49 | −0.83 * | 8 |
| MPGN | −0.38 | 20 | 0.32 | 7 | −0.04 | 16 | 0.4 | 4 |
| Nephritis | −0.69 * | 16 | −0.6 | 9 | 0.17 | 9 | ||
| Tubullar Nephritis | −0.70 * | 24 | −0.86 * | 18 | −0.46 | 16 | ||
| Healthy Subjects (n = 2) | CKD Patients (n = 8) | |
|---|---|---|
| Gender (% females) | 100 | 37.5 |
| Age (years) | 29 | 38.5 (29–50) |
| MAP (mmHg) | <90 | 89.04 (77–111) |
| Proteinuria (g/day) | <0.1 | 1.28 (0.1–2.3) |
| eGFR (mL/min/1.73 m2) | >90 | 59.34 (13–99) |
| Disease Etiology | Number of Datasets |
|---|---|
| Healthy controls | 1616 |
| Chronic kidney disease (CKD) | 865 |
| IgA nephropathy (IgAN) | 586 |
| Autosomal dominant polycystic kidney disease (ADPKD) | 290 |
| Diabetic nephropathy (DKD) | 264 |
| Nephrosclerosis | 184 |
| Focal segmental glomerulosclerosis (FSGS) | 142 |
| Membranous glomerulonephritis (MGN) | 115 |
| Minimal change disease (MCD) | 64 |
| Nephritis | 33 |
| Tubulointerstitial Nephritis | 28 |
| Membranoproliferative glomerulonephritis (MPGN) | 28 |
| eGFR > 90 mL/min/1.73 m2 (n = 12) | eGFR < 30 mL/min/1.73 m2 (n = 12) | |
|---|---|---|
| Gender (% females) | 16.7 | 75 |
| Age (years) | 38.1 (18–52) | 46.3 (18–67) |
| SBP (mmHg) | 120.3 (107–139) | 135.8 (115–164) |
| DBP (mmHg) | 77.91 (55–100) | 84.4 (72–99) |
| eGFR (mL/min/1.73 m2) | 107.8 (90–130) | 23.2 (15–29) |
| Creatinine (mmol/day) | 0.7 (0.5–0.9) | 3.0 (1.8–4.1) |
| Hemoglobin (g/dL) | 14.3 (12–16) | 13.1 (11–16) |
| Cholesterol (mg/dL) | 200.5 (155–252) | 192.3 (142–253) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lohia, S.; Latosinska, A.; Zoidakis, J.; Makridakis, M.; Mischak, H.; Glorieux, G.; Vlahou, A.; Jankowski, V. Glycosylation Analysis of Urinary Peptidome Highlights IGF2 Glycopeptides in Association with CKD. Int. J. Mol. Sci. 2023, 24, 5402. https://doi.org/10.3390/ijms24065402
Lohia S, Latosinska A, Zoidakis J, Makridakis M, Mischak H, Glorieux G, Vlahou A, Jankowski V. Glycosylation Analysis of Urinary Peptidome Highlights IGF2 Glycopeptides in Association with CKD. International Journal of Molecular Sciences. 2023; 24(6):5402. https://doi.org/10.3390/ijms24065402
Chicago/Turabian StyleLohia, Sonnal, Agnieszka Latosinska, Jerome Zoidakis, Manousos Makridakis, Harald Mischak, Griet Glorieux, Antonia Vlahou, and Vera Jankowski. 2023. "Glycosylation Analysis of Urinary Peptidome Highlights IGF2 Glycopeptides in Association with CKD" International Journal of Molecular Sciences 24, no. 6: 5402. https://doi.org/10.3390/ijms24065402