
Role of Protein Kinase A Activation in the Immune System with an Emphasis on Lipopolysaccharide-Responsive and Beige-like Anchor Protein in B Cells
 , and
, and
Abstract
:1. Introduction
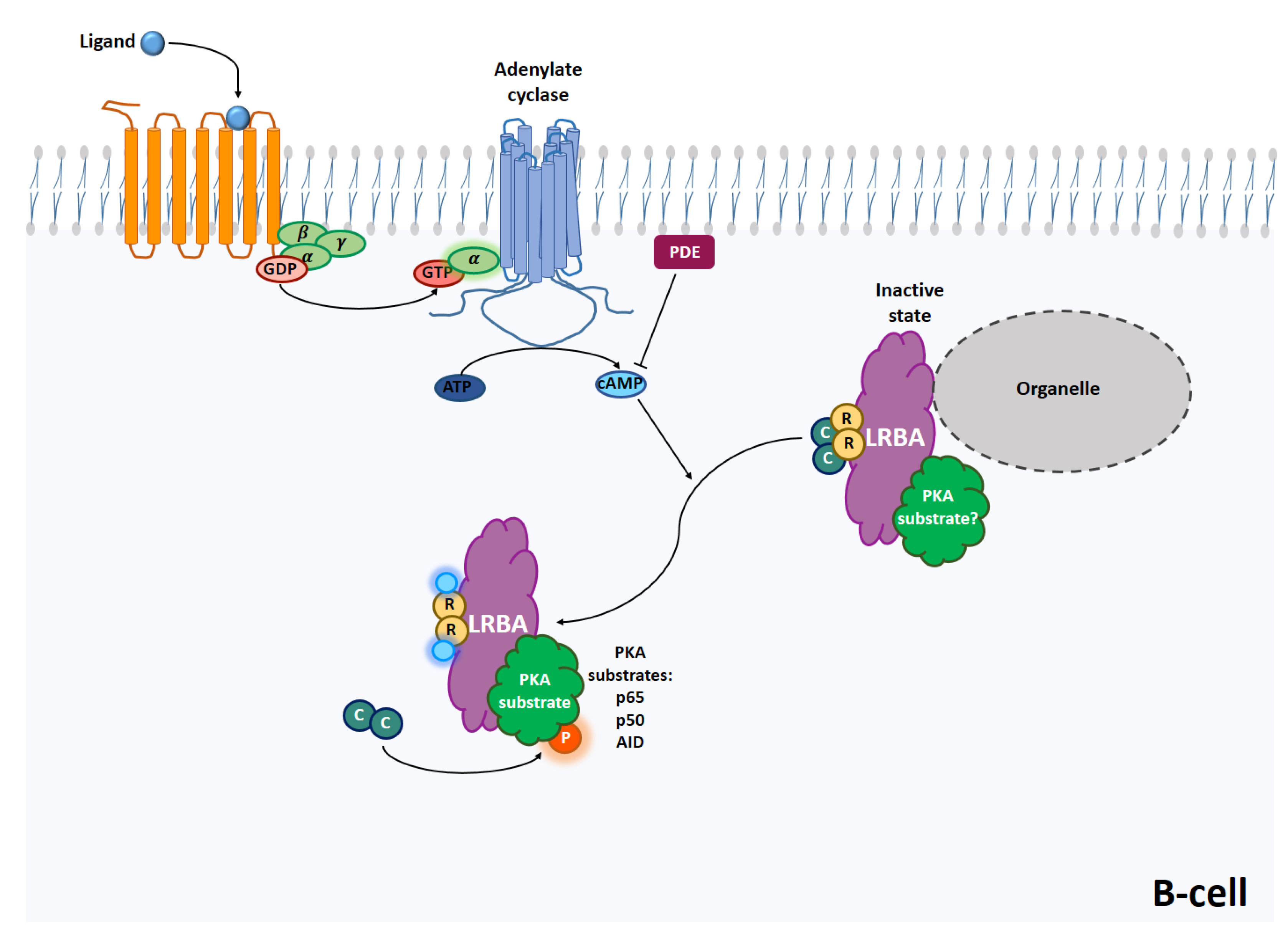
2. Protein Kinase A
2.1. PKA Signaling
2.2. Structure of PKA
2.3. PKA Catalytic Subunits
2.4. Protein Kinase A Regulatory Subunits
3. Cyclic AMP and PKA Function in the Immune System
3.1. Macrophages
3.2. Neutrophils
3.3. Natural Killer Cells
3.4. T Cells
3.5. B Cells
4. PKA Involvement in Immunological Diseases
4.1. PKA in CVID
4.2. PKA in SLE
5. AKAPs Are Involved in Immunological Processes

{kind=link}
| Name | Cell Types | Functions and References |
|---|---|---|
| Ezrin | T, dendritic, and B cells | Activation by immunoreceptors [64], antigen presentation [65], migration [68], and B cell proliferation and differentiation [71]. |
| AKAP149 | Dendritic cells | Antigen presentation [65]. |
| AKAP79 | Dendritic cells | Antigen presentation [65]. Elevated in T cells of patients with SLE [70]. |
| AKAP10 | Alveolar macrophages | LPS-induced NO synthesis, IL-10 and IL-6 synthesis mediated by PGE [69]. |
| AKAP11 | Alveolar macrophages | PGE suppression of TNF-α [69]. |
| AKAP450 | T cells | T cell motility induced via LFA-1 integrins [72]. |
| AKAP9 | T cells | Elevated in T cells of patients with SLE [70]. |
| LRBA | B cells | Physical interaction reported [11]. |
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, S.S.; Yang, J.; Wu, J.; Haste, N.M.; Radzio-Andzelm, E.; Anand, G. PKA: A portrait of protein kinase dynamics. Biochim. Biophys. Acta 2004, 1697, 259–269. [Google Scholar] [CrossRef]
- Johnson, K.R.; Nicodemus-Johnson, J.; Carnegie, G.K.; Danziger, R.S. Molecular evolution of a-kinase anchoring protein (AKAP)-7: Implications in comparative PKA compartmentalization. BMC Evol. Biol. 2012, 12, 125. [Google Scholar] [CrossRef] [PubMed]
- Colledge, M.; Scott, J.D. AKAPs: From structure to function. Trends Cell Biol. 1999, 9, 216–221. [Google Scholar] [CrossRef] [PubMed]
- Aukrust, P.; Aandahl, E.M.; Skålhegg, B.S.; Nordøy, I.; Hansson, V.; Taskén, K.; Frøland, S.S.; Müller, F. Increased activation of protein kinase A type I contributes to the T cell deficiency in common variable immunodeficiency. J. Immunol. 1999, 162, 1178–1185. [Google Scholar] [CrossRef] [PubMed]
- Holm, A.M.; Aukrust, P.; Aandahl, E.M.; Müller, F.; Taskén, K.; Frøland, S.S. Impaired secretion of IL-10 by T cells from patients with common variable immunodeficiency--involvement of protein kinase A type I. J. Immunol. 2003, 170, 5772–5777. [Google Scholar] [CrossRef]
- Mandler, R.; Birch, R.E.; Polmar, S.H.; Kammer, G.M.; Rudolph, S.A. Abnormal adenosine-induced immunosuppression and cAMP metabolism in T lymphocytes of patients with systemic lupus erythematosus. Proc. Natl. Acad. Sci. USA 1982, 79, 7542–7546. [Google Scholar] [CrossRef]
