
Complexification of In Vitro Models of Intestinal Barriers, A True Challenge for a More Accurate Alternative Approach
 , , , , and
, , , , and
Abstract
:1. Introduction
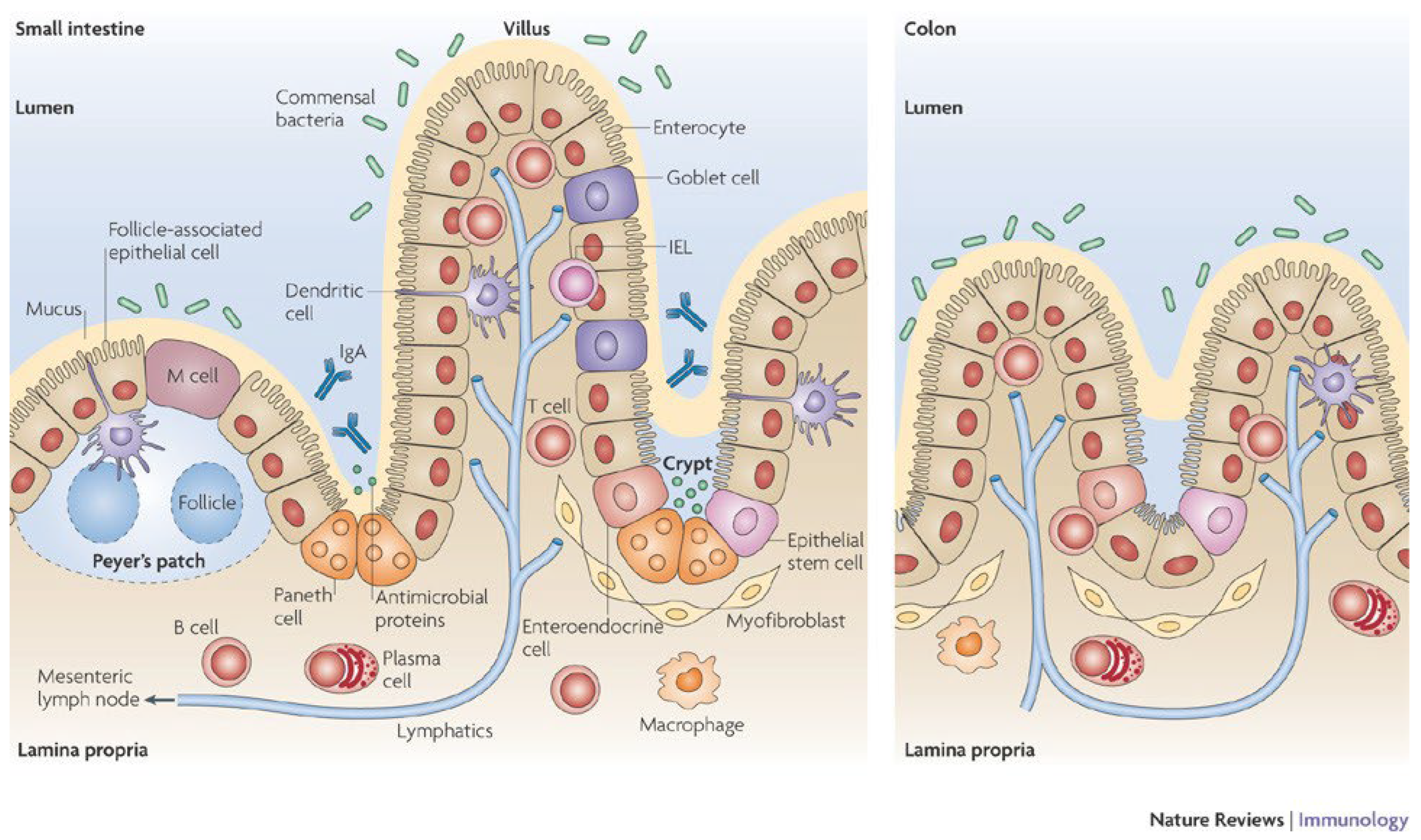
2. Organization and Functionality of the Intestinal Barrier
2.1. Role of the Intestinal Barrier
2.2. Organization of the Intestinal Barrier
2.3. Consequences of the Disruption of the Intestinal Barrier
3. Cellular Models Used to Test the Functionality of the Intestinal Barrier
3.1. Single-Cell Models Mimicking the Intestinal Physical Barrier
3.1.1. Absorptive Cells: Enterocytes and Colonocytes
Enterocyte Cell Lines Coming from Human Cancer Cells
Enterocyte Cell Lines Coming from Human Normal Cells
Colonocyte Cell Lines Coming from Human Cancer Cells
Colonocyte Cell Lines Coming from Human Normal Cells
3.1.2. Secretory Cells: Goblet, Paneth, Enteroendocrine and Tuft Cells
Mucin-Secreting Goblet Cells
Paneth Cells
Enteroendocrine Cells
3.2. Advanced Models to Investigate Gut Integrity
3.2.1. Investigation of the Epithelial Protection by the Mucus Layer
3.2.2. Investigation of the Modulation of the Innate Intestinal Immune Response of the Intestinal Barrier
3.2.3. Investigation of the Modulation of the Adaptative Intestinal Immune Response of the Intestinal Barrier
3.2.4. Investigation of the Enteroendocrine Response of the Intestinal Barrier
3.3. Advanced Models of the Gut–Lumen Interactions
3.3.1. Bioavailability Studies
3.3.2. Cytotoxicity and Inflammatory Response
3.3.3. Adhesion, Invasion and Translocation by Probiotic and Enteropathogenic Bacteria
4. Further Challenges to Obtain a More Accurate Alternative Approach
4.1. Optimizing the Co-Culture Models of the Intestinal Immune Response
4.2. Optimizing the Technical Challenges to Strengthen the Co-Culture Models
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bischoff, S.C.; Barbara, G.; Buurman, W.; Ockhuizen, T.; Schulzke, J.D.; Serino, M.; Tilg, H.; Watson, A.; Wells, J.M. Intestinal permeability—A new target for disease prevention and therapy. BMC Gastroenterol. 2014, 14, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.H.; Mushtaq, F.; Moxley-Wyles, B.; West, N.P.; Taylor, G.W.; Culmer, P.R. Real-time assessment of mechanical tissue trauma in surgery. IEEE Trans. Biomed. Eng. 2017, 64, 2384–2393. [Google Scholar] [CrossRef] [PubMed]
- Helander, H.F.; Fändriks, L. Surface area of the digestive tract—revisited. Scand. J. Gastroenterol. 2014, 49, 681–689. [Google Scholar] [CrossRef]
- Abreu, M.T. Toll-like receptor signalling in the intestinal epithelium: How bacterial recognition shapes intestinal function. Nat. Rev. Immunol. 2010, 10, 131–144. [Google Scholar] [CrossRef]
- Nicholson, J.K.; Holmes, E.; Kinross, J.; Burcelin, R.; Gibson, G.; Jia, W.; Pettersson, S. Host-Gut Microbiota Metabolic Interactions. Science 2012, 336, 1262–1267. [Google Scholar] [CrossRef]
- Robertson, R.C.; Manges, A.R.; Finlay, B.B.; Prendergast, A.J. The Human Microbiome and Child Growth – First 1000 Days and Beyond. Trends Microbiol. 2019, 27, 131–147. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Mayer, E.A.; Tillisch, K.; Gupta, A. Gut/brain axis and the microbiota. J. Clin. Investig. 2015, 125, 926–938. [Google Scholar] [CrossRef] [PubMed]
- Everson, C.A.; Toth, L.A. Systemic bacterial invasion induced by sleep deprivation. Am. J. Physiol. Integr. Comp. Physiol. 2000, 278, R905–R916. [Google Scholar] [CrossRef] [PubMed]
- Summa, K.C.; Voigt, R.M.; Forsyth, C.B.; Shaikh, M.; Cavanaugh, K.; Tang, Y.; Vitaterna, M.H.; Song, S.; Turek, F.W.; Keshavarzian, A. Disruption of the circadian clock in mice increases intestinal permeability and promotes alcohol-induced hepatic pathology and inflammation. PLoS ONE 2013, 8, e67102. [Google Scholar] [CrossRef] [Green Version]
- Hartsock, A.; Nelson, W.J. Adherens and tight junctions: Structure, function and connections to the actin cytoskeleton. Biochim. Biophys. Acta Biomembr. 2008, 1778, 660–669. [Google Scholar] [CrossRef] [PubMed]
- Lueschow, S.R.; McElroy, S.J. The Paneth cell: The curator and defender of the immature small intestine. Front. Immunol. 2020, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Price, A.E.; Shamardani, K.; Lugo, K.A.; Deguine, J.; Roberts, A.W.; Lee, B.L.; Barton, G.M. A Map of toll-like receptor expression in the intestinal epithelium reveals distinct spatial, cell type-specific, and temporal patterns. Immunity 2018, 49, 560–575. [Google Scholar] [CrossRef] [PubMed]
- Junqueira, L.C.U.; Carneiro, J. Basic Histology: Text and Atlas; McGraw-Hill: New York, NY, USA, 2005; ISBN 978-0-07-144091-2. [Google Scholar]
- Blachier, F.; de Sá Resende, A.; da Silva Fogaça Leite, G.; da Costa, A.; Lancha Junior, A.H. Colon epithelial cells luminal environment and physiopathological consequences: Impact of nutrition and exercise. Nutrire 2018, 43, 2. [Google Scholar] [CrossRef]
- Koslowski, M.; Sahin, U.; Dhaene, K.; Huber, C.; Türeci, O. MS4A12 is a colon-selective store-operated calcium channel promoting malignant cell processes. Cancer Res. 2008, 68, 3458–3466. [Google Scholar] [CrossRef]
- Johansson, M.E.V.; Sjövall, H.; Hansson, G.C. The gastrointestinal mucus system in health and disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Noah, T.K.; Donahue, B.; Shroyer, N.F. Intestinal development and differentiation. Exp. Cell Res. 2011, 317, 2702–2710. [Google Scholar] [CrossRef]
- Dillon, A.; Lo, D.D. M Cells: Intelligent engineering of mucosal immune surveillance. Front. Immunol. 2019, 10, 1499. [Google Scholar] [CrossRef]
- Corr, S.C.; CGM Gahan, C.; Hill, C. M-cells: Origin, morphology and role in mucosal immunity and microbial pathogenesis. FEMS Immunol. Med. Microbiol. 2007, 52, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Coombes, J.L.; Powrie, F. Dendritic cells in intestinal immune regulation. Nat. Rev. Immunol. 2008, 8, 435–446. [Google Scholar] [CrossRef]
- Kelsall, B. Recent progress in understanding the phenotype and function of intestinal dendritic cells and macrophages. Mucosal Immunol. 2008, 1, 460–469. [Google Scholar] [CrossRef]
- Galli, S.J.; Grimbaldeston, M.; Tsai, M. Immunomodulatory mast cells: Negative, as well as positive, regulators of immunity. Nat. Rev. Immunol. 2008, 8, 478–486. [Google Scholar] [CrossRef] [PubMed]
- Meddings, J.B.; Jarand, J.; Urbanski, S.J.; Hardin, J.; Gall, D.G. Increased gastrointestinal permeability is an early lesion in the spontaneously diabetic BB rat. Am. J. Physiol. Liver Physiol. 1999, 276, G951–G957. [Google Scholar] [CrossRef]
- Collins, S.M.; Surette, M.; Bercik, P. The interplay between the intestinal microbiota and the brain. Nat. Rev. Microbiol. 2012, 10, 735–742. [Google Scholar] [CrossRef]
- Cheng, H.; Leblond, C.P. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine I. columnar cell. Am. J. Anat. 1974, 141, 461–479. [Google Scholar] [CrossRef]
- Sambuy, Y.; De Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Janssen, A.W.F.; Duivenvoorde, L.P.M.; Rijkers, D.; Nijssen, R.; Peijnenburg, A.A.C.M.; van der Zande, M.; Louisse, J. Cytochrome P450 expression, induction and activity in human induced pluripotent stem cell-derived intestinal organoids and comparison with primary human intestinal epithelial cells and Caco-2 cells. Arch. Toxicol. 2021, 95, 907–922. [Google Scholar] [CrossRef]
- Larder, C.E.; Iskandar, M.M.; Kubow, S. Assessment of Bioavailability after In Vitro Digestion and First Pass Metabolism of Bioactive Peptides from Collagen Hydrolysates. Curr. Issues Mol. Biol. 2021, 43, 1592–1605. [Google Scholar] [CrossRef]
- Schuerer-Maly, C.C.; Eckmann, L.; Kagnoff, M.F.; Falco, M.T.; Maly, F.E. Colonic epithelial cell lines as a source of IL-8: Stimulation by inflammatory cytokines and bacterial lipopolysaccharide. Immunology 1994, 81, 85–91. [Google Scholar] [PubMed]
