
Towards a Long-Read Sequencing Approach for the Molecular Diagnosis of RPGRORF15 Genetic Variants

 ,
,  , , , , , , , , add
Show full author list
, , , , , , , , add
Show full author list
Abstract
:1. Introduction
2. Results
2.1. Study Cohort Characteristics
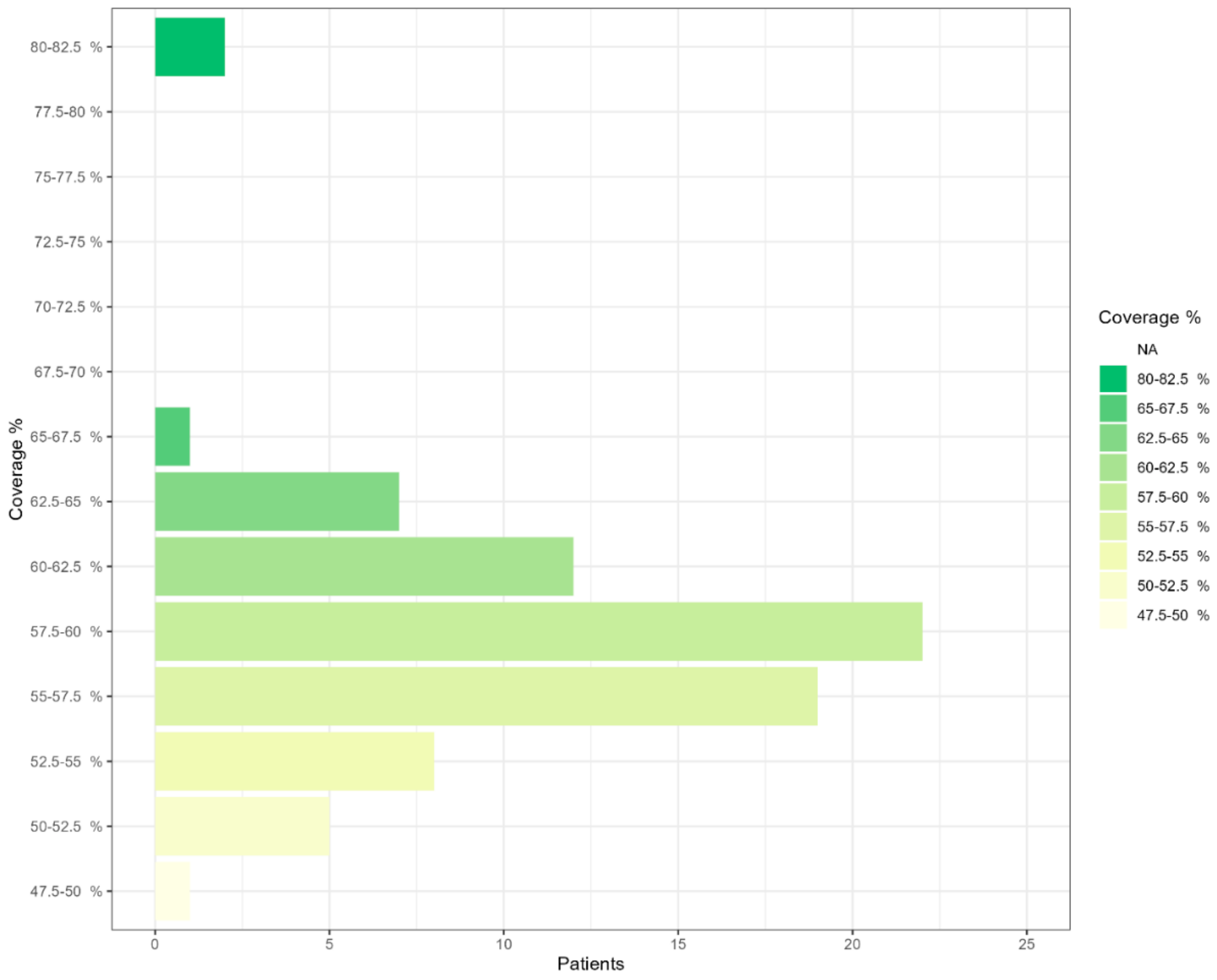
2.2. NGS Sequencing Coverage
2.3. PacBio Sequencing Results
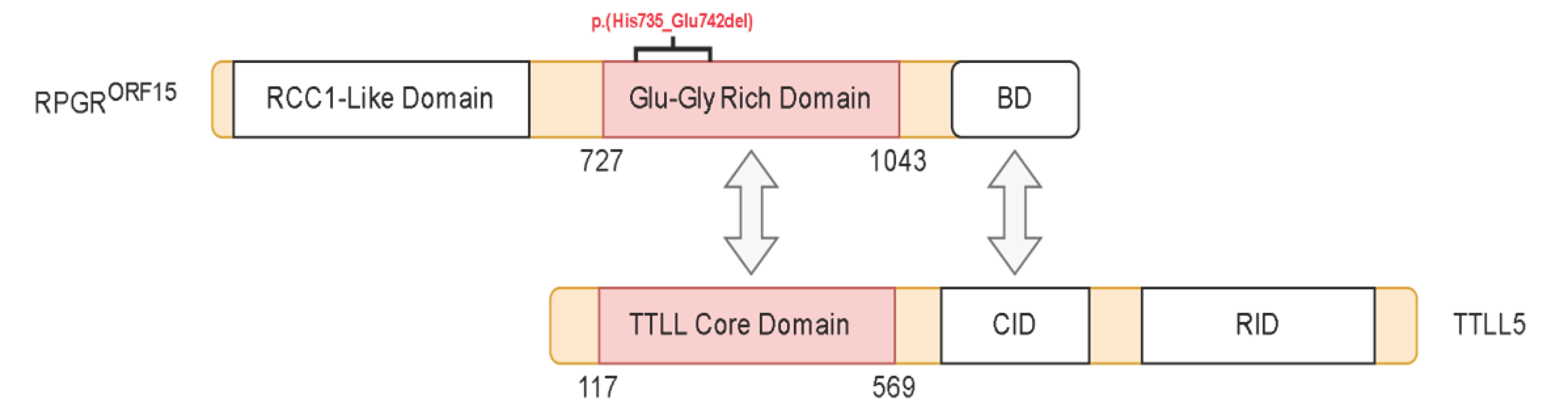
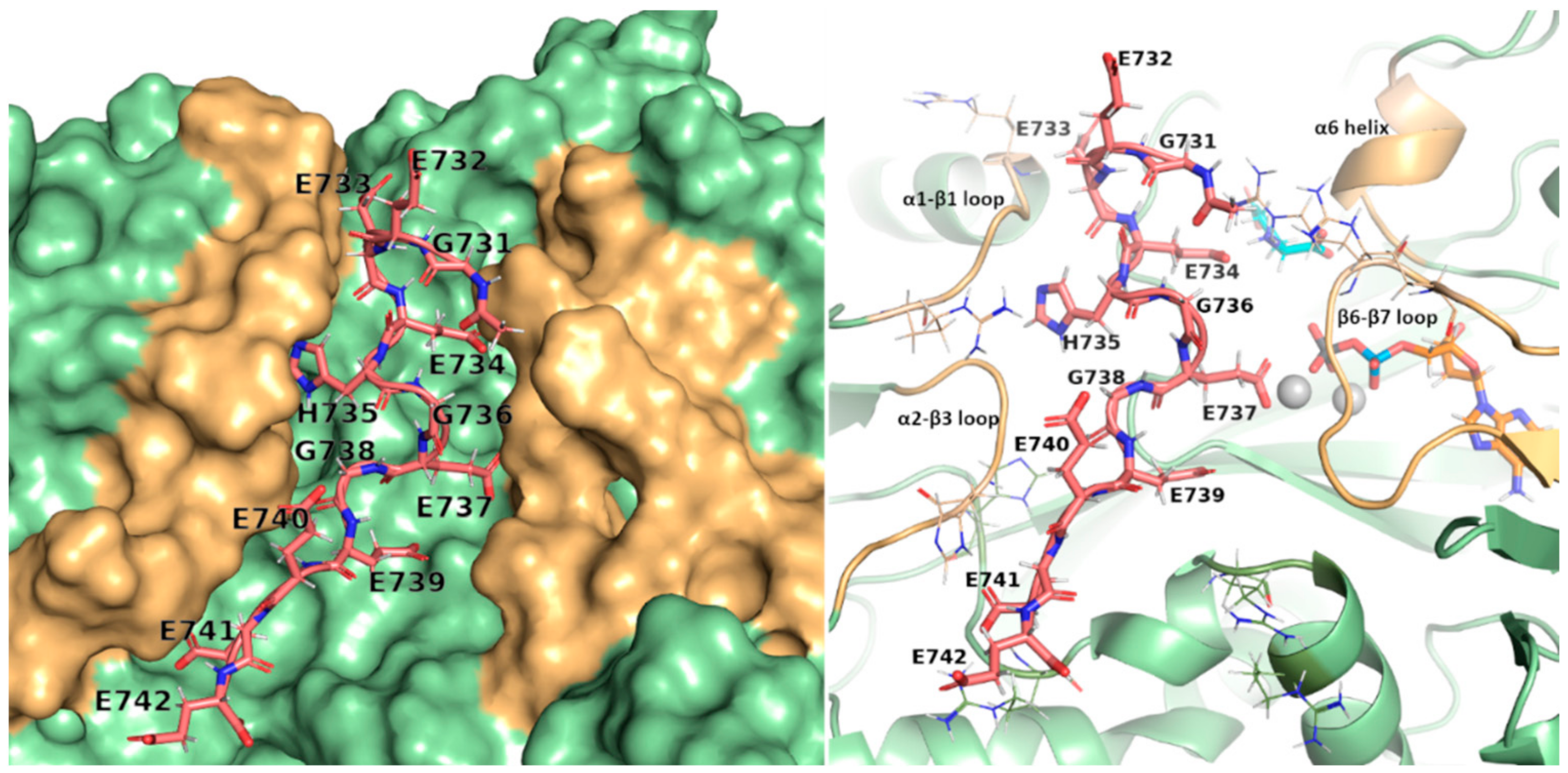
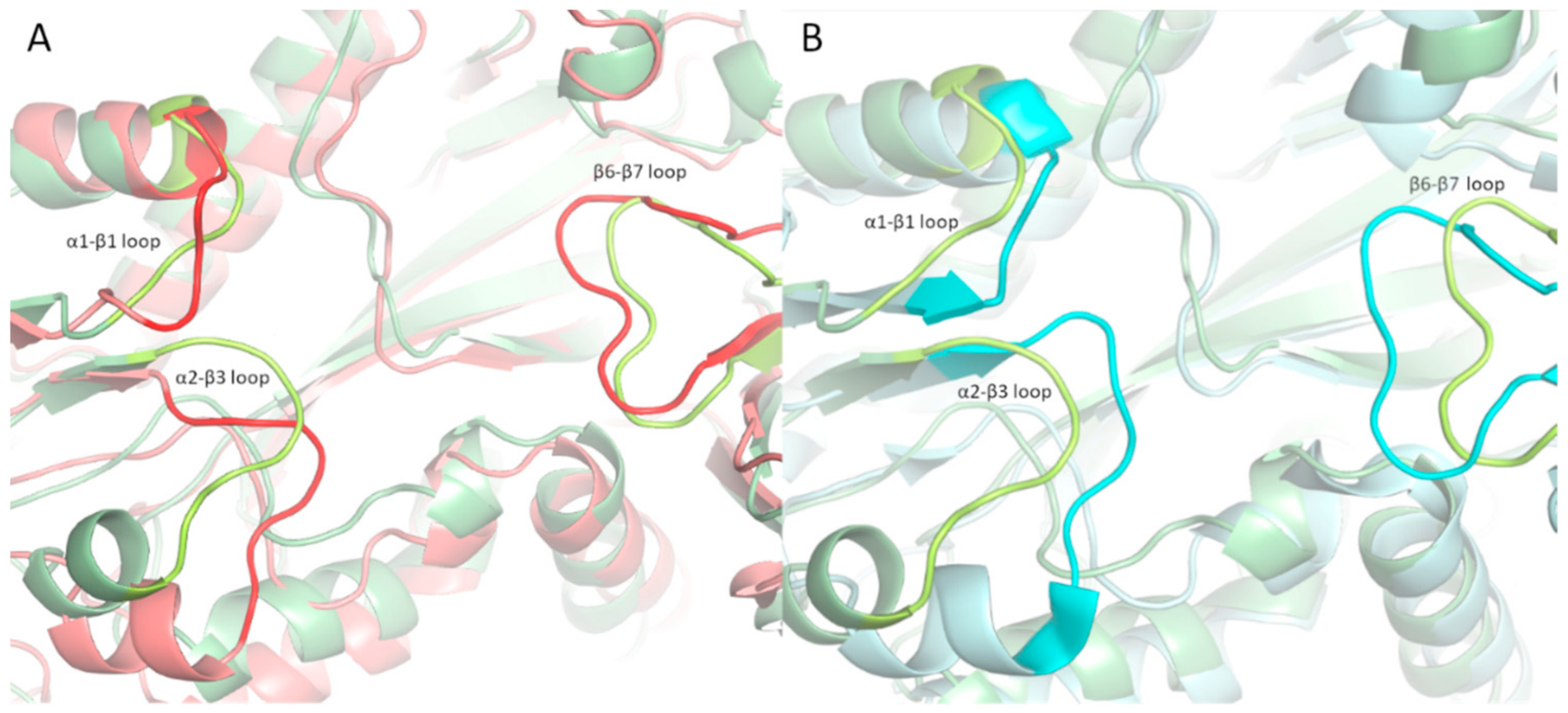
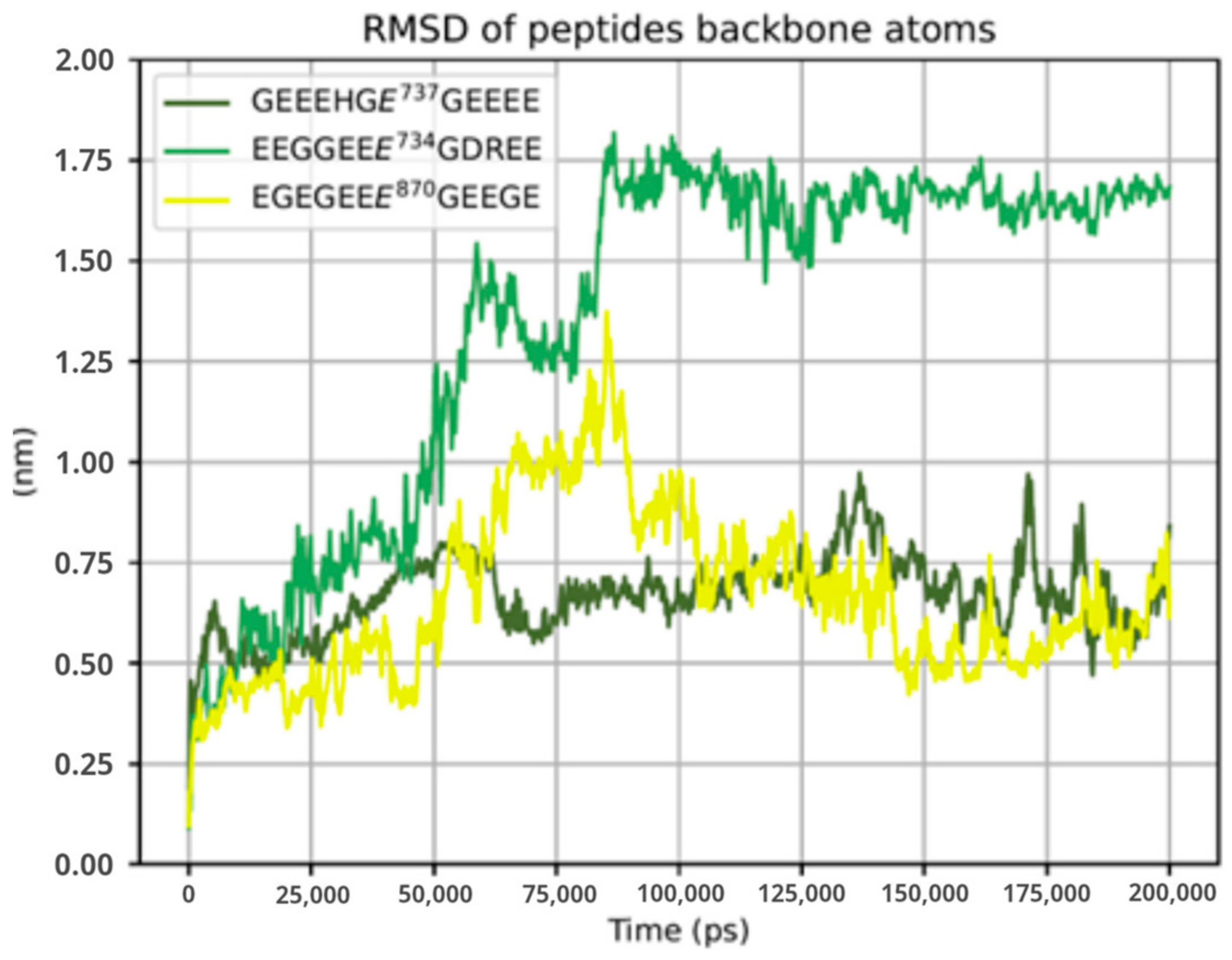
2.4. Structural Analysis of TTLL5 Core Domain Binding
3. Discussion
4. Materials and Methods
4.1. Subjects and Samples
4.2. NGS Sequencing