- Hasler, P.; Schultz, L.A.; Kammer, G.M. Defective cAMP-dependent phosphorylation of intact T lymphocytes in active systemic lupus erythematosus. Proc. Natl. Acad. Sci. USA 1990, 87, 1978–1982. [Google Scholar] [CrossRef]
- Kammer, G.M.; Smith, J.A.; Mitchell, R. Capping of human T cell specific determinants: Kinetics of capping and receptor re-expression and regulation by the cytoskeleton. J. Immunol. 1983, 130, 38–44. [Google Scholar] [CrossRef]
- Yazdani, R.; Habibi, S.; Sharifi, L.; Azizi, G.; Abolhassani, H.; Olbrich, P.; Aghamohammadi, A. Common Variable Immunodeficiency: Epidemiology, Pathogenesis, Clinical Manifestations, Diagnosis, Classification, and Management. J. Investig. Allergol. Clin. Immunol. 2020, 30, 14–34. [Google Scholar] [CrossRef]
- Wang, J.W.; Howson, J.; Haller, E.; Kerr, W.G. Identification of a novel lipopolysaccharide-inducible gene with key features of both A kinase anchor proteins and chs1/beige proteins. J. Immunol. 2001, 166, 4586–4595. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Corona, N.C.; Lopez-Ortega, O.; Flores Hermenegildo, J.M.; Berron-Ruiz, L.; Rodriguez-Alba, J.C.; Santos-Argumedo, L.; Lopez-Herrera, G. Lipopolysaccharide-responsive beige-like anchor acts as a cAMP-dependent protein kinase anchoring protein in B cells. Scand. J. Immunol. 2020, 92, e12922. [Google Scholar] [CrossRef] [PubMed]
- Dzeja, P.P.; Terzic, A. Phosphotransfer networks and cellular energetics. J. Exp. Biol. 2003, 206, 2039–2047. [Google Scholar] [CrossRef] [PubMed]
- Manning, G.; Whyte, D.B.; Martinez, R.; Hunter, T.; Sudarsanam, S. The protein kinase complement of the human genome. Science 2002, 298, 1912–1934. [Google Scholar] [CrossRef] [PubMed]
- Walsh, D.A.; Perkins, J.P.; Krebs, E.G. An adenosine 3′,5′-monophosphate-dependant protein kinase from rabbit skeletal muscle. J. Biol. Chem. 1968, 243, 3763–3765. [Google Scholar] [CrossRef]
- Bockaert, J.; Pin, J.P. Molecular tinkering of G protein-coupled receptors: An evolutionary success. EMBO J. 1999, 18, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Sunahara, R.K.; Taussig, R. Isoforms of mammalian adenylyl cyclase: Multiplicities of signaling. Mol. Interv. 2002, 2, 168–184. [Google Scholar] [CrossRef]
- Cooper, D.M.F. Regulation and organization of adenylyl cyclases and cAMP. Biochem. J. 2003, 375, 517–529. [Google Scholar] [CrossRef]
- Bender, A.T.; Beavo, J.A. Cyclic nucleotide phosphodiesterases: Molecular regulation to clinical use. Pharmacol. Rev. 2006, 58, 488–520. [Google Scholar] [CrossRef]
- Smith, F.D.; Reichow, S.L.; Esseltine, J.L.; Shi, D.; Langeberg, L.K.; Scott, J.D.; Gonen, T. Intrinsic disorder within an AKAP-protein kinase A complex guides local substrate phosphorylation. eLife 2013, 2, e01319. [Google Scholar] [CrossRef]
- Søberg, K.; Moen, L.V.; Skålhegg, B.S.; Laerdahl, J.K. Evolution of the cAMP-dependent protein kinase (PKA) catalytic subunit isoforms. PLoS ONE 2017, 12, e0181091. [Google Scholar] [CrossRef] [Green Version]
