
Spent Yeast Waste Streams as a Sustainable Source of Bioactive Peptides for Skin Applications

 , ,
, ,  , , , and
, , , and 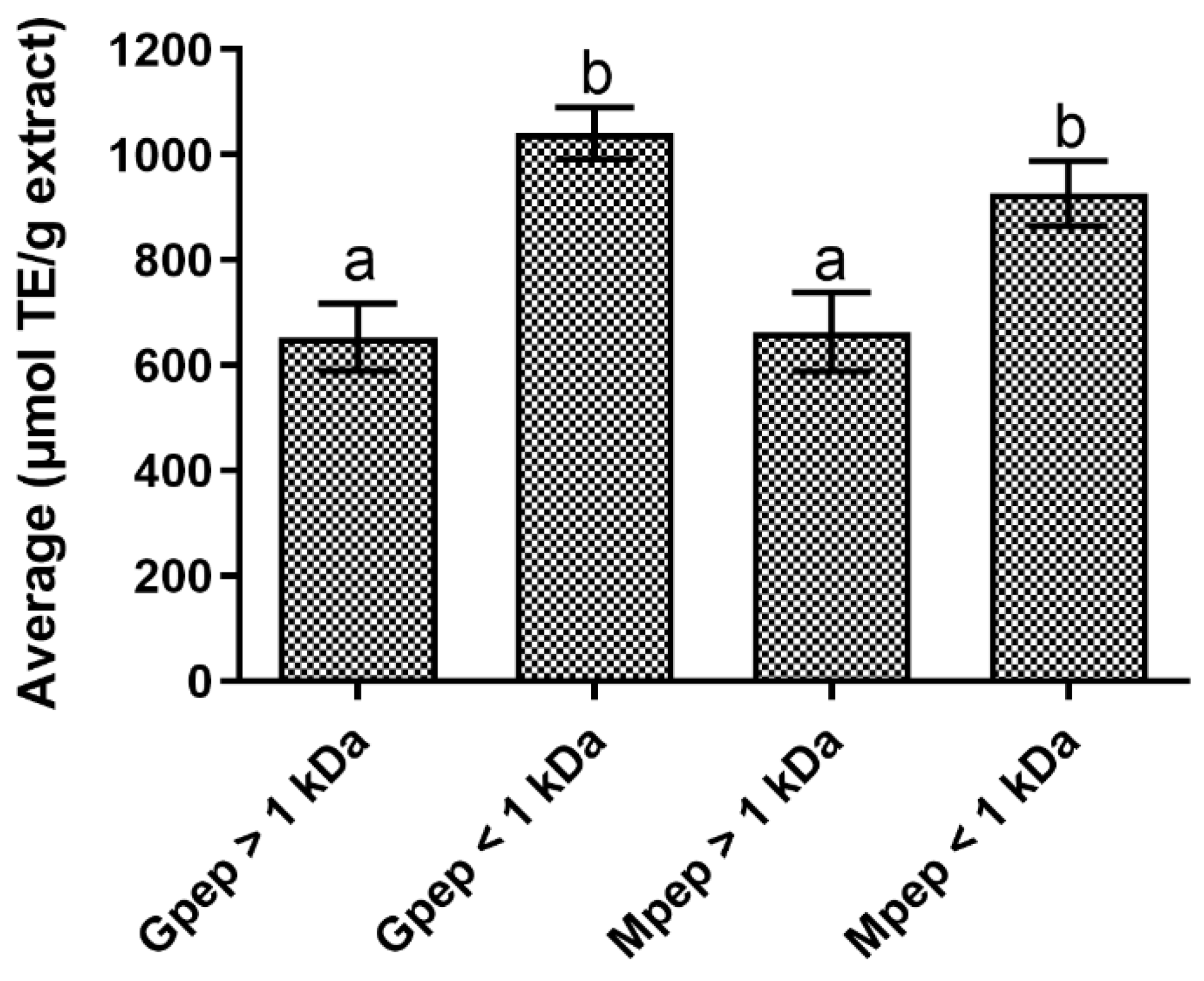{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Antioxidant Activity Results
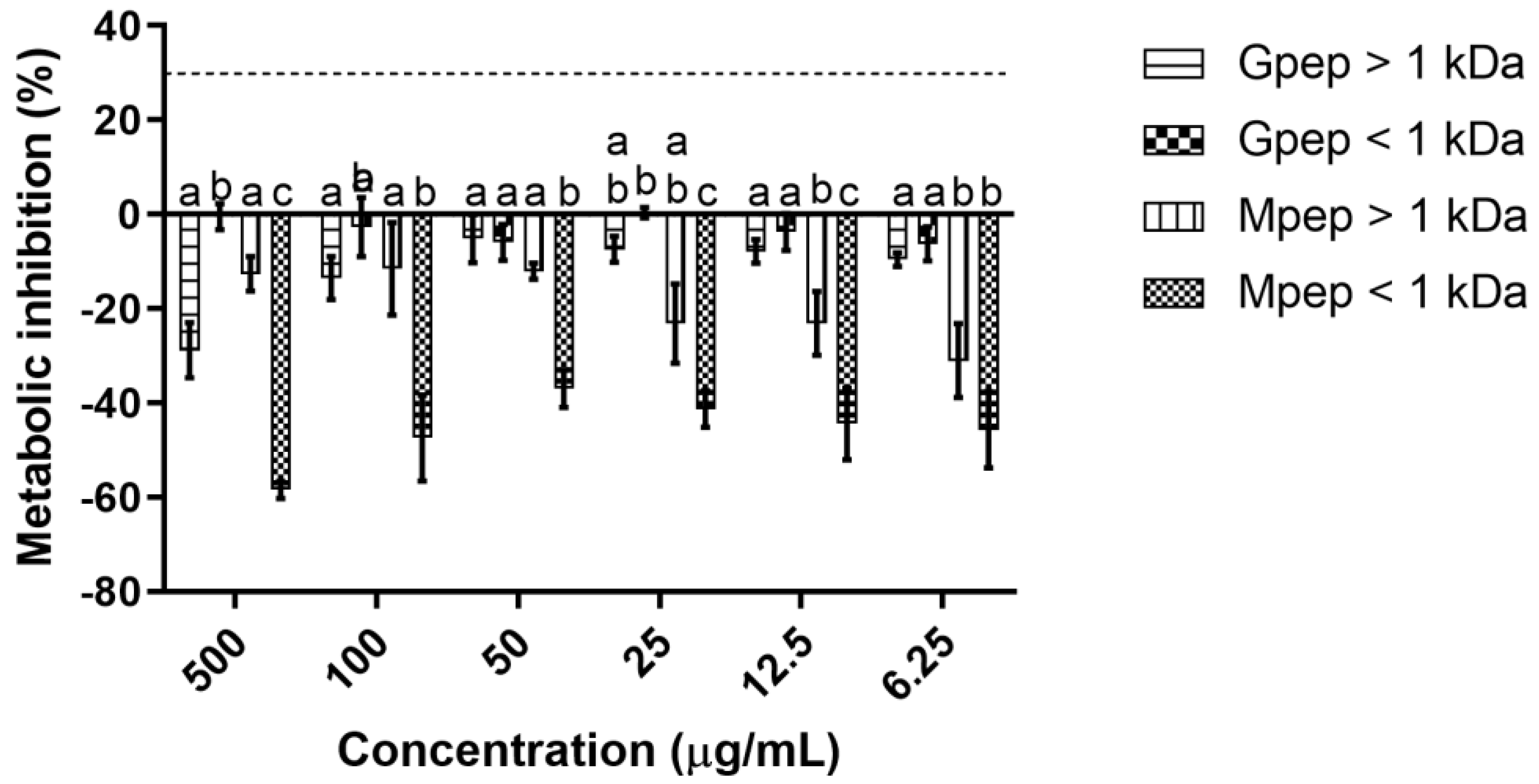
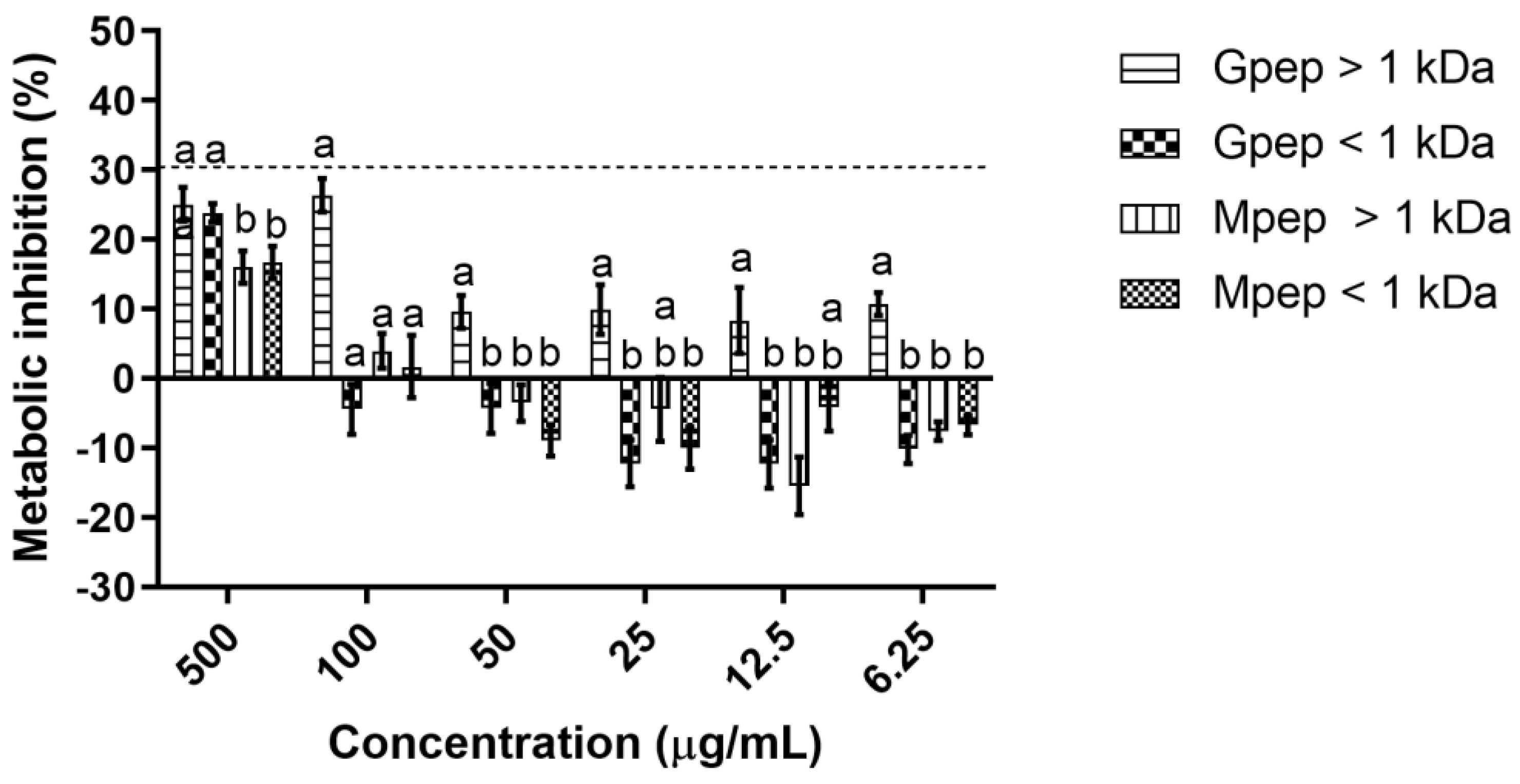
2.2. Cytototixicity Results
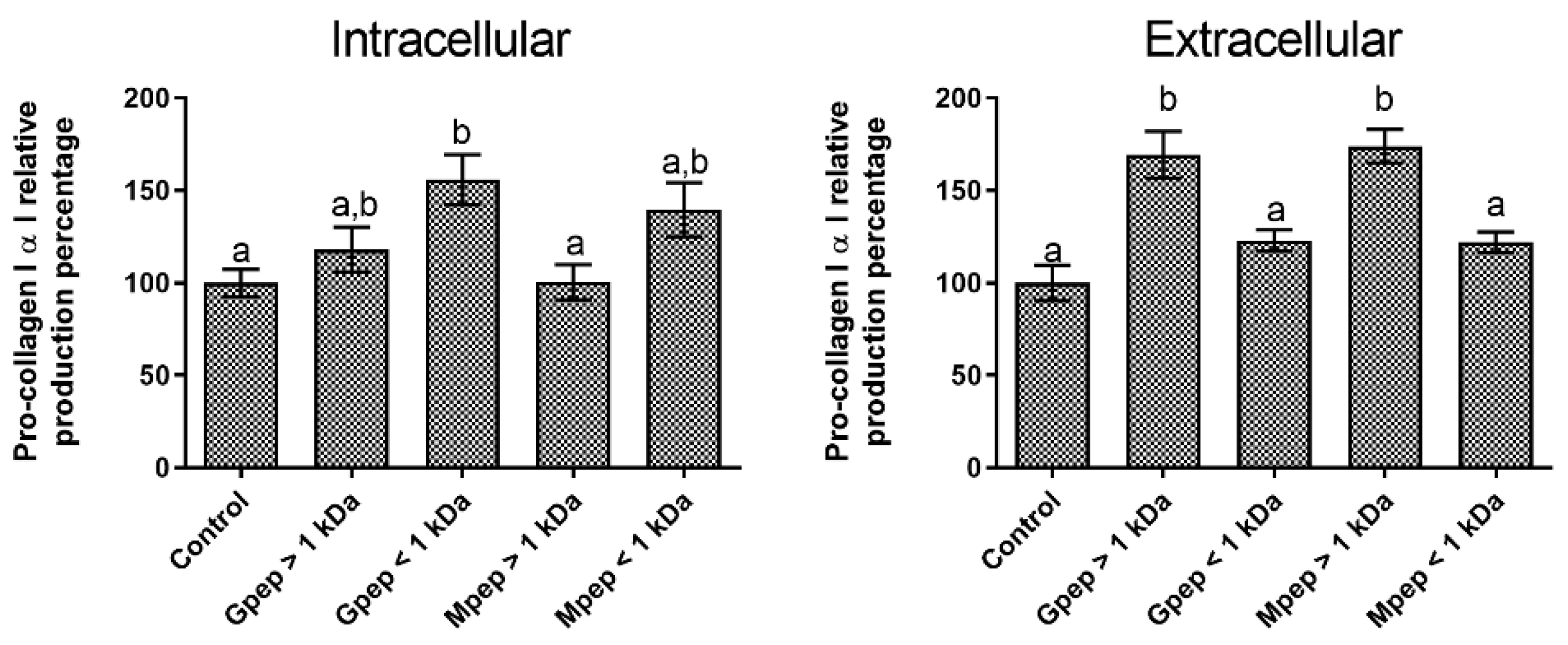
2.3. Pro-Collagen I α I Biosynthesis Results
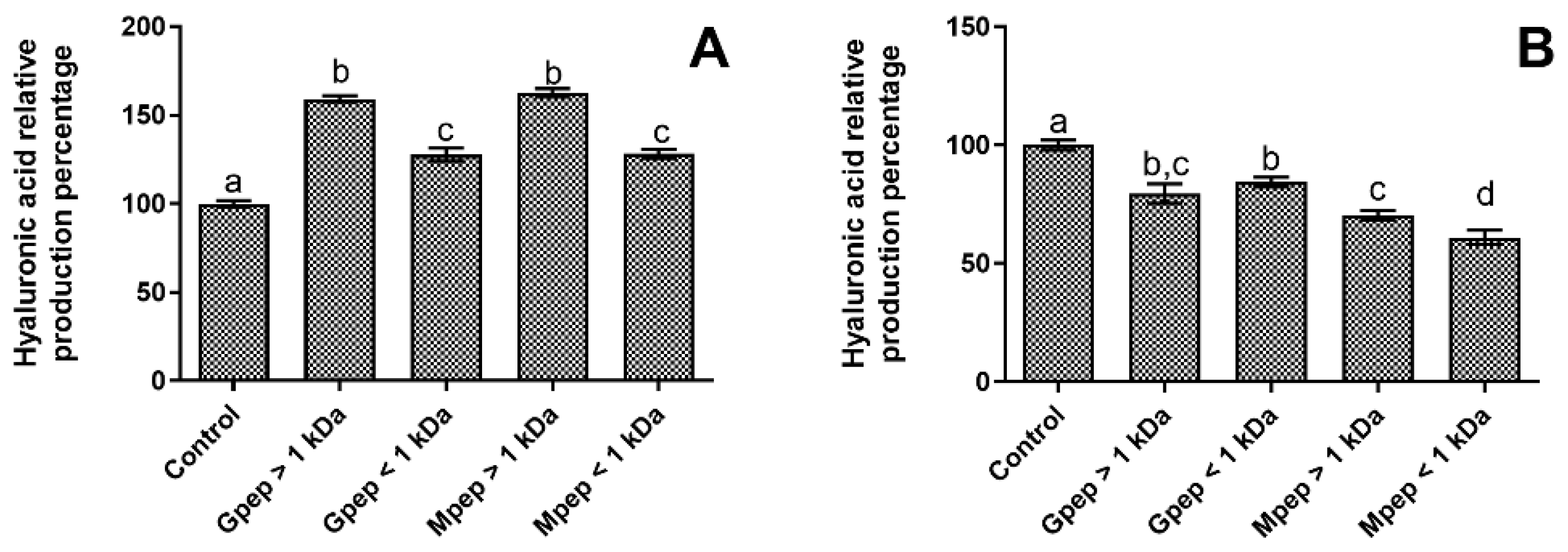
2.4. Hyaluronic Acid Biosynthesis Results
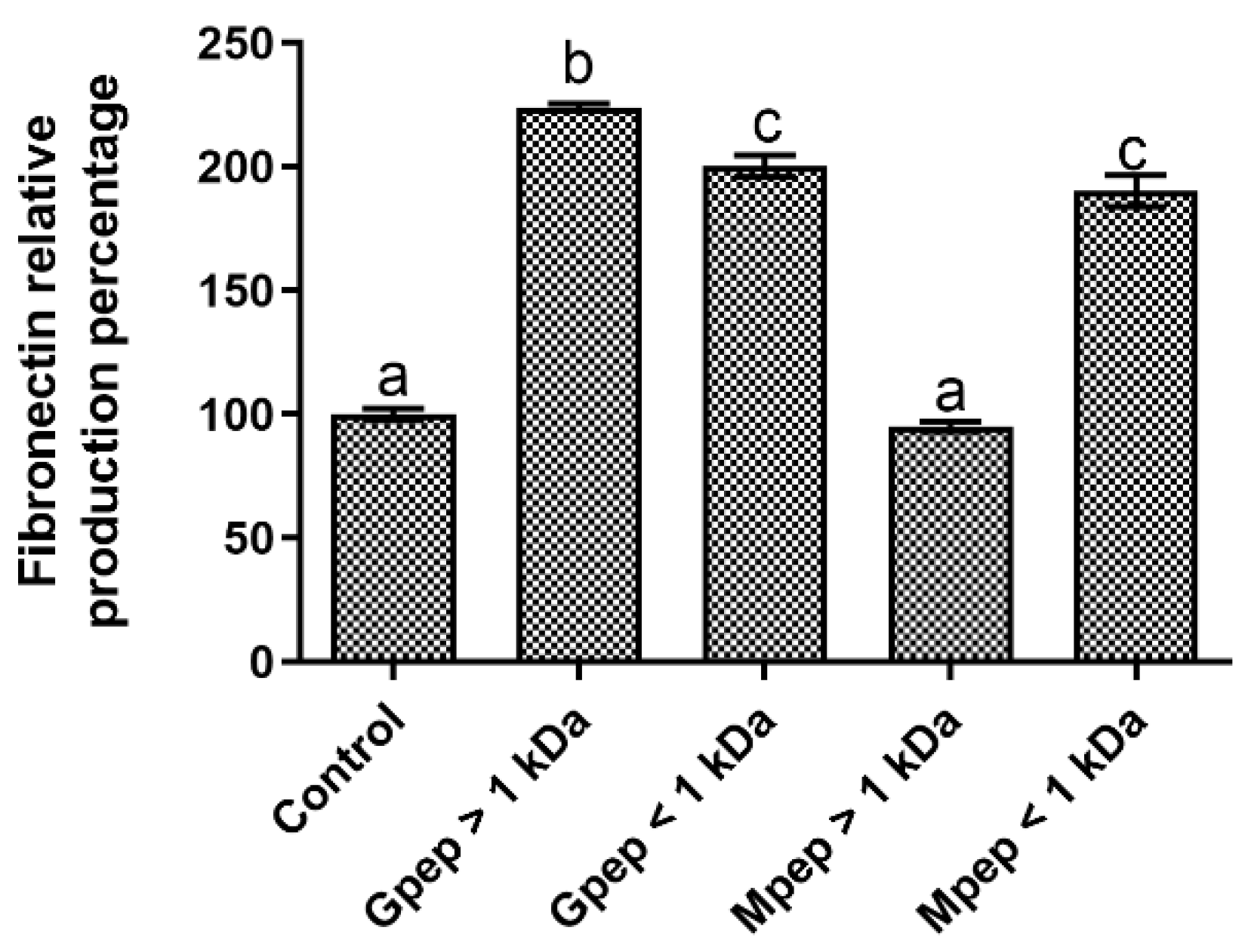
2.5. Fibronectin Biosynthesis Results
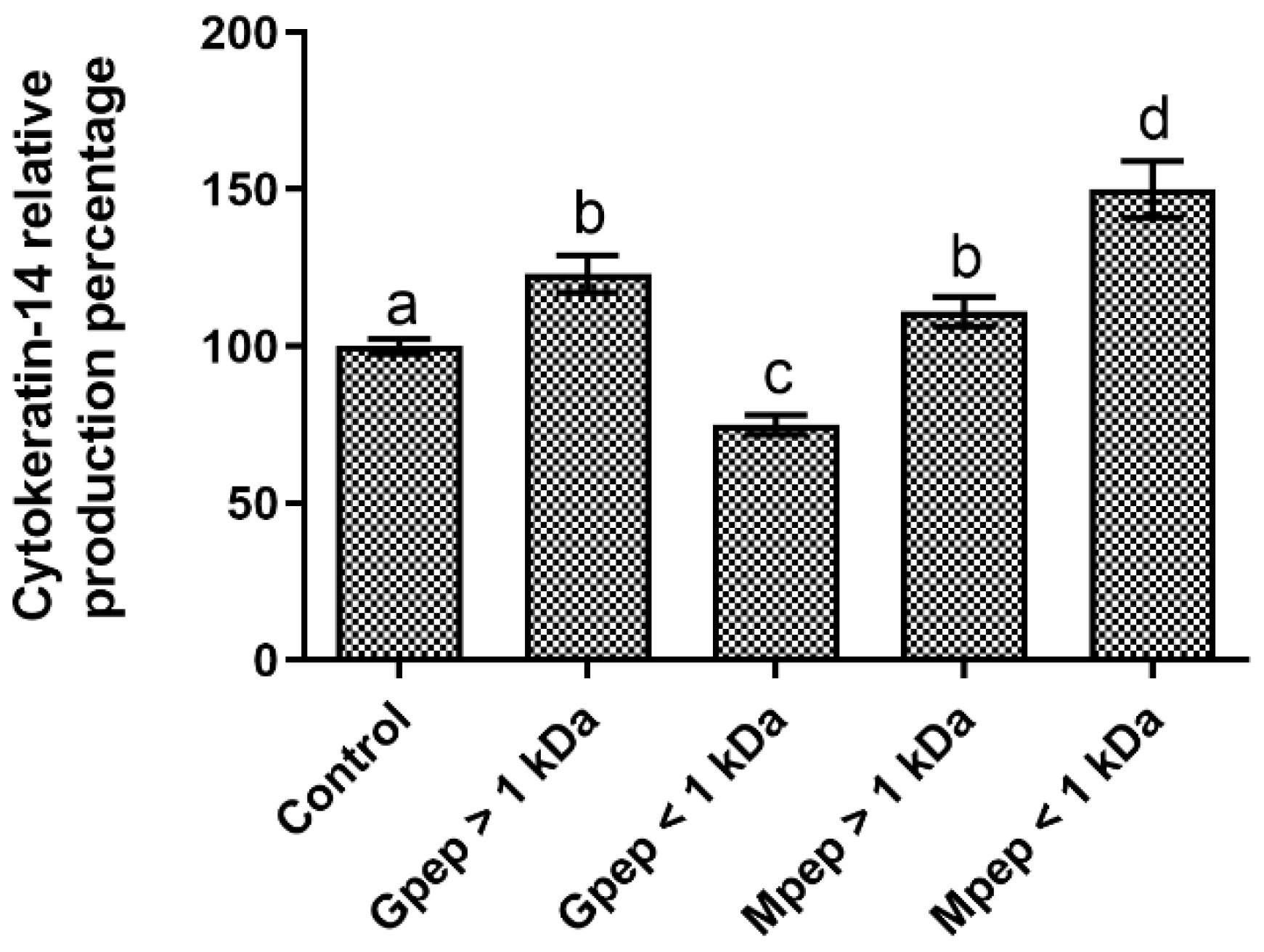
2.6. Cytokeratin-14 Biosynthesis Results
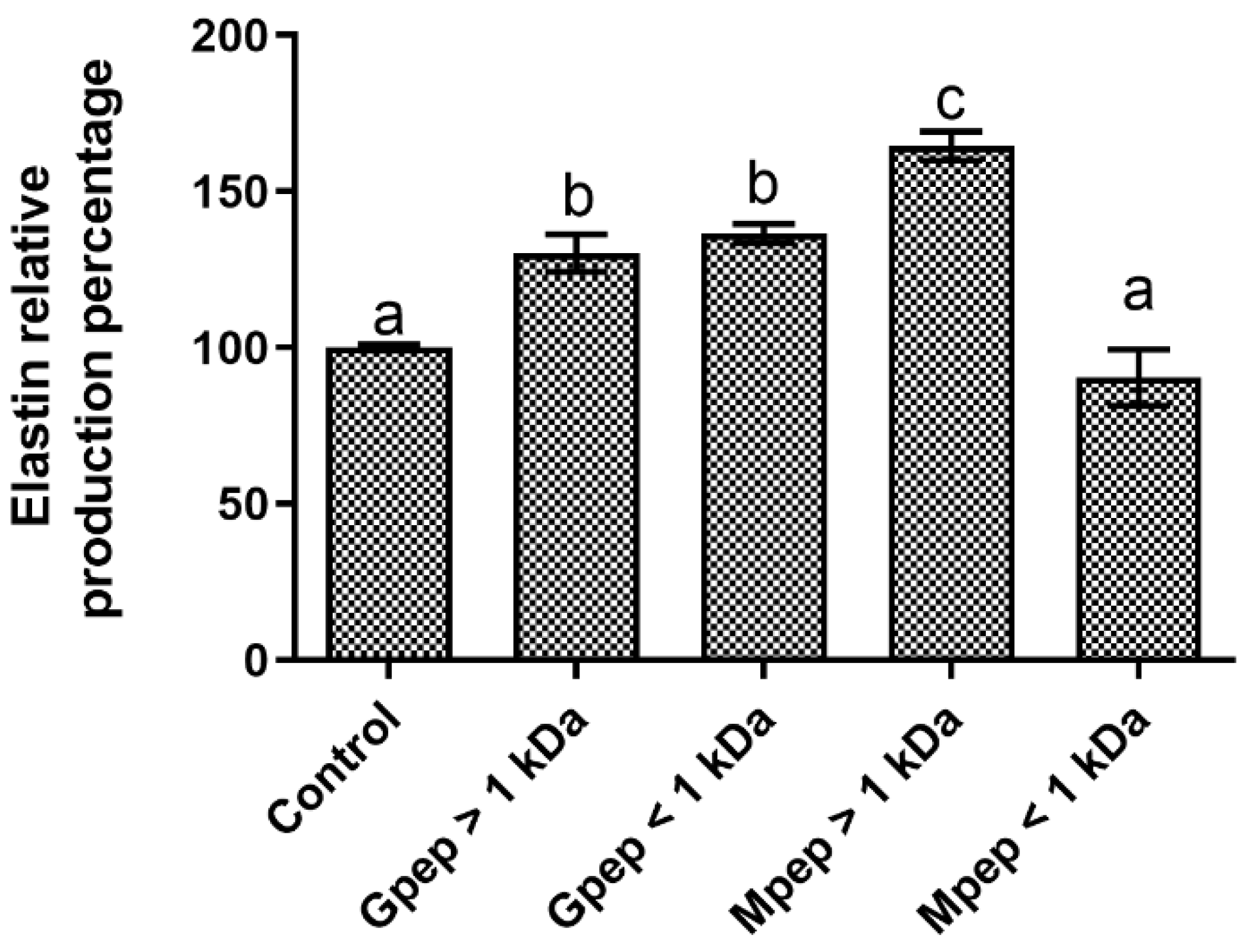
2.7. Elastin Biosynthesis Results
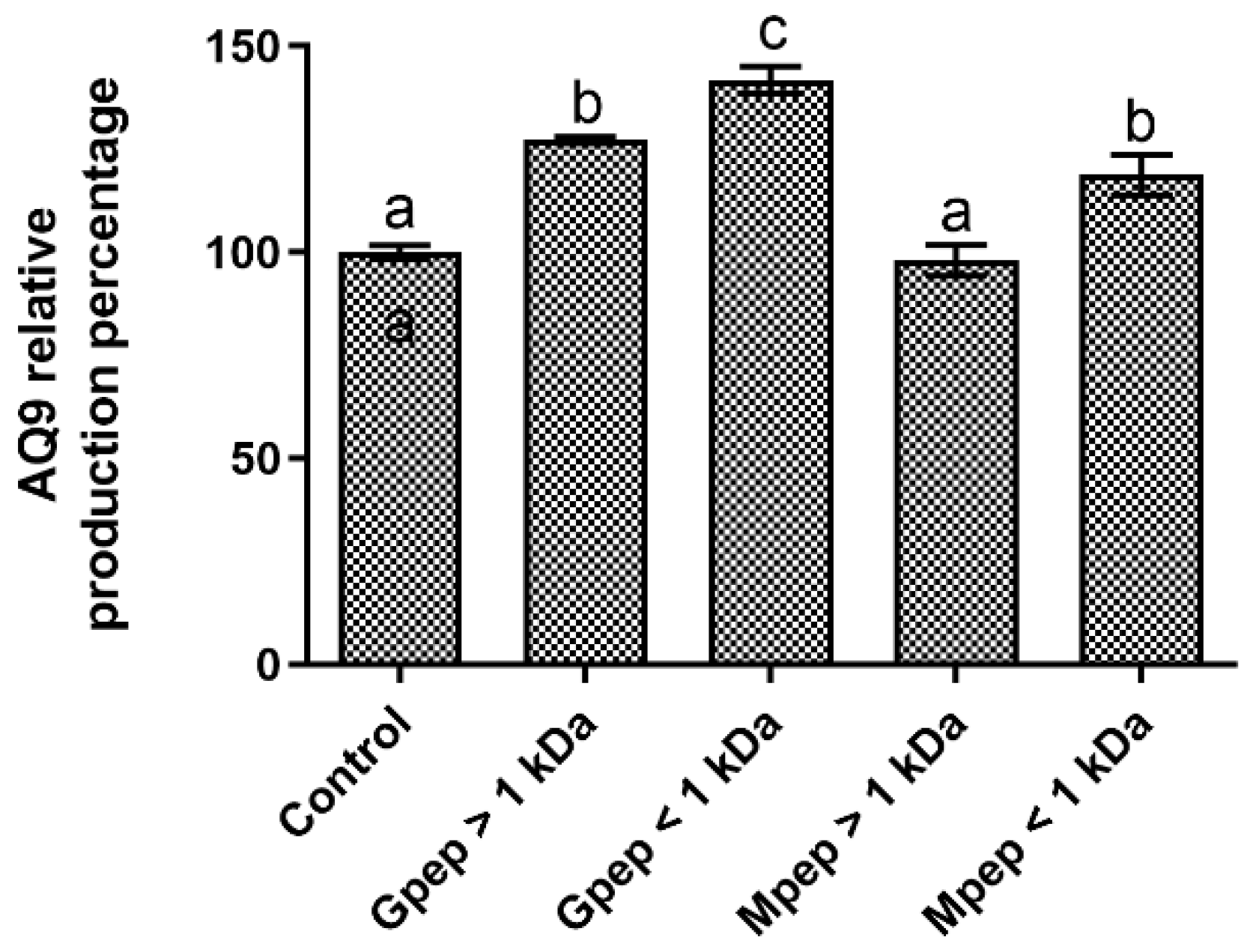
2.8. Aquaporin-9 Quantification
3. Discussion
4. Materials and Methods
4.1. Peptide Fraction Production
4.2. Antioxidant Activity
4.3. Cell lines and Culture Conditions
4.4. Evaluation of the Peptide’s Fractions Cytotoxicity
4.5. Pro-Collagen I α I Biosynthesis and Quantification
4.6. Hyaluronic acid Biosynthesis and Quantification
4.7. Fibronectin Biosynthesis and Quantification
4.8. Cytokeratin-14 Biosynthesis and Quantification
4.9. Elastin Biosynthesis and Quantification
4.10. Aquaporin 9 Biosynthesis and Quantification
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferreira, M.S.; Magalhães, M.C.; Sousa-Lobo, J.M.; Almeida, I.F. Trending Anti-Aging Peptides. Cosmetics 2020, 7, 91. [Google Scholar] [CrossRef]
