

At the Nexus between Cytoskeleton and Vacuole: How Plant Cytoskeletons Govern the Dynamics of Large Vacuoles

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
1.1. Cytoskeleton as a Rigid Yet Dynamic Intracellular Scaffold in Plant Cells
1.2. Large Vacuole, an Organelle with Versatile Functions
2. Developmental Processes
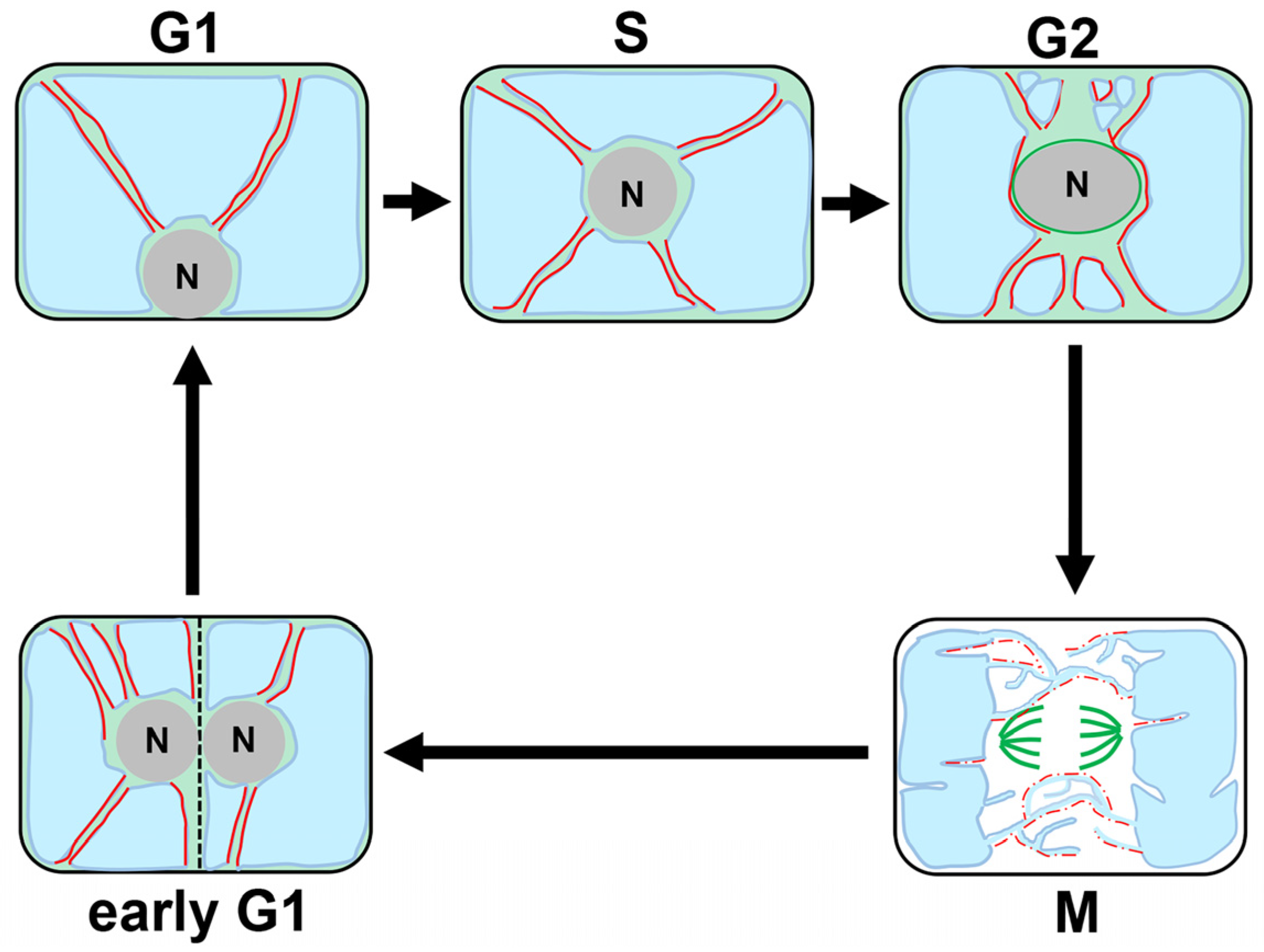
2.1. Cell Division
2.2. Cell Growth
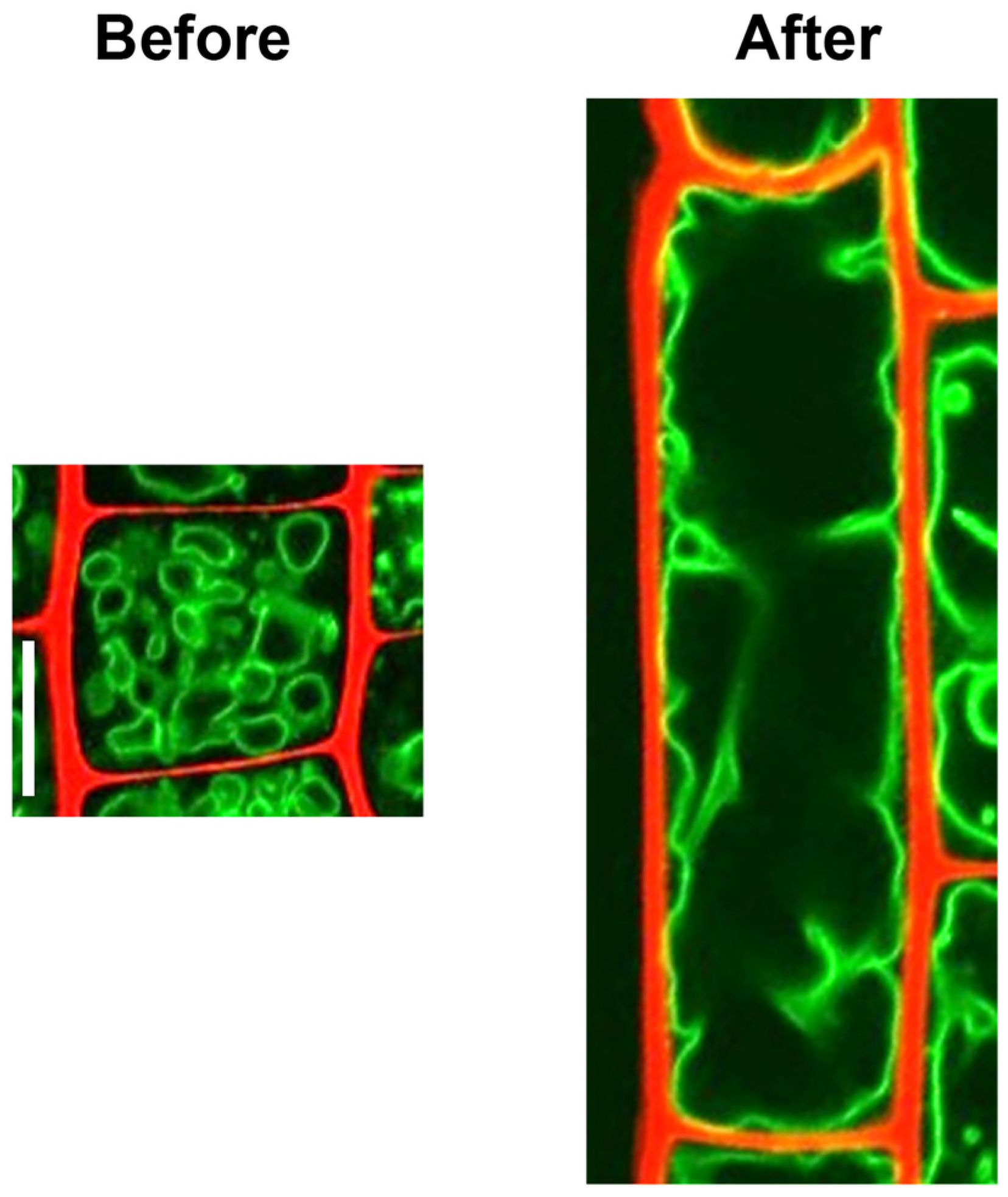
2.2.1. Diffusively Growing Cells
Root Cells
Trichome
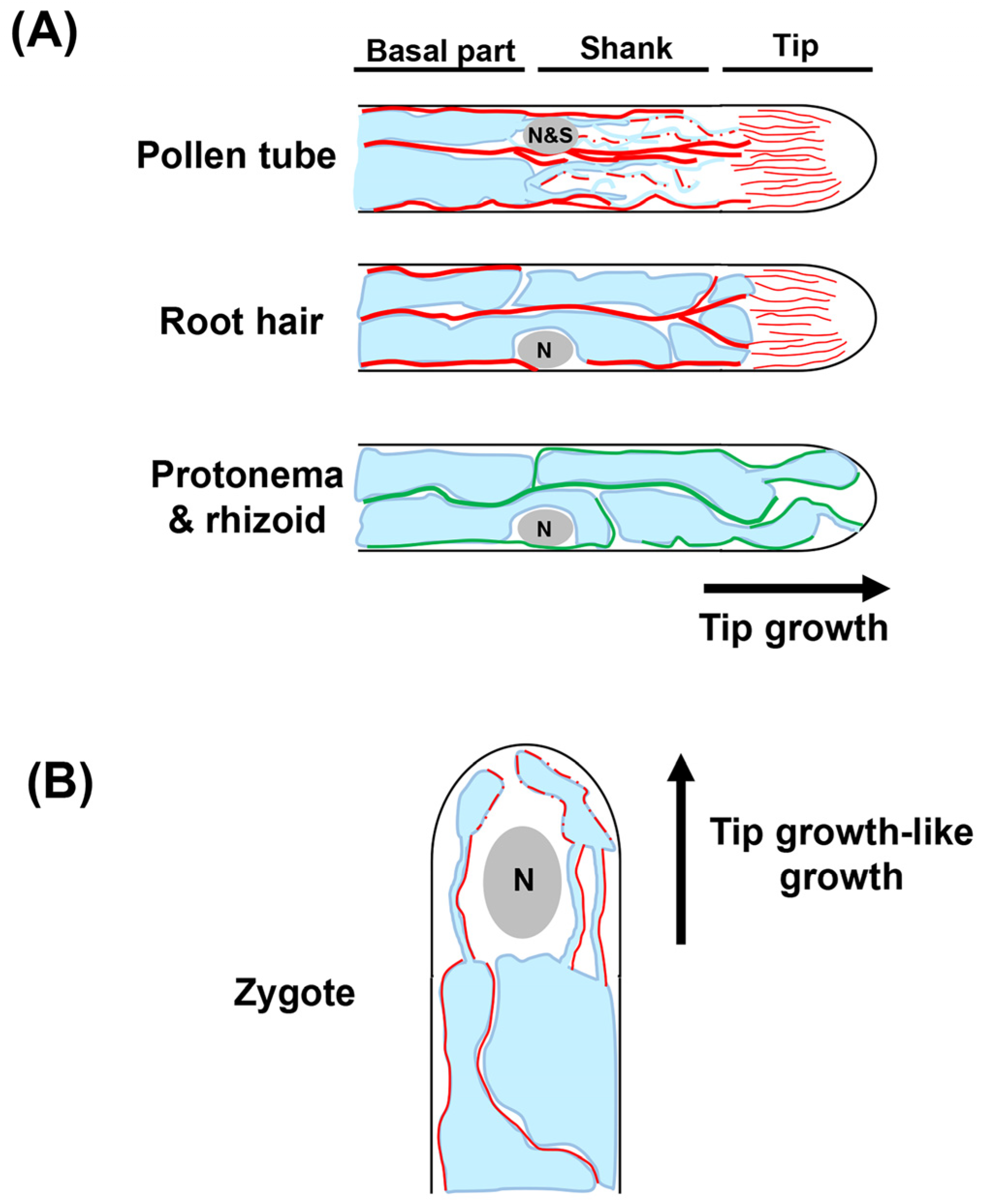
2.2.2. Tip Growing and Tip Growing-like Cells
Pollen Tube
Root Hair
Zygote
Protonema and Rhizoid in Moss
3. Responses to Environmental Stimuli
3.1. Stomatal Opening and Closure
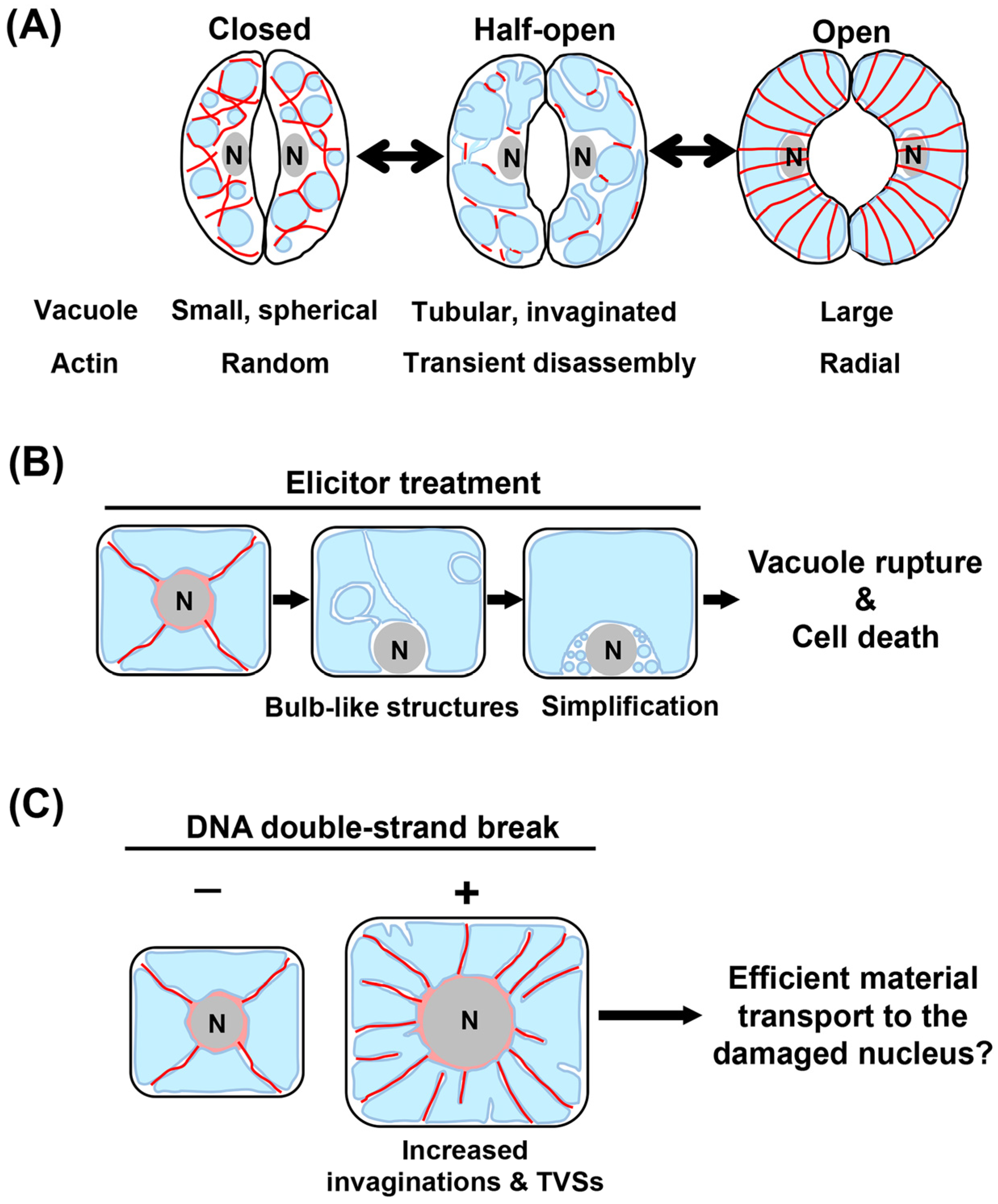
3.2. Elicitor-Induced Cell Cycle Arrest and PCD
3.3. DNA Damage-Triggered Cell Growth
4. Candidates Controlling Vacuolar Dynamics at the Nexus between Cytoskeletons and Vacuoles
4.1. Myosin
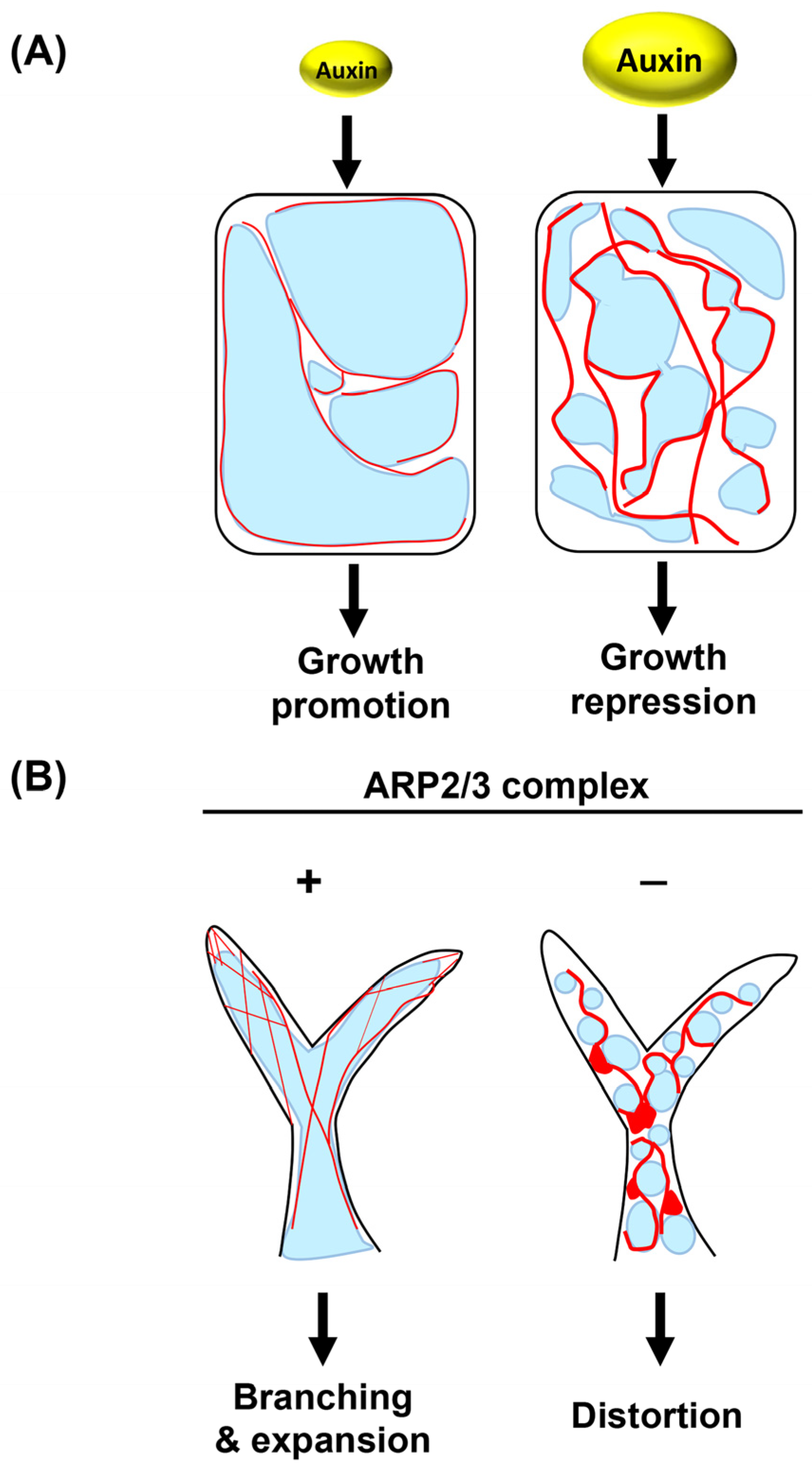
4.2. ARP2/3 Complex
4.3. The Adaptor Protein (AP) 3 Complex
4.4. NET4
5. Conclusions and Perspectives
5.1. Strategies for Studying the Roles of Vacuoles and Cytoskeletons
5.2. Strategies for Observing the Dynamics of Vacuoles and Cytoskeletons
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hohmann, T.; Dehghani, F. The cytoskeleton—A complex interacting meshwork. Cells 2019, 8, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kost, B.; Chua, N.H. The Plant Cytoskeleton: Vacuoles and Cell Walls Make the Difference. Cell 2002, 108, 9–12. [Google Scholar] [CrossRef] [Green Version]
- Coles, C.H.; Bradke, F. Coordinating Neuronal Actin-Microtubule Dynamics. Curr. Biol. 2015, 25, R677–R691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucas, J.; Geisler, M. Sequential loss of dynein sequences precedes complete loss in land plants. Plant Physiol. 2022, 189, 1237–1240. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, H.; Burkhard, P.; Aebi, U.; Herrmann, H.; Strelkov, S.V.; Burkhard, P.; Aebi, U. Intermediate filaments: Primary determinants of cell architecture and plasticity. J. Clin. Investig. 2009, 119, 1772–1783. [Google Scholar] [CrossRef] [Green Version]
- Kandasamy, M.K.; McKinney, E.C.; Roy, E.; Meagher, R.B. Plant vegetative and animal cytoplasmic actins share functional competence for spatial development with protists. Plant Cell 2012, 24, 2041–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breviario, D.; Gianì, S.; Morello, L. Multiple tubulins: Evolutionary aspects and biological implications. Plant J. 2013, 75, 202–218. [Google Scholar] [CrossRef] [PubMed]
- Utsunomiya, H.; Fujita, M.; Naito, F.; Kaneta, T. Cell cycle-dependent dynamics of a plant intermediate filament motif protein with intracellular localization related to microtubules. Protoplasma 2020, 257, 1387–1400. [Google Scholar] [CrossRef]
- Tan, X.; Li, K.; Wang, Z.; Zhu, K.; Tan, X.; Cao, J. A Review of Plant Vacuoles: Formation, Located Proteins, and Functions. Plants 2019, 8, 327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, H.; Chen, Y.; Anderson, C.T. Turgor pressure change in stomatal guard cells arises from interactions between water influx and mechanical responses of their cell walls. Quant. Plant Biol. 2022, 3, e12. [Google Scholar] [CrossRef]
