
Hyperoxidized Species of Heme Have a Potent Capacity to Induce Autoreactivity of Human IgG Antibodies

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
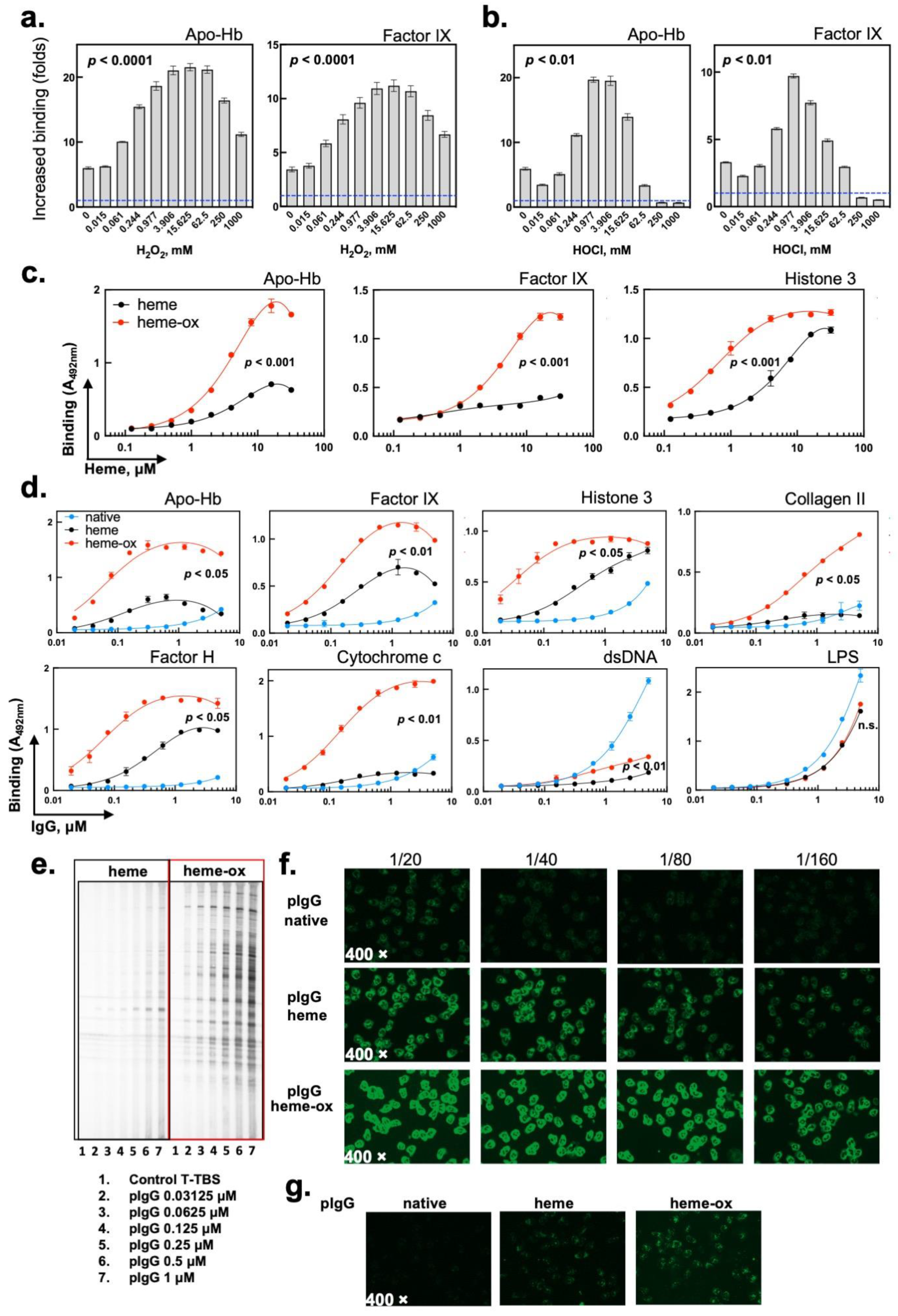
2.1. The Interaction of Heme (Fe3+) with Prooxidative Compounds Increases Its Capacity to Induce the Autoreactivity of Human IgG
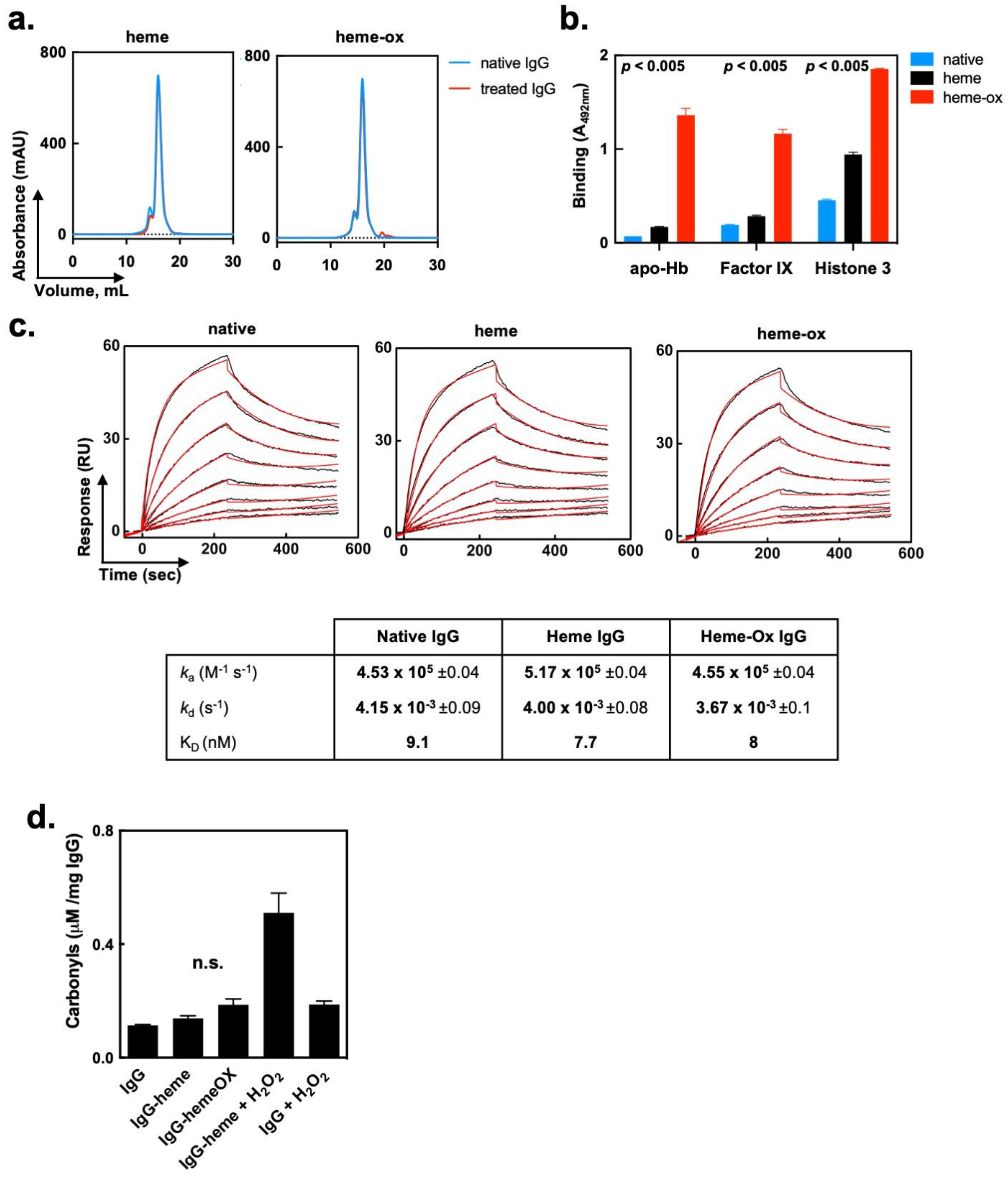
