1. Introduction
Cannabinoids are a group of substances that can bind to cannabinoid receptors (i.e., CB
1 and CB
2) and modulate the activity of the endocannabinoid system (ECS) [
1]. These can be endogenous to the body (endocannabinoids), chemically synthesized, or isolated from the
Cannabis sativa L. plant (phytocannabinoids) [
1,
2]. More than 100 different phytocannabinoids have been identified so far [
3], with THC and cannabidiol (CBD) being the most abundant cannabinoids in the plant [
4]. Other cannabinoids of the same origin include cannabigerol (CBG), cannabinol (CBN), cannabichromene (CBC), and cannabigerovarin (CBGV) [
1], albeit most research has been mainly focused on CBD and THC.
Cannabidiol has been described as exerting a variety of beneficial pharmacological effects, including anti-inflammatory, antioxidant, and neuroprotective properties [
5,
6,
7]. It is currently in the advanced stages of clinical testing for acne treatment and has also been approved for the treatment of severe seizures in epilepsy [
8,
9,
10]. Cannabidiol’s antimicrobial activity also stands out—specifically, its activity against a wide range of Gram-positive bacteria, including a variety of drug-resistant strains such as methicillin-resistant
Staphylococcus aureus (MRSA), multidrug-resistant
Streptococcus pneumoniae,
Enterococcus faecalis, and the anaerobic bacteria
Clostridioides (previously
Clostridium)
difficile and
Cutibacterium (formerly
Propionibacterium)
acnes [
11,
12,
13,
14,
15]. This effect is believed to be associated with a disruption of the bacterial membrane [
11], but further studies are still required to fully elucidate this question.
Cannabigerol acts as the precursor molecule for the most abundant phytocannabinoids, including CBD and THC. It has attracted some interest, with recent reports demonstrating it activates alpha(2)-adrenoceptors, blocks serotonin 1A (5-HT
1A) and CB
1 receptors, and binds to CB
2 receptors, potentially having neuroprotective effects [
16,
17]. Similarly to CBD, CBG has also been studied for its antibacterial properties, with studies showing activity against methicillin-resistant
S. aureus (MRSA) [
18] and planktonic growth of
Streptococcus mutans [
19]. Furthermore, CBG is also capable of interfering with the quorum sensing-mediated processes of
Vibrio harveyi, resulting in the prevention of biofilm formation [
20].
Cannabinoids’ antimicrobial effect upon key pathogens of the skin (e.g.,
Staphylococci,
Streptococci and
Cutibacterium genus) is of note, as certain inflammatory skin conditions are triggered or at higher risk of infection by
S. aureus and
S. pyogenes [
21,
22]. The association between streptococcal infection and guttate psoriasis has been well established, and disease exacerbation has been linked to skin colonization by
S. aureus and
Candida albicans [
21,
23]. Another example is atopic dermatitis, whose severity has been correlated to toxin production by
S. aureus strains, and their superantigens also have an aggravating role [
24].
Considering the current knowledge, we aimed to elucidate CBD and CBG interaction and potential antimicrobial activity upon selected microorganisms, namely on human-skin-specific microorganisms commonly associated with inflammatory skin conditions. Furthermore, the impact of these compounds on the establishment of pathogenic biofilms and their capacity to inhibit keratinocytes’ infection were also a target of this research effort. Finally, considering a potential topical use for skin conditions, dermocosmetic formulations with CBD and CBG were prepared and studied for antimicrobial preservation efficacy and for their impact upon skin microbiota and skin homeostasis.