- Schagen, S.K. Topical Peptide Treatments with Effective Anti-Aging Results. Cosmetics 2017, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Gorouhi, F.; Maibach, H.I. Role of topical peptides in preventing or treating aged skin. Int. J. Cosmet. Sci. 2009, 31, 327–345. [Google Scholar] [CrossRef]
- Mitragotri, S. Modeling skin permeability to hydrophilic and hydrophobic solutes based on four permeation pathways. J. Control. Release 2003, 86, 69–92. [Google Scholar] [CrossRef]
- Ahsan, H. The biomolecules of beauty: Biochemical pharmacology and immunotoxicology of cosmeceuticals. J. Immunoass. Immunochem. 2019, 40, 91–108. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, S.M.; Moghimi, H.R. Skin permeability, a dismissed necessity for anti-wrinkle peptide performance. Int. J. Cosmet. Sci. 2022, 44, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.S.; Pereira, J.O.; Ferreira, C.; Faustino, M.; Durão, J.; Pintado, M.E.; Carvalho, A.P. Peptide-rich extracts from spent yeast waste streams as a source of bioactive compounds for the nutraceutical market. Innov. Food Sci. Emerg. Technol. 2022, 81, 103148. [Google Scholar] [CrossRef]
- Amorim, M.; Marques, C.; Pereira, J.O.; Guardão, L.; Martins, M.J.; Osório, H.; Moura, D.; Calhau, C.; Pinheiro, H.; Pintado, M. Antihypertensive effect of spent brewer yeast peptide. Process Biochem. 2019, 76, 213–218. [Google Scholar] [CrossRef]
- Guo, H.; Guo, S.; Liu, H. Antioxidant activity and inhibition of ultraviolet radiation-induced skin damage of Selenium-rich peptide fraction from selenium-rich yeast protein hydrolysate. Bioorg. Chem. 2020, 105, 104431. [Google Scholar] [CrossRef]
- Lee, H.-S.; Jung, E.Y.; Suh, H.J. Chemical composition and anti-stress effects of yeast hydrolysate. J. Med. Food 2009, 12, 1281–1285. [Google Scholar] [CrossRef]