- Reikhardt, B.A.; Shabanov, P.D. Catalytic Subunit of PKA as a Prototype of the Eukaryotic Protein Kinase Family. Biochem. Biokhimiia 2020, 85, 409–424. [Google Scholar] [CrossRef]
- Taylor, S.S.; Wallbott, M.; Machal, E.M.F.; Søberg, K.; Ahmed, F.; Bruystens, J.; Vu, L.; Baker, B.; Wu, J.; Raimondi, F.; et al. PKA Cβ: A forgotten catalytic subunit of cAMP-dependent protein kinase opens new windows for PKA signaling and disease pathologies. Biochem. J. 2021, 478, 2101–2119. [Google Scholar] [CrossRef] [PubMed]
- Vigil, D.; Blumenthal, D.K.; Heller, W.T.; Brown, S.; Canaves, J.M.; Taylor, S.S.; Trewhella, J. Conformational differences among solution structures of the type Ialpha, IIalpha and IIbeta protein kinase A regulatory subunit homodimers: Role of the linker regions. J. Mol. Biol. 2004, 337, 1183–1194. [Google Scholar] [CrossRef] [PubMed]
- Gerlo, S.; Kooijman, R.; Beck, I.M.; Kolmus, K.; Spooren, A.; Haegeman, G. Cyclic AMP: A selective modulator of NF-κB action. Cell. Mol. Life Sci. 2011, 68, 3823–3841. [Google Scholar] [CrossRef] [PubMed]
- Cadd, G.; McKnight, G.S. Distinct patterns of cAMP-dependent protein kinase gene expression in mouse brain. Neuron 1989, 3, 71–79. [Google Scholar] [CrossRef]
- Hofmann, F.; Beavo, J.A.; Bechtel, P.J.; Krebs, E.G. Comparison of adenosine 3′:5′-monophosphate-dependent protein kinases from rabbit skeletal and bovine heart muscle. J. Biol. Chem. 1975, 250, 7795–7801. [Google Scholar] [CrossRef] [PubMed]
- Manni, S.; Mauban, J.H.; Ward, C.W.; Bond, M. Phosphorylation of the cAMP-dependent protein kinase (PKA) regulatory subunit modulates PKA-AKAP interaction, substrate phosphorylation, and calcium signaling in cardiac cells. J. Biol. Chem. 2008, 283, 24145–24154. [Google Scholar] [CrossRef]
- Banky, P.; Huang, L.J.; Taylor, S.S. Dimerization/docking domain of the type Ialpha regulatory subunit of cAMP-dependent protein kinase. Requirements for dimerization and docking are distinct but overlapping. J. Biol. Chem. 1998, 273, 35048–35055. [Google Scholar] [CrossRef]
- Døskeland, S.O.; Maronde, E.; Gjertsen, B.T. The genetic subtypes of cAMP-dependent protein kinase--functionally different or redundant? Biochim. Biophys. Acta 1993, 1178, 249–258. [Google Scholar] [CrossRef]
- Raker, V.K.; Becker, C.; Steinbrink, K. The cAMP Pathway as Therapeutic Target in Autoimmune and Inflammatory Diseases. Front. Immunol. 2016, 7, 123. [Google Scholar] [CrossRef] [Green Version]
- Papac-Milicevic, N.; Breuss, J.M.; Zaujec, J.; Ryban, L.; Plyushch, T.; Wagner, G.A.; Fenzl, S.; Dremsek, P.; Cabaravdic, M.; Steiner, M.; et al. The interferon stimulated gene 12 inactivates vasculoprotective functions of NR4A nuclear receptors. Circ. Res. 2012, 110, e50–e63. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.A.; Kelly, J.M. Structure, function and evolution of microbial adenylyl and guanylyl cyclases: Microbial purine nucleotide cyclases. Mol. Microbiol. 2004, 52, 1229–1242. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Lamichhane, G.; Gupta, R.; Nolan, S.; Bishai, W.R. Cyclic AMP intoxication of macrophages by a Mycobacterium tuberculosis adenylate cyclase. Nature 2009, 460, 98–102. [Google Scholar] [CrossRef]
