
Associations between Host Genetic Variants and Subgingival Microbiota in Patients with the Metabolic Syndrome

 , , and
, , and
Abstract
:1. Introduction
2. Results
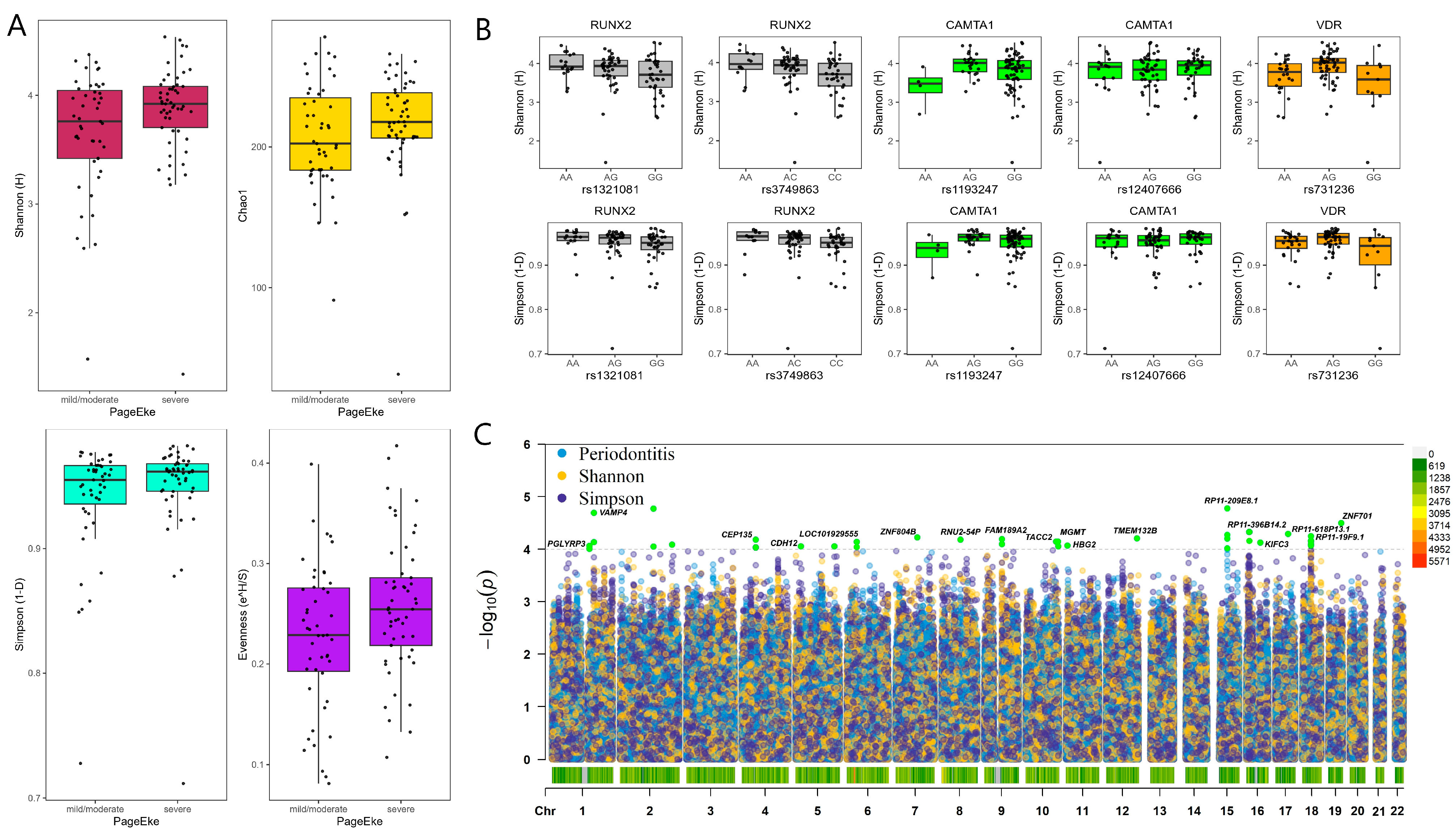
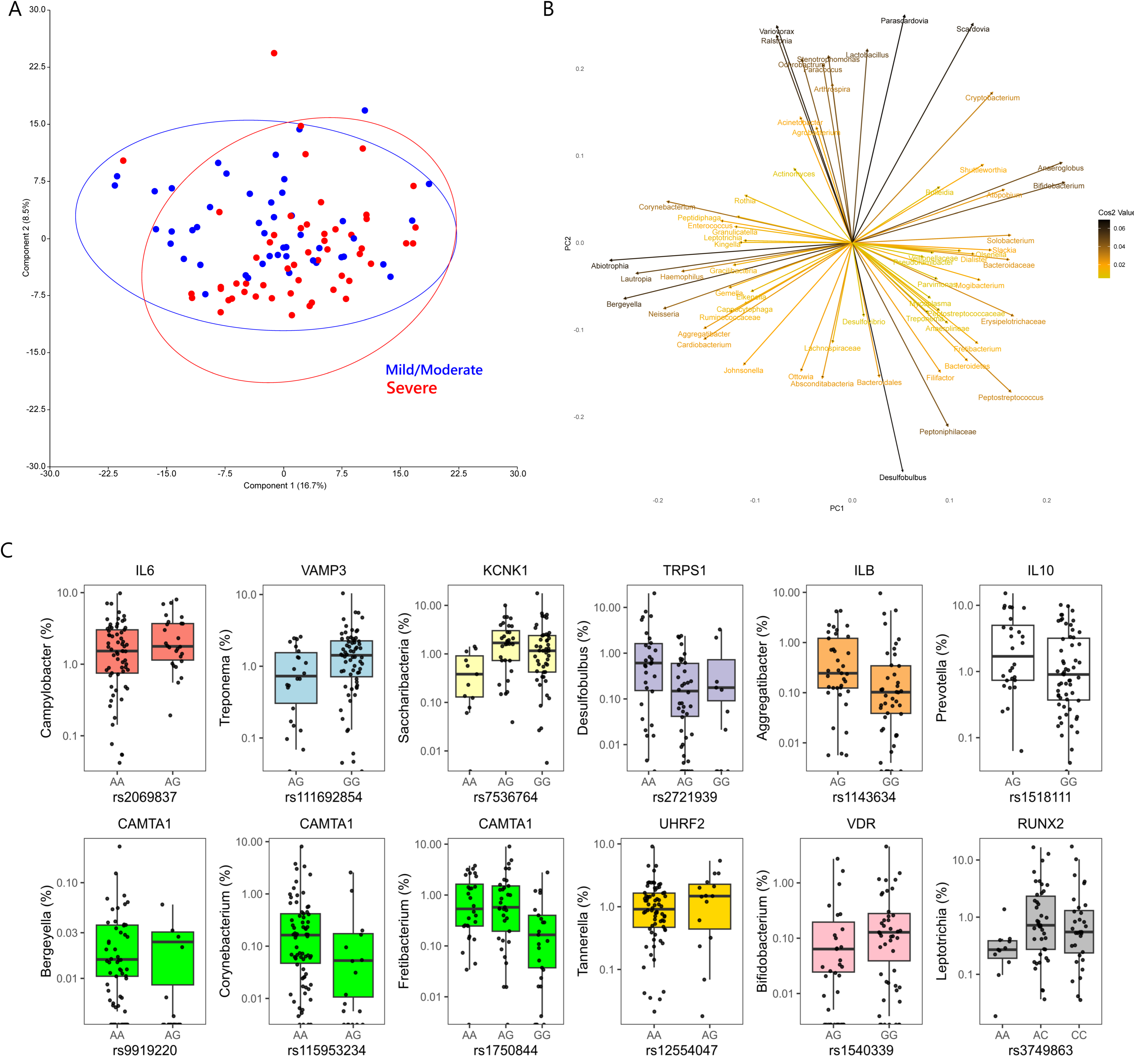
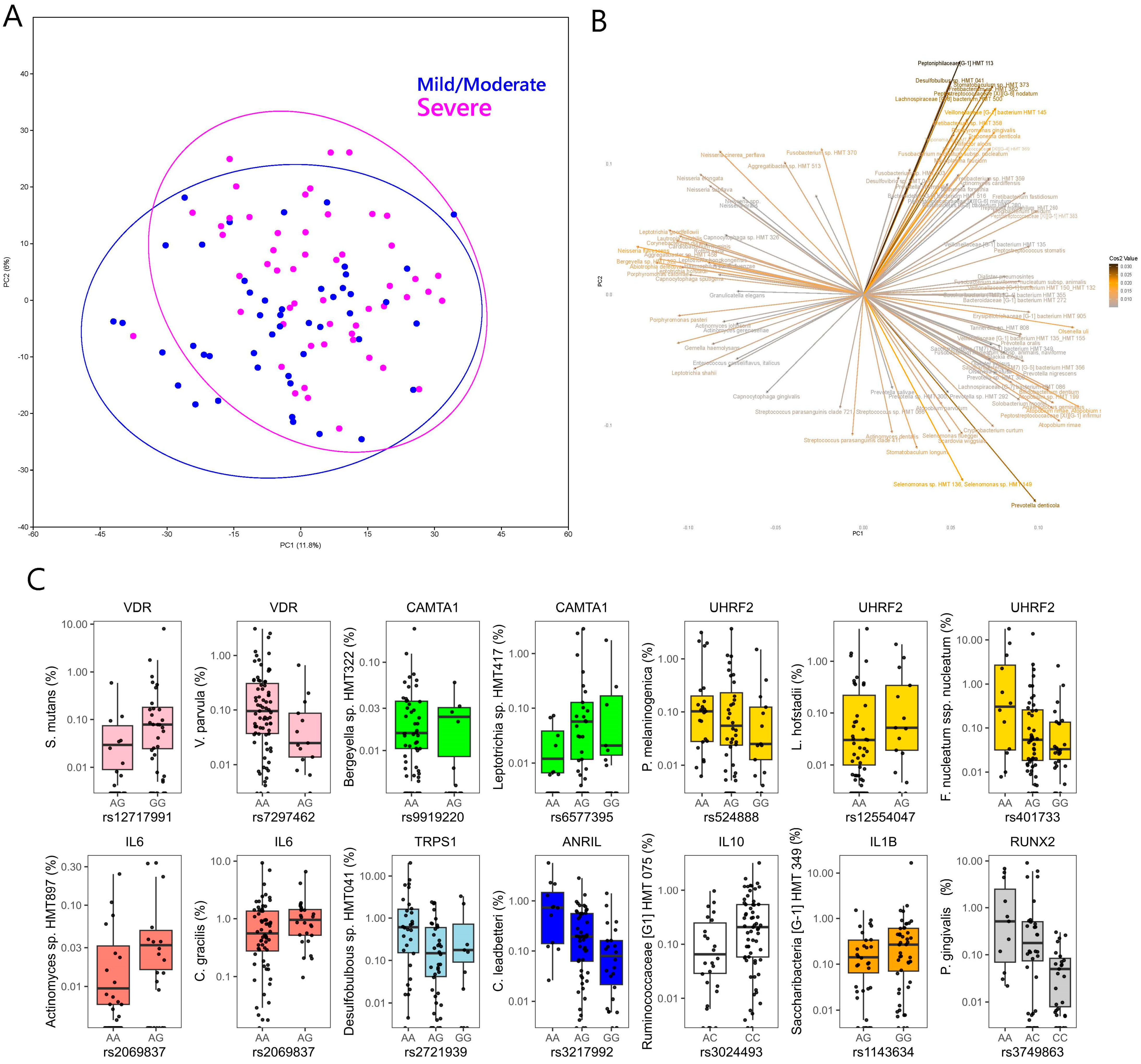
2.1. Microbial Diversity and Metabolic Syndrome
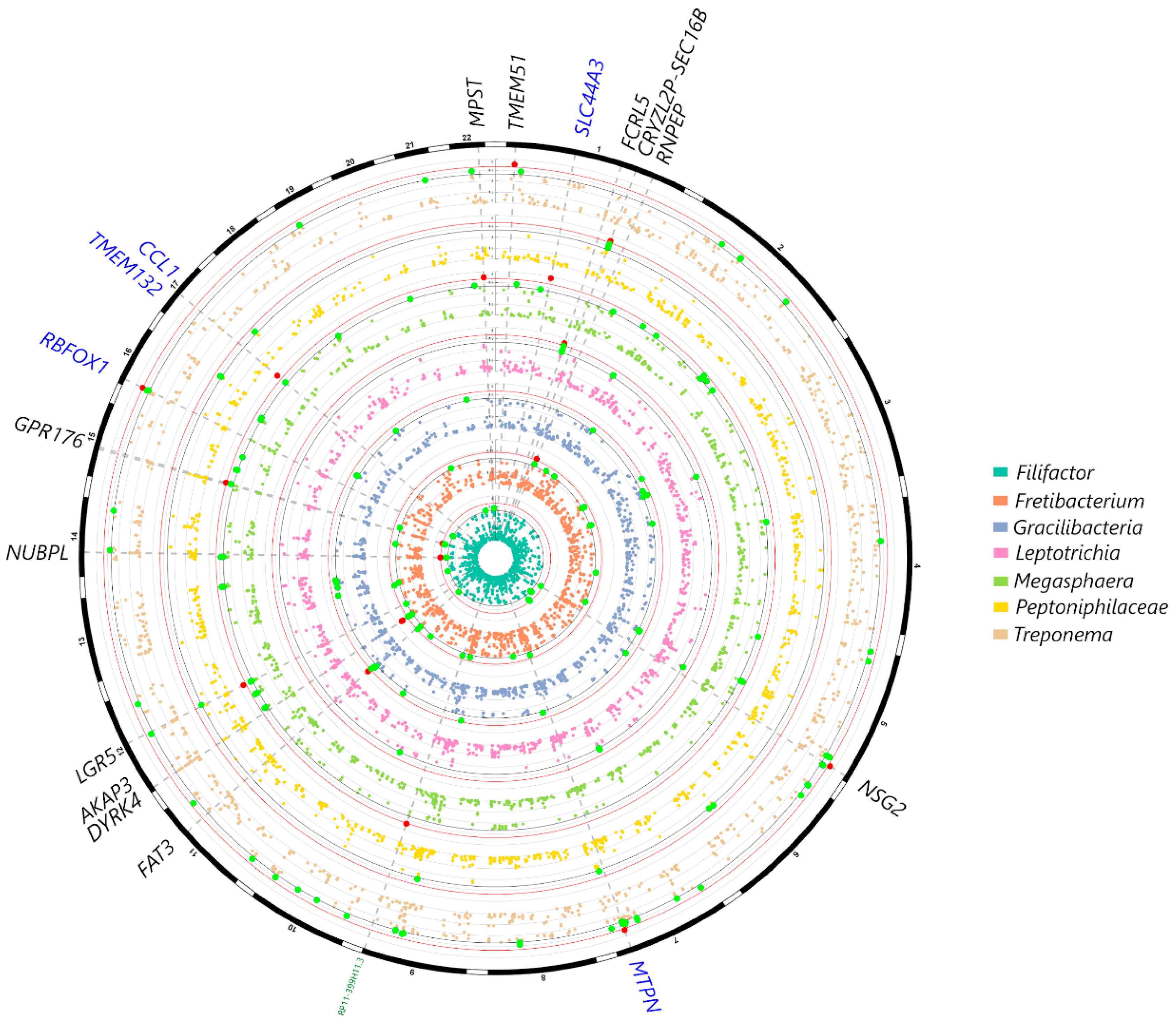
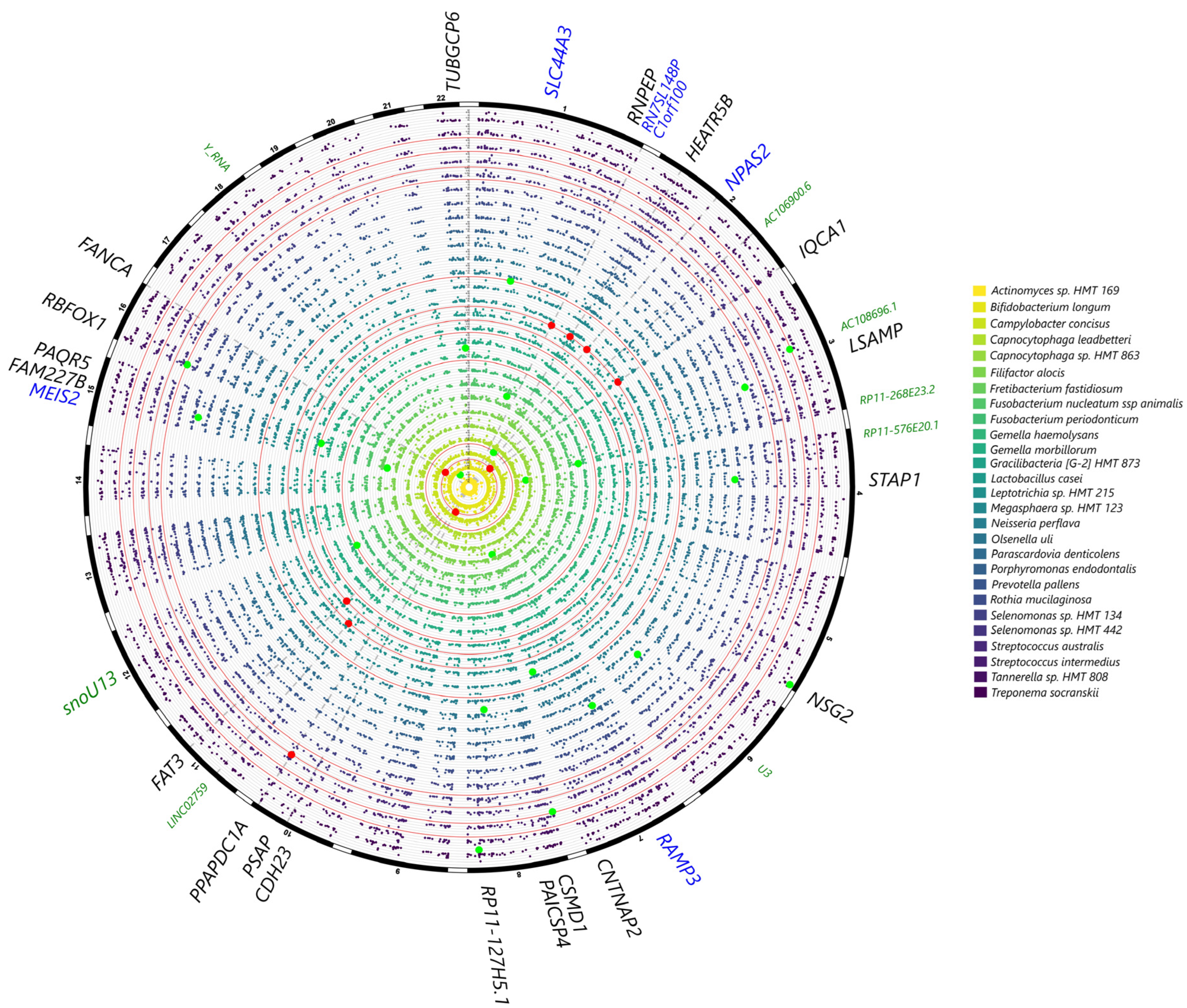
2.2. Genotype Associations with Microbial Taxa
2.2.1. Genotype Associations at the Genus Level
2.2.2. Genotype Associations at the Species Level
3. Discussion
4. Materials and Methods
4.1. Study Population
- Caucasian ethnicity;
- Age 25–75;
- Diagnosis of metabolic syndrome as defined by the revised NCEP ATP III (e.g., the presence of at least 3 of the following factors) [48]:
- Waist circumference > 102 cm for men and >88 cm for women;
- High triglycerides: ≥150 mg/dL (1.7 mmol/L), or specific treatment for this lipid abnormality;
- Low HDL cholesterol: <40 mg/dL (1.03 mmol/L) in males, <50 mg/dL (1.29 mmol/L) in females, or specific treatment for this lipid abnormality;
- High blood pressure: systolic BP ≥ 130 or diastolic BP ≥ 85 mm Hg, or treatment of previously diagnosed hypertension;