- Williams, R.; Dias, D.A.; Jayasinghe, N.; Roessner, U.; Bennett, L.E. Beta-glucan-depleted, glycopeptide-rich extracts from Brewer’s and Baker’s yeast (Saccharomyces cerevisiae) lower interferon-gamma production by stimulated human blood cells in vitro. Food Chem. 2016, 197, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Jung, E.Y.; Bae, S.H.; Kwon, K.H.; Kim, J.M.; Suh, H.J. Stimulation of osteoblastic differentiation and mineralization in MC3T3-E1 cells by yeast hydrolysate. Phytother. Res. 2011, 25, 716–723. [Google Scholar] [CrossRef]
- ISO. Biological evaluation of medical devices. In Tests for In Vitro Cytotoxicity; International Organization for Standardization: Geneva, Switzerland, 2009; p. 34. [Google Scholar]
- Montalvo, G.E.B.; Thomaz-Soccol, V.; Vandenberghe, L.P.; Carvalho, J.C.; Faulds, C.B.; Bertrand, E.; Prado, M.R.; Bonatto, S.J.; Soccol, C.R. Arthrospira maxima OF15 biomass cultivation at laboratory and pilot scale from sugarcane vinasse for potential biological new peptides production. Bioresour. Technol. 2019, 273, 103–113. [Google Scholar] [CrossRef] [Green Version]
- Schurink, M.; van Berkel, W.J.H.; Wichers, H.J.; Boeriu, C.G. Novel peptides with tyrosinase inhibitory activity. Peptides 2007, 28, 485–495. [Google Scholar] [CrossRef]
- Van Tran, V.; Moon, J.-Y.; Lee, Y.-C. Liposomes for delivery of antioxidants in cosmeceuticals: Challenges and development strategies. J. Control. Release 2019, 300, 114–140. [Google Scholar] [CrossRef] [PubMed]