- Wirth, J.J.; Kierszenbaum, F. Macrophage mediation of the inhibitory effects of elevated intracellular levels of adenosine-3′:5′ cyclic monophosphate (cAMP) on macrophage-Trypanosoma cruzi association. Int. J. Parasitol. 1984, 14, 401–404. [Google Scholar] [CrossRef]
- Negreiros-Lima, G.L.; Lima, K.M.; Moreira, I.Z.; Jardim, B.L.O.; Vago, J.P.; Galvão, I.; Teixeira, L.C.R.; Pinho, V.; Teixeira, M.M.; Sugimoto, M.A.; et al. Cyclic AMP Regulates Key Features of Macrophages via PKA: Recruitment, Reprogramming and Efferocytosis. Cells 2020, 9, 128. [Google Scholar] [CrossRef]
- Jones, S.L.; Sharief, Y. Asymmetrical protein kinase A activity establishes neutrophil cytoskeletal polarity and enables chemotaxis. J. Leukoc. Biol. 2005, 78, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Bariagaber, A.K.; Whalen, M.M. Decreased adenylyl cyclase and cAMP-dependent protein kinase activities inhibit the cytotoxic function of human natural killer cells. Hum. Immunol. 2003, 64, 866–873. [Google Scholar] [CrossRef]
- Bodor, J.; Bopp, T.; Vaeth, M.; Klein, M.; Serfling, E.; Hünig, T.; Becker, C.; Schild, H.; Schmitt, E. Cyclic AMP underpins suppression by regulatory T cells. Eur. J. Immunol. 2012, 42, 1375–1384. [Google Scholar] [CrossRef]
- Mosenden, R.; Taskén, K. Cyclic AMP-mediated immune regulation—Overview of mechanisms of action in T cells. Cell. Signal. 2011, 23, 1009–1016. [Google Scholar] [CrossRef] [PubMed]
- Shahaf, G.; Zisman-Rozen, S.; Benhamou, D.; Melamed, D.; Mehr, R. B Cell Development in the Bone Marrow Is Regulated by Homeostatic Feedback Exerted by Mature B Cells. Front. Immunol. 2016, 7, 77. [Google Scholar] [CrossRef] [Green Version]
- Klein, U.; Dalla-Favera, R. Germinal centres: Role in B-cell physiology and malignancy. Nat. Rev. Immunol. 2008, 8, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Rajewsky, K. Clonal selection and learning in the antibody system. Nature 1996, 381, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Manis, J.P.; Tian, M.; Alt, F.W. Mechanism and control of class-switch recombination. Trends Immunol. 2002, 23, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Pasqualucci, L.; Guglielmino, R.; Houldsworth, J.; Mohr, J.; Aoufouchi, S.; Polakiewicz, R.; Chaganti, R.S.K.; Dalla-Favera, R. Expression of the AID protein in normal and neoplastic B cells. Blood 2004, 104, 3318–3325. [Google Scholar] [CrossRef]
- Pasqualucci, L.; Kitaura, Y.; Gu, H.; Dalla-Favera, R. PKA-mediated phosphorylation regulates the function of activation-induced deaminase (AID) in B cells. Proc. Natl. Acad. Sci. USA 2006, 103, 395–400. [Google Scholar] [CrossRef]
