
F-Actin Organization and Epidermal Cell Morphogenesis in the Brown Alga Sargassum vulgare

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
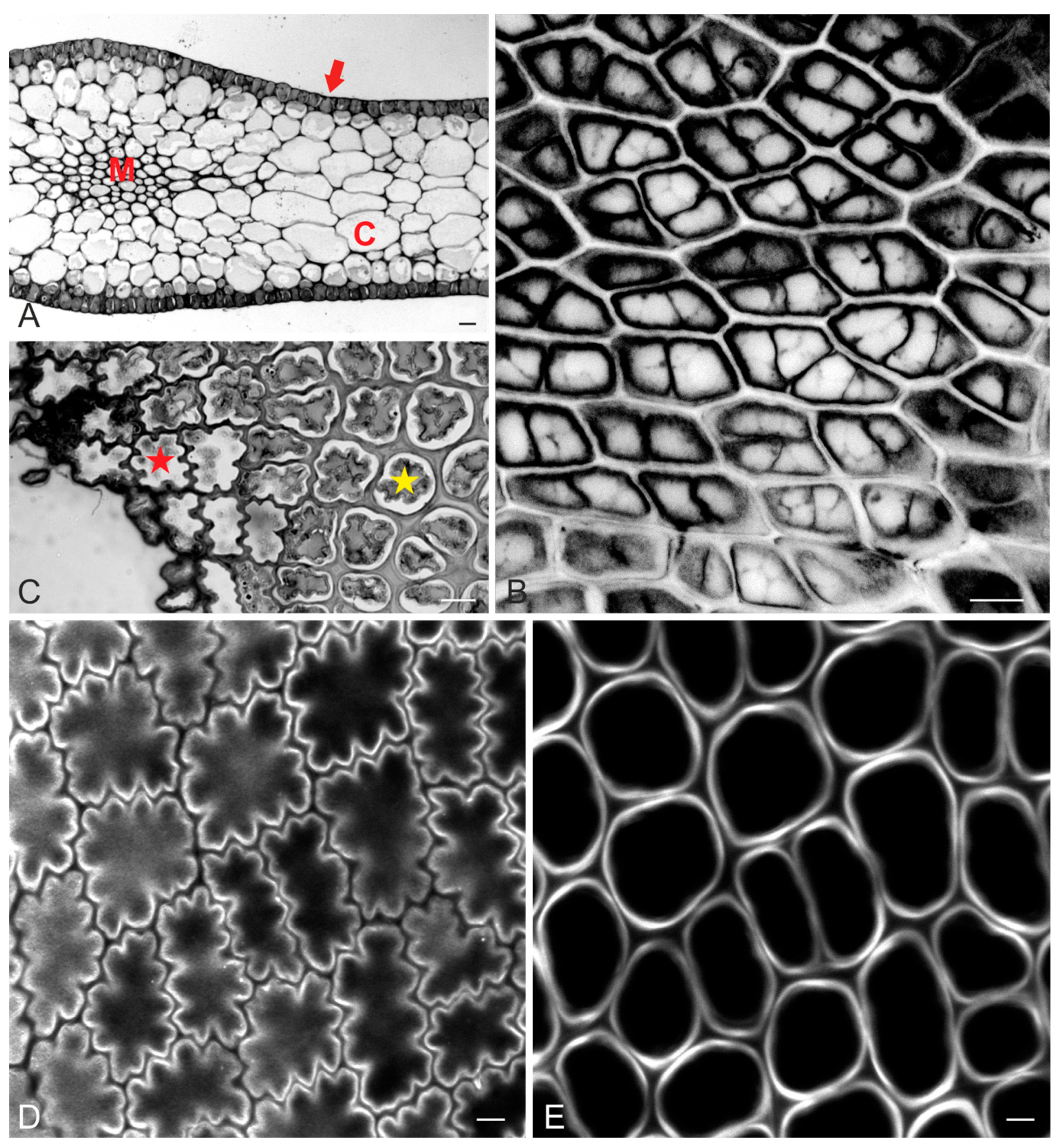
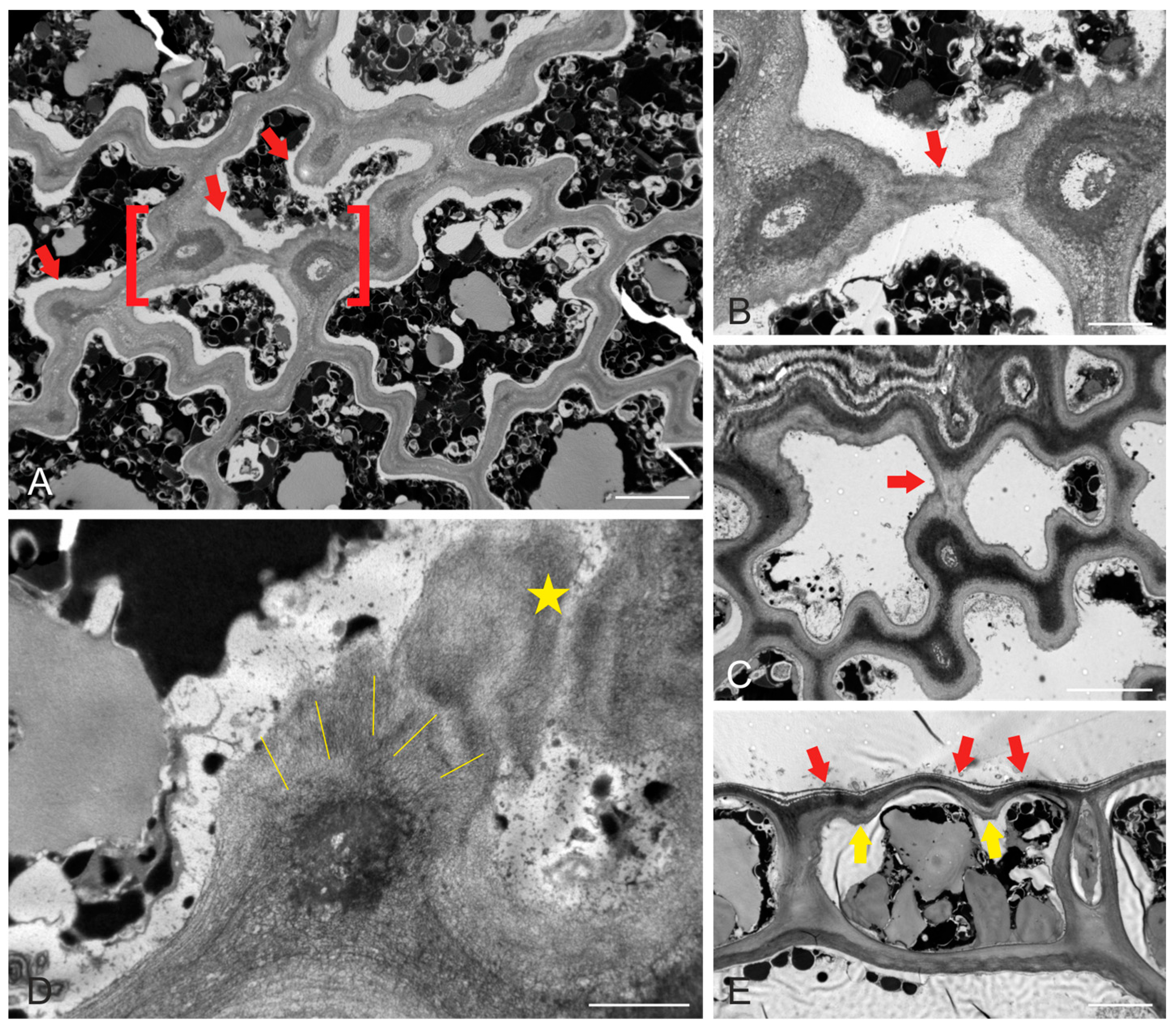
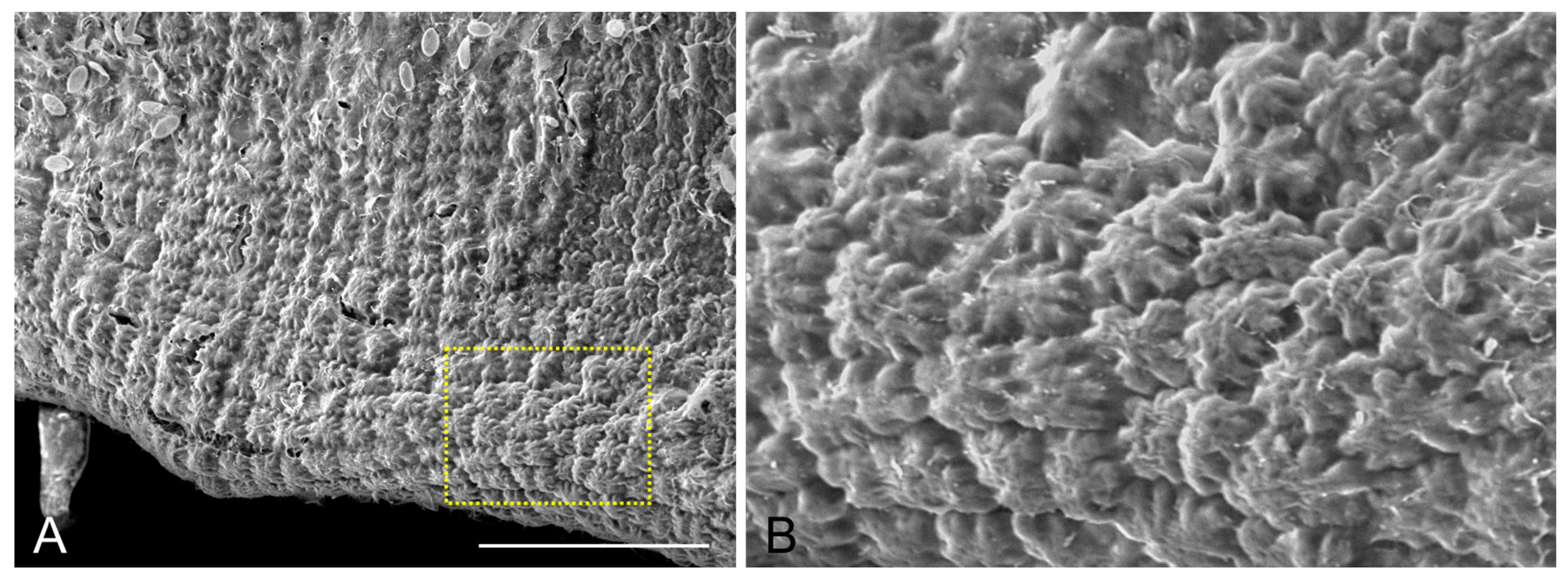
2.1. Epidermal Cell Wall and Shape
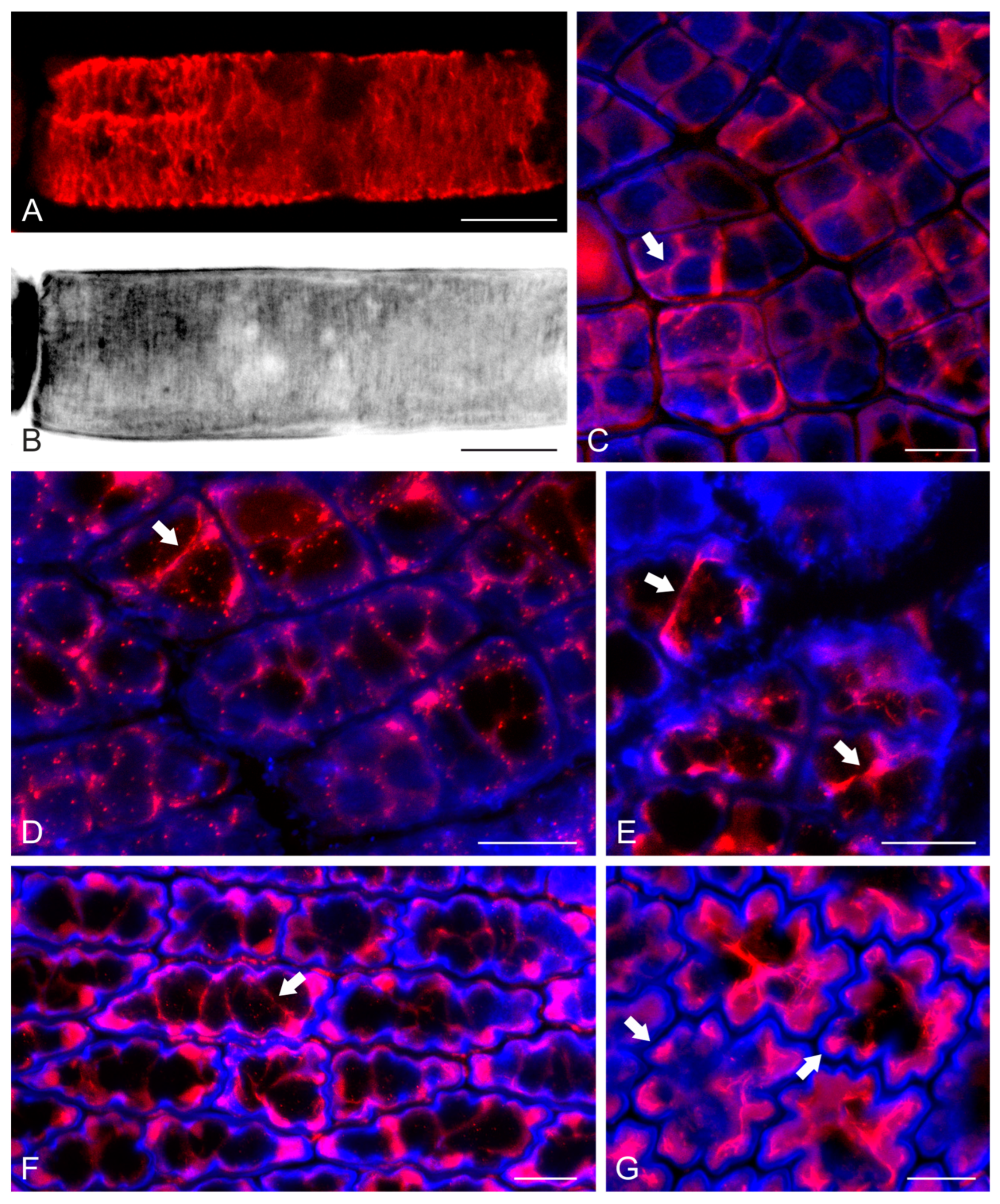
2.2. F-Actin Organization and Epidermal Cell Morphogenesis
3. Discussion
4. Materials and Methods
4.1. Sample Collection–Chemicals and Reagents
4.2. Light Microscopy and TEM
4.3. Scanning Electron Microscopy (SEM)
4.4. F-Actin and Cell Wall Fluorescent Labelling—Confocal Microscopy
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Geitmann, A.; Ortega, J.K.E. Mechanics and modeling of plant cell growth. Trends Plant Sci. 2009, 14, 467–478. [Google Scholar] [CrossRef] [PubMed]
- Wasteneys, G.O.; Fujita, M. Establishing and maintaining axial growth: Wall mechanical properties and the cytoskeleton. J. Plant Res. 2006, 119, 5–10. [Google Scholar] [CrossRef]
- Trinh, D.-C.; Alonso-Serra, J.; Asaoka, M.; Colin, L.; Cortes, M.; Malivert, A.; Takatani, S.; Zhao, F.; Traas, J.; Trehin, C.; et al. How mechanical forces shape plant organs. Curr. Biol. 2021, 31, R143–R159. [Google Scholar] [CrossRef]
