

Steroidogenic Factor 1, a Goldilocks Transcription Factor from Adrenocortical Organogenesis to Malignancy
 , , , and
, , , and
Abstract
:
1. Introduction
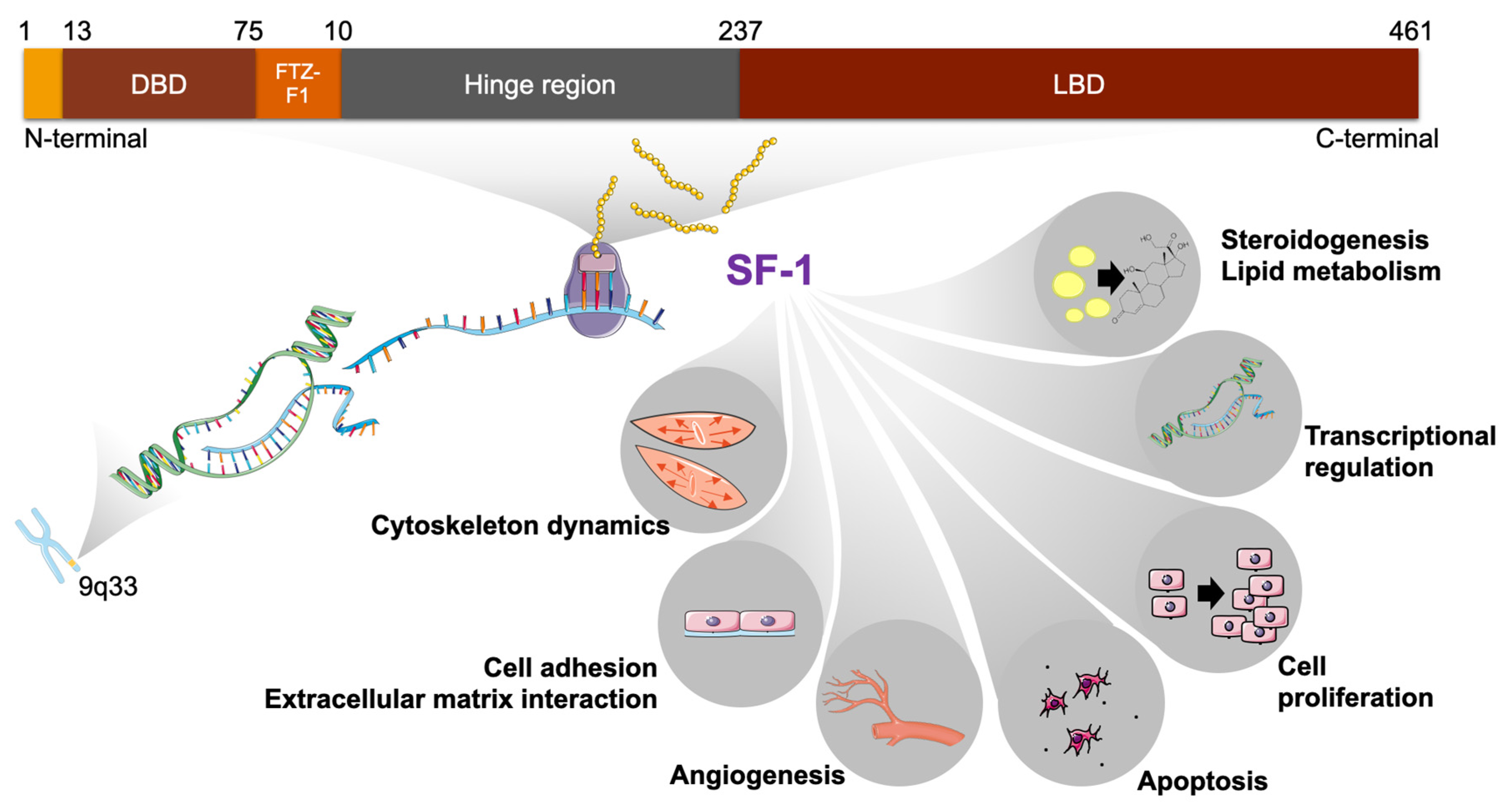
2. SF-1 Structure
3. SF-1 Expression
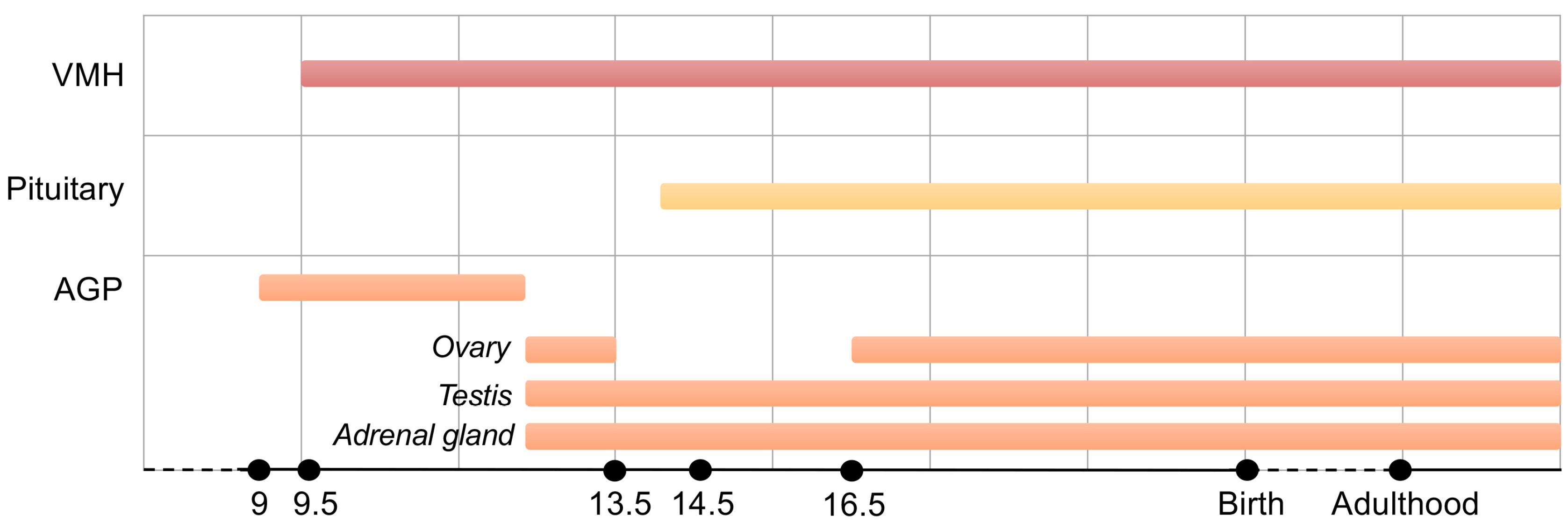
4. A Key Role for SF-1 in Adrenal Gland Organogenesis
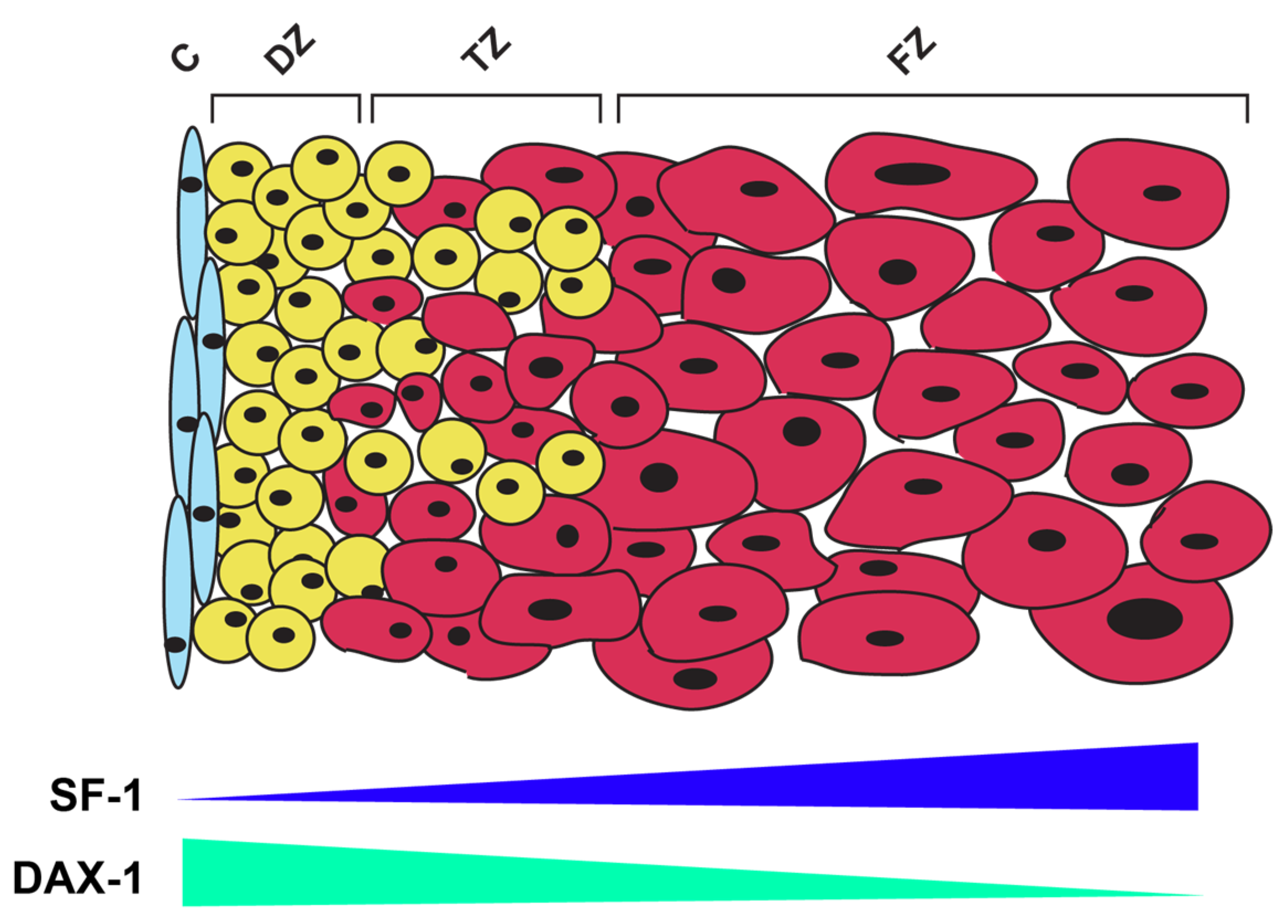
5. Regulation of SF-1 Expression and Activity in Adrenal Glands
6. SF-1 Dosage Is not Only Critical for Adrenal Development but also for Tumorigenesis
7. SF-1 Dosage-Dependent Target Genes: Their Roles in ACC and in Adrenal Gland Development
- -
- NOV/CCN3 encodes a secreted protein with pro-apoptotic properties which is a marker of the DZ in the human fetal adrenal cortex. Its expression is reduced in ACC. SF-1 overexpression in H295R cells significantly reduces NOV expression [108];
- -
- FATE1 encodes a protein enriched in mitochondria-associated membranes (MAM) which has an important role in modulating calcium transfer between the endoplasmic reticulum and mitochondria in ACC cells, being involved in the resistance to chemotherapeutic drugs [109,110]. In addition, FATE1 is a prognostic factor in ACC and a cancer-testis antigen against which an immune response is present in patients with ACC [111], making it a potential target for immunotherapy;
- -
- VAV2 encodes a guanine nucleotide exchange factor (GEF) which activates Rho family GTPases, important regulators of cell cytoskeleton, motility and invasion. VAV2 is a prognostic factor in ACC and its knockdown significantly inhibits H295R cell invasion in Matrigel activated by SF-1 overexpression [104].
8. SF-1 and the Tumor Microenvironment (TME) in ACC
9. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Berger, I.; Werdermann, M.; Bornstein, S.R.; Steenblock, C. The adrenal gland in stress—Adaptation on a cellular level. J. Steroid Biochem. Mol. Biol. 2019, 190, 198–206. [Google Scholar] [CrossRef]
- Zhang, Z.; Burch, P.E.; Cooney, A.J.; Lanz, R.B.; Pereira, F.A.; Wu, J.; Gibbs, R.A.; Weinstock, G.; Wheeler, D.A. Genomic analysis of the nuclear receptor family: New insights into structure, regulation, and evolution from the rat genome. Genome Res. 2004, 14, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Meinsohn, M.C.; Smith, O.E.; Bertolin, K.; Murphy, B.D. The orphan nuclear receptors Steroidogenic Factor-1 and Liver Receptor Homolog-1: Structure, regulation, and essential roles in mammalian reproduction. Physiol. Rev. 2019, 99, 1249–1279. [Google Scholar] [CrossRef] [PubMed]
- Lala, D.S.; Rice, D.A.; Parker, K.L. Steroidogenic factor I, a key regulator of steroidogenic enzyme expression, is the mouse homolog of fushi tarazu-factor I. Mol. Endocrinol. 1992, 6, 1249–1258. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, K.; Honda, S.; Inomata, Y.; Handa, H.; Omura, T. A common trans-acting factor, Ad4-binding protein, to the promoters of steroidogenic P-450s. J. Biol. Chem. 1992, 267, 17913–17919. [Google Scholar] [CrossRef] [PubMed]
- Parker, K.L.; Schimmer, B.P. Steroidogenic factor 1: A key determinant of endocrine development and function. Endocr. Rev. 1997, 18, 361–377. [Google Scholar] [CrossRef]
- Lalli, E.; Doghman, M.; Latre de Late, P.; El Wakil, A.; Mus-Veteau, I. Beyond steroidogenesis: Novel target genes for SF-1 discovered by genomics. Mol. Cell. Endocrinol. 2013, 371, 154–159. [Google Scholar] [CrossRef]
- Doghman, M.; Lalli, E. A matter of dosage: SF-1 in adrenocortical development and cancer. Ann. Endocrinol. 2009, 70, 148–152. [Google Scholar] [CrossRef] [PubMed]
- Achermann, J.C.; Ito, M.; Ito, M.; Hindmarsh, P.C.; Jameson, J.L. A mutation in the gene encoding Steroidogenic factor-1 causes XY sex reversal and adrenal failure in humans. Nat. Genet. 1998, 22, 125–126. [Google Scholar] [CrossRef]
- Lin, L.; Philibert, P.; Ferraz-de-Souza, B.; Kelberman, D.; Homfray, T.; Albanese, A.; Molini, V.; Sebire, N.J.; Einaudi, S.; Conway, G.S.; et al. Heterozygous missense mutations in steroidogenic factor 1 (SF1/Ad4BP, NR5A1) are associated with 46,XY disorders of sex development with normal adrenal function. J. Clin. Endocrinol. Metab. 2007, 92, 991–999. [Google Scholar] [CrossRef]
- Lin, L.; Achermann, J.C. Steroidogenic factor-1 (SF-1, Ad4BP, NR5A1) and disorders of testis development. Sex Dev. 2008, 2, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Bashamboo, A.; McElreavey, K. NR5A1/SF-1 and development and function of the ovary. Ann. Endocrinol. 2010, 71, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Bland, M.L.; Jamieson, C.A.; Akana, S.F.; Bornstein, S.R.; Eisenhofer, G.; Dallman, M.F.; Ingraham, H.A. Haploinsufficiency of Steroidogenic factor-1 in mice disrupts adrenal development leading to an impaired stress response. Proc. Natl. Acad. Sci. USA 2000, 97, 14488–14493. [Google Scholar] [CrossRef] [PubMed]