- High fasting plasma glucose: FPG ≥ 100 mg/dL (5.6 mmol/L), or previously diagnosed type 2 diabetes.
- Presence of at least 12 teeth.
- Pregnancy;
- Presence of infectious diseases such as hepatitis and HIV;
- Antibiotic pre-medication required for the performance of periodontal examination;
- Previous periodontal therapy within 6 months of the study visit.
4.2. Medical Assessment and Sampling
- Healthy/mild periodontitis: <2 sites on different teeth with CAL ≥ 4 mm or no sites with PPD ≥ 4 mm;
- Moderate periodontitis: ≥2 sites on different teeth with CAL ≥ 4 mm or one site with PPD ≥ 4 mm;
- Severe periodontitis: ≥2 sites on different teeth with CAL ≥ 6 mm and ≥1 site with PPD ≥ 4 mm.
4.3. Genotyping, Imputation and Genome-Wide Association Analysis
4.3.1. DNA Extraction and Genotyping
4.3.2. Pre-Imputation Quality Control and Imputation
4.3.3. Post-Imputation Quality Control and Statistical Analysis
4.4. Microbiome Profiling
4.4.1. DNA Extraction and Sequencing
4.4.2. Sequence Analysis and Taxonomic Classification
4.4.3. Microbial Diversity and Multivariate Analysis