3. Discussion
Cannabinoids have been described as possessing antimicrobial effects, although the mechanisms of action are not yet fully disclosed [
11]. While CBD’s potential as an antimicrobial has been extensively studied, there is a lack of studies characterizing the activity of CBG [
27]. Here, we demonstrated CBG and CBD’s strong antimicrobial activity, showing their potential as preservative agents and their interaction with human skin microorganisms. To the best of our knowledge, this is the first report with a MIC and MLC determination for both CBG and CBD against
S. pyogenes,
P. aeruginosa, and
E. coli, as well as confirmed biofilm inhibition for both
P. aeruginosa and
E. coli.
Regarding the MIC and MLC assays, our study stands in line with previous reports, where CBD showed a strong activity upon Gram-positive bacteria, albeit with higher concentrations for some of the bacteria tested [
11]. CBD’s impact upon
S. aureus has been described in concentrations ranging from 0.03 to 1.57 μM [
11,
28], while those presented here range from 10 to 75 µM. Additionally, it is interesting that the MICs and MBCs attained for the Gram-positive bacteria were not far superior to the values obtained for the antibiotics, including vancomycin, which is a last-resort antibiotic. Both MIC and MBC were determined for
C. acnes with all tested cannabinoids, except CBD Tocris. Reports have demonstrated CBD’s antimicrobial effect, and clinical trials are ongoing for acne treatment by topical application (NCT03573518) [
