
Opposite Modulation of the NMDA Receptor by Glycine and S-Ketamine and the Effects on Resting State EEG Gamma Activity: New Insights into the Glutamate Hypothesis of Schizophrenia
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Participants
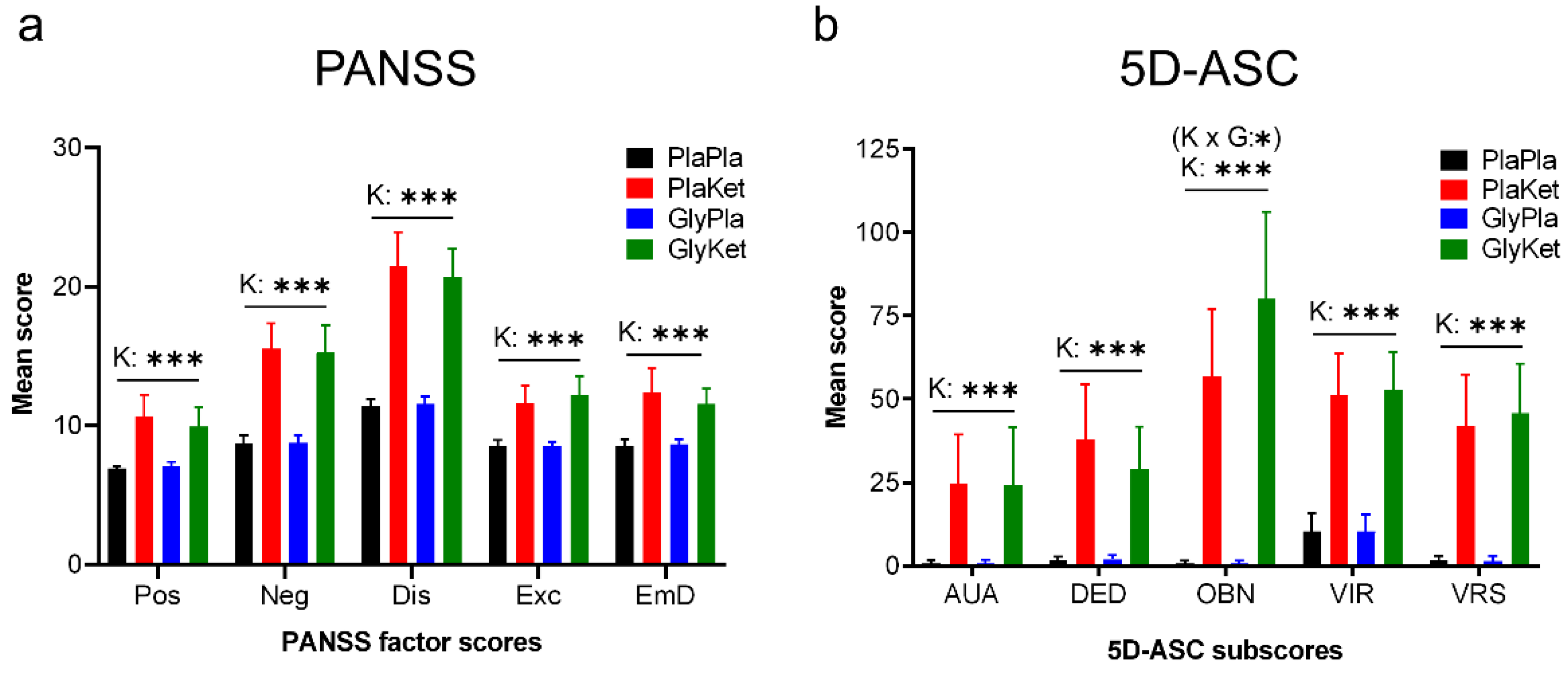
2.2. Psychometric Assessment
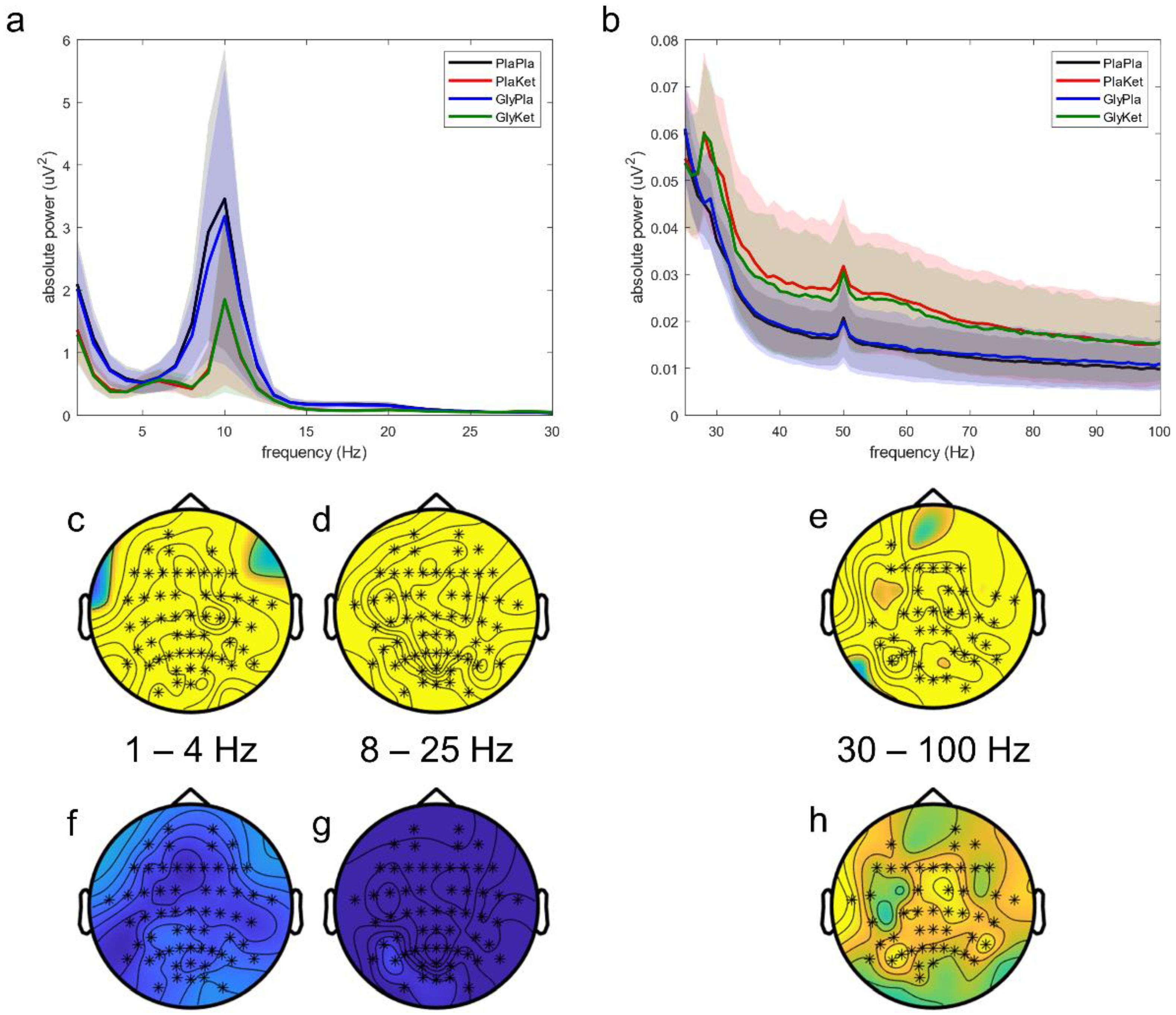
2.3. EEG Scalp Analysis
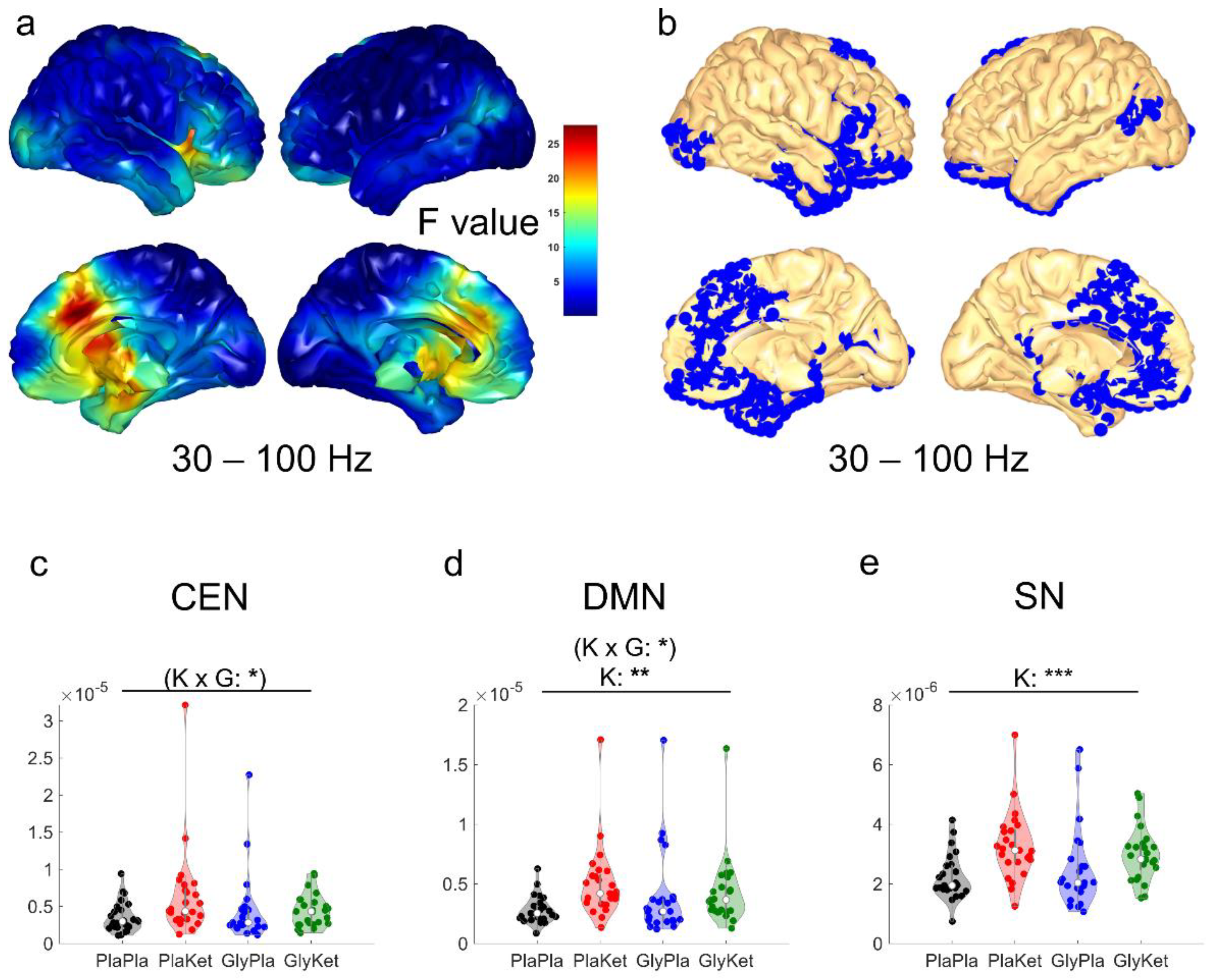
2.4. EEG Source Analysis
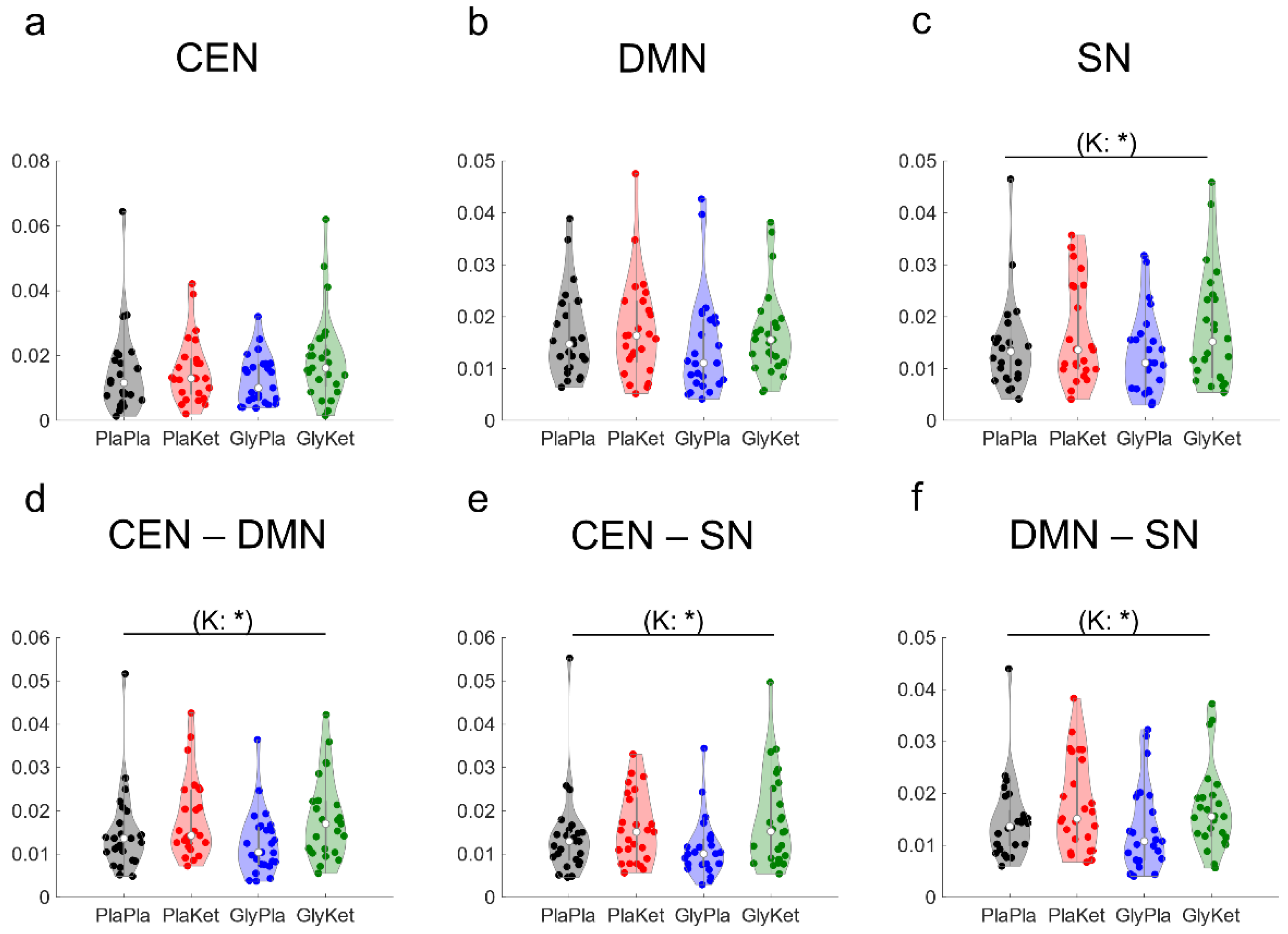
2.5. EEG Functional Connectivity Analysis
2.6. Association between the Psychometric Results and Neurophysiological Measures
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Study Design
4.3. Psychometric Assessment
4.4. EEG Aquisition
4.5. EEG Preprocessing
4.6. EEG Scalp Analysis
4.7. EEG Source Analysis
4.8. EEG Functional Conncectivity Analysis
4.9. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Miyamoto, S.; Miyake, N.; Jarskog, L.F.; Fleischhacker, W.W.; Lieberman, J.A. Pharmacological treatment of schizophrenia: A critical review of the pharmacology and clinical effects of current and future therapeutic agents. Mol. Psychiatry 2012, 17, 1206–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howes, O.; McCutcheon, R.; Stone, J. Glutamate and dopamine in schizophrenia: An update for the 21st century. J. Psychopharmacol. 2015, 29, 97–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potkin, S.G.; Kane, J.M.; Correll, C.U.; Lindenmayer, J.P.; Agid, O.; Marder, S.R.; Olfson, M.; Howes, O.D. The neurobiology of treatment-resistant schizophrenia: Paths to antipsychotic resistance and a roadmap for future research. NPJ Schizophr. 2020, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millan, M.J.; Fone, K.; Steckler, T.; Horan, W.P. Negative symptoms of schizophrenia: Clinical characteristics, pathophysiological substrates, experimental models and prospects for improved treatment. Eur. Neuropsychopharmacol. 2014, 24, 645–692. [Google Scholar] [CrossRef] [Green Version]
- Galderisi, S.; Kaiser, S.; Bitter, I.; Nordentoft, M.; Mucci, A.; Sabe, M.; Giordano, G.M.; Nielsen, M.O.; Glenthoj, L.B.; Pezzella, P.; et al. EPA guidance on treatment of negative symptoms in schizophrenia. Eur. Psychiatry 2021, 64, e21. [Google Scholar] [CrossRef]
- Correll, C.U.; Schooler, N.R. Negative Symptoms in Schizophrenia: A Review and Clinical Guide for Recognition, Assessment, and Treatment. Neuropsychiatr. Dis. Treat. 2020, 16, 519–534. [Google Scholar] [CrossRef] [Green Version]
- Poels, E.M.; Kegeles, L.S.; Kantrowitz, J.T.; Slifstein, M.; Javitt, D.C.; Lieberman, J.A.; Abi-Dargham, A.; Girgis, R.R. Imaging glutamate in schizophrenia: Review of findings and implications for drug discovery. Mol. Psychiatry 2014, 19, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Uno, Y.; Coyle, J.T. Glutamate hypothesis in schizophrenia. Psychiatry Clin. Neurosci. 2019, 73, 204–215. [Google Scholar] [CrossRef] [Green Version]
- McCutcheon, R.A.; Krystal, J.H.; Howes, O.D. Dopamine and glutamate in schizophrenia: Biology, symptoms and treatment. World Psychiatry 2020, 19, 15–33. [Google Scholar] [CrossRef] [Green Version]