- Dünser, K.; Schöller, M.; Rößling, A.K.; Löfke, C.; Xiao, N.; Pařízková, B.; Melnik, S.; Rodriguez-Franco, M.; Stöger, E.; Novák, O.; et al. Endocytic trafficking promotes vacuolar enlargements for fast cell expansion rates in plants. eLife 2022, 11, e75945. [Google Scholar] [CrossRef]
- Robert, S.; Zouhar, J.; Carter, C.; Raikhel, N. Isolation of intact vacuoles from Arabidopsis rosette leaf-derived protoplasts. Nat. Protoc. 2007, 2, 259–262. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; Cao, W.; He, Y.; Zhao, Q.; Wakazaki, M.; Zhuang, X.; Gao, J.; Zeng, Y.; Gao, C.; Ding, Y.; et al. A whole-cell electron tomography model of vacuole biogenesis in Arabidopsis root cells. Nat. Plants 2019, 5, 95–105. [Google Scholar] [CrossRef]
- Cui, Y.; Zhao, Q.; Hu, S.; Jiang, L. Vacuole Biogenesis in Plants: How Many Vacuoles, How Many Models? Trends Plant Sci. 2020, 25, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Rojo, E.; Gillmor, C.S.; Kovaleva, V.; Somerville, C.R.; Raikhel, N.V. VACUOLELESS1 Is an Essential Gene Required for Vacuole Formation and Morphogenesis in Arabidopsis. Dev. Cell 2001, 1, 303–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, L.; Liu, J.; Zhong, S.; Gu, H.; Qu, L.J. AtVPS41-mediated endocytic pathway is essential for pollen tube-stigma interaction in Arabidopsis. Proc. Natl. Acad. Sci. USA 2016, 113, 6307–6312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braidwood, L.; Breuer, C.; Sugimoto, K. My body is a cage: Mechanisms and modulation of plant cell growth. New Phytol. 2014, 201, 388–402. [Google Scholar] [CrossRef]
- Kutsuna, N.; Hasezawa, S. Dynamic organization of vacuolar and microtubule structures during cell cycle progression in synchronized tobacco BY-2 cells. Plant Cell Physiol. 2002, 43, 965–973. [Google Scholar] [CrossRef] [Green Version]
- Kutsuna, N.; Kumagai, F.; Sato, M.H.; Hasezawa, S. Three-Dimensional Reconstruction of Tubular Structure of Vacuolar Membrane Throughout Mitosis in Living Tobacco Cells. Plant Cell Physiol. 2003, 44, 1045–1054. [Google Scholar] [CrossRef] [Green Version]
- Higaki, T.; Kutsuna, N.; Okubo, E.; Sano, T.; Hasezawa, S. Actin microfilaments regulate vacuolar structures and dynamics: Dual observation of actin microfilaments and vacuolar membrane in living tobacco BY-2 cells. Plant Cell Physiol. 2006, 47, 839–852. [Google Scholar] [CrossRef] [Green Version]
- Orzechowska, M.; Stępień, K.; Kamińska, T.; Siwińska, D. Chromosome variations in regenerants of Arabidopsis thaliana derived from 2- and 6-week-old callus detected using flow cytometry and FISH analyses. Plant Cell Tissue Organ Cult. 2013, 112, 263–273. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Seo, P.J. Dynamic epigenetic changes during plant regeneration. Trends Plant Sci. 2018, 23, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Scheuring, D.; Löfke, C.; Krüger, F.; Kittelmann, M.; Eisa, A.; Hughes, L.; Smith, R.S.; Hawes, C.; Schumacher, K.; Kleine-Vehn, J. Actin-dependent vacuolar occupancy of the cell determines auxin-induced growth repression. Proc. Natl. Acad. Sci. USA 2016, 113, 452–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takatsuka, H.; Higaki, T.; Umeda, M. Actin reorganization triggers rapid cell elongation in roots. Plant Physiol. 2018, 178, 1130–1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, A.; Bannigan, A.; Sulaman, W.; Pechter, P.; Blancaflor, E.B.; Baskin, T.I. Auxin, actin and growth of the Arabidopsis thaliana primary root. Plant J. 2007, 50, 514–528. [Google Scholar] [CrossRef]
- Arieti, R.S.; Staiger, C.J. Auxin-induced actin cytoskeleton rearrangements require AUX1. New Phytol. 2020, 226, 441–459. [Google Scholar] [CrossRef] [Green Version]
- Löfke, C.; Dünser, K.; Scheuring, D.; Kleine-Vehn, J. Auxin regulates SNARE-dependent vacuolar morphology restricting cell size. eLife 2015, 4, e05868. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shen, C.; Meng, P.; Tan, G.; Lv, L. Analysis and review of trichomes in plants. BMC Plant Biol. 2021, 21, 70. [Google Scholar] [CrossRef]
- Schwab, B.; Mathur, J.; Saedler, R.; Schwarz, H.; Frey, B.; Scheidegger, C.; Hülskamp, M. Regulation of cell expansion by the DISTORTED genes in Arabidopsis thaliana: Actin controls the spatial organization of microtubules. Mol. Genet. Genom. 2003, 269, 350–360. [Google Scholar] [CrossRef]
- Li, S.; Blanchoin, L.; Yang, Z.; Lord, E.M. The Putative Arabidopsis Arp2/3 Complex Controls Leaf Cell Morphogenesis. Plant Physiol. 2003, 132, 2034–2044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, J.; Mathur, N.; Kernebeck, B.; Hülskamp, M. Mutations in actin-related proteins 2 and 3 affect cell shape development in Arabidopsis. Plant Cell 2003, 15, 1632–1645. [Google Scholar] [CrossRef] [Green Version]
- Katsaros, C.; Karyophyllis, D.; Galatis, B. Cytoskeleton and morphogenesis in brown algae. Ann. Bot. 2006, 97, 679–693. [Google Scholar] [CrossRef] [Green Version]
- Rounds, C.M.; Bezanilla, M. Growth mechanisms in tip-growing plant cells. Annu. Rev. Plant Biol. 2013, 64, 243–265. [Google Scholar] [CrossRef] [PubMed]
- Riquelme, M. Tip growth in filamentous fungi: A road trip to the apex. Annu. Rev. Microbiol. 2013, 67, 587–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephan, O.O.H. Actin fringes of polar cell growth. J. Exp. Bot. 2017, 68, 3303–3320. [Google Scholar] [CrossRef] [PubMed]
- Hepler, P.K.; Vidali, L.; Cheung, A.Y. Polarized cell growth in higher plants. Annu. Rev. Cell Dev. Biol. 2001, 17, 159–187. [Google Scholar] [CrossRef]
- Huang, J.; Dong, J.; Qu, L.J. From birth to function: Male gametophyte development in flowering plants. Curr. Opin. Plant Biol. 2021, 63, 102118. [Google Scholar] [CrossRef]