- Beuschlein, F.; Mutch, C.; Bavers, D.L.; Ulrich-Lai, Y.M.; Engeland, W.C.; Keegan, C.; Hammer, G.D. Steroidogenic factor-1 is essential for compensatory adrenal growth following unilateral adrenalectomy. Endocrinology 2002, 143, 3122–3135. [Google Scholar] [CrossRef]
- Doghman, M.; Karpova, T.; Rodrigues, G.A.; Arhatte, M.; De Moura, J.; Cavalli, L.R.; Virolle, V.; Barbry, P.; Zambetti, G.P.; Figueiredo, B.C.; et al. Increased steroidogenic factor-1 dosage triggers adrenocortical cell proliferation and cancer. Mol. Endocrinol. 2007, 21, 2968–2987. [Google Scholar] [CrossRef]
- Sbiera, S.; Schmull, S.; Assie, G.; Voelker, H.U.; Kraus, L.; Beyer, M.; Ragazzon, B.; Beuschlein, F.; Willenberg, H.S.; Hahner, S.; et al. High diagnostic and prognostic value of Steroidogenic factor-1 expression in adrenal tumors. J. Clin. Endocrinol. Metab. 2010, 95, E161–E171. [Google Scholar] [CrossRef]
- Taketo, M.; Parker, K.L.; Howard, T.A.; Tsukiyama, T.; Wong, M.; Niwa, O.; Morton, C.C.; Miron, P.M.; Seldin, M.F. Homologs of Drosophila Fushi-Tarazu factor 1 map to mouse chromosome 2 and human chromosome 9q33. Genomics 1995, 25, 565–567. [Google Scholar] [CrossRef]
- Evans, R.M. The steroid and thyroid hormone receptor superfamily. Science 1988, 240, 889–895. [Google Scholar] [CrossRef]
- Li, L.A.; Chiang, E.F.; Chen, J.C.; Hsu, N.C.; Chen, Y.J.; Chung, B.-C. Function of steroidogenic factor 1 domains in nuclear localization, transactivation, and interaction with transcription factor TFIIB and c-Jun. Mol. Endocrinol. 1999, 13, 1588–1598. [Google Scholar] [CrossRef]
- Pawlak, M.; Lefebvre, P.; Staels, B. General molecular biology and architecture of nuclear receptors. Curr. Top. Med. Chem. 2012, 12, 486–504. [Google Scholar] [CrossRef] [Green Version]
- Desclozeaux, M.; Krylova, I.N.; Horn, F.; Fletterick, R.J.; Ingraham, H.A. Phosphorylation and intramolecular stabilization of the ligand binding domain in the nuclear receptor Steroidogenic factor 1. Mol. Cell. Biol. 2002, 22, 7193–7203. [Google Scholar] [CrossRef]
- Krylova, I.N.; Sablin, E.P.; Moore, J.; Xu, R.X.; Waitt, G.M.; MacKay, J.A.; Juzumiene, D.; Bynum, J.M.; Madauss, K.; Montana, V.; et al. Structural analyses reveal phosphatidyl inositols as ligands for the NR5 orphan receptors SF-1 and LRH-1. Cell 2005, 120, 343–355. [Google Scholar] [CrossRef]
- Li, Y.; Choi, M.; Cavey, G.; Daugherty, J.; Suino, K.; Kovach, A.; Bingham, N.C.; Kliewer, S.A.; Xu, H.E. Crystallographic identification and functional characterization of phospholipids as ligands for the orphan nuclear receptor Steroidogenic factor-1. Mol. Cell 2005, 17, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, C.; Marimuthu, A.; Krupka, H.I.; Tabrizizad, M.; Shelloe, R.; Mehra, U.; Eng, K.; Nguyen, H.; Settachatgul, C.; et al. The crystal structures of human Steroidogenic factor-1 and Liver receptor homologue-1. Proc. Natl. Acad. Sci. USA 2005, 102, 7505–7510. [Google Scholar] [CrossRef] [PubMed]
- Hammer, G.D.; Krylova, I.; Zhang, Y.; Darimont, B.D.; Simpson, K.; Weigel, N.L.; Ingraham, H.A. Phosphorylation of the nuclear receptor SF-1 modulates cofactor recruitment: Integration of hormone signaling in reproduction and stress. Mol. Cell 1999, 3, 521–526. [Google Scholar] [CrossRef]
- Ikeda, Y.; Shen, W.H.; Ingraham, H.A.; Parker, K.L. Developmental expression of mouse Steroidogenic factor-1, an essential regulator of the steroid hydroxylases. Mol. Endocrinol. 1994, 8, 654–662. [Google Scholar] [CrossRef]
- Hatano, O.; Takakusu, A.; Nomura, M.; Morohashi, K. Identical origin of adrenal cortex and gonad revealed by expression profiles of Ad4BP/SF-1. Genes Cells 1996, 1, 663–671. [Google Scholar] [CrossRef]
- Val, P.; Martinez-Barbera, J.P.; Swain, A. Adrenal development is initiated by Cited2 and Wt1 through modulation of Sf-1 dosage. Development 2007, 134, 2349–2358. [Google Scholar] [CrossRef] [PubMed]
- Hanley, N.A.; Ball, S.G.; Clement-Jones, M.; Hagan, D.M.; Strachan, T.; Lindsay, S.; Robson, S.; Ostrer, H.; Parker, K.L.; Wilson, D.I. Expression of Steroidogenic factor 1 and Wilms’ tumour 1 during early human gonadal development and sex determination. Mech. Dev. 1999, 87, 175–180. [Google Scholar] [CrossRef]
- Abou Nader, N.; Boyer, A. Adrenal cortex development and maintenance: Knowledge acquired from mouse models. Endocrinology 2021, 162, bqab187. [Google Scholar] [CrossRef]