4.5. Statistical Analysis/Power Calculation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kellam:, P.; Weiss, R.A. Infectogenomics: Insights from the Host Genome into Infectious Diseases. Cell 2006, 124, 695–697. [Google Scholar] [CrossRef]
- Nibali, L.; Henderson, B.; Sadiq, S.T.; Donos, N. Genetic dysbiosis: The role of microbial insults in chronic inflammatory diseases. J. Oral Microbiol. 2014, 6, 22962. [Google Scholar] [CrossRef]
- Zoheir, N.; Kurushima, Y.; Lin, G.-H.; Nibali, L. Periodontal infectogenomics: A systematic review update of associations between host genetic variants and subgingival microbial detection. Clin. Oral Investig. 2022, 26, 2209–2221. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Lamont, R.J. Beyond the red complex and into more complexity: The polymicrobial synergy and dysbiosis (PSD) model of periodontal disease etiology. Mol. Oral Microbiol. 2012, 27, 409–419. [Google Scholar] [CrossRef]
- Socransky, S.S.; Haffajee, A.D. Periodontal microbial ecology. Periodontology 2000 2005, 38, 135–187. [Google Scholar] [CrossRef]
- Nibali, L.; Ready, D.; Parkar, M.; Brett, P.; Wilson, M.; Tonetti, M.; Griffiths, G. Gene Polymorphisms and the Prevalence of Key Periodontal Pathogens. J. Dent. Res. 2007, 86, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Nibali, L.; Donos, N.; Brett, P.M.; Parkar, M.; Ellinas, T.; Llorente, M.; Griffiths, G.S. A familial analysis of aggressive periodontitis—Clinical and genetic findings. J. Periodontal Res. 2008, 43, 627–634. [Google Scholar] [CrossRef]
- Divaris, K.; Monda, K.L.; North, K.E.; Olshan, A.F.; Reynolds, L.M.; Hsueh, W.-C.; Lange, E.M.; Moss, K.; Barros, S.P.; Weyant, R.J.; et al. Exploring the genetic basis of chronic periodontitis: A genome-wide association study. Hum. Mol. Genet. 2013, 22, 2312–2324. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Carlsson, G.; Wondimu, B.; Fahlén, A.; Karlsson-Sjöberg, J.; Andersson, M.; Engstrand, L.; Yucel-Lindberg, T.; Modéer, T.; Pütsep, K. Mutations in the ELANE Gene are Associated with Development of Periodontitis in Patients with Severe Congenital Neutropenia. J. Clin. Immunol. 2011, 31, 936–945. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yu, N.; Arce, R.M. Periodontal inflammation: Integrating genes and dysbiosis. Periodontol. 2000 2019, 82, 129–142. [Google Scholar] [CrossRef]
- Nibali, L.; Donos, N.; Terranova, V.; Di Pino, A.; Di Marca, S.; Ferrara, V.; Pisano, M.; Scicali, R.; Rabuazzo, A.M.; Purrello, F.; et al. Left ventricular geometry and periodontitis in patients with the metabolic syndrome. Clin. Oral Investig. 2018, 23, 2695–2703. [Google Scholar] [CrossRef] [PubMed]