- Bateman, E.A.L.; Ayers, L.; Sadler, R.; Lucas, M.; Roberts, C.; Woods, A.; Packwood, K.; Burden, J.; Harrison, D.; Kaenzig, N.; et al. T cell phenotypes in patients with common variable immunodeficiency disorders: Associations with clinical phenotypes in comparison with other groups with recurrent infections. Clin. Exp. Immunol. 2012, 170, 202–211. [Google Scholar] [CrossRef]
- Tsoukas, C.D.; Landgraf, B.; Bentin, J.; Valentine, M.; Lotz, M.; Vaughan, J.H.; Carson, D.A. Activation of resting T lymphocytes by anti-CD3 (T3) antibodies in the absence of monocytes. J. Immunol. 1985, 135, 1719–1723. [Google Scholar] [CrossRef]
- Iyer, S.S.; Cheng, G. Role of interleukin 10 transcriptional regulation in inflammation and autoimmune disease. Crit. Rev. Immunol. 2012, 32, 23–63. [Google Scholar] [CrossRef]
- Ameratunga, R.; Woon, S.-T.; Gillis, D.; Koopmans, W.; Steele, R. New diagnostic criteria for common variable immune deficiency (CVID), which may assist with decisions to treat with intravenous or subcutaneous immunoglobulin. Clin. Exp. Immunol. 2013, 174, 203–211. [Google Scholar] [CrossRef]
- O’Garra, A.; Vieira, P.L.; Vieira, P.; Goldfeld, A.E. IL-10-producing and naturally occurring CD4+ Tregs: Limiting collateral damage. J. Clin. Investig. 2004, 114, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Herrera, G.; Tampella, G.; Pan-Hammarström, Q.; Herholz, P.; Trujillo-Vargas, C.M.; Phadwal, K.; Simon, A.K.; Moutschen, M.; Etzioni, A.; Mory, A.; et al. Deleterious mutations in LRBA are associated with a syndrome of immune deficiency and autoimmunity. Am. J. Hum. Genet. 2012, 90, 986–1001. [Google Scholar] [CrossRef] [PubMed]
- Malisan, F.; Brière, F.; Bridon, J.M.; Harindranath, N.; Mills, F.C.; Max, E.E.; Banchereau, J.; Martinez-Valdez, H. Interleukin-10 induces immunoglobulin G isotype switch recombination in human CD40-activated naive B lymphocytes. J. Exp. Med. 1996, 183, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Fujieda, S.; Saxon, A.; Zhang, K. Direct evidence that gamma 1 and gamma 3 switching in human B cells is interleukin-10 dependent. Mol. Immunol. 1996, 33, 1335–1343. [Google Scholar] [CrossRef] [PubMed]
- Zielen, S.; Bauscher, P.; Hofmann, D.; Meuer, S.C. Interleukin 10 and immune restoration in common variable immunodeficiency. Lancet 1993, 342, 750–751. [Google Scholar] [CrossRef]
- Eisenstein, E.M.; Chua, K.; Strober, W. B cell differentiation defects in common variable immunodeficiency are ameliorated after stimulation with anti-CD40 antibody and IL-10. J. Immunol. 1994, 152, 5957–5968. [Google Scholar] [CrossRef]
- Fortuna, G.; Brennan, M.T. Systemic Lupus Erythematosus: Epidemiology, Pathophysiology, Manifestations, and Management. Dent. Clin. N. Am. 2013, 57, 631–655. [Google Scholar] [CrossRef]