- Lalli, E. Adrenocortical development and cancer: Focus on SF-1. J. Mol. Endocrinol. 2010, 44, 301–307. [Google Scholar] [CrossRef]
- Ikeda, Y.; Lala, D.S.; Luo, X.; Kim, E.; Moisan, M.P.; Parker, K.L. Characterization of the mouse FTZ-F1 gene, which encodes a key regulator of steroid hydroxylase gene expression. Mol. Endocrinol. 1993, 7, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, K.; Iida, H.; Nomura, M.; Hatano, O.; Honda, S.; Tsukiyama, T.; Niwa, O.; Hara, T.; Takakusu, A.; Shibata, Y.; et al. Functional difference between Ad4BP and ELP, and their distributions in steroidogenic tissues. Mol. Endocrinol. 1994, 8, 643–653. [Google Scholar] [CrossRef]
- Falender, A.E.; Lanz, R.; Malenfant, D.; Belanger, L.; Richards, J. Differential expression of Steroidogenic factor-1 and FTF/LRH-1 in the rodent ovary. Endocrinology 2003, 144, 3598–3610. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Shin, E.; Won, M.; Kim, J.H.; Go, H.; Kim, H.L.; Ko, J.J.; Lee, K.; Bae, J. FOXL2 interacts with steroidogenic factor-1 (SF-1) and represses SF-1-induced CYP17 transcription in granulosa cells. Mol. Endocrinol. 2010, 24, 1024–1036. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Ikeda, Y.; Parker, K.L. A cell-specific nuclear receptor is essential for adrenal and gonadal development and sexual differentiation. Cell 1994, 77, 481–490. [Google Scholar] [CrossRef]
- Shinoda, K.; Lei, H.; Yoshii, H.; Nomura, M.; Nagano, M.; Shiba, H.; Sasaki, H.; Osawa, Y.; Ninomiya, Y.; Niwa, O.; et al. Developmental defects of the ventromedial hypothalamic nucleus and pituitary gonadotroph in the Ftz-F1 disrupted mice. Dev. Dyn. 1995, 204, 22–29. [Google Scholar] [CrossRef]
- Kurrasch, D.M.; Cheung, C.C.; Lee, F.Y.; Tran, P.V.; Hata, K.; Ingraham, H.A. The neonatal ventromedial hypothalamus transcriptome reveals novel markers with spatially distinct patterning. J. Neurosci. 2007, 27, 13624–13634. [Google Scholar] [CrossRef]
- Ingraham, H.A.; Lala, D.S.; Ikeda, Y.; Luo, X.; Shen, W.H.; Nachtigal, M.W.; Abbud, R.; Nilson, J.H.; Parker, K.L. The nuclear receptor Steroidogenic factor 1 acts at multiple levels of the reproductive axis. Genes Dev. 1994, 8, 2302–2312. [Google Scholar] [CrossRef]
- Barnhart, K.M.; Mellon, P.L. The orphan nuclear receptor, Steroidogenic factor-1, regulates the glycoprotein hormone alpha-subunit gene in pituitary gonadotropes. Mol. Endocrinol. 1994, 8, 878–885. [Google Scholar] [CrossRef] [Green Version]
- Ngan, E.S.; Cheng, P.K.; Leung, P.C.; Chow, B.K. Steroidogenic factor-1 interacts with a gonadotrope-specific element within the first exon of the human gonadotropin-releasing hormone receptor gene to mediate gonadotrope-specific expression. Endocrinology 1999, 140, 2452–2462. [Google Scholar] [CrossRef] [PubMed]
- Wehrenberg, U.; Prange-Kiel, J.; Rune, G.M. Steroidogenic factor-1 expression in marmoset and rat hippocampus: Co-localization with StAR and aromatase. J. Neurochem. 2001, 76, 1879–1886. [Google Scholar] [CrossRef]
- Morohashi, K.; Tsuboi-Asai, H.; Matsushita, S.; Suda, M.; Nakashima, M.; Sasano, H.; Hataba, Y.; Li, C.L.; Fukata, J.; Irie, J.; et al. Structural and functional abnormalities in the spleen of an mFtz-F1 gene-disrupted mouse. Blood 1999, 93, 1586–1594. [Google Scholar]
- Lai, P.Y.; Wang, C.Y.; Chen, W.Y.; Kao, Y.H.; Tsai, H.M.; Tachibana, T.; Chang, W.C.; Chung, B.C. Steroidogenic Factor 1 (NR5A1) resides in centrosomes and maintains genomic stability by controlling centrosome homeostasis. Cell Death Differ. 2011, 18, 1836–1844. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Kao, Y.H.; Lai, P.Y.; Chen, W.Y.; Chung, B.C. Steroidogenic factor 1 (NR5A1) maintains centrosome homeostasis in steroidogenic cells by restricting centrosomal DNA-dependent protein kinase activation. Mol. Cell. Biol. 2013, 33, 476–484. [Google Scholar] [CrossRef] [PubMed]
- Keegan, C.E.; Hammer, G.D. Recent insights into organogenesis of the adrenal cortex. Trends Endocrinol. Metab. 2002, 13, 200–208. [Google Scholar] [CrossRef]
- Cheng, K.; Seita, Y.; Moriwaki, T.; Noshiro, K.; Sakata, Y.; Hwang, Y.S.; Torigoe, T.; Saitou, M.; Tsuchiya, H.; Iwatani, C.; et al. The developmental origin and the specification of the adrenal cortex in humans and cynomolgus monkeys. Sci. Adv. 2022, 8, eabn8485. [Google Scholar] [CrossRef]
- Zubair, M.; Parker, K.L.; Morohashi, K. Developmental links between the fetal and adult zones of the adrenal cortex revealed by lineage tracing. Mol. Cell. Biol. 2008, 28, 7030–7040. [Google Scholar] [CrossRef]