- Paradez, A.; Wright, A.; Ehrhardt, D.W. Microtubule cortical array organization and plant cell morphogenesis. Curr. Opin. Plant Biol. 2006, 9, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Hsiao, A.-S.; Huang, J.-Y. Microtubule regulation in plants: From morphological development to stress adaptation. Biomolecules 2023, 13, 627. [Google Scholar] [CrossRef]
- Elliott, A.; Shaw, S.L. Update: Plant cortical microtubule arrays. Plant Physiol. 2018, 176, 94–105. [Google Scholar] [CrossRef]
- Ledbetter, M.C.; Porter, K.R.A. ‘microtubule’ in plant cell fine structure. J. Cell Biol. 1963, 19, 239–250. [Google Scholar] [CrossRef]
- Lia, S.; Leia, L.; Somerville, C.R.; Gu, Y. Cellulose synthase interactive protein 1 (CSI1) links microtubules and cellulose synthase complexes. Proc. Natl. Acad. Sci. USA 2012, 109, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Baskin, T.I.; Gu, Y. Making parallel lines meet. Cell Adhes. Migr. 2012, 6, 404–408. [Google Scholar] [CrossRef]
- Bringmann, M.; Li, E.; Sampathkumar, A.; Kocabek, T.; Hauser, M.-T.; Persson, S. POM-POM2/CELLULOSE SYNTHASE INTERACTING1 is essential for the functional association of cellulose synthase and microtubules in Arabidopsis. Plant Cell 2012, 24, 163–177. [Google Scholar] [CrossRef]
- Klimko, M.; Korszun, S.; Bykowska, J. Comparative morphology and anatomy of the leaves of Ginkgo biloba L. cultivars. Acta Sci. Pol. Hortorum Cultus 2015, 14, 169–189. [Google Scholar]