2.2. Heme (Fe3+) and Heme-Ox Do Not Affect the Overall Molecular Integrity of IgG
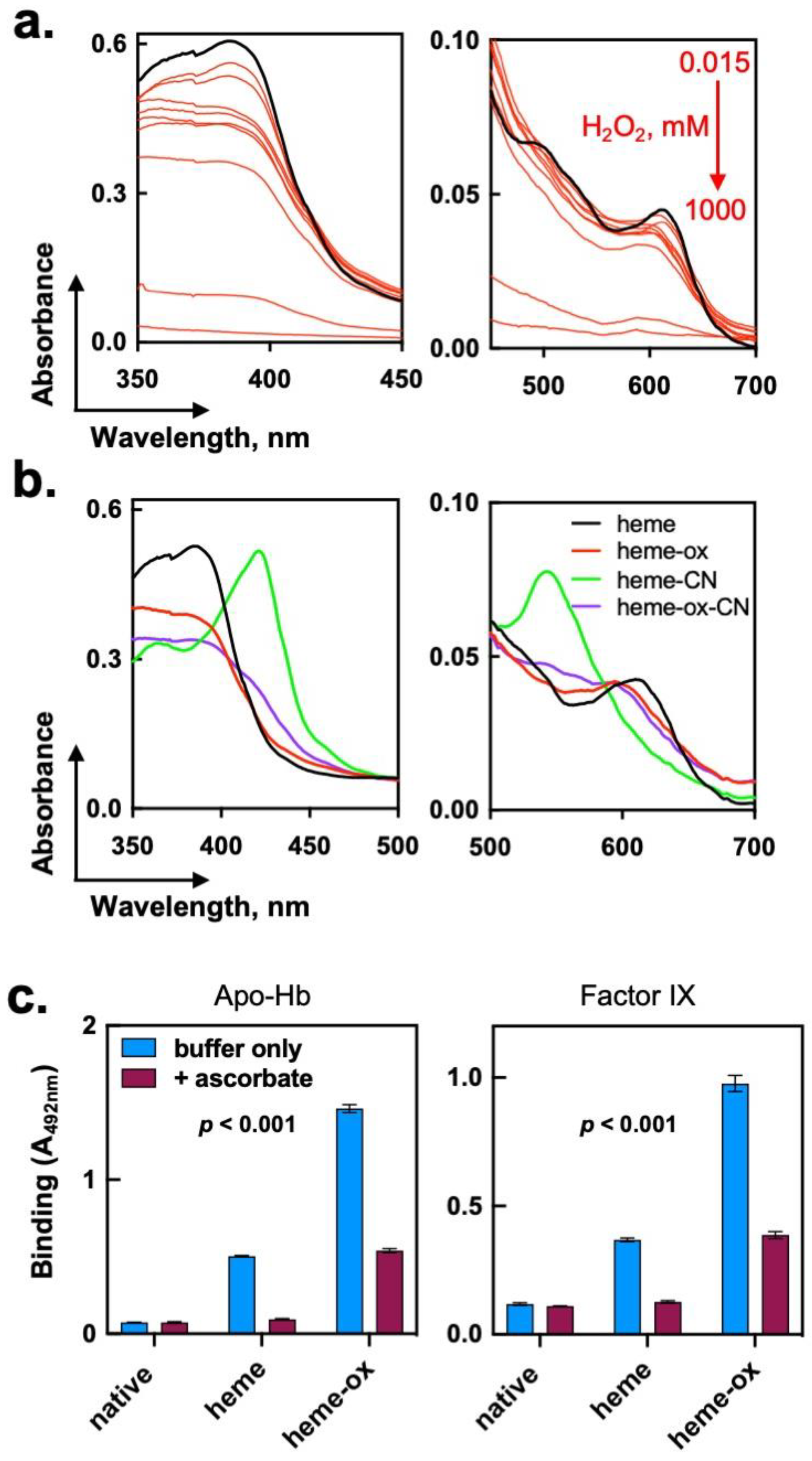
2.3. Consequences of the Exposure of Heme (Fe3+) to Hydrogen Peroxide
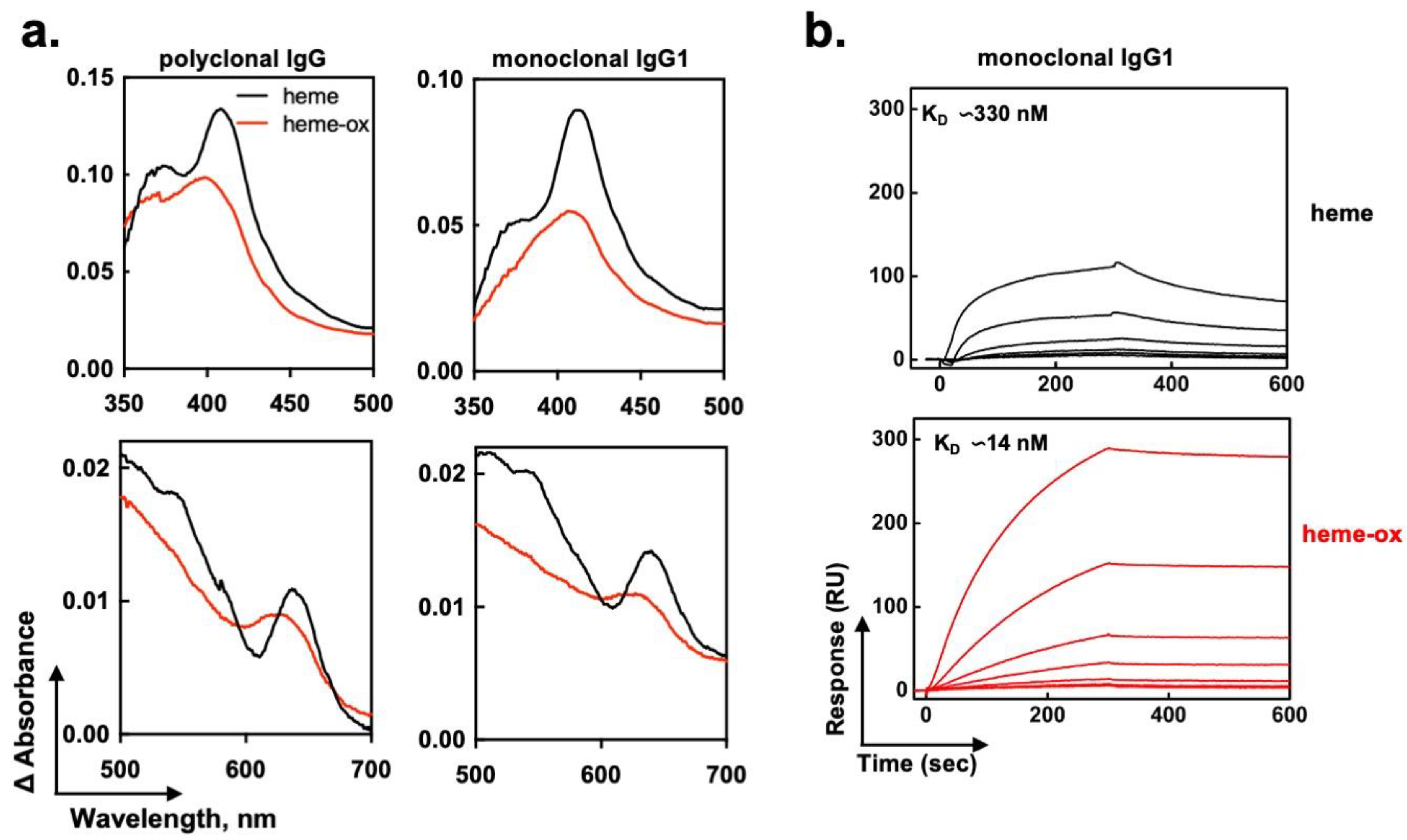
2.4. Heme-Ox Interacts with IgG in a Different Manner as Compared to Heme (Fe3+)
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. ELISA
4.3. Immunoblot
4.4. Indirect Immunofluorescence ANCA Assay
- (1)
- With regard to the preparation of samples, polyclonal IgG (PBS dialyzed) was first diluted to 10 mg/mL (67 μM) in PBS, and treated with 40 μM of final concentration of heme (Fe3+), heme-ox, or excipient only. After treatment, the preparations were stored at 4 °C until use.