- Lea, T. Caco-2 Cell Line. In The Impact of Food Bioactives on Health; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 103–111. [Google Scholar]
- Engle, M.J.; Goetz, G.S.; Alpers, D.H. Caco-2 cells express a combination of colonocyte and enterocyte phenotypes. J. Cell. Physiol. 1998, 174, 362–369. [Google Scholar] [CrossRef]
- Briske-Anderson, M.J.; Finley, J.W.; Newman, S.M. The Influence of Culture Time and Passage Number on the Morphological and Physiological Development of Caco-2 Cells. Exp. Biol. Med. 1997, 214, 248–257. [Google Scholar] [CrossRef]
- Turco, L.; Catone, T.; Caloni, F.; Di Consiglio, E.; Testai, E.; Stammati, A. Caco-2/TC7 cell line characterization for intestinal absorption: How reliable is this in vitro model for the prediction of the oral dose fraction absorbed in human? Toxicol. Vitr. 2011, 25, 13–20. [Google Scholar] [CrossRef]
- Carrière, V.; Lesuffleur, T.; Barbat, A.; Rousset, M.; Dussaulx, E.; Costet, P.; de Waziers, I.; Beaune, P.; Zweibaum, A. Expression of cytochrome P 450 3A in HT29-MTX cells and Caco-2 clone TC7. FEBS Lett. 1994, 355, 247–250. [Google Scholar] [CrossRef]
- Layunta, E.; Latorre, E.; Forcén, R.; Grasa, L.; Castro, M.; Arias, M.A.; Alcalde, A.I.; Mesonero, J.E. NOD2 Modulates Serotonin Transporter and Interacts with TLR2 and TLR4 in Intestinal Epithelial Cells. Cell. Physiol. Biochem. 2018, 47, 1217–1229. [Google Scholar] [CrossRef]
- Chantret, I.; Rodolosse, A.; Barbat, A.; Dussaulx, E.; Brot-Laroche, E.; Zweibaum, A.; Rousset, M. Differential expression of sucrase-isomaltase in clones isolated from early and late passages of the cell line Caco-2: Evidence for glucose-dependent negative regulation. J. Cell Sci. 1994, 107, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Fogh, J.; Trempe, G. New human tumor cell lines. In Human Tumor Cells In Vitro; Fogh, J., Ed.; Springer US: Boston, MA, USA, 1975; pp. 115–159. ISBN 978-1-4757-1647-4. [Google Scholar]
- del Carmen Ponce de León-Rodriguez, M.; Guyot, J.-P.; Laurent-Babot, C. Intestinal in vitro cell culture models and their potential to study the effect of food components on intestinal inflammation. Crit. Rev. Food Sci. Nutr. 2019, 59, 3648–3666. [Google Scholar] [CrossRef]
- Martínez-Maqueda, D.; Miralles, B.; Recio, I. HT29 Cell Line. In The Impact of Food Bioactives on Health; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 113–124. [Google Scholar]
- Al-mousawi, H.; O’Mara, M.; Stewart, G. Identification of the HT-29 cell line as a model for investigating MCT1 transporters in sigmoid colon adenocarcinoma. Biochem. Biophys. Res. Commun. 2020, 529, 218–223. [Google Scholar] [CrossRef] [PubMed]
- Lesuffleur, T.; Porchet, N.; Aubert, J.P.; Swallow, D.; Gum, J.R.; Kim, Y.S.; Real, F.X.; Zweibaum, A. Differential expression of the human mucin genes MUC1 to MUC5 in relation to growth and differentiation of different mucus-secreting HT-29 cell subpopulations. J. Cell Sci. 1993, 106, 771–783. [Google Scholar] [CrossRef]
- Huet, C.; Sahuquillo-Merino, C.; Coudrier, E.; Louvard, D. Absorptive and mucus-secreting subclones isolated from a multipotent intestinal cell line (HT-29) provide new models for cell polarity and terminal differentiation. J. Cell Biol. 1987, 105, 345–357. [Google Scholar] [CrossRef] [PubMed]
- Collett, A.; Sims, E.; Walker, D.; He, Y.L.; Ayrton, J.; Rowland, M.; Warhurst, G. Comparison of HT29-18-C1 and Caco-2 cell lines as models for studying intestinal paracellular drug absorption. Pharm. Res. 1996, 13, 216–221. [Google Scholar] [CrossRef]
- Montoudis, A.; Delvin, E.; Menard, D.; Beaulieu, J.-F.; Jean, D.; Tremblay, E.; Bendayan, M.; Levy, E. Intestinal-fatty acid binding protein and lipid transport in human intestinal epithelial cells. Biochem. Biophys. Res. Commun. 2006, 339, 248–254. [Google Scholar] [CrossRef]
- Perreault, N.; Beaulieu, J.-F. Use of the Dissociating Enzyme Thermolysin to Generate Viable Human Normal Intestinal Epithelial Cell Cultures. Exp. Cell Res. 1996, 224, 354–364. [Google Scholar] [CrossRef]
- Guezguez, A.; Paré, F.; Benoit, Y.D.; Basora, N.; Beaulieu, J.-F. Modulation of stemness in a human normal intestinal epithelial crypt cell line by activation of the Wnt signaling pathway. Exp. Cell Res. 2014, 322, 355–364. [Google Scholar] [CrossRef]
- Claud, E.C.; Savidge, T.; Walker, W.A. Modulation of human intestinal epithelial cell IL-8 secretion by human milk factors. Pediatr. Res. 2003, 53, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Kanwar, J.R.; Kanwar, R.K. Gut health immunomodulatory and anti-inflammatory functions of gut enzyme digested high protein micro-nutrient dietary supplement-Enprocal. BMC Immunol. 2009, 10, 7. [Google Scholar] [CrossRef] [PubMed]
- Chalphin, A.V.; Fauza, D.O. Fetal tissue. In Reference Module in Biomedical Sciences; Reis, R.L., Ed.; Elsevier: Oxford, UK, 2018; pp. 299–313. ISBN 978-0-12-813700-0. [Google Scholar]
- Procházka, E.; Melvin, S.D.; Escher, B.I.; Plewa, M.J.; Leusch, F.D.L. Global transcriptional analysis of nontransformed human intestinal epithelial cells (FHs 74 Int) after exposure to selected drinking water disinfection by-products. Environ. Health Perspect. 2019, 127, 117006. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wang, Z.; Park, H.G.; Xu, C.; Lawrence, P.; Su, X.; Wijendran, V.; Walker, A.; Kothapalli, K.S.D.; Brenna, J.T. Human fetal intestinal epithelial cells metabolize and incorporate branched chain fatty acids in a structure specific manner. Prostaglandins. Leukot. Essent. Fatty Acids 2017, 116, 32–39. [Google Scholar] [CrossRef]
- Shah, U.; Dickinson, B.L.; Blumberg, R.S.; Simister, N.E.; Lencer, W.I.; Walker, A.W. Distribution of the IgG Fc Receptor, FcRn, in the Human Fetal Intestine. Pediatr. Res. 2003, 53, 295–301. [Google Scholar] [CrossRef]
- Nanthakumar, N.N.; Fusunyan, R.D.; Sanderson, I.; Walker, W.A. Inflammation in the developing human intestine: A possible pathophysiologic contribution to necrotizing enterocolitis. Proc. Natl. Acad. Sci. USA 2000, 97, 6043–6048. [Google Scholar] [CrossRef]
- Son, H.J.; Kim, N.; Song, C.-H.; Lee, S.M.; Lee, H.-N.; Surh, Y.-J. 17β-Estradiol reduces inflammation and modulates antioxidant enzymes in colonic epithelial cells. Korean J. Intern. Med. 2020, 35, 310–319. [Google Scholar] [CrossRef]
- Packiriswamy, N.; Coulson, K.F.; Holcombe, S.J.; Sordillo, L.M. Oxidative stress-induced mitochondrial dysfunction in a normal colon epithelial cell line. World, J. Gastroenterol. 2017, 23, 3427. [Google Scholar] [CrossRef] [PubMed]
- Yoo, B.K.; Yanda, M.K.; No, Y.R.; Chris Yun, C. Human intestinal epithelial cell line SK-CO15 is a new model system to study Na +/H + exchanger 3. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Yamaura, Y.; Chapron, B.D.; Wang, Z.; Himmelfarb, J.; Thummel, K.E. Functional comparison of human colonic carcinoma cell lines and primary small intestinal epithelial cells for investigations of intestinal drug permeability and first-pass metabolism. Drug Metab Dispos 2016, 44, 329–335. [Google Scholar] [CrossRef]
- Leonard, F.; Collnot, E.-M.; Lehr, C.-M. A three-dimensional coculture of enterocytes, monocytes and dendritic cells to model inflamed intestinal mucosa in vitro. Mol. Pharm. 2010, 7, 2103–2119. [Google Scholar] [CrossRef]
- Ramzy, G.M.; Boschung, L.; Koessler, T.; Delucinge-Vivier, C.; Docquier, M.; McKee, T.A.; Rubbia-Brandt, L.; Nowak-Sliwinska, P. FOLFOXIRI resistance induction and characterization in human colorectal cancer cells. Cancers 2022, 14, 4812. [Google Scholar] [CrossRef] [PubMed]
- Sazonova, E.V.; Chesnokov, M.S.; Zhivotovsky, B.; Kopeina, G.S. Drug toxicity assessment: Cell proliferation versus cell death. Cell Death Discov. 2022, 8, 417. [Google Scholar] [CrossRef]