- Kehrer, C.; Maziashvili, N.; Dugladze, T.; Gloveli, T. Altered Excitatory-Inhibitory Balance in the NMDA-Hypofunction Model of Schizophrenia. Front. Mol. Neurosci. 2008, 1, 6. [Google Scholar] [CrossRef]
- Trubetskoy, V.; Pardinas, A.F.; Qi, T.; Panagiotaropoulou, G.; Awasthi, S.; Bigdeli, T.B.; Bryois, J.; Chen, C.Y.; Dennison, C.A.; Hall, L.S.; et al. Mapping genomic loci implicates genes and synaptic biology in schizophrenia. Nature 2022, 604, 502–508. [Google Scholar] [CrossRef] [PubMed]
- Konradi, C.; Yang, C.K.; Zimmerman, E.I.; Lohmann, K.M.; Gresch, P.; Pantazopoulos, H.; Berretta, S.; Heckers, S. Hippocampal interneurons are abnormal in schizophrenia. Schizophr. Res. 2011, 131, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Leeson, P.D.; Iversen, L.L. The glycine site on the NMDA receptor: Structure-activity relationships and therapeutic potential. J. Med. Chem. 1994, 37, 4053–4067. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Singh, V. Meta-analysis of the efficacy of adjunctive NMDA receptor modulators in chronic schizophrenia. CNS Drugs 2011, 25, 859–885. [Google Scholar] [CrossRef] [PubMed]
- Kuo, C.Y.; Lin, C.H.; Lane, H.Y. Targeting D-Amino Acid Oxidase (DAAO) for the Treatment of Schizophrenia: Rationale and Current Status of Research. CNS Drugs 2022, 36, 1143–1153. [Google Scholar] [CrossRef] [PubMed]
- Bugarski-Kirola, D.; Blaettler, T.; Arango, C.; Fleischhacker, W.W.; Garibaldi, G.; Wang, A.; Dixon, M.; Bressan, R.A.; Nasrallah, H.; Lawrie, S.; et al. Bitopertin in Negative Symptoms of Schizophrenia-Results from the Phase III FlashLyte and DayLyte Studies. Biol. Psychiatry 2017, 82, 8–16. [Google Scholar] [CrossRef]
- Sonnenschein, S.F.; Grace, A. Emerging therapeutic targets for schizophrenia: A framework for novel treatment strategies for psychosis. Expert Opin. Ther. Targets 2021, 25, 15–26. [Google Scholar] [CrossRef]
- Frohlich, J.; Van Horn, J.D. Reviewing the ketamine model for schizophrenia. J. Psychopharmacol. 2014, 28, 287–302. [Google Scholar] [CrossRef]
- Javitt, D.C.; Siegel, S.J.; Spencer, K.M.; Mathalon, D.H.; Hong, L.E.; Martinez, A.; Ehlers, C.L.; Abbas, A.I.; Teichert, T.; Lakatos, P.; et al. A roadmap for development of neuro-oscillations as translational biomarkers for treatment development in neuropsychopharmacology. Neuropsychopharmacology 2020, 45, 1411–1422. [Google Scholar] [CrossRef]
- Javitt, D.C.; Carter, C.S.; Krystal, J.H.; Kantrowitz, J.T.; Girgis, R.R.; Kegeles, L.S.; Ragland, J.D.; Maddock, R.J.; Lesh, T.A.; Tanase, C.; et al. Utility of Imaging-Based Biomarkers for Glutamate-Targeted Drug Development in Psychotic Disorders: A Randomized Clinical Trial. JAMA Psychiatry 2018, 75, 11–19. [Google Scholar] [CrossRef]
- Haaf, M.; Leicht, G.; Curic, S.; Mulert, C. Glutamatergic Deficits in Schizophrenia-Biomarkers and Pharmacological Interventions within the Ketamine Model. Curr. Pharm. Biotechnol. 2018, 19, 293–307. [Google Scholar] [CrossRef] [PubMed]
- Krystal, J.H.; Karper, L.P.; Seibyl, J.P.; Freeman, G.K.; Delaney, R.; Bremner, J.D.; Heninger, G.R.; Bowers, M.B., Jr.; Charney, D.S. Subanesthetic effects of the noncompetitive NMDA antagonist, ketamine, in humans. Psychotomimetic, perceptual, cognitive, and neuroendocrine responses. Arch. Gen. Psychiatry 1994, 51, 199–214. [Google Scholar] [CrossRef] [PubMed]