- (2)
- ANCA assay was performed by indirect immunofluorescence by using biochip of human neutrophils fixed in ethanol and formol (Euroimmun, Lübeck, Germany) according to the manufacturer’s recommendations with samples diluted from 1/20 to 1/1280 in sample buffer. Biochip reading was performed with a fluorescent microscope (Axioscope A1, Zeiss, Oberkochen, Germany) and the positivity was assessed by two independent readers in a blind manner. Identification of the antigenic target was performed by ELISA (Euroimmun) by using ANCA profile assay (proteinase 3, myeloperoxydase, elastase, cathepsin G, bacterial permeability increasing protein, lactoferrin separately) according to the manufacturer‘s procedure. Results were expressed as the ratio between the optical density (OD) of the well of each antigen to the OD of the reference well, and positivity was defined by a ratio superior to 1.
4.5. Size Exclusion Chromatography
4.6. Determination of Oxidative Modifications in IgG (Carbonyls Assay)
4.7. UV-Vis Absorbance Spectroscopy
4.8. Real-Time Interaction Analyses
4.8.1. Measurement of Binding Affinity of Fc-γ Fragments to Human FcRn
4.8.2. Measurement of Binding Affinity of Heme to Human IgG
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munro, A.W.; Girvan, H.M.; McLean, K.J.; Cheesman, M.R.; Leys, D. Heme and Hemoproteins. In Tetrapyrroles. Molecular Biology Intelligence Unit; Springer: New York, NY, USA, 2009; pp. 160–183. [Google Scholar]
- Tang, X.D.; Xu, R.; Reynolds, M.F.; Garcia, M.L.; Heinemann, S.H.; Hoshi, T. Haem can bind to and inhibit mammalian calcium-dependent Slo1 BK channels. Nature 2003, 425, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, T.; Imhof, D. Regulatory Fe(II/III) heme: The reconstruction of a molecule’s biography. ChemBioChem 2014, 15, 2024–2035. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Lengalova, A.; Martinek, V.; Martinkova, M. Heme: Emergent roles of heme in signal transduction, functional regulation and as catalytic centres. Chem. Soc. Rev. 2019, 48, 5624–5657. [Google Scholar] [CrossRef] [PubMed]
- Muller-Eberhard, U.; Javid, J.; Liem, H.H.; Hanstein, A.; Hanna, M. Plasma concentrations of hemopexin, haptoglobin and heme in patients with various hemolytic diseases. Blood 1968, 32, 811–815. [Google Scholar] [CrossRef]
- Vallelian, F.; Buehler, P.W.; Schaer, D.J. Hemolysis, free hemoglobin toxicity and scavenger protein therapeutics. Blood 2022, 140, 1837–1844. [Google Scholar] [CrossRef]
- Schaer, D.J.; Buehler, P.W.; Alayash, A.I.; Belcher, J.D.; Vercellotti, G.M. Hemolysis and free hemoglobin revisited: Exploring hemoglobin and hemin scavengers as a novel class of therapeutic proteins. Blood 2013, 121, 1276–1284. [Google Scholar] [CrossRef]
- Wagener, F.A.; Eggert, A.; Boerman, O.C.; Oyen, W.J.; Verhofstad, A.; Abraham, N.G.; Adema, G.; van Kooyk, Y.; de Witte, T.; Figdor, C.G. Heme is a potent inducer of inflammation in mice and is counteracted by heme oxygenase. Blood 2001, 98, 1802–1811. [Google Scholar] [CrossRef]
- Graca-Souza, A.V.; Arruda, M.A.; de Freitas, M.S.; Barja-Fidalgo, C.; Oliveira, P.L. Neutrophil activation by heme: Implications for inflammatory processes. Blood 2002, 99, 4160–4165. [Google Scholar] [CrossRef]
- Kumar, S.; Bandyopadhyay, U. Free heme toxicity and its detoxification systems in human. Toxicol. Lett. 2005, 157, 175–188. [Google Scholar] [CrossRef]
- Figueiredo, R.T.; Fernandez, P.L.; Mourao-Sa, D.S.; Porto, B.N.; Dutra, F.F.; Alves, L.S.; Oliveira, M.F.; Oliveira, P.L.; Graca-Souza, A.V.; Bozza, M.T. Characterization of heme as activator of Toll-like receptor 4. J. Biol. Chem. 2007, 282, 20221–20229. [Google Scholar] [CrossRef] [Green Version]
- Dutra, F.F.; Bozza, M.T. Heme on innate immunity and inflammation. Front. Pharmacol. 2014, 5, 115. [Google Scholar] [CrossRef]
- Chiabrando, D.; Vinchi, F.; Fiorito, V.; Mercurio, S.; Tolosano, E. Heme in pathophysiology: A matter of scavenging, metabolism and trafficking across cell membranes. Front. Pharmacol. 2014, 5, 61. [Google Scholar] [CrossRef]
- Dutra, F.F.; Alves, L.S.; Rodrigues, D.; Fernandez, P.L.; de Oliveira, R.B.; Golenbock, D.T.; Zamboni, D.S.; Bozza, M.T. Hemolysis-induced lethality involves inflammasome activation by heme. Proc. Natl. Acad. Sci. USA 2014, 111, E4110–E4118. [Google Scholar] [CrossRef]