- Hicks, G.R.; Rojo, E.; Hong, S.; Carter, D.G.; Raikhel, N.V. Geminating pollen has tubular vacuoles, displays highly dynamic vacuole biogenesis, and requires VACUOLESS1 for proper function. Plant Physiol. 2004, 134, 1227–1239. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.G.; Oppenheimer, D.G. Spatial control of cell expansion by the plant cytoskeleton. Annu. Rev. Cell Dev. Biol. 2005, 21, 271–295. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Jiang, Y.; Chang, M.; Liu, X.; Zhang, R.; Huang, S. Organization and regulation of the actin cytoskeleton in the pollen tube. Front. Plant Sci. 2015, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Lovy-Wheeler, A.; Cárdenas, L.; Kunkel, J.G.; Hepler, P.K. Differential organelle movement on the actin cytoskeleton in lily pollen tubes. Cell Motil. Cytoskelet. 2007, 64, 217–232. [Google Scholar] [CrossRef]
- Onelli, E.; Scali, M.; Caccianiga, M.; Stroppa, N.; Morandini, P.; Pavesi, G.; Moscatelli, A. Microtubules play a role in trafficking prevacuolar compartments to vacuoles in tobacco pollen tubes. Open Biol. 2018, 8, 180078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepler, P.K.; Winship, L.J. The pollen tube clear zone: Clues to the mechanism of polarized growth. J. Integr. Plant Biol. 2015, 57, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Takatsuka, H.; Ito, M. Cytoskeletal Control of Planar Polarity in Root Hair Development. Front. Plant Sci. 2020, 11, 1369. [Google Scholar] [CrossRef]
- Galway, M.E.; Heckman, J.W.; Schiefelbein, J.W. Growth and ultrastructure of Arabidopsis root hairs: The rhd3 mutation alters vacuole enlargement and tip growth. Planta 1997, 201, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Stefano, G.; Brandizzi, F.; Zheng, H. Arabidopsis RHD3 mediates the generation of the tubular ER network and is required for Golgi distribution and motility in plant cells. J. Cell Sci. 2011, 124, 2241–2252. [Google Scholar] [CrossRef] [Green Version]
- Tominaga, M.; Yokota, E.; Vidali, L.; Sonobe, S.; Hepler, P.K.; Shimmen, T. The role of plant villin in the organization of the actin cytoskeleton, cytoplasmic streaming and the architecture of the transvacuolar strand in root hair cells of Hydrochraris. Planta 2000, 210, 836–843. [Google Scholar] [CrossRef]
- Ueda, M.; Zhang, Z.; Laux, T. Transcriptional Activation of Arabidopsis Axis Patterning Genes WOX8/9 Links Zygote Polarity to Embryo Development. Dev. Cell 2011, 20, 264–270. [Google Scholar] [CrossRef] [Green Version]
- Gooh, K.; Ueda, M.; Aruga, K.; Park, J.; Arata, H.; Higashiyama, T.; Kurihara, D. Live-Cell Imaging and Optical Manipulation of Arabidopsis Early Embryogenesis. Dev. Cell 2015, 34, 242–251. [Google Scholar] [CrossRef] [Green Version]
- Kimata, Y.; Higaki, T.; Kawashima, T.; Kurihara, D.; Sato, Y.; Yamada, T.; Hasezawa, S.; Berger, F.; Higashiyama, T.; Ueda, M. Cytoskeleton dynamics control the first asymmetric cell division in Arabidopsis zygote. Proc. Natl. Acad. Sci. USA 2016, 113, 14157–14162. [Google Scholar] [CrossRef] [Green Version]
- Kimata, Y.; Kato, T.; Higaki, T.; Kurihara, D.; Yamada, T.; Segami, S.; Morita, M.T.; Maeshima, M.; Hasezawa, S.; Higashiyama, T.; et al. Polar vacuolar distribution is essential for accurate asymmetric division of Arabidopsis zygotes. Proc. Natl. Acad. Sci. USA 2019, 116, 2338–2343. [Google Scholar] [CrossRef] [Green Version]