- Trainer, D.L.; Kline, T.; McCabe, F.L.; Faucette, L.F.; Feild, J.; Chaikin, M.; Anzano, M.; Reiman, D.; Hoffstein, S.; Li, D.-J.; et al. Biological characterization and oncogene expression in human colorectal carcinoma cell lines. Int. J. Cancer 1988, 41, 287–296. [Google Scholar] [CrossRef]
- Chik, J.H.L.; Zhou, J.; Moh, E.S.X.; Christopherson, R.; Clarke, S.J.; Molloy, M.P.; Packer, N.H. Comprehensive glycomics comparison between colon cancer cell cultures and tumours: Implications for biomarker studies. J. Proteom. 2014, 108, 146–162. [Google Scholar] [CrossRef]
- Sharma, M.; Agrawal, S.K.; Sharma, P.R.; Chadha, B.S.; Khosla, M.K.; Saxena, A.K. Cytotoxic and apoptotic activity of essential oil from Ocimum viride towards COLO 205 cells. Food Chem. Toxicol. 2010, 48, 336–344. [Google Scholar] [CrossRef]
- Markiewicz, L.H.; Ogrodowczyk, A.M.; Wiczkowski, W.; Wróblewska, B. Phytate hydrolysate differently modulates the immune response of human healthy and cancer colonocytes to intestinal bacteria. Nutrients 2022, 14, 4234. [Google Scholar] [CrossRef]
- Crowley-Weber, C.L. Development and molecular characterization of HCT-116 cell lines resistant to the tumor promoter and multiple stress-inducer, deoxycholate. Carcinogenesis 2002, 23, 2063–2080. [Google Scholar] [CrossRef]
- Vermeulen, S.J.; Chen, T.R.; Speleman, F.; Nollet, F.; Van Roy, F.M.; Mareel, M.M. Did the Four Human Cancer Cell Lines DLD-1, HCT-15, HCT-8, and HRT-18 Originate from One and the Same Patient? Cancer Genet. Cytogenet. 1998, 107, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Mikeš, J.; Hýžďalová, M.; Kočí, L.; Jendželovský, R.; Kovaľ, J.; Vaculová, A.; Hofmanová, J.; Kozubík, A.; Fedoročko, P. Lower sensitivity of FHC fetal colon epithelial cells to photodynamic therapy compared to HT-29 colon adenocarcinoma cells despite higher intracellular accumulation of hypericin. Photochem. Photobiol. Sci. 2011, 10, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Kumar, C.K.; Moyer, M.P.; Dudeja, P.K.; Said, H.M. A protein-tyrosine kinase-regulated, pH-dependent, carrier-mediated uptake system for folate in human normal colonic epithelial cell line NCM460. J. Biol. Chem. 1997, 272, 6226–6231. [Google Scholar] [CrossRef]
- Sahi, J.; Nataraja, S.G.; Layden, T.J.; Goldstein, J.L.; Moyer, M.P.; Rao, M.C. Cl − transport in an immortalized human epithelial cell line (NCM460) derived from the normal transverse colon. Am. J. Physiol. Physiol. 1998, 275, C1048–C1057. [Google Scholar] [CrossRef] [PubMed]
- Sebens, S.; Bauer, I.; Geismann, C.; Grage-Griebenow, E.; Ehlers, S.; Kruse, M.-L.; Arlt, A.; Schäfer, H. Inflammatory macrophages induce Nrf2 transcription factor-dependent proteasome activity in colonic NCM460 cells and thereby confer anti-apoptotic protection. J. Biol. Chem. 2011, 286, 40911–40921. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.A.; Carroll, R.E.; Rasenick, M.M.; Benya, R.V. Constitutive activation of the gastrin-releasing peptide receptor expressed by the nonmalignant human colon epithelial cell line {NCM460}. J. Clin. Investig. 1997, 100, 2530–2537. [Google Scholar] [CrossRef]
- Moyer, M.P.; Manzano, L.A.; Merriman, R.L.; Stauffer, J.S.; Tanzer, L.R. NCM460, a normal human colon mucosal epithelial cell line. Vitr. Cell. Dev. Biol. Anim. 1996, 32, 315–317. [Google Scholar] [CrossRef]
- Brozek-Pluska, B. Statistics assisted analysis of Raman spectra and imaging of human colon cell lines – Label free, spectroscopic diagnostics of colorectal cancer. J. Mol. Struct. 2020, 1218, 128524. [Google Scholar] [CrossRef]
- Fogh, J.; Fogh, J.M.; Orfeo, T. One hundred and twenty-seven cultured human tumor cell lines producing tumors in nude mice. JNCI J. Natl. Cancer Inst. 1977, 59, 221–226. [Google Scholar] [CrossRef]
- Solano, L.N.; Nelson, G.L.; Ronayne, C.T.; Jonnalagadda, S.; Jonnalagadda, S.K.; Kottke, K.; Chitren, R.; Johnson, J.L.; Pandey, M.K.; Jonnalagadda, S.C.; et al. Synthesis, in vitro, and in vivo evaluation of novel {N}-phenylindazolyl diarylureas as potential anti-cancer agents. Sci. Rep. 2020, 10, 17969. [Google Scholar] [CrossRef]
- Wasserman, S.I.; Barrett, K.E.; Huott, P.A.; Beuerlein, G.; Kagnoff, M.F.; Dharmsathaphorn, K. Immune-related intestinal Cl- secretion. I. Effect of histamine on the T84 cell line. Am. J. Physiol. Physiol. 1988, 254, C53–C62. [Google Scholar] [CrossRef]
- Le Bivic, A.; Real, F.X.; Rodriguez-Boulan, E. Vectorial targeting of apical and basolateral plasma membrane proteins in a human adenocarcinoma epithelial cell line. Proc. Natl. Acad. Sci. USA 1989, 86, 9313–9317. [Google Scholar] [CrossRef]
- Lechuga, S.; Cartagena-Rivera, A.X.; Khan, A.; Crawford, B.I.; Narayanan, V.; Conway, D.E.; Lehtimäki, J.; Lappalainen, P.; Rieder, F.; Longworth, M.S.; et al. A myosin chaperone, UNC-45A, is a novel regulator of intestinal epithelial barrier integrity and repair. FASEB J. 2022, 36, e22290. [Google Scholar] [CrossRef] [PubMed]
- Strainiene, E.; Binkis, M.; Urnikyte, S.; Stankevicius, V.; Sasnauskiene, A.; Kundrotas, G.; Kazlauskas, A.; Suziedelis, K. Microenvironment dependent gene expression signatures in reprogrammed human colon normal and cancer cell lines. BMC Cancer 2018, 18, 222. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Deng, C.; Peng, M.; Li, C.; Teng, Y.; Guo, S.; Wu, T.; Yi, D.; Hou, Y. Integration of transcriptomic and proteomic analyses reveals protective mechanisms of N-acetylcysteine in indomethacin-stimulated enterocytes. J. Nutr. Biochem. 2023, 112, 109231. [Google Scholar] [CrossRef]
- Ren, H.; Li, Z.; Gao, R.; Zhao, T.; Luo, D.; Yu, Z.; Zhang, S.; Qi, C.; Wang, Y.; Qiao, H.; et al. Structural characteristics of Rehmannia glutinosa polysaccharides treated using different decolorization processes and their antioxidant effects in intestinal epithelial cells. Foods 2022, 11, 3449. [Google Scholar] [CrossRef]
- Cencič, A.; Langerholc, T. Functional cell models of the gut and their applications in food microbiology—A review. Int. J. Food Microbiol. 2010, 141, S4–S14. [Google Scholar] [CrossRef] [PubMed]
- Quaroni, A.; Wands, J.; Trelstad, R.L.; Isselbacher, K.J. Epithelioid cell cultures from rat small intestine. Characterization by morphologic and immunologic criteria. J. Cell Biol. 1979, 80, 248–265. [Google Scholar] [CrossRef]
- Rousset, M. The human colon carcinoma cell lines HT-29 and Caco-2: Two in vitro models for the study of intestinal differentiation. Biochimie 1986, 68, 1035–1040. [Google Scholar] [CrossRef]
- Hilgendorf, C.; Spahn-Langguth, H.; Regårdh, C.G.; Lipka, E.; Amidon, G.L.; Langguth, P. Caco-2 versus Caco-2/HT29-MTX Co-cultured Cell Lines: Permeabilities Via Diffusion, Inside- and Outside-Directed Carrier-Mediated Transport. J. Pharm. Sci. 2000, 89, 63–75. [Google Scholar] [CrossRef]
- Hoffmann, P.; Burmester, M.; Langeheine, M.; Brehm, R.; Empl, M.T.; Seeger, B.; Breves, G. Caco-2/HT29-MTX co-cultured cells as a model for studying physiological properties and toxin-induced effects on intestinal cells. PLoS ONE 2021, 16, e0257824. [Google Scholar] [CrossRef] [PubMed]
- Lesuffleur, T.; Barbat, A.; Dussaulx, E.; Zweibaum, A. Growth adaptation to methotrexate of HT-29 human colon carcinoma cells is associated with their ability to differentiate into columnar absorptive and mucus-secreting Cells. Cancer Res. 1990, 50, 6334–6343. [Google Scholar]
- Yuan, H.; Chen, C.-Y.; Chai, G.; Du, Y.-Z.; Hu, F.-Q. Improved transport and absorption through gastrointestinal tract by PEGylated solid lipid nanoparticles. Mol. Pharm. 2013, 10, 1865–1873. [Google Scholar] [CrossRef]
- Rutzky, L.P.; Kaye, C.I.; Siciliano, M.J.; Chao, M.; Kahan, B.D. Longitudinal karyotype and genetic signature analysis of cultured human colon adenocarcinoma cell lines LS180 and LS174T. Cancer Res. 1980, 40, 1443–1448. [Google Scholar] [PubMed]