- Freedman, B.D.; Kempna, P.B.; Carlone, D.L.; Shah, M.; Guagliardo, N.A.; Barrett, P.Q.; Gomez-Sanchez, C.E.; Majzoub, J.A.; Breault, D.T. Adrenocortical zonation results from lineage conversion of differentiated zona glomerulosa cells. Dev. Cell 2013, 26, 666–673. [Google Scholar] [CrossRef]
- Mesiano, S.; Jaffe, R.B. Developmental and functional biology of the primate fetal adrenal cortex. Endocr. Rev. 1997, 18, 378–403. [Google Scholar] [CrossRef]
- Sadovsky, Y.; Crawford, P.A.; Woodson, K.G.; Polish, J.A.; Clements, M.A.; Tourtellotte, L.M.; Simburger, K.; Milbrandt, J. Mice deficient in the orphan receptor Steroidogenic factor 1 lack adrenal glands and gonads but express P450 side-chain-cleavage enzyme in the placenta and have normal embryonic serum levels of corticosteroids. Proc. Natl. Acad. Sci. USA 1995, 92, 10939–10945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pignatti, E.; du Toit, T.; Flück, C.E. Development and function of the fetal adrenal. Rev. Endocr. Metab. Disord. 2023, 24, 5–21. [Google Scholar] [CrossRef]
- Sasaki, K.; Oguchi, A.; Cheng, K.; Murakawa, Y.; Okamoto, I.; Ohta, H.; Yabuta, Y.; Iwatani, C.; Tsuchiya, H.; Yamamoto, T.; et al. The embryonic ontogeny of the gonadal somatic cells in mice and monkeys. Cell Rep. 2021, 35, 109075. [Google Scholar] [CrossRef] [PubMed]
- Crawford, P.A.; Sadovsky, Y.; Milbrandt, J. Nuclear receptor Steroidogenic factor 1 directs embryonic stem cells toward the steroidogenic lineage. Mol. Cell. Biol. 1997, 17, 3997–4006. [Google Scholar] [CrossRef]
- Yazawa, T.; Imamichi, Y.; Miyamoto, K.; Khan, M.R.; Uwada, J.; Umezawa, A.; Taniguchi, T. Regulation of steroidogenesis, development, and cell differentiation by Steroidogenic Factor-1 and Liver Receptor Homolog-1. Zoolog. Sci. 2015, 32, 323–330. [Google Scholar] [CrossRef]
- Gondo, S.; Yanase, T.; Okabe, T.; Tanaka, T.; Morinaga, H.; Nomura, M.; Goto, K.; Nawata, H. SF-1/Ad4BP transforms primary long-term cultured bone marrow cells into ACTH-responsive steroidogenic cells. Genes Cells 2004, 9, 1239–1247. [Google Scholar] [CrossRef]
- Yazawa, T.; Mizutani, T.; Yamada, K.; Kawata, H.; Sekiguchi, T.; Yoshino, M.; Kajitani, T.; Shou, Z.; Umezawa, A.; Miyamoto, K. Differentiation of adult stem cells derived from bone marrow stroma into Leydig or adrenocortical cells. Endocrinology 2006, 147, 4104–4111. [Google Scholar] [CrossRef]
- Tanaka, T.; Gondo, S.; Okabe, T.; Ohe, K.; Shirohzu, H.; Morinaga, H.; Nomura, M.; Tani, K.; Takayanagi, R.; Nawata, H.; et al. Steroidogenic factor 1/adrenal 4 binding protein transforms human bone marrow mesenchymal cells into steroidogenic cells. J. Mol. Endocrinol. 2007, 39, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Gondo, S.; Okabe, T.; Tanaka, T.; Morinaga, H.; Nomura, M.; Takayanagi, R.; Nawata, H.; Yanase, T. Adipose tissue-derived and bone marrow-derived mesenchymal cells develop into different lineage of steroidogenic cells by forced expression of Steroidogenic factor 1. Endocrinology 2008, 149, 4717–4725. [Google Scholar] [CrossRef]
- Wei, X.; Peng, G.; Zheng, S.; Wu, X. Differentiation of umbilical cord mesenchymal stem cells into steroidogenic cells in comparison to bone marrow mesenchymal stem cells. Cell Prolif. 2012, 45, 101–110. [Google Scholar] [CrossRef]
- Ruiz-Babot, G.; Balyura, M.; Hadjidemetriou, I.; Ajodha, S.J.; Taylor, D.R.; Ghataore, L.; Taylor, N.F.; Schubert, U.; Ziegler, C.G.; Storr, H.L.; et al. Modeling congenital adrenal hyperplasia and testing interventions for adrenal insufficiency using donor-specific reprogrammed cells. Cell Rep. 2018, 22, 1236–1249. [Google Scholar] [CrossRef] [Green Version]
- Zubair, M.; Ishihara, S.; Oka, S.; Okumura, K.; Morohashi, K. Two-step regulation of Ad4BP/SF-1 gene transcription during fetal adrenal development: Initiation by a Hox-Pbx1-Prep1 complex and maintenance via autoregulation by Ad4BP/SF-1. Mol. Cell. Biol. 2006, 26, 4111–4121. [Google Scholar] [CrossRef]
- Zubair, M.; Oka, S.; Parker, K.L.; Morohashi, K. Transgenic expression of Ad4BP/SF-1 in fetal adrenal progenitor cells leads to ectopic adrenal formation. Mol. Endocrinol. 2009, 23, 1657–1667. [Google Scholar] [CrossRef] [PubMed]
- Ito, M.; Yu, R.; Jameson, J.L. DAX-1 inhibits SF-1-mediated transactivation via a carboxy-terminal domain that is deleted in adrenal hypoplasia congenita. Mol. Cell. Biol. 1997, 17, 1476–1483. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.B.; Lebedeva, L.A.; Suzawa, M.; Wadekar, S.A.; Desclozeaux, M.; Ingraham, H.A. The DEAD-box protein DP103 (Ddx20 or Gemin-3) represses orphan nuclear receptor activity via SUMO modification. Mol. Cell. Biol. 2005, 25, 1879–1890. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.Y.; Faivre, E.J.; Suzawa, M.; Lontok, E.; Ebert, D.; Cai, F.; Belsham, D.D.; Ingraham, H.A. Eliminating SF-1 (NR5A1) sumoylation in vivo results in ectopic hedgehog signaling and disruption of endocrine development. Dev. Cell 2011, 21, 315–327. [Google Scholar] [CrossRef]