- Zanoli, L.; Ozturk, K.; Cappello, M.; Inserra, G.; Geraci, G.; Tuttolomondo, A.; Torres, D.; Pinto, A.; Duminuco, A.; Riguccio, G.; et al. Inflammation and Aortic Pulse Wave Velocity: A Multicenter Longitudinal Study in Patients With Inflammatory Bowel Disease. J. Am. Heart Assoc. 2019, 8, e010942. [Google Scholar] [CrossRef] [PubMed]
- Nibali, L.; Tatarakis, N.; Needleman, I.; Tu, Y.-K.; D’Aiuto, F.; Rizzo, M.; Donos, N. Association Between Metabolic Syndrome and Periodontitis: A Systematic Review and Meta-analysis. J. Clin. Endocrinol. Metab. 2013, 98, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Suwanprasit, W.; Lertpimonchai, A.; Thienpramuk, L.; Vathesatogkit, P.; Sritara, P.; Tamsailom, S. Metabolic syndrome and severe periodontitis were associated in Thai adults: A cross-sectional study. J. Periodontol. 2021, 92, 1420–1429. [Google Scholar] [CrossRef] [PubMed]
- Lourenςo, T.G.B.; Spencer, S.J.; Alm, E.J.; Colombo, A.P.V. Defining the gut microbiota in individuals with periodontal diseases: An exploratory study. J. Oral Microbiol. 2018, 10, 1487741. [Google Scholar] [CrossRef]