- Lorenzo, J.M.; Munekata, P.E.S.; Gómez, B.; Barba, F.J.; Mora, L.; Pérez-Santaescolástica, C.; Toldrá, F. Bioactive peptides as natural antioxidants in food products—A review. Trends Food Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Mirzaei, M.; Shavandi, A.; Mirdamadi, S.; Soleymanzadeh, N.; Motahari, P.; Mirdamadi, N.; Moser, M.; Subra, G.; Alimoradi, H.; Goriely, S. Bioactive peptides from yeast: A comparative review on production methods, bioactivity, structure-function relationship, and stability. Trends Food Sci. Technol. 2021, 118, 297–315. [Google Scholar] [CrossRef]
- Ribeiro-Oliveira, R.; Martins, Z.E.; Sousa, J.B.; Ferreira, I.M.P.L.V.O.; Diniz, C. The health-promoting potential of peptides from brewing by-products: An up-to-date review. Trends Food Sci. Technol. 2021, 118, 143–153. [Google Scholar] [CrossRef]
- Ciurko, D.; Łaba, W.; Żarowska, B.; Janek, T. Enzymatic hydrolysis using bacterial cultures as a novel method for obtaining antioxidant peptides from brewers’ spent grain. RSC Adv. 2021, 11, 4688–4700. [Google Scholar] [CrossRef]
- Schaich, K.M.; Tian, X.; Xie, J. Hurdles and pitfalls in measuring antioxidant efficacy: A critical evaluation of ABTS, DPPH, and ORAC assays. J. Funct. Foods 2015, 14, 111–125. [Google Scholar] [CrossRef]
- Marson, G.V.; de Castro, R.J.S.; Machado, M.T.d.C.; da Silva Zandonadi, F.; Barros, H.D.d.F.Q.; Maróstica Júnior, M.R.; Sussulini, A.; Hubinger, M.D. Proteolytic enzymes positively modulated the physicochemical and antioxidant properties of spent yeast protein hydrolysates. Process Biochem. 2020, 91, 34–45. [Google Scholar] [CrossRef]
- Lima, T.N.; Pedriali Moraes, C.A. Bioactive Peptides: Applications and Relevance for Cosmeceuticals. Cosmetics 2018, 5, 21. [Google Scholar] [CrossRef] [Green Version]