- Xing, Y.; Morohashi, K.I.; Ingraham, H.A.; Hammer, G.D. Timing of adrenal regression controlled by synergistic interaction between Sf1 SUMOylation and Dax1. Development 2017, 144, 3798–3807. [Google Scholar] [CrossRef]
- Yu, R.N.; Ito, M.; Saunders, T.L.; Camper, S.A.; Jameson, J.L. Role of Ahch in gonadal development and gametogenesis. Nat. Genet. 1998, 20, 353–357. [Google Scholar] [CrossRef]
- Fatchiyah; Zubair, M.; Shima, Y.; Oka, S.; Ishihara, S.; Fukui-Katoh, Y.; Morohashi, K. Differential gene dosage effects of Ad4BP/SF-1 on target tissue development. Biochem. Biophys. Res. Commun. 2006, 341, 1036–1045. [Google Scholar] [CrossRef]
- Katoh-Fukui, Y.; Owaki, A.; Toyama, Y.; Kusaka, M.; Shinohara, Y.; Maekawa, M.; Toshimori, K.; Morohashi, K. Mouse Polycomb M33 is required for splenic vascular and adrenal gland formation through regulating Ad4BP/SF1 expression. Blood 2005, 106, 1612–1620. [Google Scholar] [CrossRef] [PubMed]
- Guasti, L.; Candy Sze, W.C.; McKay, T.; Grose, R.; King, P.J. FGF signalling through Fgfr2 isoform IIIb regulates adrenal cortex development. Mol. Cell. Endocrinol. 2013, 371, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, Y.; Tanaka, S.S.; Yamaguchi, Y.L.; Kobayashi, H.; Kuroki, S.; Tachibana, M.; Shinomura, M.; Kanai, Y.; Morohashi, K.; Kawakami, K.; et al. Homeoproteins Six1 and Six4 regulate male sex determination and mouse gonadal development. Dev. Cell 2013, 26, 416–430. [Google Scholar] [CrossRef]
- Pitetti, J.L.; Calvel, P.; Romero, Y.; Conne, B.; Truong, V.; Papaioannou, M.D.; Schaad, O.; Docquier, M.; Herrera, P.L.; Wilhelm, D.; et al. Insulin and IGF1 receptors are essential for XX and XY gonadal differentiation and adrenal development in mice. PLoS Genet. 2013, 9, e1003160. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, C.A.; Selleri, L.; Cleary, M.L. Pbx1 is essential for adrenal development and urogenital differentiation. Genesis 2003, 37, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Morohashi, K.; Zubair, M. The fetal and adult adrenal cortex. Mol. Cell. Endocrinol. 2011, 336, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y.; Zubair, M.; Ishihara, S.; Shinohara, Y.; Oka, S.; Kimura, S.; Okamoto, S.; Minokoshi, Y.; Suita, S.; Morohashi, K. Ventromedial hypothalamic nucleus-specific enhancer of Ad4BP/SF-1 gene. Mol. Endocrinol. 2005, 19, 2812–2823. [Google Scholar] [CrossRef] [PubMed]
- Shima, Y.; Zubair, M.; Komatsu, T.; Oka, S.; Yokoyama, C.; Tachibana, T.; Hjalt, T.A.; Drouin, J.; Morohashi, K. Pituitary homeobox 2 regulates adrenal4 binding protein/steroidogenic factor-1 gene transcription in the pituitary gonadotrope through interaction with the intronic enhancer. Mol. Endocrinol. 2008, 22, 1633–1646. [Google Scholar] [CrossRef]
- Ikeda, Y.; Swain, A.; Weber, T.J.; Hentges, K.E.; Zanaria, E.; Lalli, E.; Tamai, K.T.; Sassone-Corsi, P.; Lovell-Badge, R.; Camerino, G.; et al. Steroidogenic factor 1 and Dax-1 colocalize in multiple cell lineages: Potential links in endocrine development. Mol. Endocrinol. 1996, 10, 1261–1272. [Google Scholar] [CrossRef]
- Lalli, E. Role of orphan nuclear receptor DAX-1/NR0B1 in development, physiology and disease. Adv. Biol. 2014, 2014, 582749. [Google Scholar] [CrossRef]
- Zazopoulos, E.; Lalli, E.; Stocco, D.M.; Sassone-Corsi, P. DNA binding and transcriptional repression by DAX-1 blocks steroidogenesis. Nature 1997, 390, 311–315. [Google Scholar] [CrossRef]
- Wang, Q.; Lan, Y.; Cho, E.S.; Maltby, K.M.; Jiang, R. Odd-skipped related 1 (Odd1) is an essential regulator of heart and urogenital development. Dev. Biol. 2005, 288, 582–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishinakamura, R.; Matsumoto, Y.; Nakao, K.; Nakamura, K.; Sato, A.; Copeland, N.G.; Gilbert, D.J.; Jenkins, N.A.; Scully, S.; Lacey, D.L.; et al. Murine homolog of SALL1 is essential for ureteric bud invasion in kidney development. Development 2001, 128, 3105–3115. [Google Scholar] [CrossRef]