- Kammer, G.M.; Mitchell, E. Impaired mobility of human T lymphocyte surface molecules during inactive systemic lupus erythematosus. Relationship to a defective cAMP pathway. Arthritis Rheum. 1988, 31, 88–98. [Google Scholar] [CrossRef]
- Kammer, G.M.; Khan, I.U.; Malemud, C.J. Deficient type I protein kinase A isozyme activity in systemic lupus erythematosus T lymphocytes. J. Clin. Investig. 1994, 94, 422–430. [Google Scholar] [CrossRef]
- Kammer, G.M.; Laxminarayana, D.; Khan, I.U. Mechanisms of deficient type I protein kinase A activity in lupus T lymphocytes. Int. Rev. Immunol. 2004, 23, 225–244. [Google Scholar] [CrossRef]
- Pawson, T.; Scott, J.D. Signaling through scaffold, anchoring, and adaptor proteins. Science 1997, 278, 2075–2080. [Google Scholar] [CrossRef] [Green Version]
- Wong, W.; Scott, J.D. AKAP signalling complexes: Focal points in space and time. Nat. Rev. Mol. Cell Biol. 2004, 5, 959–970. [Google Scholar] [CrossRef] [PubMed]
- Newlon, M.G.; Roy, M.; Morikis, D.; Hausken, Z.E.; Coghlan, V.; Scott, J.D.; Jennings, P.A. The molecular basis for protein kinase A anchoring revealed by solution NMR. Nat. Struct. Biol. 1999, 6, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Skalhegg, B.S.; Tasken, K. Specificity in the cAMP/PKA signaling pathway. Differential expression, regulation, and subcellular localization of subunits of PKA. Front. Biosci. J. Virtual Libr. 2000, 5, D678–D693. [Google Scholar] [CrossRef]
- Gupta, N.; Wollscheid, B.; Watts, J.D.; Scheer, B.; Aebersold, R.; DeFranco, A.L. Quantitative proteomic analysis of B cell lipid rafts reveals that ezrin regulates antigen receptor-mediated lipid raft dynamics. Nat. Immunol. 2006, 7, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Schillace, R.V.; Miller, C.L.; Carr, D.W. AKAPs in lipid rafts are required for optimal antigen presentation by dendritic cells. Immunol. Cell Biol. 2011, 89, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Matsui, K.; Parameswaran, N.; Bagheri, N.; Willard, B.; Gupta, N. Proteomics analysis of the ezrin interactome in B cells reveals a novel association with Myo18aα. J. Proteome Res. 2011, 10, 3983–3992. [Google Scholar] [CrossRef]
- Hsu, R.-M.; Hsieh, Y.-J.; Yang, T.-H.; Chiang, Y.-C.; Kan, C.-Y.; Lin, Y.-T.; Chen, J.-T.; Yu, J.-S. Binding of the extreme carboxyl-terminus of PAK-interacting exchange factor β (βPIX) to myosin 18A (MYO18A) is required for epithelial cell migration. Biochim. Biophys. Acta 2014, 1843, 2513–2527. [Google Scholar] [CrossRef]
- Pore, D.; Gupta, N. The ezrin-radixin-moesin family of proteins in the regulation of B-cell immune response. Crit. Rev. Immunol. 2015, 35, 15–31. [Google Scholar] [CrossRef]
- Kim, S.-H.; Serezani, C.H.; Okunishi, K.; Zaslona, Z.; Aronoff, D.M.; Peters-Golden, M. Distinct protein kinase A anchoring proteins direct prostaglandin E2 modulation of Toll-like receptor signaling in alveolar macrophages. J. Biol. Chem. 2011, 286, 8875–8883. [Google Scholar] [CrossRef]