- Soares, M.P.; Bozza, M.T. Red alert: Labile heme is an alarmin. Curr. Opin. Immunol. 2016, 38, 94–100. [Google Scholar] [CrossRef]
- Larsen, R.; Gozzelino, R.; Jeney, V.; Tokaji, L.; Bozza, F.A.; Japiassu, A.M.; Bonaparte, D.; Cavalcante, M.M.; Chora, A.; Ferreira, A.; et al. A central role for free heme in the pathogenesis of severe sepsis. Sci. Transl. Med. 2010, 2, 51ra71. [Google Scholar] [CrossRef]
- Larsen, R.; Gouveia, Z.; Soares, M.P.; Gozzelino, R. Heme cytotoxicity and the pathogenesis of immune-mediated inflammatory diseases. Front. Pharmacol. 2012, 3, 77. [Google Scholar] [CrossRef]
- Chen, G.; Zhang, D.; Fuchs, T.A.; Manwani, D.; Wagner, D.D.; Frenette, P.S. Heme-induced neutrophil extracellular traps contribute to the pathogenesis of sickle cell disease. Blood 2014, 123, 3818–3827. [Google Scholar] [CrossRef]
- Belcher, J.D.; Chen, C.; Nguyen, J.; Milbauer, L.; Abdulla, F.; Alayash, A.I.; Smith, A.; Nath, K.A.; Hebbel, R.P.; Vercellotti, G.M. Heme triggers TLR4 signaling leading to endothelial cell activation and vaso-occlusion in murine sickle cell disease. Blood 2014, 123, 377–390. [Google Scholar] [CrossRef]
- Gbotosho, O.T.; Kapetanaki, M.G.; Kato, G.J. The Worst Things in Life are Free: The Role of Free Heme in Sickle Cell Disease. Front. Immunol. 2020, 11, 561917. [Google Scholar] [CrossRef]
- Roumenina, L.T.; Rayes, J.; Lacroix-Desmazes, S.; Dimitrov, J.D. Heme: Modulator of Plasma Systems in Hemolytic Diseases. Trends Mol. Med. 2016, 22, 200–213. [Google Scholar] [CrossRef]
- Pawluczkowycz, A.W.; Lindorfer, M.A.; Waitumbi, J.N.; Taylor, R.P. Hematin promotes complement alternative pathway-mediated deposition of C3 activation fragments on human erythrocytes: Potential implications for the pathogenesis of anemia in malaria. J. Immunol. 2007, 179, 5543–5552. [Google Scholar] [CrossRef] [PubMed]
- Frimat, M.; Tabarin, F.; Dimitrov, J.D.; Poitou, C.; Halbwachs-Mecarelli, L.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement activation by heme as a secondary hit for atypical hemolytic uremic syndrome. Blood 2013, 122, 282–292. [Google Scholar] [CrossRef] [PubMed]
- Merle, N.S.; Grunenwald, A.; Rajaratnam, H.; Gnemmi, V.; Frimat, M.; Figueres, M.L.; Knockaert, S.; Bouzekri, S.; Charue, D.; Noe, R.; et al. Intravascular hemolysis activates complement via cell-free heme and heme-loaded microvesicles. JCI Insight 2018, 3, e96910. [Google Scholar] [CrossRef] [PubMed]
- Merle, N.S.; Paule, R.; Leon, J.; Daugan, M.; Robe-Rybkine, T.; Poillerat, V.; Torset, C.; Fremeaux-Bacchi, V.; Dimitrov, J.D.; Roumenina, L.T. P-selectin drives complement attack on endothelium during intravascular hemolysis in TLR-4/heme-dependent manner. Proc. Natl. Acad. Sci. USA 2019, 116, 6280–6285. [Google Scholar] [CrossRef]
- McIntyre, J.A. The appearance and disappearance of antiphospholipid autoantibodies subsequent to oxidation—Reduction reactions. Thromb. Res. 2004, 114, 579–587. [Google Scholar] [CrossRef]
- McIntyre, J.A.; Wagenknecht, D.R.; Faulk, W.P. Autoantibodies unmasked by redox reactions. J. Autoimmun. 2005, 24, 311–317. [Google Scholar] [CrossRef]
- McIntyre, J.A.; Wagenknecht, D.R.; Faulk, W.P. Redox-reactive autoantibodies: Detection and physiological relevance. Autoimmun. Rev. 2006, 5, 76–83. [Google Scholar] [CrossRef]
- Dimitrov, J.D.; Roumenina, L.T.; Doltchinkova, V.R.; Mihaylova, N.M.; Lacroix-Desmazes, S.; Kaveri, S.V.; Vassilev, T.L. Antibodies use heme as a cofactor to extend their pathogen elimination activity and to acquire new effector functions. J. Biol. Chem. 2007, 282, 26696–26706. [Google Scholar] [CrossRef]
- Dimitrov, J.D.; Planchais, C.; Scheel, T.; Ohayon, D.; Mesnage, S.; Berek, C.; Kaveri, S.V.; Lacroix-Desmazes, S. A cryptic polyreactive antibody recognizes distinct clades of HIV-1 glycoprotein 120 by an identical binding mechanism. J. Biol. Chem. 2014, 289, 17767–17779. [Google Scholar] [CrossRef]
- McIntyre, J.A.; Faulk, W.P. Redox-reactive autoantibodies: Biochemistry, characterization, and specificities. Clin. Rev. Allergy Immunol. 2009, 37, 49–54. [Google Scholar] [CrossRef]
- Lecerf, M.; Scheel, T.; Pashov, A.D.; Jarossay, A.; Ohayon, D.; Planchais, C.; Mesnage, S.; Berek, C.; Kaveri, S.V.; Lacroix-Desmazes, S.; et al. Prevalence and gene characteristics of antibodies with cofactor-induced HIV-1 specificity. J. Biol. Chem. 2015, 290, 5203–5213. [Google Scholar] [CrossRef]