- Gandal, M.J.; Edgar, J.C.; Klook, K.; Siegel, S.J. Gamma synchrony: Towards a translational biomarker for the treatment-resistant symptoms of schizophrenia. Neuropharmacology 2012, 62, 1504–1518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curic, S.; Leicht, G.; Thiebes, S.; Andreou, C.; Polomac, N.; Eichler, I.C.; Eichler, L.; Zolliner, C.; Gallinat, J.; Steinmann, S.; et al. Reduced auditory evoked gamma-band response and schizophrenia-like clinical symptoms under subanesthetic ketamine. Neuropsychopharmacology 2019, 44, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Uhlhaas, P.J.; Singer, W. Abnormal neural oscillations and synchrony in schizophrenia. Nat. Rev. Neurosci. 2010, 11, 100–113. [Google Scholar] [CrossRef]
- Singer, W. Neuronal synchrony: A versatile code for the definition of relations? Neuron 1999, 24, 49–65. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.-J. Neurophysiological and computational principles of cortical rhythms in cognition. Physiol. Rev. 2010, 90, 1195–1268. [Google Scholar] [CrossRef]
- Sohal, V.S.; Zhang, F.; Yizhar, O.; Deisseroth, K. Parvalbumin neurons and gamma rhythms enhance cortical circuit performance. Nature 2009, 459, 698–702. [Google Scholar] [CrossRef] [Green Version]
- Spencer, K.M. Time to be spontaneous: A renaissance of intrinsic brain activity in psychosis research? Biol. Psychiatry 2014, 76, 434–435. [Google Scholar] [CrossRef]
- Spencer, K.M. Baseline gamma power during auditory steady-state stimulation in schizophrenia. Front. Hum. Neurosci. 2011, 5, 190. [Google Scholar] [CrossRef]
- Baldeweg, T.; Spence, S.; Hirsch, S.R.; Gruzelier, J. Gamma-band electroencephalographic oscillations in a patient with somatic hallucinations. Lancet 1998, 352, 620–621. [Google Scholar] [CrossRef]
- Baradits, M.; Kakuszi, B.; Balint, S.; Fullajtar, M.; Mod, L.; Bitter, I.; Czobor, P. Alterations in resting-state gamma activity in patients with schizophrenia: A high-density EEG study. Eur. Arch. Psychiatry Clin. Neurosci. 2019, 269, 429–437. [Google Scholar] [CrossRef]
- Bianciardi, B.; Uhlhaas, P.J. Do NMDA-R antagonists re-create patterns of spontaneous gamma-band activity in schizophrenia? A systematic review and perspective. Neurosci. Biobehav. Rev. 2021, 124, 308–323. [Google Scholar] [CrossRef]
- Muthukumaraswamy, S.D.; Shaw, A.D.; Jackson, L.E.; Hall, J.; Moran, R.; Saxena, N. Evidence that Subanesthetic Doses of Ketamine Cause Sustained Disruptions of NMDA and AMPA-Mediated Frontoparietal Connectivity in Humans. J. Neurosci. 2015, 35, 11694–11706. [Google Scholar] [CrossRef] [Green Version]
- Rivolta, D.; Heidegger, T.; Scheller, B.; Sauer, A.; Schaum, M.; Birkner, K.; Singer, W.; Wibral, M.; Uhlhaas, P.J. Ketamine dysregulates the amplitude and connectivity of high-frequency oscillations in cortical-subcortical networks in humans: Evidence from resting-state magnetoencephalography-recordings. Schizophr. Bull. 2015, 41, 1105–1114. [Google Scholar] [CrossRef] [Green Version]