29,
30]. Nevertheless, this is the first study reporting CBG’s interaction with this bacterium. Concerning the yeast tested, although Feldman et al. [
31] demonstrated CBD’s ability to inhibit
C. albicans biofilm formation and to disrupt mature biofilm through a multitarget course of action, the author reported neither MIC nor MLC for planktonic
C. albicans. Moreover, no studies have shown CBG’s impact on yeast or fungi, with this being the first time reported.
MIC and MLC values for the same microorganism differed from cannabinoid to cannabinoid. This may be due to differences in the strains used, the methodology employed, or the cannabinoids themselves, as different origins/extraction and purification methods can considerably impact results. For instance, all cannabinoids tested revealed a strong antimicrobial activity, although the values demonstrated in the antimicrobial results differed from molecule to molecule, sometimes significantly. For example, MIC for
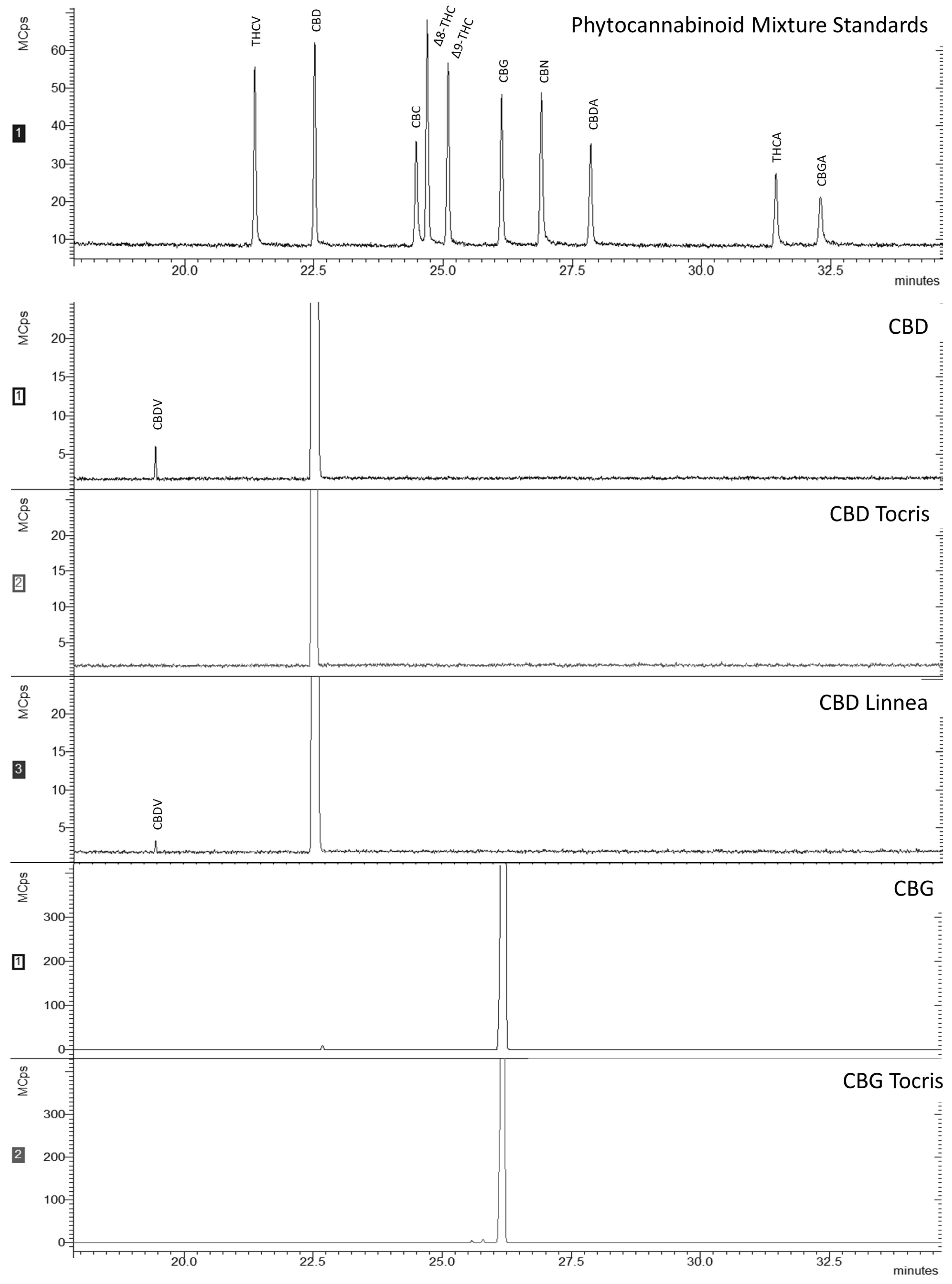
S. aureus was 10 µM for CBD and CBD Linnea (both purified from hemp seeds), whereas for CBD Tocris (prepared by chemical synthesis), it was 7 times higher (75 µM). For CBG Tocris, also prepared by chemical synthesis, MIC was 10 µM, while for CBG, it was 2.5 times higher (25 µM, the concentration corresponding to the subsequent twofold dilution). Those differences are probably due to the different sources of CBDs and to the nature of their impurities. This indication is corroborated by the GC-MS results (