- Panteris, E.; Apostolakos, P.; Galatis, B. Microtubules and morphogenesis in ordinary epidermal cells of Vigna sinensis leaves. Protoplasma 1993, 174, 91–100. [Google Scholar] [CrossRef]
- Panteris, E.; Apostolakos, P.; Galatis, B. Microtubule organization and cell morphogenesis in two semi-lobed cell types of Adiantum capillus-veneris L. leaflets. New Phytol. 1993, 125, 509–520. [Google Scholar] [CrossRef]
- Panteris, E.; Apostolakos, P.; Galatis, B. Sinuous ordinary epidermal cells: Behind several patterns of waviness, a common morphogenetic mechanism. New Phytol. 1994, 127, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Pigg, K.B. Anatomicaly preserved Glossopteris foliage from the central Transantarctic Mountains. Rev. Palaeobot. Palynol. 1990, 66, 105–127. [Google Scholar] [CrossRef]
- Vőfély, R.V.; Gallagher, J.; Pisano, G.D.; Bartlett, M.; Braybrook, S.A. Of puzzles and pavements: A quantitative exploration of leaf epidermal cell shape. New Phytol. 2019, 221, 540–552. [Google Scholar] [CrossRef]
- Panteris, E.; Galatis, B. The morphogenesis of lobed plant cells in the mesophyll and epidermis: Organization and distinct roles of cortical microtubules and actin filaments. New Phytol. 2005, 167, 721–732. [Google Scholar] [CrossRef]
- Smith, L.G.; Oppenheimer, D. G Spatial control of cell expansion by the plant cytoskeleton. Annu. Rev. Cell Dev. Biol. 2005, 21, 271–295. [Google Scholar] [CrossRef]