- Olsen, I.; Yamazaki, K. Can oral bacteria affect the microbiome of the gut? J. Oral Microbiol. 2019, 11, 1586422. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Li, L.; Zhang, Y.; Wang, M.; Chen, F.; Ge, S.; Chen, B.; Yan, F. Periodontitis may induce gut microbiota dysbiosis via salivary microbiota. Int. J. Oral Sci. 2022, 14, 32. [Google Scholar] [CrossRef]
- Page, R.C.; Eke, P.I. Case Definitions for Use in Population-Based Surveillance of Periodontitis. J. Periodontol. 2007, 78, 1387–1399. [Google Scholar] [CrossRef]
- Tu, Y.; Ling, X.; Chen, Y.; Wang, Y.; Zhou, N.; Chen, H. Effect of S. Mutans and S. Sanguinis on Growth and Adhesion of P. Gingivalis and Their Ability to Adhere to Different Dental Materials. Experiment 2017, 23, 4539–5445. [Google Scholar] [CrossRef]
- Luppens, S.B.I.; Kara, D.; Bandounas, L.; Jonker, M.J.; Wittink, F.R.A.; Bruning, O.; Breit, T.M.; Cate, J.M.T.; Crielaard, W. Effect of Veillonella parvula on the antimicrobial resistance and gene expression of Streptococcus mutans grown in a dual-species biofilm. Oral Microbiol. Immunol. 2008, 23, 183–189. [Google Scholar] [CrossRef]