- Restu, W.K.; Nishida, Y.; Yamamoto, S.; Ishii, J.; Maruyama, T. Short Oligopeptides for Biocompatible and Biodegradable Supramolecular Hydrogels. Langmuir 2018, 34, 8065–8074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirzaei, M.; Mirdamadi, S.; Safavi, M. Antioxidant activity and protective effects of Saccharomyces cerevisiae peptide fractions against H2O2-induced oxidative stress in Caco-2 cells. J. Food Meas. Charact. 2019, 13, 2654–2662. [Google Scholar] [CrossRef]
- Moharamzadeh, K.; Van Noort, R.; Brook, I.M.; Scutt, A.M. Cytotoxicity of resin monomers on human gingival fibroblasts and HaCaT keratinocytes. Dent. Mater. 2007, 23, 40–44. [Google Scholar] [CrossRef]
- Damour, O.; Zhi Hua, S.; Lasne, F.; Villain, M.; Rousselle, P.; Collombel, C. Cytotoxicity evaluation of antiseptics and antibiotics on cultured human fibroblasts and keratinocytes. Burns 1992, 18, 479–485. [Google Scholar] [CrossRef]
- Wiegand, C.; Hipler, U.C. Evaluation of Biocompatibility and Cytotoxicity Using Keratinocyte and Fibroblast Cultures. Skin Pharmacol. Physiol. 2009, 22, 74–82. [Google Scholar] [CrossRef]
- Costa, E.M.; Pereira, C.F.; Ribeiro, A.A.; Casanova, F.; Freixo, R.; Pintado, M.; Ramos, O.L. Characterization and Evaluation of Commercial Carboxymethyl Cellulose Potential as an Active Ingredient for Cosmetics. Appl. Sci. 2022, 12, 6560. [Google Scholar] [CrossRef]
- Apone, F.; Barbulova, A.; Colucci, M.G. Plant and Microalgae Derived Peptides Are Advantageously Employed as Bioactive Compounds in Cosmetics. Front. Plant Sci. 2019, 10, 756. [Google Scholar] [CrossRef] [Green Version]
- Zaky, A.A.; Simal-Gandara, J.; Eun, J.-B.; Shim, J.-H.; Abd El-Aty, A.M. Bioactivities, Applications, Safety, and Health Benefits of Bioactive Peptides From Food and By-Products: A Review. Front. Nutr. 2022, 8, 815640. [Google Scholar] [CrossRef]