- Armour, W.J.; Barton, D.A.; Law, A.M.K.; Overall, R.L. Differential growth in periclinal and anticlinal walls during lobe formation in Arabidopsis cotyledon pavement cells. Plant Cell 2015, 27, 2484–2500. [Google Scholar] [CrossRef]
- Qian, P.; Hou, S.; Guo, G. Molecular mechanisms controlling pavement cell shape in Arabidopsis leaves. Plant Cell Rep. 2009, 28, 1147–1157. [Google Scholar] [CrossRef]
- Szymanski, D.B. The kinematics and mechanics of leaf expansion: New pieces to the Arabidopsis puzzle. Curr. Opin. Plant Biol. 2014, 22, 141–148. [Google Scholar] [CrossRef]
- Eng, R.C.; Sampathkumar, A. Getting into shape: The mechanics behind plant morphogenesis. Curr. Opin. Plant Biol. 2018, 46, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Sapala, A.; Runions, A.; Smith, R.S. Mechanics, geometry and genetics of epidermal cell shape regulation: Different pieces of the same puzzle. Curr. Opin. Plant Biol. 2019, 47, 1–8. [Google Scholar] [CrossRef]
- Lin, W.; Yang, Z. Unlocking the mechanisms behind the formation of interlocking pavement cells. Curr. Opin. Plant Biol. 2020, 57, 142–154. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Jobert, F.; Rahneshan, Z.; Doyle, S.M.; Robert, S. Solving the puzzle of shape formation in plant epidermal pavement cells. Annu. Rev. Plant Biol. 2021, 72, 525–550. [Google Scholar] [CrossRef]