- Rensing, S.A. Plant Evo–Devo: How Tip Growth Evolved. Curr. Biol. 2016, 26, R1228–R1230. [Google Scholar] [CrossRef] [Green Version]
- Bibeau, J.P.; Galotto, G.; Wu, M.; Tüzel, E.; Vidali, L. Quantitative cell biology of tip growth in moss. Plant Mol. Biol. 2021, 107, 227–244. [Google Scholar] [CrossRef]
- Oda, Y.; Hirata, A.; Sano, T.; Fujita, T.; Hiwatashi, Y.; Sato, Y.; Kadota, A.; Hasebe, M.; Hasezawa, S. Microtubules regulate dynamic organization of vacuoles in physcomitrella patens. Plant Cell Physiol. 2009, 50, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Buckley, T.N. The control of stomata by water balance. New Phytol. 2005, 168, 275–292. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.L.; Arce Cubas, L.; Gray, J.E.; Hepworth, C. The influence of stomatal morphology and distribution on photosynthetic gas exchange. Plant J. 2020, 101, 768–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, X.Q.; Wang, X.L.; Ren, F.; Chen, J.; Wang, X.C. Dynamics of vacuoles and actin filaments in guard cells and their roles in stomatal movement. Plant Cell Environ. 2009, 32, 1108–1116. [Google Scholar] [CrossRef] [PubMed]
- Beguerisse-Diaz, M.; Hernández-Gómez, M.C.; Lizzul, A.M.; Barahona, M.; Desikan, R. Compound stress response in stomatal closure: A mathematical model of ABA and ethylene interaction in guard cells. BMC Syst. Biol. 2012, 6, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.J.; Ren, F.; Gao, X.Q.; Wei, P.C.; Wang, X.C. The reorganization of actin filaments is required for vacuolar fusion of guard cells during stomatal opening in Arabidopsis. Plant Cell Environ. 2013, 36, 484–497. [Google Scholar] [CrossRef]
- Hwang, J.U.; Lee, Y. Abscisic acid-induced actin reorganization in guard cells of dayflower is mediated by cytosolic calcium levels and by protein kinase and protein phosphatase activities. Plant Physiol. 2001, 125, 2120–2128. [Google Scholar] [CrossRef] [Green Version]
- Lemichez, E.; Wu, Y.; Sanchez, J.P.; Mettouchi, A.; Mathur, J.; Chua, N.H. Inactivation of AtRac1 by abscisic acid is essential for stomatal closure. Genes Dev. 2001, 15, 1808–1816. [Google Scholar] [CrossRef] [Green Version]
- MacRobbie, E.A.C.; Kurup, S. Signalling mechanisms in the regulation of vacuolar ion release in guard cells. New Phytol. 2007, 175, 630–640. [Google Scholar] [CrossRef]
- Gao, X.Q.; Chen, J.; Wei, P.C.; Ren, F.; Chen, J.; Wang, X.C. Array and distribution of actin filaments in guard cells contribute to the determination of stomatal aperture. Plant Cell Rep. 2008, 27, 1655–1665. [Google Scholar] [CrossRef]
- Hwang, J.-U.; Eun, S.-O.; Lee, Y. Structure and Function of Actin Filaments in Mature Guard Cells. Actin A Dyn. Framew. Mult. Plant Cell Funct. 2000, 89, 427–436. [Google Scholar] [CrossRef]