- de la Salle, S.; Choueiry, J.; Shah, D.; Bowers, H.; McIntosh, J.; Ilivitsky, V.; Knott, V. Effects of ketamine on resting-state EEG activity and their relationship to perceptual/dissociative symptoms in healthy humans. Front. Pharmacol. 2016, 7, 348. [Google Scholar] [CrossRef] [Green Version]
- Hudson, M.R.; Rind, G.; O’Brien, T.J.; Jones, N.C. Reversal of evoked gamma oscillation deficits is predictive of antipsychotic activity with a unique profile for clozapine. Transl. Psychiatry 2016, 6, e784. [Google Scholar] [CrossRef]
- Kocsis, B. Differential role of NR2A and NR2B subunits in N-methyl-D-aspartate receptor antagonist-induced aberrant cortical gamma oscillations. Biol. Psychiatry 2012, 71, 987–995. [Google Scholar] [CrossRef] [Green Version]
- Bressler, S.L. Large-scale cortical networks and cognition. Brain Res. Brain Res. Rev. 1995, 20, 288–304. [Google Scholar] [CrossRef]
- Nekovarova, T.; Fajnerova, I.; Horacek, J.; Spaniel, F. Bridging disparate symptoms of schizophrenia: A triple network dysfunction theory. Front. Behav. Neurosci. 2014, 8, 171. [Google Scholar] [CrossRef]
- Bressler, S.L.; Menon, V. Large-scale brain networks in cognition: Emerging methods and principles. Trends Cogn. Sci. 2010, 14, 277–290. [Google Scholar] [CrossRef] [PubMed]
- Menon, V. Large-scale brain networks and psychopathology: A unifying triple network model. Trends Cogn. Sci. 2011, 15, 483–506. [Google Scholar] [CrossRef] [PubMed]
- Haaf, M.; Curic, S.; Steinmann, S.; Rauh, J.; Leicht, G.; Mulert, C. Glycine attenuates impairments of stimulus-evoked gamma oscillations in the ketamine model of schizophrenia. Neuroimage 2022, 251, 119004. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Balla, A.; Sershen, H.; Sehatpour, P.; Lakatos, P.; Javitt, D.C. Rodent mismatch negativity/theta neuro-oscillatory response as a translational neurophysiological biomarker for N-Methyl-D-Aspartate receptor-based new treatment development in schizophrenia. Neuropsychopharmacology 2018, 43, 571–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curic, S.; Andreou, C.; Nolte, G.; Steinmann, S.; Thiebes, S.; Polomac, N.; Haaf, M.; Rauh, J.; Leicht, G.; Mulert, C. Ketamine alters functional gamma and theta resting-state connectivity in healthy humans: Implications for schizophrenia treatment targeting the glutamate system. Front. Psychiatry 2021, 12, 671007. [Google Scholar] [CrossRef]
- Zacharias, N.; Musso, F.; Muller, F.; Lammers, F.; Saleh, A.; London, M.; de Boer, P.; Winterer, G. Ketamine effects on default mode network activity and vigilance: A randomized, placebo-controlled crossover simultaneous fMRI/EEG study. Hum. Brain Mapp. 2020, 41, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Nugent, A.C.; Ballard, E.D.; Gould, T.D.; Park, L.T.; Moaddel, R.; Brutsche, N.E.; Zarate, C.A. Ketamine has distinct electrophysiological and behavioral effects in depressed and healthy subjects. Mol. Psychiatry 2019, 24, 1040–1052. [Google Scholar] [CrossRef]
- de la Salle, S.; Choueiry, J.; Shah, D.; Bowers, H.; McIntosh, J.; Ilivitsky, V.; Carroll, B.; Knott, V. Resting-state functional EEG connectivity in salience and default mode networks and their relationship to dissociative symptoms during NMDA receptor antagonism. Pharmacol. Biochem. Behav. 2021, 201, 173092. [Google Scholar] [CrossRef]