- Tsai, W.C.; Hsu, C.C.; Chung, C.Y.; Lin, M.S.; Li, S.L.; Pang, J.H.S. The pentapeptide KTTKS promoting the expressions of type I collagen and transforming growth factor-β of tendon cells. J. Orthop. Res. 2007, 25, 1629–1634. [Google Scholar] [CrossRef]
- Edgar, S.; Hopley, B.; Genovese, L.; Sibilla, S.; Laight, D.; Shute, J. Effects of collagen-derived bioactive peptides and natural antioxidant compounds on proliferation and matrix protein synthesis by cultured normal human dermal fibroblasts. Sci. Rep. 2018, 8, 10474. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Reddi, S.; Devi, S.; Mada, S.B.; Kapila, R.; Kapila, S. Nrf2 dependent antiaging effect of milk-derived bioactive peptide in old fibroblasts. J. Cell. Biochem. 2019, 120, 9677–9691. [Google Scholar] [CrossRef] [PubMed]
- Offengenden, M.; Chakrabarti, S.; Wu, J. Chicken collagen hydrolysates differentially mediate anti-inflammatory activity and type I collagen synthesis on human dermal fibroblasts. Food Sci. Hum. Wellness 2018, 7, 138–147. [Google Scholar] [CrossRef]
- Benjakul, S.; Karnjanapratum, S.; Visessanguan, W. Hydrolysed collagen from Lates calcarifer skin: Its acute toxicity and impact on cell proliferation and collagen production of fibroblasts. Int. J. Food Sci. Technol. 2018, 53, 1871–1879. [Google Scholar] [CrossRef]