Figure 1), as the spectra for CBD and CBD Linnea also had a peak pertaining to CBDV. CBDV is a propyl analogue of CBD, also found in
C. sativa L. [
32,
33]. Nevertheless, it has been demonstrated that the differences in MIC (or MLC) values are recurrent in studies with cannabinoids, especially when different origins and purity levels, combinations with antibiotics, or cannabis extracts are tested [
12].
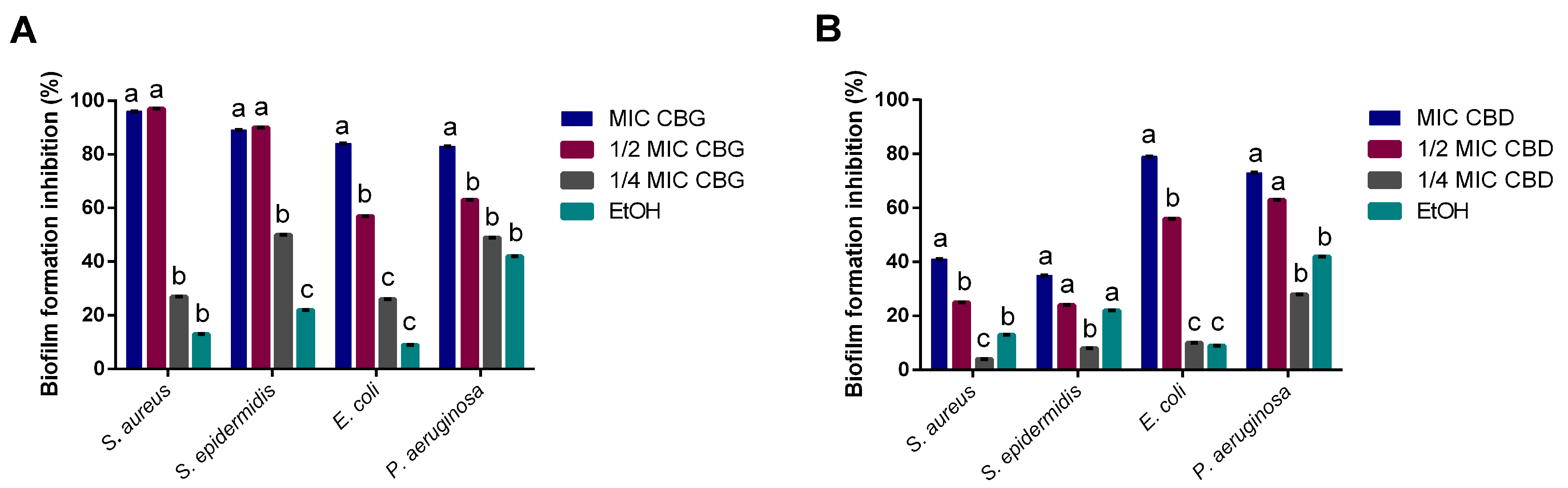
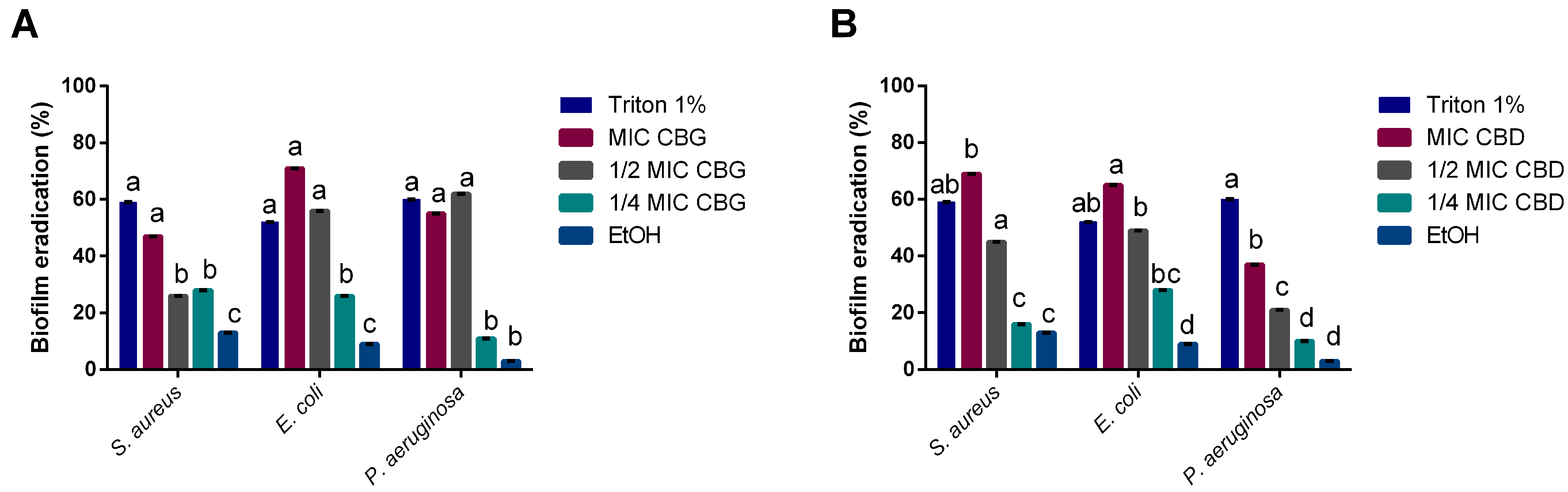
Cannabinoids’ impact on bacterial biofilms was also studied, as the capacity to inhibit biofilm formation or eradicate mature biofilms is of clinical relevance [
34]. CBD’s moderate ability to inhibit biofilm formation for
S. aureus diverges from what has been found in previous reports [
11], showing only a 40% of biomass reduction. This could be a result of the strain used (
S. aureus ATCC 6538™ vs.
S. aureus ATCC 25923™) or the concentrations tested, as the ones that had an effect were much higher than the MIC obtained for
S. aureus in this study. However, it did lead to high inhibitions of biofilm formation for
E. coli and
P. aeruginosa at MIC levels. On the other hand, CBG demonstrated a strong inhibitory activity upon all four tested microorganisms at MICs, standing in line with previous reports of its antibiofilm activity on Gram-positive bacteria [
18,
19]. However, we also observed the capacity of CBG to inhibit Gram-negative biofilm formation, which has not been reported yet.