- Gagnon, J.; Brubaker, P.L. NCI-H716 Cells. In The Impact of Food Bioactives on Health; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 221–228. [Google Scholar]
- Park, J.G.; Kramer, B.S.; Steinberg, S.M.; Carmichael, J.; Collins, J.M.; Minna, J.D.; Gazdar, A.F. Chemosensitivity testing of human colorectal carcinoma cell lines using a tetrazolium-based colorimetric assay. Cancer Res. 1987, 47, 5875–5879. [Google Scholar] [PubMed]
- Reimer, R.A.; Darimont, C.; Gremlich, S.; Nicolas-Métral, V.; Rüegg, U.T.; Macé, K. A human cellular model for studying the regulation of glucagon-like peptide-1 secretion. Endocrinology 2001, 142, 4522–4528. [Google Scholar] [CrossRef]
- Jang, H.-J.; Kokrashvili, Z.; Theodorakis, M.J.; Carlson, O.D.; Kim, B.-J.; Zhou, J.; Kim, H.H.; Xu, X.; Chan, S.L.; Juhaszova, M.; et al. Gut-expressed gustducin and taste receptors regulate secretion of glucagon-like peptide-1. Proc. Natl. Acad. Sci. USA 2007, 104, 15069–15074. [Google Scholar] [CrossRef]
- Larraufie, P.; Martin-Gallausiaux, C.; Lapaque, N.; Dore, J.; Gribble, F.M.; Reimann, F.; Blottiere, H.M. SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 2018, 8, 1–9. [Google Scholar] [CrossRef]
- McCarthy, T.; Green, B.D.; Calderwood, D.; Gillespie, A.; Cryan, J.F.; Giblin, L. STC-1 Cells. In The Impact of Food Bioactives on Health; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 211–220. ISBN 978-3-319-16104-4. [Google Scholar]
- Rindi, G.; Grant, S.G.; Yiangou, Y.; Ghatei, M.A.; Bloom, S.R.; Bautch, V.L.; Solcia, E.; Polak, J.M. Development of neuroendocrine tumors in the gastrointestinal tract of transgenic mice. {Heterogeneity} of hormone expression. Am. J. Pathol. 1990, 136, 1349–1363. [Google Scholar]
- Gil-Lozano, M.; Brubaker, P.L. Murine GLUTag Cells. In The Impact of Food Bioactives on Health; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 229–238. [Google Scholar]
- Brubaker, P.L.; Drucker, D.J.; Greenberg, G.R. Synthesis and secretion of somatostatin-28 and -14 by fetal rat intestinal cells in culture. Am. J. Physiol. Liver Physiol. 1990, 258, G974–G981. [Google Scholar] [CrossRef]
- Drucker, D.J.; Jin, T.; Asa, S.L.; Young, T.A.; Brubaker, P.L. Activation of proglucagon gene transcription by protein kinase-A in a novel mouse enteroendocrine cell line. Mol. Endocrinol. 1994, 8, 1646–1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Khan, W. Goblet cells and Mucins: Role in innate defense in enteric infections. Pathogens 2013, 2, 55–70. [Google Scholar] [CrossRef]
- Xiong, W.; Ma, H.; Zhang, Z.; Jin, M.; Wang, J.; Xu, Y.; Wang, Z. The protective effect of icariin and phosphorylated icariin against LPS-induced intestinal goblet cell dysfunction. Innate Immun. 2020, 26, 97–106. [Google Scholar] [CrossRef]
- Klinkspoor, J.H.; Mok, K.S.; Van Klinken, B.J.-W.; Tytgat, G.N.J.; Lee, S.P.; Groen, A.K. Mucin secretion by the human colon cell line LS174T is regulated by bile salts. Glycobiology 1999, 9, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Bu, X.-D.; Li, N.; Tian, X.-Q.; Huang, P.-L. Caco-2 and LS174T cell lines provide different models for studying mucin expression in colon cancer. Tissue Cell 2011, 43, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Corridoni, D.; Pastorelli, L.; Mattioli, B.; Locovei, S.; Ishikawa, D.; Arseneau, K.O.; Chieppa, M.; Cominelli, F.; Pizarro, T.T. Probiotic Bacteria Regulate Intestinal Epithelial Permeability in Experimental Ileitis by a TNF-Dependent Mechanism. PLoS ONE 2012, 7, e42067. [Google Scholar] [CrossRef]
- Navabi, N.; McGuckin, M.A.; Lindén, S.K. Gastrointestinal cell lines form polarized epithelia with an adherent mucus layer when cultured in Semi-wet interfaces with mechanical stimulation. PLoS ONE 2013, 8, e68761. [Google Scholar] [CrossRef]
- Clevers, H.C.; Bevins, C.L. Paneth cells: Maestros of the small intestinal crypts. Annu. Rev. Physiol. 2013, 75, 289–311. [Google Scholar] [CrossRef]
- Sato, T.; van Es, J.H.; Snippert, H.J.; Stange, D.E.; Vries, R.G.; van den Born, M.; Barker, N.; Shroyer, N.F.; van de Wetering, M.; Clevers, H. Paneth cells constitute the niche for Lgr5 stem cells in intestinal crypts. Nature 2011, 469, 415–418. [Google Scholar] [CrossRef]
- Jones, C.; Avino, M.; Giroux, V.; Boudreau, F. HNF4α acts as upstream functional regulator of intestinal Wnt3 and paneth cell fate. Cell. Mol. Gastroenterol. Hepatol. 2023, 15, 593–612. [Google Scholar] [CrossRef] [PubMed]
- Adriaenssens, A.E.; Reimann, F.; Gribble, F.M. Distribution and Stimulus Secretion Coupling of Enteroendocrine Cells along the Intestinal Tract. In Comprehensive Physiology; Wiley: Hoboken, NJ, USA, 2018; pp. 1603–1638. [Google Scholar]
- Fitzgerald, K.A.; Malhotra, M.; Curtin, C.M.; O’ Brien, F.J.; O’ Driscoll, C.M. Life in 3D is never flat: 3D models to optimise drug delivery. J. Control. Release 2015, 215, 39–54. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.-M.; Elisia, I.; Kitts, D.D. Defining conditions for the co-culture of Caco-2 and HT29-MTX cells using Taguchi design. J. Pharmacol. Toxicol. Methods 2010, 61, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Nollevaux, G.; Devillé, C.; El Moualij, B.; Zorzi, W.; Deloyer, P.; Schneider, Y.-J.; Peulen, O.; Dandrifosse, G. Development of a serum-free co-culture of human intestinal epithelium cell-lines (Caco-2/HT29-5M21). BMC Cell Biol. 2006, 7, 20. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.G. Organophosphorus compounds at 80: Some old and new issues. Toxicol. Sci. 2018, 162, 24–35. [Google Scholar] [CrossRef]
- Murata, M. Browning and pigmentation in food through the Maillard reaction. Glycoconj. J. 2021, 38, 283–292. [Google Scholar] [CrossRef]
- Van der Post, C.R.S.; Carneiro, F. Goblet Cells. In Pathology of the Gastrointestinal Tract. Encyclopedia of Pathology; Carneiro, F., Chaves, P., Ensari, A., Eds.; Springer: Cham, Switzerland, 2017; pp. 316–319. [Google Scholar]
- Kwak, Y.-K.; Vikström, E.; Magnusson, K.-E.; Vécsey-Semjén, B.; Colque-Navarro, P.; Möllby, R. The Staphylococcus aureus alpha-toxin perturbs the barrier function in Caco-2 epithelial cell monolayers by altering junctional integrity. Infect. Immun. 2012, 80, 1670–1680. [Google Scholar] [CrossRef]
- Gagnon, M.; Zihler Berner, A.; Chervet, N.; Chassard, C.; Lacroix, C. Comparison of the Caco-2, HT-29 and the mucus-secreting HT29-MTX intestinal cell models to investigate Salmonella adhesion and invasion. J. Microbiol. Methods 2013, 94, 274–279. [Google Scholar] [CrossRef] [PubMed]
- Behrens, I.; Stenberg, P.; Artursson, P.; Kissel, T. Transport of lipophilic drug molecules in a new mucus-secreting cell culture model based on HT29-MTX cells. Pharm. Res. 2001, 18, 1138–1145. [Google Scholar] [CrossRef]
- Kaulmann, A.; André, C.M.; Schneider, Y.-J.; Hoffmann, L.; Bohn, T. Carotenoid and polyphenol bioaccessibility and cellular uptake from plum and cabbage varieties. Food Chem. 2016, 197, 325–332. [Google Scholar] [CrossRef]
- Selby-Pham, S.N.B.; Osborne, S.A.; Howell, K.S.; Dunshea, F.R.; Bennett, L.E. Transport rates of dietary phytochemicals in cell monolayers is inversely correlated with absorption kinetics in humans. J. Funct. Foods 2017, 39, 206–214. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Tom, B.H.; Rutzky, L.P.; Jakstys, M.M.; Oyasu, R.; Kaye, C.I.; Kahan, B.D. Human colonic adenocarcinoma cells. In Vitro 1976, 12, 180–191. [Google Scholar] [CrossRef]
- Van Klinken, B.J.-W.; Oussoren, E.; Weenink, J.-J.; Strous, G.J.; Büller, H.A.; Dekker, J.; Einerhand, A.W.C. The human intestinal cell lines Caco-2 and LS174T as models to study cell-type specific mucin expression. Glycoconj. J. 1996, 13, 757–768. [Google Scholar] [CrossRef]
- Damiano, S.; Sasso, A.; De Felice, B.; Di Gregorio, I.; La Rosa, G.; Lupoli, G.A.; Belfiore, A.; Mondola, P.; Santillo, M. Quercetin increases MUC2 and MUC5AC gene expression and secretion in intestinal goblet cell-like LS174T via PLC/PKCα/ERK1-2 pathway. Front. Physiol. 2018, 9, 357. [Google Scholar] [CrossRef]