- Nakchum, L.; Kim, S.M. Preparation of squid skin collagen hydrolysate as an antihyaluronidase, antityrosinase, and antioxidant agent. Prep. Biochem. Biotechnol. 2016, 46, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Norzagaray-Valenzuela, C.D.; Valdez-Ortiz, A.; Shelton, L.M.; Jiménez-Edeza, M.; Rivera-López, J.; Valdez-Flores, M.A.; Germán-Báez, L.J. Residual biomasses and protein hydrolysates of three green microalgae species exhibit antioxidant and anti-aging activity. J. Appl. Phycol. 2017, 29, 189–198. [Google Scholar] [CrossRef]
- Farwick, M.; Grether-Beck, S.; Marini, A.; Maczkiewitz, U.; Lange, J.; Köhler, T.; Lersch, P.; Falla, T.; Felsner, I.; Brenden, H. Bioactive tetrapeptide GEKG boosts extracellular matrix formation: In vitro and in vivo molecular and clinical proof. Exp. Dermatol. 2011, 20, 602–604. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Luo, D.; Qu, W.; Chen, D.; Hong, Y.; Sheng, J.; Yang, X.; Liu, W. Nanoliposomes codelivering bioactive peptides produce enhanced anti-aging effect in human skin. J. Drug Deliv. Sci. Technol. 2020, 57, 101693. [Google Scholar] [CrossRef]
- De Lucia, A.; Zappelli, C.; Angelillo, M.; Langelotti, A.; Fogliano, V.; Cucchiara, M. A novel biotechnological active ingredient, derived from the microalga Spirulina, increases hydration and reduces osmotic stress in skin cells. H&PC Today 2018, 13, 60. [Google Scholar]
- Nicolas-Espinosa, J.; Yepes-Molina, L.; Carvajal, M. Bioactive peptides from broccoli stems strongly enhance regenerative keratinocytes by stimulating controlled proliferation. Pharm. Biol. 2022, 60, 235–246. [Google Scholar] [CrossRef] [PubMed]
- Ikarashi, N.; Kaneko, M.; Wakana, D.; Shinozaki, Y.; Tabata, K.; Nishinaka, Y.; Yoshida, R.; Watanabe, T.; Wakui, N.; Kon, R.; et al. Effect of Chimpi, dried citrus peel, on aquaporin-3 expression in HaCaT human epidermal keratinocytes. Mol. Biol. Rep. 2022, 49, 10175–10181. [Google Scholar] [CrossRef]