Our results indicate that CBG has a stronger antimicrobial potential than CBD. CBG and CBD have slight structural differences, namely the alicyclic ring in CBD that in CBG forms an alkyl chain [
13]. This structural difference may explain the discrepancies in the results obtained since these molecules may interact differently with bacterial membrane receptors. Aqawi, Sionov, Gallily, Friedman, and Steinberg [
19] demonstrated that CBG alters the membrane properties by inducing membrane hyperpolarization, decreasing the membrane fluidity while increasing its permeability. Likewise, these molecules seem to lead to a gradient disruption associated with a loss of membrane integrity [
11]. Farha, El-Halfawy, Gale, MacNair, Carfrae, Zhang, Jentsch, Magolan, and Brown [
18] reported that CBG exerts its bactericidal activity by acting on bacteria’s inner membrane. Moreover, the Gram-negative bacteria outer membrane hinders the uptake of both molecules, explaining the discrepancies between MICs for Gram-positive and Gram-negative bacteria tested [
11]. Besides exerting an effect upon planktonic bacteria, CBD and CBG exhibited interesting results regarding biofilm formation and destruction. Reports have suggested CBG and CBD’s ability to act directly on metabolic pathways responsible for regulating biofilms, whilst suppressing metabolic activity and reducing the expression of fundamental genes [