- Liu, S.; Chen, M.; Wang, Y.; Zhou, X.; Peng, X.; Ren, B.; Li, M.; Cheng, L. Effect of Veillonella parvula on the physiological activity of Streptococcus mutans. Arch. Oral Biol. 2020, 109, 104578. [Google Scholar] [CrossRef]
- Van Dyke, T.E.; Bartold, P.M.; Reynolds, E.C. The Nexus Between Periodontal Inflammation and Dysbiosis. Front. Immunol. 2020, 11, 511. [Google Scholar] [CrossRef]
- Kurushima, Y.; Wells, P.; Bowyer, R.; Zoheir, N.; Doran, S.; Richardson, J.; Sprockett, D.; Relman, D.; Steves, C.; Nibali, L. Host Genotype Links to Salivary and Gut Microbiota by Periodontal Status. J. Dent. Res. 2022, 102, 146–156. [Google Scholar] [CrossRef] [PubMed]
- Torrungruang, K.; Chantarangsu, S.; Sura, T.; Thienpramuk, L. Interplay between vitamin D receptor FokI polymorphism and smoking influences Porphyromonas gingivalis proportions in subgingival plaque. J. Clin. Periodontol. 2020, 47, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Nibali, L.; Madden, I.; Chillida, F.F.; Heitz-Mayfield, L.; Brett, P.; Donos, N. IL6 −174 Genotype Associated with Aggregatibacter actinomycetemcomitans in Indians. Oral Dis. 2010, 17, 232–237. [Google Scholar] [CrossRef]