- Doghman, M.; El Wakil, A.; Cardinaud, B.; Thomas, E.; Wang, J.; Zhao, W.; Peralta-Del Valle, M.H.; Figueiredo, B.C.; Zambetti, G.P.; Lalli, E. Regulation of insulin-like growth factor-mammalian target of rapamycin signaling by microRNA in childhood adrenocortical tumors. Cancer Res. 2010, 70, 4666–4675. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.Y.; Lee, W.C.; Hsu, N.C.; Huang, F.; Chung, B.C. SUMO modification of repression domains modulates function of nuclear receptor 5A1 (Steroidogenic factor-1). J. Biol. Chem. 2004, 279, 38730–38735. [Google Scholar] [CrossRef]
- Chen, W.Y.; Juan, L.J.; Chung, B.C. SF-1 (nuclear receptor 5A1) activity is activated by cyclic AMP via p300-mediated recruitment to active foci, acetylation, and increased DNA binding. Mol. Cell. Biol. 2005, 25, 10442–10453. [Google Scholar] [CrossRef] [PubMed]
- Jacob, A.L.; Lund, J.; Martinez, P.; Hedin, L. Acetylation of steroidogenic factor 1 protein regulates its transcriptional activity and recruits the coactivator GCN5. J. Biol. Chem. 2001, 276, 37659–37664. [Google Scholar] [CrossRef]
- Fan, W.; Yanase, T.; Wu, Y.; Kawate, H.; Saitoh, M.; Oba, K.; Nomura, M.; Okabe, T.; Goto, K.; Yanagisawa, J.; et al. Protein kinase A potentiates adrenal 4 binding protein/steroidogenic factor 1 transactivation by reintegrating the subcellular dynamic interactions of the nuclear receptor with its cofactors, general control nonderepressed-5/transformation/transcription domain-associated protein, and suppressor, dosage-sensitive sex reversal-1: A laser confocal imaging study in living KGN cells. Mol. Endocrinol. 2004, 18, 127–141. [Google Scholar] [CrossRef]
- Winnay, J.N.; Hammer, G.D. Adrenocorticotropic hormone-mediated signaling cascades coordinate a cyclic pattern of steroidogenic factor 1-dependent transcriptional activation. Mol. Endocrinol. 2006, 20, 147–166. [Google Scholar] [CrossRef]
- Urs, A.N.; Dammer, E.; Sewer, M.B. Sphingosine regulates the transcription of CYP17 by binding to Steroidogenic factor-1. Endocrinology 2006, 147, 5249–5258. [Google Scholar] [CrossRef]
- Li, D.; Urs, A.N.; Allegood, J.; Leon, A.; Merrill, A.H., Jr.; Sewer, M.B. Cyclic AMP-stimulated interaction between steroidogenic factor 1 and diacylglycerol kinase theta facilitates induction of CYP17. Mol. Cell. Biol. 2007, 27, 6669–6685. [Google Scholar] [CrossRef]
- Chen, W.Y.; Weng, J.H.; Huang, C.C.; Chung, B.C. Histone deacetylase inhibitors reduce steroidogenesis through SCF-mediated ubiquitination and degradation of steroidogenic factor 1 (NR5A1). Mol. Cell. Biol. 2007, 27, 7284–7290. [Google Scholar] [CrossRef] [Green Version]
- Fowkes, R.C.; Desclozeaux, M.; Patel, M.V.; Aylwin, S.J.; King, P.; Ingraham, H.A.; Burrin, J.M. Steroidogenic factor-1 and the gonadotrope-specific element enhance basal and pituitary adenylate cyclase-activating polypeptide-stimulated transcription of the human glycoprotein hormone alpha-subunit gene in gonadotropes. Mol. Endocrinol. 2003, 17, 2177–2188. [Google Scholar] [CrossRef]
- Lewis, A.E.; Rusten, M.; Hoivik, E.A.; Vikse, E.L.; Hansson, M.L.; Wallberg, A.E.; Bakke, M. Phosphorylation of Steroidogenic factor 1 is mediated by cyclin-dependent kinase 7. Mol. Endocrinol. 2008, 22, 91–104. [Google Scholar] [CrossRef]
- Campbell, L.A.; Faivre, E.J.; Show, M.D.; Ingraham, J.G.; Flinders, J.; Gross, J.D.; Ingraham, H.A. Decreased recognition of SUMO-sensitive target genes following modification of SF-1 (NR5A1). Mol. Cell. Biol. 2008, 28, 7476–7486. [Google Scholar] [CrossRef]
- Yang, W.H.; Heaton, J.H.; Brevig, H.; Mukherjee, S.; Iñiguez-Lluhí, J.A.; Hammer, G.D. SUMOylation inhibits SF-1 activity by reducing CDK7-mediated serine 203 phosphorylation. Mol. Cell. Biol. 2009, 29, 613–625. [Google Scholar] [CrossRef]
- Suntharalingham, J.P.; Buonocore, F.; Duncan, A.J.; Achermann, J.C. DAX-1 (NR0B1) and steroidogenic factor-1 (SF-1, NR5A1) in human disease. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 607–619. [Google Scholar] [CrossRef]
- Biason-Lauber, A.; Schoenle, E.J. Apparently normal ovarian differentiation in a prepubertal girl with transcriptionally inactive steroidogenic factor 1 (NR5A1/SF-1) and adrenocortical insufficiency. Am. J. Hum. Genet. 2000, 67, 1563–1568. [Google Scholar] [CrossRef]
- Custódio, G.; Parise, G.A.; Kiesel Filho, N.; Komechen, H.; Sabbaga, C.C.; Rosati, R.; Grisa, L.; Parise, I.Z.; Pianovski, M.A.; Fiori, C.M.; et al. Impact of neonatal screening and surveillance for the TP53 R337H mutation on early detection of childhood adrenocortical tumors. J. Clin. Oncol. 2013, 31, 2619–2626. [Google Scholar] [CrossRef]
- Figueiredo, B.C.; Stratakis, C.A.; Sandrini, R.; DeLacerda, L.; Pianovski, M.A.; Giatzakis, C.; Young, H.M.; Haddad, B.R. Comparative genomic hybridization analysis of adrenocortical tumors of childhood. J. Clin. Endocrinol. Metab. 1999, 84, 1116–1121. [Google Scholar] [CrossRef]