- Béduneau, A.; Tempesta, C.; Fimbel, S.; Pellequer, Y.; Jannin, V.; Demarne, F.; Lamprecht, A. A tunable Caco-2/HT29-MTX co-culture model mimicking variable permeabilities of the human intestine obtained by an original seeding procedure. Eur. J. Pharm. Biopharm. 2014, 87, 290–298. [Google Scholar] [CrossRef]
- Kernéis, S.; Caliot, E.; Stubbe, H.; Bogdanova, A.; Kraehenbuhl, J.-P.; Pringault, E. Molecular studies of the intestinal mucosal barrier physiopathology using cocultures of epithelial and immune cells: A technical update. Microbes Infect. 2000, 2, 1119–1124. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Berghaus, L.J.; Moore, J.N.; Hurley, D.J.; Vandenplas, M.L.; Fortes, B.P.; Wolfert, M.A.; Boons, G.-J. Innate immune responses of primary murine macrophage-lineage cells and RAW 264.7 cells to ligands of toll-like receptors 2, 3, and 4. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 443–454. [Google Scholar] [CrossRef]
- Lee, J.M.; Yim, M.-J.; Kim, H.-S.; Ko, S.-C.; Kim, J.-Y.; Oh, G.-W.; Baek, K.; Lee, D.-S. Antioxidant, anti-inflammatory, and antibacterial activities of a 70% ethanol-Symphyocladia linearis extract. Fish. Aquat. Sci. 2022, 25, 579–586. [Google Scholar] [CrossRef]
- Chanput, W.; Peters, V.; Wichers, H. THP-1 and U937 Cells. In The Impact of Food Bioactives on Health; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 147–159. ISBN 978-3-319-16104-4. [Google Scholar]
- García-Rodríguez, A.; Vila, L.; Cortés, C.; Hernández, A.; Marcos, R. Exploring the usefulness of the complex in vitro intestinal epithelial model Caco-2/HT29/Raji-B in nanotoxicology. Food Chem. Toxicol. 2018, 113, 162–170. [Google Scholar] [CrossRef] [PubMed]
- Crotty Alexander, L.E.; Marsh, B.J.; Timmer, A.M.; Lin, A.E.; Zainabadi, K.; Czopik, A.; Guarente, L.; Nizet, V. Myeloid Cell Sirtuin-1 Expression Does Not Alter Host Immune Responses to Gram-Negative Endotoxemia or Gram-Positive Bacterial Infection. PLoS ONE 2013, 8, e84481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ibañez, P.; Núñez-Sánchez, M.A.; Oliva-Bolarín, A.; Martínez-Sánchez, M.A.; Ramos-Molina, B.; Ruiz-Alcaraz, A.J.; Moreno, D.A. Anti-inflammatory potential of digested Brassica sprout extracts in human macrophage-like HL-60 cells. Food Funct. 2023, 14, 112–121. [Google Scholar] [CrossRef]
- Kämpfer, A.A.M.; Urbán, P.; Gioria, S.; Kanase, N.; Stone, V.; Kinsner-Ovaskainen, A. Development of an in vitro co-culture model to mimic the human intestine in healthy and diseased state. Toxicol. Vitr. 2017, 45, 31–43. [Google Scholar] [CrossRef]
- Smythies, L.E.; Sellers, M.; Clements, R.H.; Mosteller-Barnum, M.; Meng, G.; Benjamin, W.H.; Orenstein, J.M.; Smith, P.D. Human intestinal macrophages display profound inflammatory anergy despite avid phagocytic and bacteriocidal activity. J. Clin. Investig. 2005, 115, 66–75. [Google Scholar] [CrossRef]
- Kaulmann, A.; Legay, S.; Schneider, Y.-J.; Hoffmann, L.; Bohn, T. Inflammation related responses of intestinal cells to plum and cabbage digesta with differential carotenoid and polyphenol profiles following simulated gastrointestinal digestion. Mol. Nutr. Food Res. 2016, 60, 992–1005. [Google Scholar] [CrossRef] [PubMed]
- Susewind, J.; de Souza Carvalho-Wodarz, C.; Repnik, U.; Collnot, E.-M.; Schneider-Daum, N.; Griffiths, G.W.; Lehr, C.-M. A 3D co-culture of three human cell lines to model the inflamed intestinal mucosa for safety testing of nanomaterials. Nanotoxicology 2016, 10, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Wang, D.; Sui, Z.; Qi, X.; Ji, L.; Wang, X.; Yang, L. Development of an improved three-dimensional in vitro intestinal mucosa model for drug absorption evaluation. Tissue Eng. Part C Methods 2013, 19, 708–719. [Google Scholar] [CrossRef]
- Kim, K.-M.; Kim, Y.-S.; Lim, J.Y.; Min, S.J.; Ko, H.-C.; Kim, S.-J.; Kim, Y. Intestinal anti-inflammatory activity of Sasa quelpaertensis leaf extract by suppressing lipopolysaccharide-stimulated inflammatory mediators in intestinal epithelial Caco-2 cells co-cultured with RAW 264.7 macrophage cells. Nutr. Res. Pract. 2015, 9, 3–10. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Yamabe, M.; Yamaguchi, Y.; Kobayashi, Y.; Konno, T.; Tada, K. Establishment and characterization of a human acute monocytic leukemia cell line (THP-1). Int. J. Cancer 1980, 26, 171–176. [Google Scholar] [CrossRef]
- Taciak, B.; Białasek, M.; Braniewska, A.; Sas, Z.; Sawicka, P.; Kiraga, Ł.; Rygiel, T.; Król, M. Evaluation of phenotypic and functional stability of RAW 264.7 cell line through serial passages. PLoS ONE 2018, 13, e0198943. [Google Scholar] [CrossRef]
- Tanoue, T.; Nishitani, Y.; Kanazawa, K.; Hashimoto, T.; Mizuno, M. In vitro model to estimate gut inflammation using co-cultured Caco-2 and RAW264.7 cells. Biochem. Biophys. Res. Commun. 2008, 374, 565–569. [Google Scholar] [CrossRef] [Green Version]
- Gullberg, E.; Leonard, M.; Karlsson, J.; Hopkins, A.M.; Brayden, D.; Baird, A.W.; Artursson, P. Expression of specific markers and particle transport in a new human intestinal M-cell model. Biochem. Biophys. Res. Commun. 2000, 279, 808–813. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.; Park, H.-E.; Soh, S.H.; Im, Y.B.; Yoo, H.S. Induction of Th2 response through TLR2-mediated MyD88-dependent pathway in human microfold cells stimulated with chitosan nanoparticles loaded with Brucella abortus Mdh. Microb. Pathog. 2020, 142, 104040. [Google Scholar] [CrossRef]
- Tonry, J.H.; Popov, S.G.; Narayanan, A.; Kashanchi, F.; Hakami, R.M.; Carpenter, C.; Bailey, C.; Chung, M.-C. In vivo murine and in vitro M-like cell models of gastrointestinal anthrax. Microbes Infect. 2013, 15, 37–44. [Google Scholar] [CrossRef]
- Kesisoglou, F.; Schmiedlin-Ren, P.; Fleisher, D.; Zimmermann, E.M. Adenoviral transduction of enterocytes and M-Cells using in vitro models based on Caco-2 cells: The Coxsackievirus and Adenovirus Receptor (CAR) mediates both apical and basolateral transduction. Mol. Pharm. 2010, 7, 619–629. [Google Scholar] [CrossRef] [PubMed]
- des Rieux, A.; Fievez, V.; Théate, I.; Mast, J.; Préat, V.; Schneider, Y.-J. An improved in vitro model of human intestinal follicle-associated epithelium to study nanoparticle transport by M cells. Eur. J. Pharm. Sci. 2007, 30, 380–391. [Google Scholar] [CrossRef] [PubMed]
- Corr, S.; Hill, C.; Gahan, C.G.M. An in vitro cell-culture model demonstrates internalin- and hemolysin-independent translocation of Listeria monocytogenes across M-cells. Microb. Pathog. 2006, 41, 241–250. [Google Scholar] [CrossRef]
- Masuda, K.; Kajikawa, A.; Igimi, S. Establishment and evaluation of an in vitro M Cell model using C2BBe1 cells and Raji cells. Biosci. Microflora 2011, 30, 37–44. [Google Scholar] [CrossRef]
- Ahmad, T.; Gogarty, M.; Walsh, E.G.; Brayden, D.J. A comparison of three Peyer’s patch “M-like” cell culture models: Particle uptake, bacterial interaction, and epithelial histology. Eur. J. Pharm. Biopharm. 2017, 119, 426–436. [Google Scholar] [CrossRef]
- Schimpel, C.; Teubl, B.; Absenger, M.; Meindl, C.; Fröhlich, E.; Leitinger, G.; Zimmer, A.; Roblegg, E. Development of an advanced intestinal in vitro triple culture permeability model to study transport of nanoparticles. Mol. Pharm. 2014, 11, 808–818. [Google Scholar] [CrossRef]
- Antunes, F.; Andrade, F.; Araújo, F.; Ferreira, D.; Sarmento, B. Establishment of a triple co-culture in vitro cell models to study intestinal absorption of peptide drugs. Eur. J. Pharm. Biopharm. 2013, 83, 427–435. [Google Scholar] [CrossRef]
- Araújo, F.; Pereira, C.; Costa, J.; Barrias, C.; Granja, P.L.; Sarmento, B. In vitro M-like cells genesis through a tissue-engineered triple-culture intestinal model. J. Biomed. Mater. Res. Part B Appl. Biomater. 2016, 104, 782–788. [Google Scholar] [CrossRef]