4.3. SMRTbell Library Preparation and TGS Sequencing
4.4. Bioinformatics and Genetic Variants Classification
4.5. Molecular Modeling of TTLL5-RPGRORF15 Interactions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, T.; Chitnis, N.; Monos, D.; Dinh, A. Next-generation sequencing technologies: An overview. Hum. Immunol. 2021, 82, 801–811. [Google Scholar] [CrossRef]
- van Dijk, E.L.; Jaszczyszyn, Y.; Naquin, D.; Thermes, C. The Third Revolution in Sequencing Technology. Trends Genet. 2018, 34, 666–681. [Google Scholar] [CrossRef]
- Logsdon, G.A.; Vollger, M.R.; Eichler, E.E. Long-read human genome sequencing and its applications. Nat. Rev. Genet. 2020, 21, 597–614. [Google Scholar] [CrossRef] [PubMed]
- Nanopore Sequencing Accuracy. Available online: https://nanoporetech.com/accuracy (accessed on 21 March 2023).
- Nguyen, X.-T.-A.; Talib, M.; van Schooneveld, M.J.; Brinks, J.; Ten Brink, J.; Florijn, R.J.; Wijnholds, J.; Verdijk, R.M.; Bergen, A.A.; Boon, C.J. RPGR-Associated Dystrophies: Clinical, Genetic, and Histopathological Features. Int. J. Mol. Sci. 2020, 21, 835. [Google Scholar] [CrossRef] [PubMed]
- Nassisi, M.; De Bartolo, G.; Mohand-Said, S.; Condroyer, C.; Antonio, A.; Lancelot, M.E.; Bujakowska, K.; Smirnov, V.; Pugliese, T.; Neidhardt, J.; et al. Retrospective Natural History Study of RPGR-Related Cone- and Cone-Rod Dystrophies While Expanding the Mutation Spectrum of the Disease. Int. J. Mol. Sci. 2022, 23, 7189. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Park, J.H.; Gumerson, J.; Wu, Z.; Swaroop, A.; Qian, H.; Roll-Mecak, A.; Li, T. Loss of RPGR glutamylation underlies the pathogenic mechanism of retinal dystrophy caused by TTLL5 mutations. Proc. Natl. Acad. Sci. USA 2016, 113, E2925–E2934. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.P.W.; Lamey, T.M.; Wang, N.K.; Duan, J.; Zhou, W.; McLaren, T.L.; Thompson, J.A.; Ruddle, J.; De Roach, J.N. Development of High-Throughput Clinical Testing of RPGR ORF15 Using a Large Inherited Retinal Dystrophy Cohort. Investig. Opthalmology Vis. Sci. 2018, 59, 4434. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.-B.; Hayakawa, M.; Murakami, A.; Nao-i, N. RCC1-Like Domain and ORF15: Essentials in RPGR Gene. In Retinal Degenerative Diseases; Springer: Boston, MA, USA, 2006; pp. 29–33. [Google Scholar] [CrossRef]
- Maggi, J.; Roberts, L.; Koller, S.; Rebello, G.; Berger, W.; Ramesar, R. De Novo Assembly-Based Analysis of RPGR Exon ORF15 in an Indigenous African Cohort Overcomes Limitations of a Standard Next-Generation Sequencing (NGS) Data Analysis Pipeline. Genes 2020, 11, 800. [Google Scholar] [CrossRef]
- The Human Gene Mutation Database. Available online: https://www.hgmd.cf.ac.uk/ac/index.php (accessed on 14 April 2023).