- Arikan, M.K.; Metin, B.; Metin, S.Z.; Tulay, E.E.; Tarhan, N. High Frequencies in QEEG Are Related to the Level of Insight in Patients with Schizophrenia. Clin. EEG Neurosci. 2018, 49, 316–320. [Google Scholar] [CrossRef]
- Grent-′t-Jong, T.; Gross, J.; Goense, J.; Wibral, M.; Gajwani, R.; Gumley, A.I.; Lawrie, S.M.; Schwannauer, M.; Schultze-Lutter, F.; Schroder, T.N.; et al. Resting-state gamma-band power alterations in schizophrenia reveal E/I-balance abnormalities across illness-stages. Elife 2018, 7, e37799. [Google Scholar] [CrossRef]
- Hirano, Y.; Oribe, N.; Kanba, S.; Onitsuka, T.; Nestor, P.G.; Spencer, K.M. Spontaneous gamma activity in schizophrenia. JAMA Psychiatry 2015, 72, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Kissler, J.; Muller, M.M.; Fehr, T.; Rockstroh, B.; Elbert, T. MEG gamma band activity in schizophrenia patients and healthy subjects in a mental arithmetic task and at rest. Clin. Neurophysiol. 2000, 111, 2079–2087. [Google Scholar] [CrossRef] [Green Version]
- Garakh, Z.; Zaytseva, Y.; Kapranova, A.; Fiala, O.; Horacek, J.; Shmukler, A.; Gurovich, I.Y.; Strelets, V.B. EEG correlates of a mental arithmetic task in patients with first episode schizophrenia and schizoaffective disorder. Clin. Neurophysiol. 2015, 126, 2090–2098. [Google Scholar] [CrossRef]
- Hu, W.; MacDonald, M.L.; Elswick, D.E.; Sweet, R.A. The glutamate hypothesis of schizophrenia: Evidence from human brain tissue studies. Ann. N. Y. Acad. Sci. 2015, 1338, 38–57. [Google Scholar] [CrossRef] [Green Version]
- Guyon, N.; Zacharias, L.R.; Fermino de Oliveira, E.; Kim, H.; Leite, J.P.; Lopes-Aguiar, C.; Carlen, M. Network asynchrony underlying increased broadband gamma power. J. Neurosci. 2021, 41, 2944–2963. [Google Scholar] [CrossRef]
- Northoff, G. Is schizophrenia a spatiotemporal disorder of the brain’s resting state? World Psychiatry 2015, 14, 34–35. [Google Scholar] [CrossRef] [Green Version]
- Lavigne, K.M.; Menon, M.; Woodward, T.S. Functional brain networks underlying evidence integration and delusions in schizophrenia. Schizophr. Bull. 2020, 46, 175–183. [Google Scholar] [CrossRef]
- Liu, H.; Kaneko, Y.; Ouyang, X.; Li, L.; Hao, Y.; Chen, E.Y.; Jiang, T.; Zhou, Y.; Liu, Z. Schizophrenic patients and their unaffected siblings share increased resting-state connectivity in the task-negative network but not its anticorrelated task-positive network. Schizophr. Bull. 2012, 38, 285–294. [Google Scholar] [CrossRef] [Green Version]
- Supekar, K.; Cai, W.; Krishnadas, R.; Palaniyappan, L.; Menon, V. Dysregulated brain dynamics in a triple-network saliency model of schizophrenia and its relation to psychosis. Biol. Psychiatry 2019, 85, 60–69. [Google Scholar] [CrossRef]
- Manoliu, A.; Riedl, V.; Zherdin, A.; Muhlau, M.; Schwerthoffer, D.; Scherr, M.; Peters, H.; Zimmer, C.; Forstl, H.; Bauml, J.; et al. Aberrant dependence of default mode/central executive network interactions on anterior insular salience network activity in schizophrenia. Schizophr. Bull. 2014, 40, 428–437. [Google Scholar] [CrossRef]