- Higaki, T.; Kutsuna, N.; Sano, T.; Kondo, N.; Hasezawa, S. Quantification and cluster analysis of actin cytoskeletal structures in plant cells: Role of actin bundling in stomatal movement during diurnal cycles in Arabidopsis guard cells. Plant J. 2010, 61, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Eisinger, W.; Ehrhardt, D.; Briggs, W. Microtubules are essential for guard-cell function in Vicia and Arabidopsis. Mol. Plant 2012, 5, 601–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dou, L.; He, K.; Peng, J.; Wang, X.; Mao, T. The E3 ligase MREL57 modulates microtubule stability and stomatal closure in response to ABA. Nat. Commun. 2021, 12, 2181. [Google Scholar] [CrossRef]
- Burke, R.; Schwarze, J.; Sherwood, O.L.; Jnaid, Y.; McCabe, P.F.; Kacprzyk, J. Stressed to Death: The Role of Transcription Factors in Plant Programmed Cell Death Induced by Abiotic and Biotic Stimuli. Front. Plant Sci. 2020, 11, 1235. [Google Scholar] [CrossRef]
- Qi, F.; Zhang, F. Cell Cycle Regulation in the Plant Response to Stress. Front. Plant Sci. 2020, 10, 1765. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M. Programmed cell death in development and defense. Plant Physiol. 2001, 125, 94–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara-Nishimura, I.; Hatsugai, N. The role of vacuole in plant cell death. Cell Death Differ. 2011, 18, 1298–1304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatsugai, N.; Kuroyanagi, M.; Yamada, K.; Meshi, T.; Tsuda, S.; Kondo, M.; Nishimura, M.; Hara-Nishimura, I. A plant vacuolar protease, VPE, mediates, virus-induced hypersensitive cell death. Science 2004, 305, 855–858. [Google Scholar] [CrossRef] [PubMed]
- Tavernier, E.; Wendehenne, D.; Blein, J.P.; Pugin, A. Involvement of Free Calcium in Action of Cryptogein, a Proteinaceous Elicitor of Hypersensitive Reaction in Tobacco Cells. Plant Physiol. 1995, 109, 1025–1031. [Google Scholar] [CrossRef] [Green Version]
- Binet, M.N.; Humbert, C.; Lecourieux, D.; Vantard, M.; Pugin, A. Disruption of Microtubular Cytoskeleton Induced by Cryptogein, an Elicitor of Hypersensitive Response in Tobacco Cells. Plant Physiol. 2001, 125, 564. [Google Scholar] [CrossRef] [Green Version]
- Kadota, Y.; Watanabe, T.; Fujii, S.; Higashi, K.; Sano, T.; Nagata, T.; Hasezawa, S.; Kuchitsu, K. Crosstalk between elicitor-induced cell death and cell cycle regulation in tobacco BY-2 cells. Plant J. 2004, 40, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Higaki, T.; Kadota, Y.; Goh, T.; Hayashi, T.; Kutsuna, N.; Sano, T.; Hasezawa, S.; Kuchitsu, K. Vacuolar and cytoskeletal dynamics during elicitor-induced programmed cell death in tobacco BY-2 cells. Plant Signal. Behav. 2008, 3, 700–703. [Google Scholar] [CrossRef] [Green Version]
- Ohno, R.; Kadota, Y.; Fujii, S.; Sekine, M.; Umeda, M.; Kuchitsu, K. Cryptogein-induced cell cycle arrest at G2 phase is associated with inhibition of cyclin-dependent kinases, suppression of expression of cell cycle-related genes and protein degradation in synchronized tobacco by-2 cells. Plant Cell Physiol. 2011, 52, 922–932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takatsuka, H.; Shibata, A.; Umeda, M. Genome maintenance mechanisms at the chromatin level. Int. J. Mol. Sci. 2021, 22, 10384. [Google Scholar] [CrossRef]