- Pérez-Lorenzo, M.J.; Galindo, M.; García-González, A.J.; Criado, G. Increased expression of A-kinase anchoring proteins in T cells from systemic lupus erythematosus patients. J. Transl. Med. 2010, 8, P49. [Google Scholar] [CrossRef] [Green Version]
- Pore, D.; Parameswaran, N.; Matsui, K.; Stone, M.B.; Saotome, I.; McClatchey, A.I.; Veatch, S.L.; Gupta, N. Ezrin tunes the magnitude of humoral immunity. J. Immunol. 2013, 191, 4048–4058. [Google Scholar] [CrossRef] [PubMed]
- El Din El Homasany, B.S.; Volkov, Y.; Takahashi, M.; Ono, Y.; Keryer, G.; Delouvée, A.; Looby, E.; Long, A.; Kelleher, D. The scaffolding protein CG-NAP/AKAP450 is a critical integrating component of the LFA-1-induced signaling complex in migratory T cells. J. Immunol. 2005, 175, 7811–7818. [Google Scholar] [CrossRef] [PubMed]
- Gebauer, D.; Li, J.; Jogl, G.; Shen, Y.; Myszka, D.G.; Tong, L. Crystal structure of the PH-BEACH domains of human LRBA/BGL. Biochemistry 2004, 43, 14873–14880. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-W.; Lockey, R. Lipopolysaccharide-responsive beige-like anchor (LRBA), a novel regulator of human immune disorders. Austin J. Clin. Immunol. 2014, 1, 1–9. [Google Scholar]
- Hara, Y.; Ando, F.; Oikawa, D.; Ichimura, K.; Yanagawa, H.; Sakamaki, Y.; Nanamatsu, A.; Fujiki, T.; Mori, S.; Suzuki, S.; et al. LRBA is essential for urinary concentration and body water homeostasis. Proc. Natl. Acad. Sci. USA 2022, 119, e2202125119. [Google Scholar] [CrossRef]
- Lo, B.; Zhang, K.; Lu, W.; Zheng, L.; Zhang, Q.; Kanellopoulou, C.; Zhang, Y.; Liu, Z.; Fritz, J.M.; Marsh, R.; et al. AUTOIMMUNE DISEASE. Patients with LRBA deficiency show CTLA4 loss and immune dysregulation responsive to abatacept therapy. Science 2015, 349, 436–440. [Google Scholar] [CrossRef] [Green Version]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Pérez, D.; Santos-Argumedo, L.; Rodríguez-Alba, J.C.; López-Herrera, G. Role of Protein Kinase A Activation in the Immune System with an Emphasis on Lipopolysaccharide-Responsive and Beige-like Anchor Protein in B Cells. Int. J. Mol. Sci. 2023, 24, 3098. https://doi.org/10.3390/ijms24043098
Pérez-Pérez D, Santos-Argumedo L, Rodríguez-Alba JC, López-Herrera G. Role of Protein Kinase A Activation in the Immune System with an Emphasis on Lipopolysaccharide-Responsive and Beige-like Anchor Protein in B Cells. International Journal of Molecular Sciences. 2023; 24(4):3098. https://doi.org/10.3390/ijms24043098
Chicago/Turabian StylePérez-Pérez, Daniela, Leopoldo Santos-Argumedo, Juan Carlos Rodríguez-Alba, and Gabriela López-Herrera. 2023. "Role of Protein Kinase A Activation in the Immune System with an Emphasis on Lipopolysaccharide-Responsive and Beige-like Anchor Protein in B Cells" International Journal of Molecular Sciences 24, no. 4: 3098. https://doi.org/10.3390/ijms24043098