- Panteris, E.; Adamakis, I.-D.S. Double puzzle: Morphogenesis of the bi-layered leaf adaxial epidermis of Magnolia grandiflora. Plants 2022, 11, 3437. [Google Scholar] [CrossRef] [PubMed]
- Katsaros, C.; Karyophyllis, D.; Galatis, B. Cytoskeleton and morphogenesis in brown algae. Ann. Bot. 2006, 97, 679–693. [Google Scholar] [CrossRef]
- Karyophyllis, D.; Katsaros, C.; Galatis, B. F-actin involvement in apical cell morphogenesis of Sphacelaria rigidula (Phaeophyceae): Mutual alignment between cortical actin filaments and cellulose microfibrils. Eur. J. Phycol. 2000, 35, 195–203. [Google Scholar] [CrossRef]
- Katsaros, C.; Karyophyllis, D.; Galatis, B. F-actin cytoskeleton and cell wall morphogenesis in brown algae. Cell Biol. Int. 2003, 27, 209–210. [Google Scholar] [CrossRef]
- Sampathkumar, A.; Krupinski, P.; Wightman, R.; Milani, P.; Berquand, A.; Boudaoud, A.; Hamant, O.; Jönsson, H.; Meyerowitz, E.M. Subcellular and supracellular mechanical stress prescribes cytoskeleton behavior in Arabidopsis cotyledon pavement cells. eLife 2014, 3, e01967. [Google Scholar] [CrossRef] [PubMed]
- Sapala, A.; Runions, A.; Routier-Kierzkowska, A.-L.; Das Gupta, M.; Hong, L.; Hofhuis, H.; Verger, S.; Mosca, G.; Li, C.-B.; Hay, A.; et al. Why plants make puzzle cells, and how their shape emerges. eLife 2018, 7, e32794. [Google Scholar]