- Adachi, S.; Minamisawa, K.; Okushima, Y.; Inagaki, S.; Yoshiyama, K.; Kondou, Y.; Kaminuma, E.; Kawashima, M.; Toyoda, T.; Matsui, M.; et al. Programmed induction of endoreduplication by DNA double-strand breaks in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 10004–10009. [Google Scholar] [CrossRef] [Green Version]
- Perrot-Rechenmann, C. Cellular Responses to Auxin: Division versus Expansion. Cold Spring Harb. Perspect. Biol. 2010, 2, a001446. [Google Scholar] [CrossRef]
- Hasegawa, J.; Higaki, T.; Hamamura, Y.; Kurihara, D.; Kutsuna, N.; Higashiyama, T.; Hasezawa, S.; Matsunaga, S. Increase in invaginated vacuolar membrane structure caused by plant cell expansion by genotoxic stress induced by DNA double-strand breaks. Cytologia 2014, 79, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Carter, C.; Pan, S.; Zouhar, J.; Avila, E.L.; Girke, T.; Raikhel, N.V. The vegetative vacuole proteome of Arabidopsis thaliana reveals predicted and unexpected proteins. Plant Cell 2004, 16, 3285–3303. [Google Scholar] [CrossRef] [Green Version]
- Van Der Honing, H.S.; De Ruijter, N.C.A.; Emons, A.M.C.; Ketelaar, T. Actin and myosin regulate cytoplasm stiffness in plant cells: A study using optical tweezers. New Phytol. 2010, 185, 90–102. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.L.; Catlett, N.L.; Weisman, L.S. Actin and Myosin Function in Directed Vacuole Movement during Cell Division in Saccharomyces cerevisiae. J. Cell Biol. 1996, 135, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, F.; Kauffman, E.J.; Novak, J.L.; Nau, J.J.; Catlett, N.L.; Weisman, L.S. Regulated degradation of a class V myosin receptor directs movement of the yeast vacuole. Nature 2003, 422, 87–92. [Google Scholar] [CrossRef]
- Ishikawa, K.; Catlett, N.L.; Novak, J.L.; Tang, F.; Nau, J.J.; Weisman, L.S. Identification of an organelle-specific myosin V receptor. J. Cell Biol. 2003, 160, 887–897. [Google Scholar] [CrossRef] [Green Version]
- Mathur, J. The ARP2/3 complex: Giving plant cells a leading edge. BioEssays 2005, 27, 377–387. [Google Scholar] [CrossRef]
- Goley, E.D.; Welch, M.D. The ARP2/3 complex: An actin nucleator comes of age. Nat. Rev. Mol. Cell Biol. 2006, 7, 713–726. [Google Scholar] [CrossRef]
- García-González, J.; Kebrlová, Š.; Semerák, M.; Lacek, J.; Kotannal Baby, I.; Petrášek, J.; Schwarzerová, K. Arp2/3 Complex Is Required for Auxin-Driven Cell Expansion Through Regulation of Auxin Transporter Homeostasis. Front. Plant Sci. 2020, 11, 486. [Google Scholar] [CrossRef]
- Eitzen, G.; Wang, L.; Thorngren, N.; Wickner, W. Remodeling of organelle-bound actin is required for yeast vacuole fusion. J. Cell Biol. 2002, 158, 669–679. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Guo, X. Adaptor protein complexes and intracellular transport. Biosci. Rep. 2014, 34, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Jiang, Y.; Wang, X.; Huang, S.; Yuan, M.; Guo, Y. AP3M harbors actin filament binding activity that is crucial for vacuole morphology and stomatal closure in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 18132–18141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowles, C.R.; Odorizzi, G.; Payne, G.S.; Emr, S.D. The AP-3 adaptor complex is essential for cargo-selective transport to the yeast vacuole. Cell 1997, 91, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deeks, M.J.; Calcutt, J.R.; Ingle, E.K.S.; Hawkins, T.J.; Chapman, S.; Richardson, A.C.; Mentlak, D.A.; Dixon, M.R.; Cartwright, F.; Smertenko, A.P.; et al. A superfamily of actin-binding proteins at the actin-membrane nexus of higher plants. Curr. Biol. 2012, 22, 1595–1600. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, S.; Eisa, A.; Kleine-Vehn, J.; Scheuring, D. NET4 Modulates the compactness of vacuoles in Arabidopsis thaliana. Int. J. Mol. Sci. 2019, 20, 4752. [Google Scholar] [CrossRef] [Green Version]
- Surpin, M.; Zheng, H.; Morita, M.T.; Saito, C.; Avila, E.; Blakeslee, J.J.; Bandyopadhyay, A.; Kovaleva, V.; Carter, D.; Murphy, A.; et al. The VTI family of SNARE proteins is necessary for plant viability and mediates different protein transport pathways. Plant Cell 2003, 15, 2885–2899. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Ye, L.; Lyu, M.; Ursache, R.; Löytynoja, A.; Mähönen, A.P. An inducible genome editing system for plants. Nat. Plants 2020, 6, 766–772. [Google Scholar] [CrossRef]
- Komis, G.; Šamajová, O.; Ovečka, M.; Šamaj, J. Super-resolution Microscopy in Plant Cell Imaging. Trends Plant Sci. 2015, 20, 834–843. [Google Scholar] [CrossRef]
- Komis, G.; Mistrik, M.; Šamajová, O.; Doskočilová, A.; Ovečka, M.; Illés, P.; Bartek, J.; Šamaj, J. Dynamics and organization of cortical microtubules as revealed by superresolution structured illumination microscopy. Plant Physiol. 2014, 165, 129–148. [Google Scholar] [CrossRef] [Green Version]
- Dong, B.; Yang, X.; Zhu, S.; Bassham, D.C.; Fang, N. Stochastic Optical Reconstruction Microscopy Imaging of Microtubule Arrays in Intact Arabidopsis thaliana Seedling Roots. Sci. Rep. 2015, 5, 15694. [Google Scholar] [CrossRef] [Green Version]
- Cope, J.; Gilbert, S.; Rayment, I.; Mastronarde, D.; Hoenger, A. Cryo-electron tomography of microtubule-kinesin motor complexes. J. Struct. Biol. 2010, 170, 257–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, E.; Jacob, S.; Nemethova, M.; Resch, G.P.; Small, J.V. Electron tomography reveals unbranched networks of actin filaments in lamellipodia. Nat. Cell Biol. 2010, 12, 429–435. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takatsuka, H.; Higaki, T.; Ito, M. At the Nexus between Cytoskeleton and Vacuole: How Plant Cytoskeletons Govern the Dynamics of Large Vacuoles. Int. J. Mol. Sci. 2023, 24, 4143. https://doi.org/10.3390/ijms24044143
Takatsuka H, Higaki T, Ito M. At the Nexus between Cytoskeleton and Vacuole: How Plant Cytoskeletons Govern the Dynamics of Large Vacuoles. International Journal of Molecular Sciences. 2023; 24(4):4143. https://doi.org/10.3390/ijms24044143
Chicago/Turabian StyleTakatsuka, Hirotomo, Takumi Higaki, and Masaki Ito. 2023. "At the Nexus between Cytoskeleton and Vacuole: How Plant Cytoskeletons Govern the Dynamics of Large Vacuoles" International Journal of Molecular Sciences 24, no. 4: 4143. https://doi.org/10.3390/ijms24044143