- Araújo, F.; Sarmento, B. Towards the characterization of an in vitro triple co-culture intestine cell model for permeability studies. Int. J. Pharm. 2013, 458, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, S.; Devlin, A. Intestinal co-culture system to study TGR5 agonism and gut restriction. Bio-Protocol 2021, 11, e3948. [Google Scholar] [CrossRef]
- Tang, S.-C.; Sambanis, A. Differential rAAV2 transduction efficiencies and insulin secretion profiles in pure and co-culture models of human enteroendocrine L-cells and enterocytes. J. Gene Med. 2004, 6, 1003–1013. [Google Scholar] [CrossRef]
- Gautier, T.; Fahet, N.; Tamanai-Shacoori, Z.; Oliviero, N.; Blot, M.; Sauvager, A.; Burel, A.; Gall, S.D.-L.; Tomasi, S.; Blat, S.; et al. Roseburia intestinalis modulates PYY expression in a new a multicellular model including enteroendocrine cells. Microorganisms 2022, 10, 2263. [Google Scholar] [CrossRef] [PubMed]
- Alminger, M.; Svelander, C.; Wellner, A.; Martinez-Tomas, R.; Bialek, L.; Larque, E.; Perez-Llamas, F. Applicability of in vitro models in predicting the in vivo bioavailability of lycopene and β-carotene from differently processed soups. Food Nutr. Sci. 2012, 3, 477–489. [Google Scholar] [CrossRef]
- Wu, T.; Grootaert, C.; Voorspoels, S.; Jacobs, G.; Pitart, J.; Kamiloglu, S.; Possemiers, S.; Heinonen, M.; Kardum, N.; Glibetic, M.; et al. Aronia (Aronia melanocarpa) phenolics bioavailability in a combined in vitro digestion/Caco-2 cell model is structure and colon region dependent. J. Funct. Foods 2017, 38, 128–139. [Google Scholar] [CrossRef]
- Déat, E.; Blanquet-Diot, S.; Jarrige, J.-F.; Denis, S.; Beyssac, E.; Alric, M. Combining the Dynamic TNO-gastrointestinal tract system with a Caco-2 cell culture model: Application to the assessment of lycopene and α-tocopherol bioavailability from a whole food. J. Agric. Food Chem. 2009, 57, 11314–11320. [Google Scholar] [CrossRef]
- Ye, D.; López Mármol, Á.; Lenz, V.; Muschong, P.; Wilhelm-Alkubaisi, A.; Weinheimer, M.; Koziolek, M.; Sauer, K.A.; Laplanche, L.; Mezler, M. Mucin-Protected Caco-2 Assay to Study Drug Permeation in the Presence of Complex Biorelevant Media. Pharmaceutics 2022, 14, 699. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, R.; Vissenaekens, H.; Pitart, J.; Romo-Vaquero, M.; Espín, J.C.; Grootaert, C.; Selma, M.V.; Raes, K.; Smagghe, G.; Possemiers, S.; et al. Gastrointestinal simulation model TWIN-SHIME shows differences between human urolithin-metabotypes in gut microbiota composition, pomegranate polyphenol metabolism, and transport along the intestinal tract. J. Agric. Food Chem. 2017, 65, 5480–5493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kortman, G.A.M.; Dutilh, B.E.; Maathuis, A.J.H.; Engelke, U.F.; Boekhorst, J.; Keegan, K.P.; Nielsen, F.G.G.; Betley, J.; Weir, J.C.; Kingsbury, Z.; et al. Microbial metabolism shifts towards an adverse profile with supplementary iron in the TIM-2 In vitro model of the Human colon. Front. Microbiol. 2016, 6, 1481. [Google Scholar] [CrossRef] [PubMed]
- Réquilé, M.; Gonzàlez Alvarez, D.O.; Delanaud, S.; Rhazi, L.; Bach, V.; Depeint, F.; Khorsi-Cauet, H. Use of a combination of in vitro models to investigate the impact of chlorpyrifos and inulin on the intestinal microbiota and the permeability of the intestinal mucosa. Environ. Sci. Pollut. Res. 2018, 25, 22529–22540. [Google Scholar] [CrossRef] [PubMed]
- Wu, T. Bioavailability and Bioactivity of Aronia polyphenols Using In Vitro Digestion Models Combined with a Caco-2/Endothelial Cell Co-Culture Model; Ghent University: Ghent, Belgium, 2018. [Google Scholar]
- Banskota, S.; Regmi, S.C.; Gautam, J.; Gurung, P.; Lee, Y.-J.; Ku, S.K.; Lee, J.-H.; Lee, J.; Chang, H.W.; Park, S.J.; et al. Serotonin disturbs colon epithelial tolerance of commensal E. coli by increasing NOX2-derived superoxide. Free Radic. Biol. Med. 2017, 106, 196–207. [Google Scholar] [CrossRef]
- Barnett, A.; Roy, N.; Cookson, A.; McNabb, W. Metabolism of caprine milk carbohydrates by probiotic bacteria and Caco-2:HT29–MTX epithelial co-cultures and their impact on intestinal barrier Integrity. Nutrients 2018, 10, 949. [Google Scholar] [CrossRef]
- Hsieh, C.-Y.; Osaka, T.; Moriyama, E.; Date, Y.; Kikuchi, J.; Tsuneda, S. Strengthening of the intestinal epithelial tight junction by Bifidobacterium bifidum. Physiol. Rep. 2015, 3, e12327. [Google Scholar] [CrossRef]
- Burkholder, K.M.; Fletcher, D.H.; Gileau, L.; Kandolo, A. Lactic acid bacteria decrease Salmonella enterica Javiana virulence and modulate host inflammation during infection of an intestinal epithelial cell line. Pathog. Dis. 2019, 77, ftz025. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Prete, R.; Battista, N.; Corsetti, A. Adhesion properties of food-associated Lactobacillus plantarum strains on human intestinal epithelial cells and modulation of IL-8 release. Front. Microbiol. 2018, 9, 2392. [Google Scholar] [CrossRef]
- Rocha-Mendoza, D.; Kosmerl, E.; Miyagusuku-Cruzado, G.; Giusti, M.M.; Jiménez-Flores, R.; García-Cano, I. Growth of lactic acid bacteria in milk phospholipids enhances their adhesion to Caco-2 cells. J. Dairy Sci. 2020, 103, 7707–7718. [Google Scholar] [CrossRef]
- Garriga, M.; Rubio, R.; Aymerich, T.; Ruas-Madiedo, P. Potentially probiotic and bioprotective lactic acid bacteria starter cultures antagonise the Listeria monocytogenes adhesion to HT29 colonocyte-like cells. Benef. Microbes 2015, 6, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.; Sharma, P.; Kalia, N.; Singh, J.; Kaur, S. Anti-biofilm properties of the fecal probiotic Lactobacilli against Vibrio spp. Front. Cell. Infect. Microbiol. 2018, 8, 120. [Google Scholar] [CrossRef] [Green Version]
- Beatty, J.K.; Akierman, S.V.; Motta, J.-P.; Muise, S.; Workentine, M.L.; Harrison, J.J.; Bhargava, A.; Beck, P.L.; Rioux, K.P.; McKnight, G.W.; et al. Giardia duodenalis induces pathogenic dysbiosis of human intestinal microbiota biofilms. Int. J. Parasitol. 2017, 47, 311–326. [Google Scholar] [CrossRef]
- Shil, A.; Chichger, H. Artificial sweeteners negatively regulate pathogenic characteristics of two model gut bacteria, E. coli and E. faecalis. Int. J. Mol. Sci. 2021, 22, 5228. [Google Scholar] [CrossRef] [PubMed]
- Beamer, M.A.; Zamora, C.; Kalinoski, A.L.N.; Fernando, V.; Sharma, V.; Furuta, S. Novel 3D Flipwell system that models gut mucosal microenvironment for studying interactions between gut microbiota, epithelia and immunity. Sci. Rep. 2023, 13, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Clark, E.C. Glutamine deprivation facilitates tumour necrosis factor induced bacterial translocation in Caco-2 cells by depletion of enterocyte fuel substrate. Gut 2003, 52, 224–230. [Google Scholar] [CrossRef] [PubMed]
- Overman, A.; Bumrungpert, A.; Kennedy, A.; Martinez, K.; Chuang, C.-C.; West, T.; Dawson, B.; Jia, W.; McIntosh, M. Polyphenol-rich grape powder extract (GPE) attenuates inflammation in human macrophages and in human adipocytes exposed to macrophage-conditioned media. Int. J. Obes. 2010, 34, 800–808. [Google Scholar] [CrossRef]
- Overman, A.; Chuang, C.-C.; McIntosh, M. Quercetin attenuates inflammation in human macrophages and adipocytes exposed to macrophage-conditioned media. Int. J. Obes. 2011, 35, 1165–1172. [Google Scholar] [CrossRef]
- Gillies, P.J.; Bhatia, S.K.; Belcher, L.A.; Hannon, D.B.; Thompson, J.T.; Vanden Heuvel, J.P. Regulation of inflammatory and lipid metabolism genes by eicosapentaenoic acid-rich oil. J. Lipid Res. 2012, 53, 1679–1689. [Google Scholar] [CrossRef]