- Panteris, E.; Galatis, B.; Quader, H.; Apostolakos, P. Cortical actin filament organization in developing and functioning stomatal complexes of Zea mays and Triticum turgidum. Cell Motil. Cytoskelet. 2007, 64, 531–548. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, P.; Giannoutsou, E.; Panteris, E.; Galatis, B.; Apostolakos, P. Local differentiation of cell wall matrix polysaccharides in sinuous pavement cells: Its possible involvement in the flexibility of cell shape. Plant Biol. 2018, 20, 223–237. [Google Scholar] [CrossRef] [PubMed]
- Vogel, S. Drag and reconfiguration of broad leaves in high winds. J. Exp. Bot. 1989, 40, 941–948. [Google Scholar] [CrossRef]
- Niklas, K.J. A mechanical perspective on foliage leaf form and function. New Phytol. 1999, 143, 19–31. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panteris, E.; Pappas, D. F-Actin Organization and Epidermal Cell Morphogenesis in the Brown Alga Sargassum vulgare. Int. J. Mol. Sci. 2023, 24, 13234. https://doi.org/10.3390/ijms241713234
Panteris E, Pappas D. F-Actin Organization and Epidermal Cell Morphogenesis in the Brown Alga Sargassum vulgare. International Journal of Molecular Sciences. 2023; 24(17):13234. https://doi.org/10.3390/ijms241713234
Chicago/Turabian StylePanteris, Emmanuel, and Dimitris Pappas. 2023. "F-Actin Organization and Epidermal Cell Morphogenesis in the Brown Alga Sargassum vulgare" International Journal of Molecular Sciences 24, no. 17: 13234. https://doi.org/10.3390/ijms241713234