- Lecerf, M.; Kanyavuz, A.; Rossini, S.; Dimitrov, J.D. Interaction of clinical-stage antibodies with heme predicts their physiochemical and binding qualities. Commun. Biol. 2021, 4, 391. [Google Scholar] [CrossRef]
- Silva, D.G.H.; Belini Junior, E.; de Almeida, E.A.; Bonini-Domingos, C.R. Oxidative stress in sickle cell disease: An overview of erythrocyte redox metabolism and current antioxidant therapeutic strategies. Free Radic. Biol. Med. 2013, 65, 1101–1109. [Google Scholar] [CrossRef]
- Voskou, S.; Aslan, M.; Fanis, P.; Phylactides, M.; Kleanthous, M. Oxidative stress in beta-thalassaemia and sickle cell disease. Redox Biol. 2015, 6, 226–239. [Google Scholar] [CrossRef]
- Nagababu, E.; Rifkind, J.M. Heme degradation by reactive oxygen species. Antioxid. Redox Signal. 2004, 6, 967–978. [Google Scholar] [CrossRef]
- Nagababu, E.; Fabry, M.E.; Nagel, R.L.; Rifkind, J.M. Heme degradation and oxidative stress in murine models for hemoglobinopathies: Thalassemia, sickle cell disease and hemoglobin C disease. Blood Cells Mol. Dis. 2008, 41, 60–66. [Google Scholar] [CrossRef]
- Kremer, M.L. The reaction of hemin with H2O2. Eur. J. Biochem. 1989, 185, 651–658. [Google Scholar] [CrossRef]
- Jones, P.; Mantle, D.; Wilson, I. Peroxidase-like activities of iron(III)-porphyrins: Kinetics of the reduction of a peroxidatically active derivative of deuteroferriheme by phenols. J. Inorg. Biochem. 1982, 17, 293–304. [Google Scholar] [CrossRef]
- Vlasova, I.I. Peroxidase Activity of Human Hemoproteins: Keeping the Fire under Control. Molecules 2018, 23, 2561. [Google Scholar] [CrossRef]
- Alayash, A.I. Oxidation reactions of cellular and acellular hemoglobins: Implications for human health. Front. Med. Technol. 2022, 4, 1068972. [Google Scholar] [CrossRef]
- Giulivi, C.; Cadenas, E. The reaction of ascorbic acid with different heme iron redox states of myoglobin. Antioxidant and prooxidant aspects. FEBS Lett. 1993, 332, 287–290. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.B.; Hatzikonstantinou, H.; Herries, D.G. The role of peroxide in haem degradation. A study of the oxidation of ferrihaems by hydrogen peroxide. Biochem. J. 1978, 174, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, W.H.; Harris, T.M.; Guengerich, F.P. Characterization of the enzymatic and nonenzymatic peroxidative degradation of iron porphyrins and cytochrome P-450 heme. Biochemistry 1985, 24, 3254–3263. [Google Scholar] [CrossRef] [PubMed]
- Ortiz de Montellano, P.R. Free radical modification of prosthetic heme groups. Pharmacol. Ther. 1990, 48, 95–120. [Google Scholar] [CrossRef]
- Nagababu, E.; Rifkind, J.M. Formation of fluorescent heme degradation products during the oxidation of hemoglobin by hydrogen peroxide. Biochem. Biophys. Res. Commun. 1998, 247, 592–596. [Google Scholar] [CrossRef] [PubMed]
- Nagababu, E.; Chrest, F.J.; Rifkind, J.M. The origin of red cell fluorescence caused by hydrogen peroxide treatment. Free Radic. Biol. Med. 2000, 29, 659–663. [Google Scholar] [CrossRef]
- Goodarzi, M.; Moosavi-Movahedi, A.A.; Habibi-Rezaei, M.; Shourian, M.; Ghourchian, H.; Ahmad, F.; Farhadi, M.; Saboury, A.A.; Sheibani, N. Hemoglobin fructation promotes heme degradation through the generation of endogenous reactive oxygen species. Spectrochim. Acta A Mol Biomol. Spectrosc. 2014, 130, 561–567. [Google Scholar] [CrossRef]
- Seidel, R.A.; Claudel, T.; Schleser, F.A.; Ojha, N.K.; Westerhausen, M.; Nietzsche, S.; Sponholz, C.; Cuperus, F.; Coldewey, S.M.; Heinemann, S.H.; et al. Impact of higher-order heme degradation products on hepatic function and hemodynamics. J. Hepatol. 2017, 67, 272–281. [Google Scholar] [CrossRef]
- Joerk, A.; Seidel, R.A.; Walter, S.G.; Wiegand, A.; Kahnes, M.; Klopfleisch, M.; Kirmse, K.; Pohnert, G.; Westerhausen, M.; Witte, O.W.; et al. Impact of heme and heme degradation products on vascular diameter in mouse visual cortex. J. Am. Heart Assoc. 2014, 3, e001220. [Google Scholar] [CrossRef]
- Ritter, M.; Neupane, S.; Seidel, R.A.; Steinbeck, C.; Pohnert, G. In vivo and in vitro identification of Z-BOX C—A new bilirubin oxidation end product. Org. Biomol. Chem. 2018, 16, 3553–3555. [Google Scholar] [CrossRef]