- Jabaud, J.; Ckless, K. Inflammation, immunity and redox signaling—Chapter 7. In Inflammation, Chronic Diseases and Cancer: Cell and Molecular Biology, Immunology and Clinical Bases; BoD—Books on Demand: Rijeka, Croatia, 2012; ISBN 978-953-51-0102-4. [Google Scholar]
- Neustock, P.; Brand, J.M.; Kruse, A.; Kirchner, H. Cytokine production of the human monocytic cell line Mono Mac 6 in comparison to mature monocytes in peripheral blood mononuclear cells. Immunobiology 1993, 188, 293–302. [Google Scholar] [CrossRef]
- Coluccia, A.; Matti, F.; Zhu, X.; Lussi, A.; Stähli, A.; Sculean, A.; Eick, S. In vitro study on green propolis as a potential ingredient of oral health care products. Antibiotics 2022, 11, 1764. [Google Scholar] [CrossRef] [PubMed]
- Plantinga, M.; de Haar, C.; Nierkens, S. Dendritic Cells. In The Impact of Food Bioactives on Health; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 181–196. ISBN 978-3-319-15791-7. [Google Scholar]
- Koeffler, H.P.; Bar-Eli, M.; Territo, M.C. Phorbol ester effect on differentiation of human myeloid leukemia cell lines blocked at different stages of maturation. Cancer Res. 1981, 41, 919–926. [Google Scholar] [PubMed]
- Plante, A.M.; McCarthy, A.L.; Lacey, S.; O’Halloran, F. Investigating the bioactive properties of cheese-fruit combinations following in vitro digestion using an elderly model. Curr. Res. Nutr. Food Sci. J. 2021, 9, 465–478. [Google Scholar] [CrossRef]
- Chen, J.L.; Nong, G.M. Advances in application of jurkat cell model in research on infectious diseases. Chin. J. Contemp. Pediatr. 2018, 20, 236–242. [Google Scholar] [CrossRef]
- Abraham, R.T.; Weiss, A. Jurkat T cells and development of the T-cell receptor signalling paradigm. Nat. Rev. Immunol. 2004, 4, 301–308. [Google Scholar] [CrossRef]
- Devriese, S.; Van den Bossche, L.; Van Welden, S.; Holvoet, T.; Pinheiro, I.; Hindryckx, P.; De Vos, M.; Laukens, D. T84 monolayers are superior to Caco-2 as a model system of colonocytes. Histochem. Cell Biol. 2017, 148, 85–93. [Google Scholar] [CrossRef]
- Pizarroso, N.; Fuciños, P.; Gonçalves, C.; Pastrana, L.; Amado, I. A review on the role of food-derived bioactive molecules and the microbiota–gut–brain axis in satiety regulation. Nutrients 2021, 13, 632. [Google Scholar] [CrossRef]
- Kauffman, A.L.; Gyurdieva, A.V.; Mabus, J.R.; Ferguson, C.; Yan, Z.; Hornby, P.J. Alternative functional in vitro models of human intestinal epithelia. Front. Pharmacol. 2013, 4, 79. [Google Scholar] [CrossRef]
- Jochems, P.; Garssen, J.; van Keulen, A.; Masereeuw, R.; Jeurink, P. Evaluating human intestinal cell lines for studying dietary protein absorption. Nutrients 2018, 10, 322. [Google Scholar] [CrossRef]
- Park, N.-Y.; Koh, A. From the Dish to the Real World: Modeling Interactions between the Gut and Microorganisms in Gut Organoids by Tailoring the Gut Milieu. Int. J. Stem Cells 2022, 15, 70–84. [Google Scholar] [CrossRef]
- Co, J.Y.; Margalef-Català, M.; Li, X.; Mah, A.T.; Kuo, C.J.; Monack, D.M.; Amieva, M.R. Controlling Epithelial Polarity: A Human Enteroid Model for Host-Pathogen Interactions. Cell Rep. 2019, 26, 2509–2520. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yang, N.; Chen, J.; Huang, X.; Zhang, N.; Yang, S.; Liu, G.; Liu, G. Next-Generation Porcine Intestinal Organoids: An Apical-Out Organoid Model for Swine Enteric Virus Infection and Immune Response Investigations. J. Virol. 2020, 94, e01006-20. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Cell Line | Origins | Culture Conditions | Use | Specificities | Reference | |
|---|---|---|---|---|---|---|
| Enterocytes | Caco-2 (HBT-37) | Human colorectal adenocarcinoma | RPMI/DMEM + 10% FBS maintenance 1:10 dilution upon reaching 80% confluence culture on inserts: 4 × 105 cells/cm2 spontaneous differentiation when reaching confluence (14–21 d) | Permeability, absorption, transepithelial transport after oral intake | Apical brush border with microvilli and tight junctions brush border activity: lactase, aminopeptidase N, sucrase isomaltase, dipeptidylpeptidase IV No mucus secretion Molecular transport: amino acids, sugars (GLUT1, GLUT2, GLUT3, GLUT5, SGLT1), vitamins (B12, D3), hormones, xenobiotics (P-gp, MRP), but low PepT1 Drug metabolization: aminopeptidase, esterase, sulfatase, glucuronosyltransferase, CYP1A1 or CYP1B1 but not CYP3A4 or CYP2B6 Inflammation: IL6, IL8, TNFa, TGFb1, TLSP, IL15, low TLR-4 | [27,28,29,30,31,32,33] |
| Caco-2/TC7 | Clone of Caco-2 from late passage (198) | DMEM 10% FBSDMEM + HEPES + 10% FBS low glucose consumer, differentiation faster than parental line (20 d vs. 30 d), homogeneous population | Intestinal drug permeability and absorption studies | Well-organized brush border and high sucrase isomaltase, DPP-IV TLR2 + TLR4 Molecular transport: SGLT1, GLUT2, GLUT5 remain high but GLUT1 and GLUT3 decrease after confluence, good P-gp Drug metabolization: increased DMT1, glucuronosyltransferases, low transferrin receptors, CYP1A1 but no CYP3A4 (except MTX-induced) | [27,34,35,36,37,38,39] | |
| Caco-2/C2BBE1 | Clone of Caco-2 | Glutamin-induced carcinogenesis | Good level of sucrase isomaltase, well-developed brush border, all microvillar proteins expressed including myosin I Molecular transport: folate receptor, vit C channel | [27] | ||
| Caco-2/15 | Clone of Caco-2 vit D3-sensititve | Inhibition of cell growth in response to vit D3 High sucrase isomaltase activity | [27] | |||
| HT29 | Human colon adenocarcinoma | DMEM + 10% FBS McCoy’s 5A + 10% FBS; differentiation in glucose-free medium very similar to Caco-2 after 30 d | Cell differentiation, food digestion and bioavailability Drugs and food transport Intestinal immune response to bacterial infection, microorganism survival, adhesion or invasion | Considered a pluripotent intestinal cell line Brush border: aminopeptidase N, dipeptidyl peptidase-IV and alkaline phosphatase and villin, low sucrase isomaltase, no lactase MUC1 expression except at late confluency Cytokines: TNFα, IL1β, IL6, IL8, IL15, VEGF, GCF, GMCF, IL3 No PYY and NPY receptors MCT1 transport Impaired glucose metabolism; brush-border-associated hydrolases | [30,37,38,39,40,41,42] | |
| HT29-18C | Clone of HT29- | DMEM + 10% FBS + HEPES | Linear model for paracellular transport route close to ileum | Slow acquisition of electrical resistance over 24 days on inserts, polarization but poorly developped microvillus membrane | [43,44] | |
| HIEC-6 | Human fetal cell line small intestine, normal | OptiMEM + HEPES + 4% FBS 8–10 d for differentiation | Physiological and pathophysiological conditions and interaction between intestinal epithelium and mycotoxins | Non-malignant Non-tumorigenic mimics in vivo condition better than Caco-2 PepT1 transporter Lipid-binding transport proteins iFABP | [29,45,46,47] | |
| tsFHIFHs-74-Int | Human fetal small intestine epithelial cells (transformed with SV40) | OptiMEM + HEPES + 4% FBS DMEM + HEPES + 7% FBS | Non-malignant At 37 °C, the SV40 antigen breaks down, leading to growth arrest and acquisition of an enterocyte-like phenotype, although they remain morphologically immature and do not form tight junctions or polarize | [48,49,50,51] | ||
| H4 | Human fetal primary intestine cell line | DMEM + 10% FBS DMEM + HEPES + 10% FBS + insulin | Inflammation response Physiopathology of necrotized enterocolitis Fatty acids transport and absorption studies | Non-malignant Highly tolerant Express more IL-8 than Caco-2 with and without LPS Not forming tight junctions or polarize | [48,52,53,54] | |
| CCD 841 Con = CRL1790 | Normal fetal human colon tissue | DMEM/MEM + 10% FBS MEM + 3% FBS | Inflammatory mechanisms in colonic epithelial cells | Non-malignant, no keratin Stretched epithelial-like cells growing as a flattened and disorganized layer in culture Secretion TNFα, IL1β, IL8 in response to bacterial treatment | [55,56] | |