19,
31]. Furthermore, CBG has been described as interfering with quorum sensing (QS) mechanisms [
20], even with no detectable MIC. QS mechanisms are the basis for the development of biofilms, with these mechanisms differing extensively from Gram-positive to Gram-negative bacteria [
35]. One of the pathways through which Gram-negative bacteria form biofilm is the acylated homoserine lactone (AHL) pathway [
35]. There could be an interaction between this pathway and CBD and CBG, which could justify how these cannabinoids exerted biofilm inhibitory activity at sub-MIC values. On a similar note, CBD has been reported to modify the architecture of fungal biofilm through the reduction in exopolysaccharide (EPS) production and consequent thickness of the biofilm [
31].
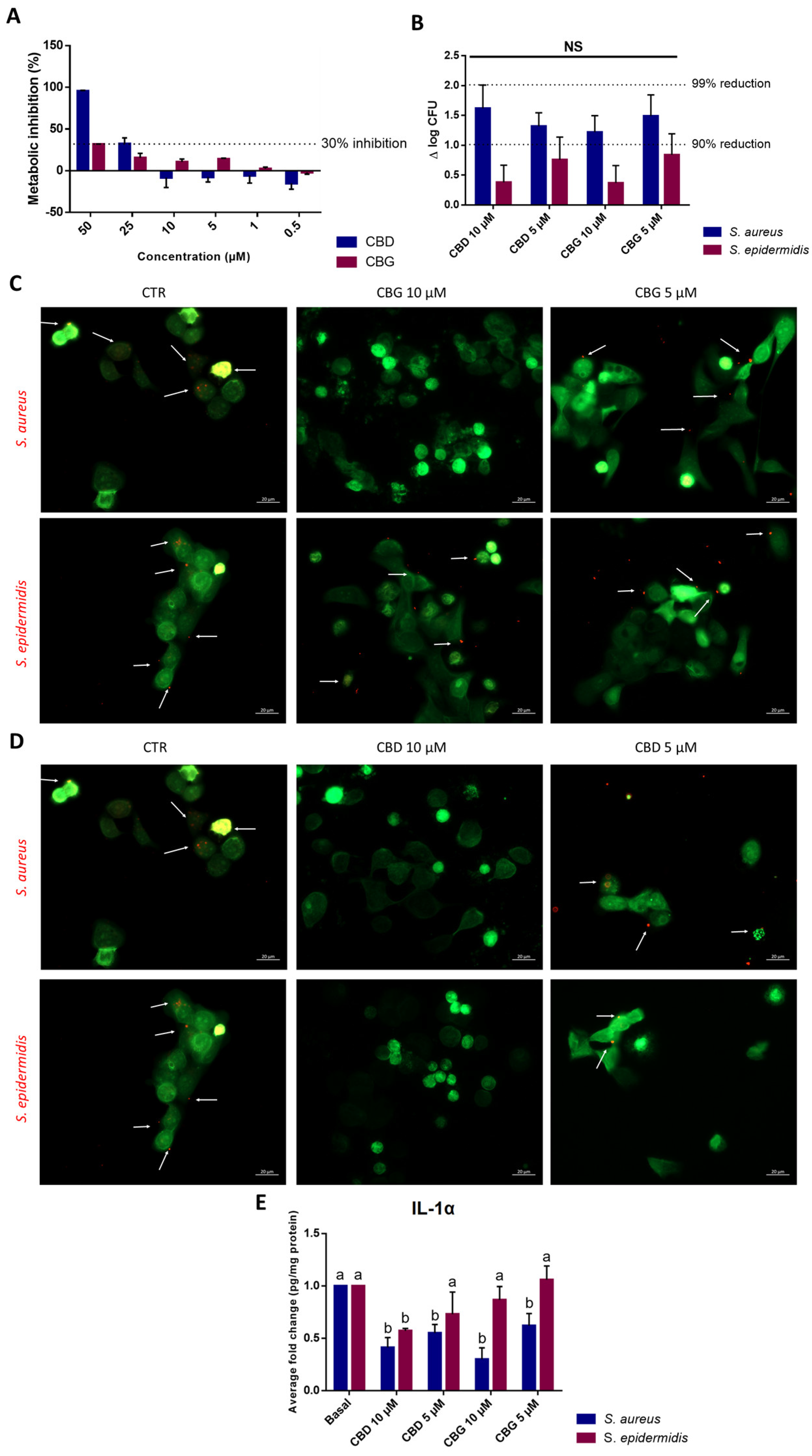
S. epidermidis is more abundantly found in the healthy skin microbiota, while
S. aureus is more frequently associated with a dysbiosis state and different skin disorders [
36]. Moreover, the adherence of bacteria to epithelial cells is an essential step for colonization and infection. Concerning the evaluation of cannabinoids’ impact upon bacterial adhesion to keratinocytes, it is interesting to note that both cannabinoids exerted stronger inhibition against