- Figueiredo, B.C.; Cavalli, L.R.; Pianovski, M.A.; Lalli, E.; Sandrini, R.; Ribeiro, R.C.; Zambetti, G.; DeLacerda, L.; Rodrigues, G.A.; Haddad, B.R. Amplification of the Steroidogenic factor 1 gene in childhood adrenocortical tumors. J. Clin. Endocrinol. Metab. 2005, 90, 615–619. [Google Scholar] [CrossRef]
- Pianovski, M.A.; Cavalli, L.R.; Figueiredo, B.C.; Santos, S.C.; Doghman, M.; Ribeiro, R.C.; Oliveira, A.G.; Michalkiewicz, E.; Rodrigues, G.A.; Zambetti, G.; et al. SF-1 overexpression in childhood adrenocortical tumours. Eur. J. Cancer 2006, 42, 1040–1043. [Google Scholar] [CrossRef]
- Almeida, M.Q.; Soares, I.C.; Ribeiro, T.C.; Fragoso, M.C.; Marins, L.V.; Wakamatsu, A.; Ressio, R.A.; Nishi, M.Y.; Jorge, A.A.; Lerario, A.M.; et al. Steroidogenic factor 1 overexpression and gene amplification are more frequent in adrenocortical tumors from children than from adults. J. Clin. Endocrinol. Metab. 2010, 95, 1458–1462. [Google Scholar] [CrossRef]
- Latre de Late, P.; El Wakil, A.; Jarjat, M.; de Krijger, R.R.; Heckert, L.L.; Naquet, P.; Lalli, E. Vanin-1 inactivation antagonizes the development of adrenocortical neoplasia in Sf-1 transgenic mice. Endocrinology 2014, 155, 2349–2354. [Google Scholar] [CrossRef]
- Ruggiero, C.; Doghman-Bouguerra, M.; Sbiera, S.; Sbiera, I.; Parsons, M.; Ragazzon, B.; Morin, A.; Robidel, E.; Favier, J.; Bertherat, J.; et al. Dosage-dependent regulation of VAV2 expression by Steroidogenic factor-1 drives adrenocortical carcinoma cell invasion. Sci. Signal. 2017, 10, eaal2464. [Google Scholar] [CrossRef]
- Doghman, M.; Cazareth, J.; Douguet, D.; Madoux, F.; Hodder, P.; Lalli, E. Inhibition of adrenocortical carcinoma cell proliferation by steroidogenic factor-1 inverse agonists. J. Clin. Endocrinol. Metab. 2009, 94, 178–183. [Google Scholar] [CrossRef]
- West, A.N.; Neale, G.A.; Pounds, S.; Figueiredo, B.C.; Rodriguez Galindo, C.; Pianovski, M.A.; Oliveira Filho, A.G.; Malkin, D.; Lalli, E.; Ribeiro, R.; et al. Gene expression profiling of childhood adrenocortical tumors. Cancer Res. 2007, 67, 600–608. [Google Scholar] [CrossRef]
- Doghman, M.; Figueiredo, B.C.; Volante, M.; Papotti, M.; Lalli, E. Integrative analysis of SF-1 transcription factor dosage impact on genome-wide binding and gene expression regulation. Nucleic Acids Res. 2013, 41, 8896–8907. [Google Scholar] [CrossRef]
- Doghman, M.; Arhatte, M.; Thibout, H.; Rodrigues, G.; De Moura, J.; Grosso, S.; West, A.N.; Laurent, M.; Mas, J.C.; Bongain, A.; et al. Nephroblastoma overexpressed/cysteine-rich protein 61/connective tissue growth factor/nephroblastoma overexpressed gene-3 (NOV/CCN3), a selective adrenocortical cell proapoptotic factor, is down-regulated in childhood adrenocortical tumors. J. Clin. Endocrinol. Metab. 2007, 92, 3253–3260. [Google Scholar] [CrossRef]
- Doghman-Bouguerra, M.; Granatiero, V.; Sbiera, S.; Sbiera, I.; Lacas-Gervais, S.; Brau, F.; Fassnacht, M.; Rizzuto, R.; Lalli, E. FATE1 antagonizes calcium- and drug-induced apoptosis by uncoupling ER and mitochondria. EMBO Rep. 2016, 7, 1264–1280. [Google Scholar] [CrossRef]
- Doghman-Bouguerra, M.; Lalli, E. ER-mitochondria interactions: Both strength and weakness within cancer cells. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 650–662. [Google Scholar] [CrossRef]
- Doghman-Bouguerra, M.; Finetti, P.; Durand, N.; Parise, I.Z.S.; Sbiera, S.; Cantini, G.; Canu, L.; Hescot, S.; Figueiredo, M.M.O.; Komechen, H.; et al. Cancer-testis antigen FATE1 expression in adrenocortical tumors is associated with a pervasive autoimmune response and is a marker of malignancy in adult, but not children, ACC. Cancers 2020, 12, 689. [Google Scholar] [CrossRef] [Green Version]
- Ruggiero, C.; Doghman, M.; Lalli, E. How genomic studies have improved our understanding of the mechanisms of transcriptional regulation by NR5A nuclear receptors. Mol. Cell. Endocrinol. 2015, 408, 138–144. [Google Scholar] [CrossRef]
- Battista, M.C.; Otis, M.; Côté, M.; Laforest, A.; Peter, M.; Lalli, E.; Gallo-Payet, N. Extracellular matrix and hormones modulate DAX-1 localization in the human fetal adrenal gland. J. Clin. Endocrinol. Metab. 2005, 90, 5426–5431. [Google Scholar] [CrossRef]
- Sakata, Y.; Cheng, K.; Mayama, M.; Seita, Y.; Detlefsen, A.J.; Mesaros, C.A.; Penning, T.M.; Shishikura, K.; Yang, W.; Auchus, R.J.; et al. Reconstitution of human adrenocortical specification and steroidogenesis using induced pluripotent stem cells. Dev. Cell 2022, 57, 2566–2583.e8. [Google Scholar] [CrossRef]