- Joerk, A.; Ritter, M.; Langguth, N.; Seidel, R.A.; Freitag, D.; Herrmann, K.H.; Schaefgen, A.; Ritter, M.; Gunther, M.; Sommer, C.; et al. Propentdyopents as Heme Degradation Intermediates Constrict Mouse Cerebral Arterioles and Are Present in the Cerebrospinal Fluid of Patients with Subarachnoid Hemorrhage. Circ. Res. 2019, 124, e101–e114. [Google Scholar] [CrossRef]
- Silva, G.; Jeney, V.; Chora, A.; Larsen, R.; Balla, J.; Soares, M.P. Oxidized hemoglobin is an endogenous proinflammatory agonist that targets vascular endothelial cells. J. Biol. Chem. 2009, 284, 29582–29595. [Google Scholar] [CrossRef]
- Maitra, D.; Byun, J.; Andreana, P.R.; Abdulhamid, I.; Saed, G.M.; Diamond, M.P.; Pennathur, S.; Abu-Soud, H.M. Mechanism of hypochlorous acid-mediated heme destruction and free iron release. Free Radic. Biol. Med. 2011, 51, 364–373. [Google Scholar] [CrossRef]
- Hadzhieva, M.; Vassilev, T.L.; Roumenina, L.T.; Bayry, J.; Kaveri, S.V.; Lacroix-Desmazes, S.; Dimitrov, J.D. Mechanism and Functional Implications of the Heme-Induced Binding Promiscuity of IgE. Biochemistry 2015, 54, 2061–2072. [Google Scholar] [CrossRef]
- Gorshkova, E.N.; Lecerf, M.; Astrakhantseva, I.V.; Vasilenko, E.A.; Starkina, O.V.; Ilyukina, N.A.; Dimitrova, P.A.; Dimitrov, J.D.; Vassilev, T.L. Induced antigen-binding polyreactivity in human serum IgA. Immunobiology 2022, 227, 152213. [Google Scholar] [CrossRef]
- Gorshkova, E.N.; Pashova, S.; Vasilenko, E.A.; Tchurina, T.S.; Razzorenova, E.A.; Starkina, O.V.; Dimitrova, P.; Pashov, A.; Vassilev, T.L. Induced Polyspecificity of Human Secretory Immunoglobulin A Antibodies: Is It Possible to Improve Their Ability to Bind Pathogens? Pharmacology 2022, 107, 341–350. [Google Scholar] [CrossRef]
- Kanyavuz, A.; Marey-Jarossay, A.; Lacroix-Desmazes, S.; Dimitrov, J.D. Breaking the law: Unconventional strategies for antibody diversification. Nat. Rev. Immunol. 2019, 19, 355–368. [Google Scholar] [CrossRef]
- Wilkinson, I.V.L.; Bottlinger, M.; El Harraoui, Y.; Sieber, S.A. Profiling the Heme-Binding Proteomes of Bacteria Using Chemical Proteomics. Angew. Chem. Int. Ed. 2022, e202212111. [Google Scholar] [CrossRef]
- Roumenina, L.T.; Dimitrov, J.D. Assessment of the breadth of binding promiscuity of heme towards human proteins. Biol. Chem. 2022, 403, 1083–1090. [Google Scholar] [CrossRef]
- Homan, R.A.; Jadhav, A.M.; Conway, L.P.; Parker, C.G. A Chemical Proteomic Map of Heme-Protein Interactions. J. Am. Chem. Soc. 2022, 144, 15013–15019. [Google Scholar] [CrossRef]
- Dimitrov, J.D.; Ivanovska, N.D.; Lacroix-Desmazes, S.; Doltchinkova, V.R.; Kaveri, S.V.; Vassilev, T.L. Ferrous ions and reactive oxygen species increase antigen-binding and anti-inflammatory activities of immunoglobulin G. J. Biol. Chem. 2006, 281, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Lecerf, M.; Lacombe, R.; Kanyavuz, A.; Dimitrov, J.D. Functional Changes of Therapeutic Antibodies upon Exposure to Pro-Oxidative Agents. Antibodies 2022, 11, 11. [Google Scholar] [CrossRef] [PubMed]
- Wiatr, M.; Merle, N.S.; Boudhabhay, I.; Poillerat, V.; Rossini, S.; Lecerf, M.; Kaveri, S.V.; Lacroix-Desmazes, S.; Roumenina, L.T.; Dimitrov, J.D. Anti-inflammatory activity of intravenous immunoglobulin through scavenging of heme. Mol. Immunol. 2019, 111, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Minneci, P.C.; Deans, K.J.; Zhi, H.; Yuen, P.S.; Star, R.A.; Banks, S.M.; Schechter, A.N.; Natanson, C.; Gladwin, M.T.; Solomon, S.B. Hemolysis-associated endothelial dysfunction mediated by accelerated NO inactivation by decompartmentalized oxyhemoglobin. J. Clin. Investig. 2005, 115, 3409–3417. [Google Scholar] [CrossRef] [PubMed]
- Vona, R.; Sposi, N.M.; Mattia, L.; Gambardella, L.; Straface, E.; Pietraforte, D. Sickle Cell Disease: Role of Oxidative Stress and Antioxidant Therapy. Antioxidants 2021, 10, 296. [Google Scholar] [CrossRef]
- Cao, H.; Vickers, M.A. Oxidative stress, malaria, sickle cell disease, and innate immunity. Trends Immunol. 2021, 42, 849–851. [Google Scholar] [CrossRef]
- Biswal, S.; Rizwan, H.; Pal, S.; Sabnam, S.; Parida, P.; Pal, A. Oxidative stress, antioxidant capacity, biomolecule damage, and inflammation symptoms of sickle cell disease in children. Hematology 2019, 24, 1–9. [Google Scholar] [CrossRef]