- Ensembl. Available online: https://www.ensembl.org/index.html (accessed on 14 April 2023).
- Yokoyama, A.; Maruiwa, F.; Hayakawa, M.; Kanai, A.; Vervoort, R.; Wright, A.F.; Yamada, K.; Niikawa, N.; Naōi, N. Three novel mutations of the RPGR gene exon ORF15 in three Japanese families with X-linked retinitis pigmentosa. Am. J. Med. Genet. 2001, 104, 232–238. [Google Scholar] [CrossRef]
- Oh, J.K.; Del Valle, J.G.V.; de Carvalho, J.R.L.; Sun, Y.J.; Levi, S.R.; Ryu, J.; Yang, J.; Nagasaki, T.; Emanuelli, A.; Rasool, N.; et al. Expanding the phenotype of TTLL5-associated retinal dystrophy: A case series. Orphanet J. Rare Dis. 2022, 17, 146. [Google Scholar] [CrossRef]
- Smirnov, V.; Grunewald, O.; Muller, J.; Zeitz, C.; Obermaier, C.D.; Devos, A.; Pelletier, V.; Bocquet, B.; Andrieu, C.; Bacquet, J.-L.; et al. Novel TTLL5 Variants Associated with Cone-Rod Dystrophy and Early-Onset Severe Retinal Dystrophy. Int. J. Mol. Sci. 2021, 22, 6410. [Google Scholar] [CrossRef]
- Del Pozo-Valero, M.; Riveiro-Alvarez, R.; Martin-Merida, I.; Blanco-Kelly, F.; Swafiri, S.; Lorda-Sanchez, I.; Trujillo-Tiebas, M.J.; Carreño, E.; Jimenez-Rolando, B.; Garcia-Sandoval, B.; et al. Impact of Next Generation Sequencing in Unraveling the Genetics of 1036 Spanish Families with Inherited Macular Dystrophies. Investig. Ophthalmol Vis. Sci 2022, 63, 11. [Google Scholar] [CrossRef]
- Mahalingan, K.K.; Keenan, E.K.; Strickland, M.; Li, Y.; Liu, Y.; Ball, H.L.; Tanner, M.E.; Tjandra, N.; Roll-Mecak, A. Structural basis for polyglutamate chain initiation and elongation by TTLL family enzymes. Nat. Struct. Mol. Biol. 2020, 27, 802–813. [Google Scholar] [CrossRef]
- Megaw, R.D.; Soares, D.C.; Wright, A.F. RPGR: Its role in photoreceptor physiology, human disease, and future therapies. Exp. Eye Res. 2015, 138, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Hussain, H.B.; Wilson, K.A.; Wetmore, S.D. Serine and Cysteine π-Interactions in Nature: A Comparison of the Frequency, Structure, and Stability of Contacts Involving Oxygen and Sulfur. Aust. J. Chem. 2015, 68, 385. [Google Scholar] [CrossRef]
- Natarajan, K.; Gadadhar, S.; Souphron, J.; Magiera, M.M.; Janke, C. Molecular interactions between tubulin tails and glutamylases reveal determinants of glutamylation patterns. EMBO Rep. 2017, 18, 1013–1026. [Google Scholar] [CrossRef]
- Colombo, L.; Maltese, P.E.; Castori, M.; El Shamieh, S.; Zeitz, C.; Audo, I.; Zulian, A.; Marinelli, C.; Benedetti, S.; Costantini, A.; et al. Molecular Epidemiology in 591 Italian Probands with Nonsyndromic Retinitis Pigmentosa and Usher Syndrome. Investig. Opthalmology Vis. Sci. 2021, 62, 13. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Xie, D.; Wang, L.; Kuang, Y.; Luo, S.; Ren, L.; Li, D.; Mao, A.; Li, J.; Chen, L.; et al. Application of third-generation sequencing for genetic testing of thalassemia in Guizhou Province, Southwest China. Hematology 2022, 27, 1305–1311. [Google Scholar] [CrossRef]
- Hassan, S.; Bahar, R.; Johan, M.F.; Hashim, E.K.M.; Abdullah, W.Z.; Esa, E.; Hamid, F.S.A.; Zulkafli, Z. Next-Generation Sequencing (NGS) and Third-Generation Sequencing (TGS) for the Diagnosis of Thalassemia. Diagnostics 2023, 13, 373. [Google Scholar] [CrossRef] [PubMed]
- dbSNP. Available online: https://www.ncbi.nlm.nih.gov/snp/ (accessed on 21 March 2023).