- Wilding, J.L.; Bodmer, W.F. Cancer cell lines for drug discovery and development. Cancer Res. 2014, 74, 2377–2384. [Google Scholar] [CrossRef]
- Fletcher, M.N.; Castro, M.A.; Wang, X.; de Santiago, I.; O’Reilly, M.; Chin, S.F.; Rueda, O.M.; Caldas, C.; Ponder, B.A.; Markowetz, F.; et al. Master regulators of FGFR2 signalling and breast cancer risk. Nat. Commun. 2013, 4, 2464. [Google Scholar] [CrossRef]
- Castro, M.A.; de Santiago, I.; Campbell, T.M.; Vaughn, C.; Hickey, T.E.; Ross, E.; Tilley, W.D.; Markowetz, F.; Ponder, B.A.; Meyer, K.B. Regulators of genetic risk of breast cancer identified by integrative network analysis. Nat. Genet. 2016, 48, 12–21. [Google Scholar] [CrossRef]
- Zheng, S.; Cherniack, A.D.; Dewal, N.; Moffitt, R.A.; Danilova, L.; Murray, B.A.; Lerario, A.M.; Else, T.; Knijnenburg, T.A.; Ciriello, G.; et al. Comprehensive pan-genomic characterization of adrenocortical carcinoma. Cancer Cell 2016, 29, 723–736. [Google Scholar] [CrossRef]
- Muzzi, J.C.D.; Magno, J.M.; Souza, J.S.; Alvarenga, L.M.; de Moura, J.F.; Figueiredo, B.C.; Castro, M.A.A. Comprehensive characterization of the regulatory landscape of adrenocortical carcinoma: Novel transcription factors and targets associated with prognosis. Cancers 2022, 14, 5279. [Google Scholar] [CrossRef]
- Assié, G.; Letouzé, E.; Fassnacht, M.; Jouinot, A.; Luscap, W.; Barreau, O.; Omeiri, H.; Rodriguez, S.; Perlemoine, K.; René-Corail, F.; et al. Integrated genomic characterization of adrenocortical carcinoma. Nat. Genet. 2014, 46, 607–612. [Google Scholar] [CrossRef]
- Landwehr, L.S.; Altieri, B.; Schreiner, J.; Sbiera, I.; Weigand, I.; Kroiss, M.; Fassnacht, M.; Sbiera, S. Interplay between glucocorticoids and tumor-infiltrating lymphocytes on the prognosis of adrenocortical carcinoma. J. Immunother. Cancer 2020, 8, e000469. [Google Scholar] [CrossRef]
- Muzzi, J.C.D.; Magno, J.M.; Cardoso, M.A.; de Moura, J.; Castro, M.A.A.; Figueiredo, B.C. Adrenocortical carcinoma steroid profiles: In silico pan-cancer analysis of TCGA data uncovers immunotherapy targets for potential improved outcomes. Front. Endocrinol. 2021, 12, 672319. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Model | Adrenal Phenotype | Observed in the Model | Reference |
|---|---|---|---|
| Cited2−/− | Absence of adrenal glands | Lack of Sf-1 expression | [28] |
| Cited2+/−; Wt1+/− | Marked decrease in adrenal size at E11.5 | Decrease in Sf-1 expression due to activation of Nr5a1 promoter by Cited2 et Wt1 | [28] |
| Six1−/−; Six4−/− | Smaller adrenal glands than wild type counterparts | Reduced Nr5a1 expression due to the ability of Six1 and Six4 to bind to the Nr5a1 promoter | [72] |
| M33−/− | Underdeveloped adrenal glands | Decreased Nr5a1 expression through indirect binding to the Nr5a1 locus | [70] |
| Insr−/−; Igf1r−/− | Adrenal agenesis for the majority of mutant animals; drastic reduction in adrenal size for a small number of mutants | Altered expression of the core AGP program including a reduction in Nr5a1 transcript levels | [73] |
| Pbx1−/− | Absence of adrenal glands | Decrease in SF-1 expression in Pbx1-/- embryos correlated with the absence of the adrenocortical cell population | [74] |
| Fgfr2b−/− | Adrenal hypoplasia | Lack of FGFR2b splice variant which causes reduced Sf-1 mRNA levels and reduction in inner cortical cell proliferation | [71] |
| Sf12KR/2KR | Smaller adrenal glands and delayed postnatal regression of adrenal X-zone | SUMOylation-deficiency leading to persistent FadE activity in X-zone | [66,67] |
| Dax1−/y | Delayed postnatal regression of adrenal X-zone | Dax1-deficiency leading to persistent FadE activity in X-zone due to its repressor role towards Sf-1 | [67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Relav, L.; Doghman-Bouguerra, M.; Ruggiero, C.; Muzzi, J.C.D.; Figueiredo, B.C.; Lalli, E. Steroidogenic Factor 1, a Goldilocks Transcription Factor from Adrenocortical Organogenesis to Malignancy. Int. J. Mol. Sci. 2023, 24, 3585. https://doi.org/10.3390/ijms24043585
Relav L, Doghman-Bouguerra M, Ruggiero C, Muzzi JCD, Figueiredo BC, Lalli E. Steroidogenic Factor 1, a Goldilocks Transcription Factor from Adrenocortical Organogenesis to Malignancy. International Journal of Molecular Sciences. 2023; 24(4):3585. https://doi.org/10.3390/ijms24043585
Chicago/Turabian StyleRelav, Lauriane, Mabrouka Doghman-Bouguerra, Carmen Ruggiero, João C. D. Muzzi, Bonald C. Figueiredo, and Enzo Lalli. 2023. "Steroidogenic Factor 1, a Goldilocks Transcription Factor from Adrenocortical Organogenesis to Malignancy" International Journal of Molecular Sciences 24, no. 4: 3585. https://doi.org/10.3390/ijms24043585