- Andreou, C.; Nolte, G.; Leicht, G.; Polomac, N.; Hanganu-Opatz, I.L.; Lambert, M.; Engel, A.K.; Mulert, C. Increased resting-state gamma-band connectivity in first-episode schizophrenia. Schizophr. Bull. 2015, 41, 930–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, G.; Daverio, A.; Ferrentino, F.; Santarnecchi, E.; Ciabattini, F.; Monaco, L.; Lisi, G.; Barone, Y.; Di Lorenzo, C.; Niolu, C.; et al. Altered resting-state EEG source functional connectivity in schizophrenia: The effect of illness duration. Front. Hum. Neurosci. 2015, 9, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leicht, G.; Kirsch, V.; Giegling, I.; Karch, S.; Hantschk, I.; Moller, H.J.; Pogarell, O.; Hegerl, U.; Rujescu, D.; Mulert, C. Reduced early auditory evoked gamma-band response in patients with schizophrenia. Biol. Psychiatry 2010, 67, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Leicht, G.; Vauth, S.; Polomac, N.; Andreou, C.; Rauh, J.; Mussmann, M.; Karow, A.; Mulert, C. EEG-informed fMRI reveals a disturbed gamma-band-specific network in subjects at high risk for psychosis. Schizophr. Bull. 2016, 42, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Leicht, G.; Andreou, C.; Polomac, N.; Lanig, C.; Schottle, D.; Lambert, M.; Mulert, C. Reduced auditory evoked gamma band response and cognitive processing deficits in first episode schizophrenia. World J. Biol. Psychiatry 2015, 16, 387–397. [Google Scholar] [CrossRef]
- Spencer, K.M.; Niznikiewicz, M.A.; Shenton, M.E.; McCarley, R.W. Sensory-evoked gamma oscillations in chronic schizophrenia. Biol. Psychiatry 2008, 63, 744–747. [Google Scholar] [CrossRef] [Green Version]
- Shaw, A.D.; Knight, L.; Freeman, T.C.A.; Williams, G.M.; Moran, R.J.; Friston, K.J.; Walters, J.T.R.; Singh, K.D. Oscillatory, Computational, and Behavioral Evidence for Impaired GABAergic Inhibition in Schizophrenia. Schizophr. Bull. 2020, 46, 345–353. [Google Scholar] [CrossRef]
- Kantrowitz, J.T.; Grinband, J.; Goff, D.C.; Lahti, A.C.; Marder, S.R.; Kegeles, L.S.; Girgis, R.R.; Sobeih, T.; Wall, M.M.; Choo, T.H.; et al. Proof of mechanism and target engagement of glutamatergic drugs for the treatment of schizophrenia: RCTs of pomaglumetad and TS-134 on ketamine-induced psychotic symptoms and pharmacoBOLD in healthy volunteers. Neuropsychopharmacology 2020, 45, 1842–1850. [Google Scholar] [CrossRef]
- Mehta, M.A.; Schmechtig, A.; Kotoula, V.; McColm, J.; Jackson, K.; Brittain, C.; Tauscher-Wisniewski, S.; Kinon, B.J.; Morrison, P.D.; Pollak, T.; et al. Group II metabotropic glutamate receptor agonist prodrugs LY2979165 and LY2140023 attenuate the functional imaging response to ketamine in healthy subjects. Psychopharmacology 2018, 235, 1875–1886. [Google Scholar] [CrossRef]
- Greenwood, L.M.; Leung, S.; Michie, P.T.; Green, A.; Nathan, P.J.; Fitzgerald, P.; Johnston, P.; Solowij, N.; Kulkarni, J.; Croft, R.J. The effects of glycine on auditory mismatch negativity in schizophrenia. Schizophr. Res. 2018, 191, 61–69. [Google Scholar] [CrossRef]