- ClinVar. Available online: https://www.ncbi.nlm.nih.gov/clinvar/ (accessed on 21 March 2023).
- Procedure & Checklist—Preparing SMRTbell® Libraries using PacBio® Barcoded Universal Primers for Multiplexing Amplicons. Available online: https://www.pacb.com/wp-content/uploads/Procedure-Checklist-Preparing-SMRTbell-Libraries-using-PacBio-Barcoded-Universal-Primers-for-Multiplexing-Amplicons.pdf (accessed on 21 March 2023).
- Mareso, C.; Albion, E.; Cozza, W.; Tanzi, B.; Cecchin, S.; Gisondi, P.; Michelini, S.; Bellinato, F.; Michelini, S.; Michelini, S.; et al. Optimization of long-range PCR protocol to prepare filaggrin exon 3 libraries for PacBio long-read sequencing. Mol. Biol. Rep. 2023, 50, 3119–3127. [Google Scholar] [CrossRef]
- Primer3web. Available online: https://primer3.ut.ee (accessed on 21 March 2023).
- Primer-BLAST. Available online: https://www.ncbi.nlm.nih.gov/tools/primer-blast (accessed on 21 March 2023).
- UCSC In-Silico PCR. Available online: https://genome.ucsc.edu/cgi-bin/hgPcr (accessed on 21 March 2023).
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and guidelines for the interpretation of sequence variants: A joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- VarSome. Available online: https://varsome.com/ (accessed on 21 March 2023).
- GnomAD. Available online: https://gnomad.broadinstitute.org/ (accessed on 21 March 2023).
- Cristofoli, F.; Sorrentino, E.; Guerri, G.; Miotto, R.; Romanelli, R.; Zulian, A.; Cecchin, S.; Paolacci, S.; Miertus, J.; Bertelli, M.; et al. Variant Selection and Interpretation: An Example of Modified VarSome Classifier of ACMG Guidelines in the Diagnostic Setting. Genes 2021, 12, 1885. [Google Scholar] [CrossRef] [PubMed]
- Mattassi, R.; Manara, E.; Colombo, P.G.; Manara, S.; Porcella, A.; Bruno, G.; Bruson, A.; Bertelli, M. Variant Discovery in Patients with Mendelian Vascular Anomalies by Next-Generation Sequencing and Their Use in Patient Clinical Management. J. Vasc. Surg. 2018, 67, 922–932.e11. [Google Scholar] [CrossRef] [PubMed]
- Marceddu, G.; Dallavilla, T.; Guerri, G.; Zulian, A.; Marinelli, C.; Bertelli, M. Analysis of Machine Learning Algorithms as Integrative Tools for Validation of next Generation Sequencing Data. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 8139–8147. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, E.; Albion, E.; Modena, C.; Daja, M.; Cecchin, S.; Paolacci, S.; Miertus, J.; Bertelli, M.; Maltese, P.E.; Chiurazzi, P.; et al. PacMAGI: A pipeline including accurate indel detection for the analysis of PacBio sequencing data applied to RPE65. Gene 2022, 832, 146554. [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- MAGI-Dock. Available online: https://github.com/gjonwick/MAGI-Dock (accessed on 29 May 2023).