S. aureus than against
S. epidermidis. As such, the antimicrobial potential and inhibitory activity upon
S. aureus adhesion to skin cells demonstrated by both CBD and CBG, associated with the anti-inflammatory potential as seen on the reduction in IL-1α levels and as described by several authors [
5,
17,
37], could prove a useful alternative to ameliorate symptoms and prevent infections in patients suffering from skin disorders.
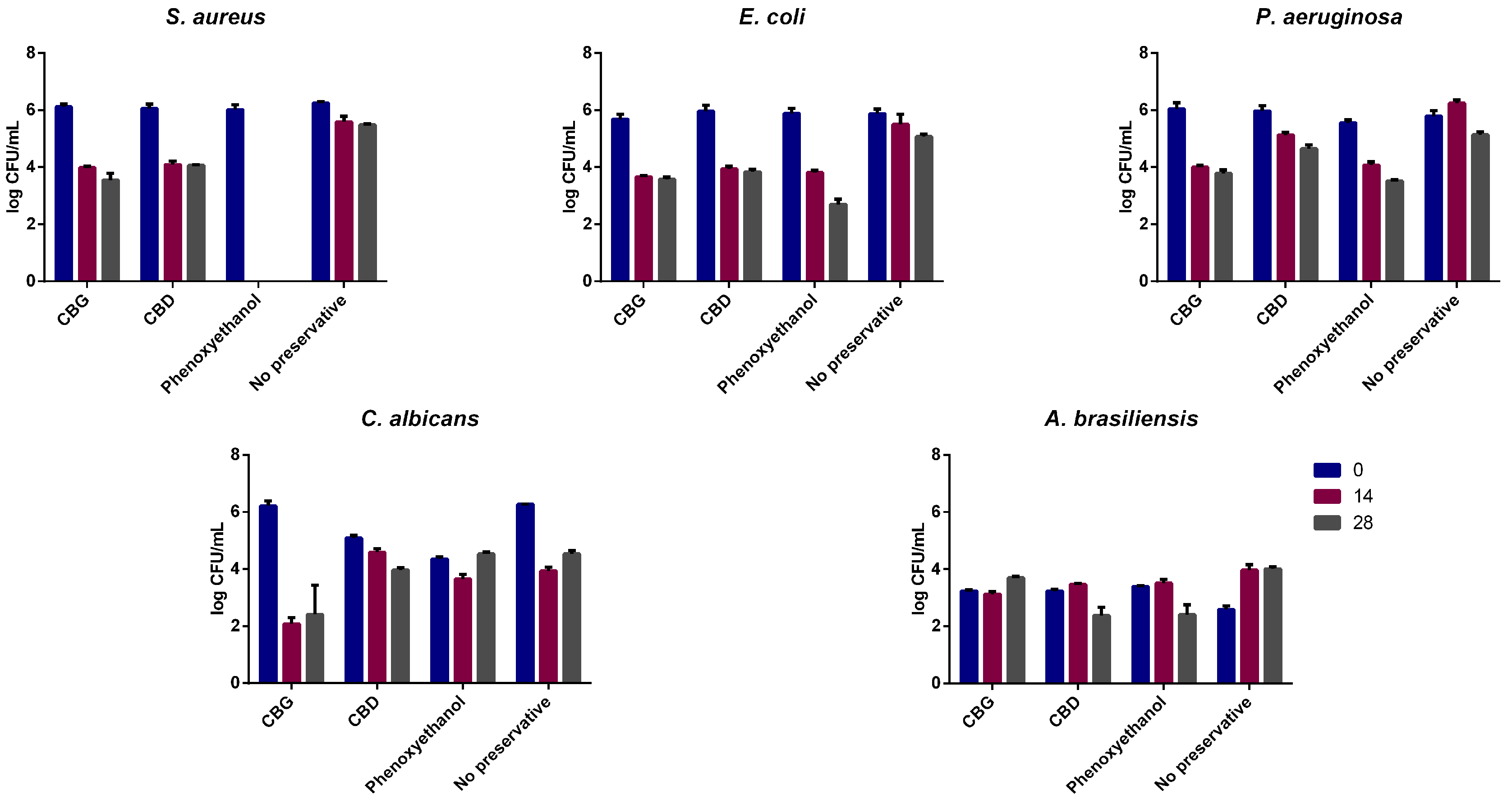
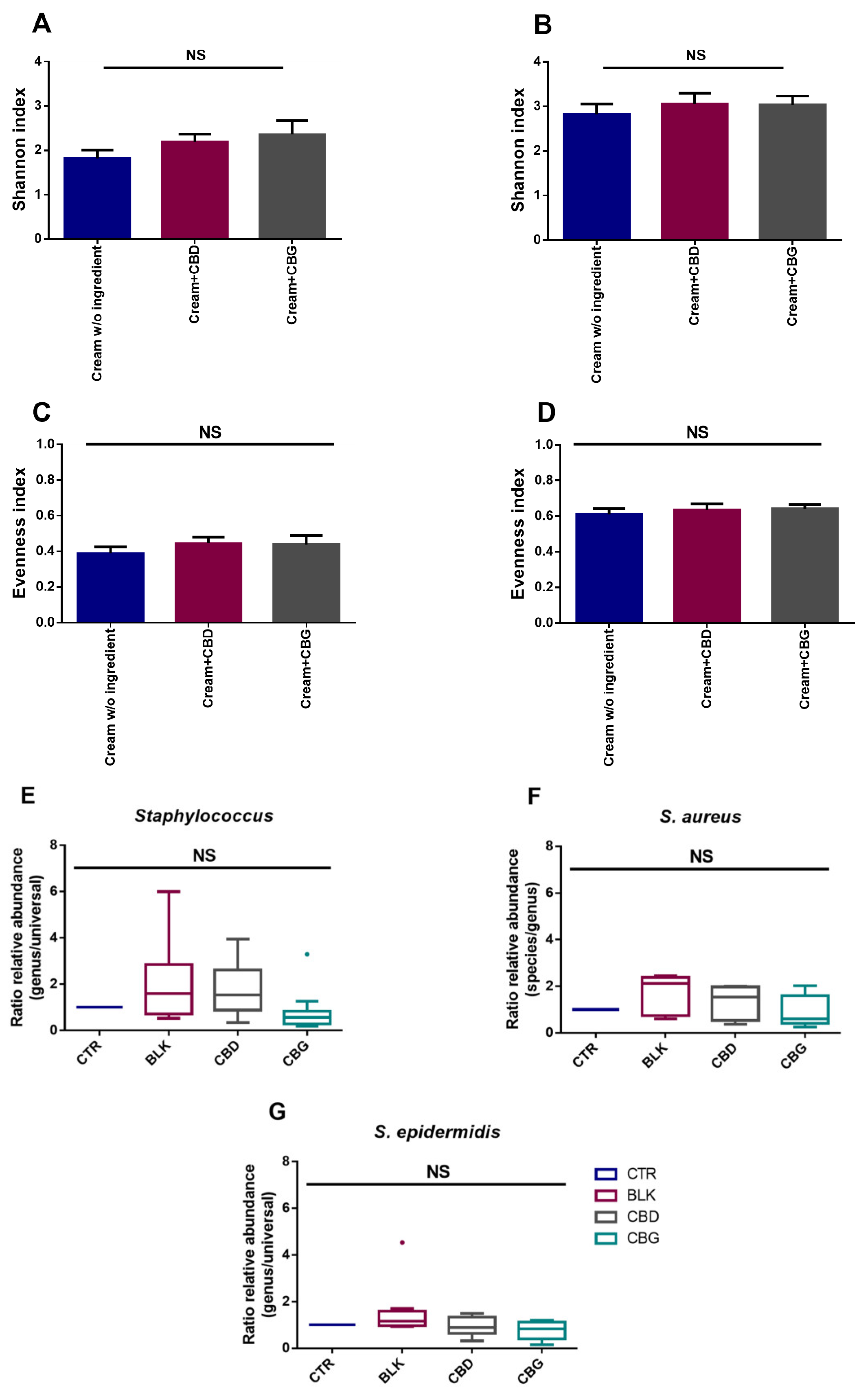
Due to the antimicrobial potential of these cannabinoids and considering a topical/dermatological application, CBD and CBG were also studied for their dermocosmetic formulations’ preservative potential. To the best of our knowledge, this is the first report concerning the use of both cannabinoids as a preservative in a dermocosmetic formulation aimed to be applied topically. CBG yielded better results than CBD as, although CBD passed the criteria for yeasts and molds, it did not lead to a reduction of ≥2 log for P. aeruginosa, even if it had a similar effect to CBG on the MIC/MBC assays. As cannabinoids have also been described as possessing anti-inflammatory activity, their use as multifunctional ingredients in dermatological formulations stands as a possibility. Regarding the skin microbiota, the assays performed demonstrated that both CBD and CBG formulations are microbiota-friendly, not having a significant impact on the alpha diversity of the samples. This is a good indication of the potential use of these cannabinoids in topical applications since they seem to have no significant impact on the skin microbiota of volunteers without skin diseases diagnosed. To understand in more detail their impact on the skin microbiota at a species level, qPCR was performed focusing on detecting S. aureus and S. epidermidis. Although no significant changes between donors were found for either genus or species, there seems to be a decrease in ratio regarding the relative abundance when CBD and CBG are added. Additionally, as the donors did not present skin conditions, only in 4 out of 12 donors was S. aureus detected, which limits the analysis of these cannabinoids’ effect on microorganisms typical of unhealthy skin. As such, the evaluation of phytocannabinoids’ impact on unhealthy skin microbiota, which demonstrates a hegemony of S. aureus, could be of interest in the future.

 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