- Amulic, B.; Cazalet, C.; Hayes, G.L.; Metzler, K.D.; Zychlinsky, A. Neutrophil function: From mechanisms to disease. Annu. Rev. Immunol. 2012, 30, 459–489. [Google Scholar] [CrossRef]
- Quismorio, F.P., Jr.; Johnson, C. Serum autoantibodies in patients with sickle cell anemia. Am. J. Med. Sci. 1984, 287, 13–15. [Google Scholar] [CrossRef]
- Zouali, M.; Druilhe, P.; Eyquem, A. IgG-subclass expression of anti-DNA and anti-ribonucleoprotein autoantibodies in human malaria. Clin. Exp. Immunol. 1986, 66, 273–278. [Google Scholar]
- Baethge, B.A.; Bordelon, T.R.; Mills, G.M.; Bowen, L.M.; Wolf, R.E.; Bairnsfather, L. Antinuclear antibodies in sickle cell disease. Acta Haematol. 1990, 84, 186–189. [Google Scholar] [CrossRef]
- Kucuk, O.; Gilman-Sachs, A.; Beaman, K.; Lis, L.J.; Westerman, M.P. Antiphospholipid antibodies in sickle cell disease. Am. J. Hematol. 1993, 42, 380–383. [Google Scholar] [CrossRef]
- Jakobsen, P.H.; Morris-Jones, S.D.; Hviid, L.; Theander, T.G.; Hoier-Madsen, M.; Bayoumi, R.A.; Greenwood, B.M. Anti-phospholipid antibodies in patients with Plasmodium falciparum malaria. Immunology 1993, 79, 653–657. [Google Scholar]
- Bhatnagar, H.; Kala, S.; Sharma, L.; Jain, S.; Kim, K.S.; Pal, R. Serum and organ-associated anti-hemoglobin humoral autoreactivity: Association with anti-Sm responses and inflammation. Eur. J. Immunol. 2011, 41, 537–548. [Google Scholar] [CrossRef]
- Toly-Ndour, C.; Rouquette, A.M.; Obadia, S.; M’Bappe, P.; Lionnet, F.; Hagege, I.; Boussa-Khettab, F.; Tshilolo, L.; Girot, R. High titers of autoantibodies in patients with sickle-cell disease. J. Rheumatol. 2011, 38, 302–309. [Google Scholar] [CrossRef]
- Hommel, B.; Charuel, J.L.; Jaureguiberry, S.; Arnaud, L.; Courtin, R.; Kassab, P.; Prendki, V.; Paris, L.; Ghillani-Dalbin, P.; Thellier, M.; et al. Chronic malaria revealed by a new fluorescence pattern on the antinuclear autoantibodies test. PLoS ONE 2014, 9, e88548. [Google Scholar] [CrossRef]
- Ward, P.A.; Kibukamusoke, J.W. Evidence for soluble immune complexes in the pathogenesis of the glomerulonephritis of quartan malaria. Lancet 1969, 1, 283–285. [Google Scholar] [CrossRef]
- Houba, V. The relation between immunofluorescence patterns of glomerular deposits and subclasses of IgG in patients with nephropathies associated with malaria. Ann. N. Y. Acad. Sci. 1975, 254, 332–333. [Google Scholar] [CrossRef]
- Eisenhut, M. Auto-antibodies and glomerulonephritis in Plasmodium falciparum malaria. Autoimmunity 2010, 43, 640–641. [Google Scholar] [CrossRef]
- Rossini, S.; Noe, R.; Daventure, V.; Lecerf, M.; Justesen, S.; Dimitrov, J.D. V Region of IgG Controls the Molecular Properties of the Binding Site for Neonatal Fc Receptor. J. Immunol. 2020, 205, 2850–2860. [Google Scholar] [CrossRef]
- Andersen, J.T.; Justesen, S.; Berntzen, G.; Michaelsen, T.E.; Lauvrak, V.; Fleckenstein, B.; Buus, S.; Sandlie, I. A strategy for bacterial production of a soluble functional human neonatal Fc receptor. J. Immunol. Methods 2008, 331, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.T.; Justesen, S.; Fleckenstein, B.; Michaelsen, T.E.; Berntzen, G.; Kenanova, V.E.; Daba, M.B.; Lauvrak, V.; Buus, S.; Sandlie, I. Ligand binding and antigenic properties of a human neonatal Fc receptor with mutation of two unpaired cysteine residues. FEBS J. 2008, 275, 4097–4110. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiatr, M.; Hadzhieva, M.; Lecerf, M.; Noé, R.; Justesen, S.; Lacroix-Desmazes, S.; Dragon-Durey, M.-A.; Dimitrov, J.D. Hyperoxidized Species of Heme Have a Potent Capacity to Induce Autoreactivity of Human IgG Antibodies. Int. J. Mol. Sci. 2023, 24, 3416. https://doi.org/10.3390/ijms24043416
Wiatr M, Hadzhieva M, Lecerf M, Noé R, Justesen S, Lacroix-Desmazes S, Dragon-Durey M-A, Dimitrov JD. Hyperoxidized Species of Heme Have a Potent Capacity to Induce Autoreactivity of Human IgG Antibodies. International Journal of Molecular Sciences. 2023; 24(4):3416. https://doi.org/10.3390/ijms24043416
Chicago/Turabian StyleWiatr, Marie, Maya Hadzhieva, Maxime Lecerf, Rémi Noé, Sune Justesen, Sébastien Lacroix-Desmazes, Marie-Agnès Dragon-Durey, and Jordan D. Dimitrov. 2023. "Hyperoxidized Species of Heme Have a Potent Capacity to Induce Autoreactivity of Human IgG Antibodies" International Journal of Molecular Sciences 24, no. 4: 3416. https://doi.org/10.3390/ijms24043416