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Case Subjects (n = 75) | |
|---|---|---|
| Age (years) | Mean | 51 ± 17 |
| Median | 52 ± 17 | |
| Females/Males | 35/40 (47%/53%) | |
| Diagnosis | Hereditary non-syndromic retinal dystrophies | 5 (7%) |
| Retinitis pigmentosa | 56 (75%) | |
| Cone dystrophy | 8 (10%) | |
| Macular distrophy | 6 (8%) | |
| Age of onset (years) | Mean | 25 ± 16 |
| Median | 24 ± 16 | |
| Unknown | n = 8 | |
| Familiarity | Familiar | 26 (35%) |
| Sporadic | 40 (53%) | |
| Unknown | 9 (12%) |
| ID | Nucleotide Variant | rsID | Verdict | Zygosity |
|---|---|---|---|---|
| 1 | NM_001034853.2:c.2919_2940dup | / | Likely Pathogenic | 0/1 |
| 2 | NM_001034853.2:c.2203_2226del | rs768423834 | Uncertain Significance | 0/1 |
| 3 | NM_001034853.2:c.2203_2226del | rs768423834 | Uncertain Significance | 1/1 |
| 4 | NM_001034853.2:c.2820_2841dup | / | Likely Pathogenic | 0/1 |
| 5 | NM_001034853:c.1871A>C | / | Uncertain Significance | 0/1 |
| 6 | NM_001034853:c.3423G>T | / | Uncertain Significance | 1/1 |
| 7 | NM_001034853.2:c.3262_3263insA | / | Likely Pathogenic | 0/1 |
| 8 | NM_001034853.2:c.2820_2841dup | / | Likely Pathogenic | 0/1 |
| Characteristics | Patients with RPGRORF15 Mutation (n = 8) | Case Subjects (n = 67) | p-Value | |
|---|---|---|---|---|
| Age (years) | Mean | 35 ± 11 | 53 ± 15 | <0.05 |
| Median | 36 ± 11 | 53 ± 15 | ||
| Females/Males | 4/4 (50%/50%) | 31/36 (46%/54%) | - | |
| Diagnosis | Hereditary non-syndromic retinal dystrophies | 0 (0%) | 5 (7%) | - |
| Retinitis pigmentosa | 3 (37.5%) | 53 (79%) | ||
| Cone dystrophy | 2 (25%) | 6 (9%) | ||
| Macular dystrophy | 3 (37.5%) | 3 (5%) | ||
| Age of onset (years) | Mean | 13 ±11 | 27 ± 16 | <0.05 |
| Median | 10 ± 11 | 27 ± 16 | ||
| Unknown | n = 0 | n = 8 | ||
| Familiarity | Familiar | 2 (25%) | 24 (36%) | - |
| Sporadic | 3 (37.5%) | 37 (55%) | ||
| Unknown | 3 (37.5%) | 6 (9%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonetti, G.; Cozza, W.; Bernini, A.; Kaftalli, J.; Mareso, C.; Cristofoli, F.; Medori, M.C.; Colombo, L.; Martella, S.; Staurenghi, G.; et al. Towards a Long-Read Sequencing Approach for the Molecular Diagnosis of RPGRORF15 Genetic Variants. Int. J. Mol. Sci. 2023, 24, 16881. https://doi.org/10.3390/ijms242316881
Bonetti G, Cozza W, Bernini A, Kaftalli J, Mareso C, Cristofoli F, Medori MC, Colombo L, Martella S, Staurenghi G, et al. Towards a Long-Read Sequencing Approach for the Molecular Diagnosis of RPGRORF15 Genetic Variants. International Journal of Molecular Sciences. 2023; 24(23):16881. https://doi.org/10.3390/ijms242316881
Chicago/Turabian StyleBonetti, Gabriele, William Cozza, Andrea Bernini, Jurgen Kaftalli, Chiara Mareso, Francesca Cristofoli, Maria Chiara Medori, Leonardo Colombo, Salvatore Martella, Giovanni Staurenghi, and et al. 2023. "Towards a Long-Read Sequencing Approach for the Molecular Diagnosis of RPGRORF15 Genetic Variants" International Journal of Molecular Sciences 24, no. 23: 16881. https://doi.org/10.3390/ijms242316881