- Kantrowitz, J.T.; Epstein, M.L.; Lee, M.; Lehrfeld, N.; Nolan, K.A.; Shope, C.; Petkova, E.; Silipo, G.; Javitt, D.C. Improvement in mismatch negativity generation during d-serine treatment in schizophrenia: Correlation with symptoms. Schizophr. Res. 2018, 191, 70–79. [Google Scholar] [CrossRef]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Baker, R.; Dunbar, G.C. The Mini-International Neuropsychiatric Interview (M.I.N.I.): The development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J. Clin. Psychiatry 1998, 59 (Suppl. 20), 22–33; quiz 34–57. [Google Scholar]
- Raine, A. The SPQ: A scale for the assessment of schizotypal personality based on DSM-III-R criteria. Schizophr. Bull. 1991, 17, 555–564. [Google Scholar] [CrossRef] [Green Version]
- D’Souza, D.C.; Gil, R.; Cassello, K.; Morrissey, K.; Abi-Saab, D.; White, J.; Sturwold, R.; Bennett, A.; Karper, L.P.; Zuzarte, E.; et al. IV glycine and oral D-cycloserine effects on plasma and CSF amino acids in healthy humans. Biol. Psychiatry 2000, 47, 450–462. [Google Scholar] [CrossRef]
- Schmidt, A.; Bachmann, R.; Kometer, M.; Csomor, P.A.; Stephan, K.E.; Seifritz, E.; Vollenweider, F.X. Mismatch negativity encoding of prediction errors predicts S-ketamine-induced cognitive impairments. Neuropsychopharmacology 2012, 37, 865–875. [Google Scholar] [CrossRef] [Green Version]
- Feng, N.; Vollenweider, F.X.; Minder, E.I.; Rentsch, K.; Grampp, T.; Vonderschmitt, D.J. Development of a gas chromatography-mass spectrometry method for determination of ketamine in plasma and its application to human samples. Ther. Drug Monit. 1995, 17, 95–100. [Google Scholar] [CrossRef]
- Umbricht, D.; Schmid, L.; Koller, R.; Vollenweider, F.X.; Hell, D.; Javitt, D.C. Ketamine-induced deficits in auditory and visual context-dependent processing in healthy volunteers: Implications for models of cognitive deficits in schizophrenia. Arch. Gen. Psychiatry 2000, 57, 1139–1147. [Google Scholar] [CrossRef] [Green Version]
- Kay, S.R.; Fiszbein, A.; Opler, L.A. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr. Bull. 1987, 13, 261–276. [Google Scholar] [CrossRef]
- Thiebes, S.; Leicht, G.; Curic, S.; Steinmann, S.; Polomac, N.; Andreou, C.; Eichler, I.; Eichler, L.; Zollner, C.; Gallinat, J.; et al. Glutamatergic deficit and schizophrenia-like negative symptoms: New evidence from ketamine-induced mismatch negativity alterations in healthy male humans. J. Psychiatry Neurosci. 2017, 42, 160187. [Google Scholar] [CrossRef] [Green Version]
- Thiebes, S.; Steinmann, S.; Curic, S.; Polomac, N.; Andreou, C.; Eichler, I.C.; Eichler, L.; Zollner, C.; Gallinat, J.; Leicht, G.; et al. Alterations in interhemispheric gamma-band connectivity are related to the emergence of auditory verbal hallucinations in healthy subjects during NMDA-receptor blockade. Neuropsychopharmacology 2018, 43, 1608–1615. [Google Scholar] [CrossRef] [Green Version]
- van der Gaag, M.; Hoffman, T.; Remijsen, M.; Hijman, R.; de Haan, L.; van Meijel, B.; van Harten, P.N.; Valmaggia, L.; de Hert, M.; Cuijpers, A.; et al. The five-factor model of the Positive and Negative Syndrome Scale II: A ten-fold cross-validation of a revised model. Schizophr. Res. 2006, 85, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, A. The standardized psychometric assessment of altered states of consciousness (ASCs) in humans. Pharmacopsychiatry 1998, 31 (Suppl. 2), 80–84. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, A.; Lamparter, D.; Maurer, M. 5D-ABZ: Fragebogen zur Erfassung Aussergewöhnlicher Bewusstseinszustände. In Eine Kurze Einführung; PSIN PLUS: Zurich, Switzerland, 2006. [Google Scholar]