- Fournière, M.; Bedoux, G.; Lebonvallet, N.; Leschiera, R.; Le Goff-Pain, C.; Bourgougnon, N.; Latire, T. Poly- and Oligosaccharide Ulva sp. Fractions from Enzyme-Assisted Extraction Modulate the Metabolism of Extracellular Matrix in Human Skin Fibroblasts: Potential in Anti-Aging Dermo-Cosmetic Applications. Mar. Drugs 2021, 19, 156. [Google Scholar] [CrossRef] [PubMed]
- Coscueta, E.R.; Campos, D.A.; Osório, H.; Nerli, B.B.; Pintado, M. Enzymatic soy protein hydrolysis: A tool for biofunctional food ingredient production. Food Chem. X 2019, 1, 100006. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa, E.M.; Oliveira, A.S.; Silva, S.; Ribeiro, A.B.; Pereira, C.F.; Ferreira, C.; Casanova, F.; Pereira, J.O.; Freixo, R.; Pintado, M.E.; et al. Spent Yeast Waste Streams as a Sustainable Source of Bioactive Peptides for Skin Applications. Int. J. Mol. Sci. 2023, 24, 2253. https://doi.org/10.3390/ijms24032253
Costa EM, Oliveira AS, Silva S, Ribeiro AB, Pereira CF, Ferreira C, Casanova F, Pereira JO, Freixo R, Pintado ME, et al. Spent Yeast Waste Streams as a Sustainable Source of Bioactive Peptides for Skin Applications. International Journal of Molecular Sciences. 2023; 24(3):2253. https://doi.org/10.3390/ijms24032253
Chicago/Turabian StyleCosta, Eduardo M., Ana Sofia Oliveira, Sara Silva, Alessandra B. Ribeiro, Carla F. Pereira, Carlos Ferreira, Francisca Casanova, Joana O. Pereira, Ricardo Freixo, Manuela E. Pintado, and et al. 2023. "Spent Yeast Waste Streams as a Sustainable Source of Bioactive Peptides for Skin Applications" International Journal of Molecular Sciences 24, no. 3: 2253. https://doi.org/10.3390/ijms24032253