| Colonocytes | SK-CO15 | Colorectal adenocarcinoma | DMEM + Ham’s F-12 + 10% FBS | Model for Na(+)/h(+) exchange Cell migration Epithelial cyst formation Epithelial barrier function Transport studies Regulation of epithelial junctions | Expression of NHE1, 2, 8 and 3 and of NHERF1 and NHERF2 Polarized distribution similar to original glycoprotein antigens in human colon High expression of NHE3 No expression of sucrase-isomaltase, aminopeptidase N, lactase, glucoamylase Absence of brush border, low alkaline-phosphatase and villin levels Few and disorganised microvilli | [57] |
| T84 | Lung metastasis colon carcinoma | DMEM/Ham’s F-12 + 10% FBS or 5% NBS | Hormonal control of colon cell growth Electrolytes transport Intestinal permeability Inflammatory response Bacteria/enterocyte interaction | Shorter microvilli Tight junction structure, basal CYP3A4 expression controversial but PXP-induced PgP efflux No IL8 secretion in response to LPS but ok after IL1β | [30,39,58,59] | |
| SW620 | Dukes C colorectal cancer patient, Stage 3 | DMEM/10%FBS, McCoy’s 5A + 10% FBS | Drug toxicity, cancer research | Keratin IL8 secretion in response to LPS and IL1β Lower expression of MUC2, MUC6 and MUC5B with high expression of MUC1 | [60,61,62,63] | |
| Colo-205 | Ascitic fluid derived from colon cancer Dukes’ type D | RPMI-1640 + 10% FBS | Cancer research and toxicology | Keratin It has mixed growth properties: adherent and suspension | [62,64] | |
| HCT116 | Colon cancer Stage 1 (primary) | RPMI + 10% FBS DMEM + 10% FBS McCoy’s 5A + 10% FBS | Cancer research and toxicology | Expression of apoptosis genes: Grp78, Bcl-2, NF-kappaB(p50), NF-kappaB(p65), thioredoxin peroxidase (peroxiredoxin) 2, peroxiredoxin 4, maspin, guanylate cyclase activating protein-1, PKCzeta, EGFR, Ras family members, PKA, PI(4,5)K, TRAF2 and BIRC1 (IAP protein) Mutation in codon 13 of the ras-proto-oncogene | [60,64,65,66] | |
| HRT18 | Large intestine adenocarcinoma | RPMI-+ 10% horse serum | Cancer research and toxicology | Identical to HCT-8 cell line Epitheloid morphotype | [67] | |
| FHC (CRL1831) | Human fetal colon cells | Ham’s F12 + DMEM, + HEPES +10% FBS, + cholera toxin + insulin, + transferrin + hydrocortisone | [68] | |||
| NCM460 | Human normal colonic epithelial cell line | M3 medium + 10% FBS | Intestinal barrier Model for transverse colonic crypts | Immortalized Proteasome activity; characteristics of native colonocytes Gastrin-releasing peptide receptor | [69,70,71,72,73] | |
| CCD-18co = CRL-1459 | Infant colon non-cancerous | DMEM/EMEM + 10% FBS | Cancer research and toxicology | Fibroblast morphology Senesce at about 42 PDL | [74] |
| Cell Lines | Origins | Culture Conditions | Use | Specificities | Reference | |
|---|---|---|---|---|---|---|
| Goblet cells | HT29-MTX | HT29 + stepwise adaptation with increased concentration of Methotrexate | DMEM + 10% FBS | Mucus-inducing properties of food compounds, microorganism adhesion survival, transport studies | MTX-induced differentiation into entirely differentiated goblet-cell-like phenotype Mucin: MUC1 and MUC5 but low MUC2 Passive diffusion CYP3A4 in apical localization | [35,39,85,86,87,88,89] |
| LS180 | Colon adenocarcinoma stage 2 (primary) | DMEM/Ham’s F12 + 10% FBS MEM 10% FBS | Mucin synthesis studies | Mucus-secreting cell line Expression of mucin Basal and inducible CYP3A4 activity but poor tight junction structure | [90] | |
| LS174T | Trypsined variant from LS180 | DMEM + 10% FBS | Chemical, physiological, pharmacologic and immunologic characteristics of neoplastic colonic cells | Abundant microvilli Intracytoplasmic mucin vacuoles High level of carcinoembryonic antigens High expression of MUC2, MUC6 and MUC5B with lower expression of MUC13 | [63] | |
| Enteroendocrine cell lines | NCI-H716 | Lymphoblast morphology isolated from colorectal adenocarcinoma | RPMI + 10% FBS-Need of Matrigel for adherence | Human intestinal L cell GLP-1 secretion assay in response to nutrients | Muscarinic receptors PYY and GLP-1 secretion | [91,92,93,94] |
| HuTu-80 | Duodenum cancerous | DMEM/F-12 + 10% FBS | Cancer research | The cells express receptors for bombesin at up to 6000 sites per cell PYY expression is reduced for butyrate or propionate stimulation | [95] | |
| STC-1 | Murine duodenal enteroendocrine tumor L-cell Duodenal proinsulin-polyoma (transformed SV40) | DMEM + 17.5% FBS | Representative of L-cell Used to study GLP-1 secretion | Enteroendocrine cell model Express and secrete CCK, PYY, pancreatic polypeptide, neurotensin, GLP-1, GLP-2, oxyntomodulin Poor secretion of GIP | [96,97] | |
| GLUTtag | Murine colon tumor expressing SV40 large T antigen under control of proglucagon promoter (transformed SV40) | DMEM + 10% FBS | GLP-1 secretion | GLUT and SGLT1 glucose transporters to stimulate GLUT secretion GPR40, GPR120, TGR5, GPR43 | [98,99,100] |
| Cell Line | Origins | Culture Conditions | Use | Specificities | References |
|---|---|---|---|---|---|
| THP-1 | Human monocytic leukemia | RPMI1640 + 10% FBS | Immune modulation and inflammation studies | PMA-induced differentiation into an adherent macrophage phenotype and further polarized into M1 and M2 macrophage cells; culture in serum-free medium + IL4, GM-CSF, TNFα and ionomycim to differentiate into dendritic-cell-expressing CD83, CD80, CD86, CD40, CD206, CD209, CD120a, CD120b + intracellular IL10 | [131] |
| RAW264.7 | Murine tumour induced by the Abelson leukaemia virus | RPMI/DMEM + FBS | Oxidative stress | Closely mimic bone-marrow-derived macrophages in terms of cell surface receptors and response to microbial ligands via TLRs IL12 NO | [129] |
| Raji | B-cell Human Burkitt’s Lymphoma | DMEM without Pyruvate + 10% FBS + plasmocin | Useful model for drug delivery studies | In co-culture, transform Caco-2 to reach morphological and functional features of M cell Fc receptors for binding immunoglobulin IL-10 TNF-α INF-γ lack of differentiation | [39,132] |
| HL-60 | Promyeoloblasts from peripheral blood with promyelocytic leukemia | RPMI-1640 + 10% FBS/DMSO 1,25% for neutrophilic differentiation | Immune disorder and immunology research | Pro-monocytic cell line differentiating into macrophage or dendritic cells in the presence of butyrate, hypoxanthine, phorbol myristic acid, DMSO, actinomycin D and retinoic acid stimulation; phagocytic activity, Fc expression | [133,134] |
| U937 | Human leukemia from histiocytic lymphoma | Differentiation into macrophages and mature monocytes with PMA-treatment (48h). Differentiating into DC-like cells by exposure to Ep1, B | Study of anti-inflammatory properties of food bioactive, monocyte–endothelium mechanism of attachment, monocyte–macrophage differentiation | Pro-monocytic cell line that can differentiate into macrophage or dendritic cells with more passage stability than THP-1 | [131] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haddad, M.J.; Sztupecki, W.; Delayre-Orthez, C.; Rhazi, L.; Barbezier, N.; Depeint, F.; Anton, P.M. Complexification of In Vitro Models of Intestinal Barriers, A True Challenge for a More Accurate Alternative Approach. Int. J. Mol. Sci. 2023, 24, 3595. https://doi.org/10.3390/ijms24043595
Haddad MJ, Sztupecki W, Delayre-Orthez C, Rhazi L, Barbezier N, Depeint F, Anton PM. Complexification of In Vitro Models of Intestinal Barriers, A True Challenge for a More Accurate Alternative Approach. International Journal of Molecular Sciences. 2023; 24(4):3595. https://doi.org/10.3390/ijms24043595
Chicago/Turabian StyleHaddad, Michelle J., Wendy Sztupecki, Carine Delayre-Orthez, Larbi Rhazi, Nicolas Barbezier, Flore Depeint, and Pauline M. Anton. 2023. "Complexification of In Vitro Models of Intestinal Barriers, A True Challenge for a More Accurate Alternative Approach" International Journal of Molecular Sciences 24, no. 4: 3595. https://doi.org/10.3390/ijms24043595