- Jobert, M.; Wilson, F.J.; Ruigt, G.S.; Brunovsky, M.; Prichep, L.S.; Drinkenburg, W.H.; IPEG Pharmaco-EEG Guideline Committee. Guidelines for the recording and evaluation of pharmaco-EEG data in man: The International Pharmaco-EEG Society (IPEG). Neuropsychobiology 2012, 66, 201–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullen, T.R.; Kothe, C.A.; Chi, Y.M.; Ojeda, A.; Kerth, T.; Makeig, S.; Jung, T.P.; Cauwenberghs, G. Real-time neuroimaging and cognitive monitoring using wearable dry EEG. IEEE Trans. Biomed. Eng. 2015, 62, 2553–2567. [Google Scholar] [CrossRef] [Green Version]
- Pion-Tonachini, L.; Kreutz-Delgado, K.; Makeig, S. ICLabel: An automated electroencephalographic independent component classifier, dataset, and website. Neuroimage 2019, 198, 181–197. [Google Scholar] [CrossRef] [Green Version]
- Perrin, F.; Pernier, J.; Bertrand, O.; Echallier, J.F. Spherical splines for scalp potential and current density mapping. Electroencephalogr. Clin. Neurophysiol. 1989, 72, 184–187. [Google Scholar] [CrossRef]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.M. FieldTrip: Open source software for advanced analysis of MEG, EEG, and invasive electrophysiological data. Comput. Intell. Neurosci. 2011, 2011, 156869. [Google Scholar] [CrossRef]
- Pascual-Marqui, R.D. Discrete, 3D distributed, linear imaging methods of electric neuronal activity. Part 1: Exact, zero error localization. arXiv 2007, arXiv:0710.3341. [Google Scholar]
- Pascual-Marqui, R.D.; Lehmann, D.; Koukkou, M.; Kochi, K.; Anderer, P.; Saletu, B.; Tanaka, H.; Hirata, K.; John, E.R.; Prichep, L.; et al. Assessing interactions in the brain with exact low-resolution electromagnetic tomography. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 3768–3784. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haaf, M.; Curic, S.; Rauh, J.; Steinmann, S.; Mulert, C.; Leicht, G. Opposite Modulation of the NMDA Receptor by Glycine and S-Ketamine and the Effects on Resting State EEG Gamma Activity: New Insights into the Glutamate Hypothesis of Schizophrenia. Int. J. Mol. Sci. 2023, 24, 1913. https://doi.org/10.3390/ijms24031913
Haaf M, Curic S, Rauh J, Steinmann S, Mulert C, Leicht G. Opposite Modulation of the NMDA Receptor by Glycine and S-Ketamine and the Effects on Resting State EEG Gamma Activity: New Insights into the Glutamate Hypothesis of Schizophrenia. International Journal of Molecular Sciences. 2023; 24(3):1913. https://doi.org/10.3390/ijms24031913
Chicago/Turabian StyleHaaf, Moritz, Stjepan Curic, Jonas Rauh, Saskia Steinmann, Christoph Mulert, and Gregor Leicht. 2023. "Opposite Modulation of the NMDA Receptor by Glycine and S-Ketamine and the Effects on Resting State EEG Gamma Activity: New Insights into the Glutamate Hypothesis of Schizophrenia" International Journal of Molecular Sciences 24, no. 3: 1913. https://doi.org/10.3390/ijms24031913