- Schaefer, A.S.; Bochenek, G.; Jochens, A.; Ellinghaus, D.; Dommisch, H.; Güzeldemir-Akçakanat, E.; Graetz, C.; Harks, I.; Jockel-Schneider, Y.; Weinspach, K.; et al. Genetic Evidence for PLASMINOGEN as a Shared Genetic Risk Factor of Coronary Artery Disease and Periodontitis. Circ. Cardiovasc. Genet. 2015, 8, 159–167. [Google Scholar] [CrossRef]
- Aarabi, G.; Zeller, T.; Seedorf, H.; Reissmann, D.; Heydecke, G.; Schaefer, A.; Seedorf, U. Genetic Susceptibility Contributing to Periodontal and Cardiovascular Disease. J. Dent. Res. 2017, 96, 610–617. [Google Scholar] [CrossRef]
- Li, N.; Li, Y.; Hu, J.; Wu, Y.; Yang, J.; Fan, H.; Li, L.; Luo, D.; Ye, Y.; Gao, Y.; et al. A Link Between Mitochondrial Dysfunction and the Immune Microenvironment of Salivary Glands in Primary Sjogren’s Syndrome. Front. Immunol. 2022, 13, 845209. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Hong, J.; Sharma, A.; Wang, B. Streptococcus cristatus ArcA interferes with Porphyromonas gingivalis pathogenicity in mice. J. Periodontal Res. 2012, 47, 578–583. [Google Scholar] [CrossRef]
- Ho, M.-H.; Lamont, R.J.; Xie, H. Identification of Streptococcus cristatus peptides that repress expression of virulence genes in Porphyromonas gingivalis. Sci. Rep. 2017, 7, 1413. [Google Scholar] [CrossRef]
- Karched, M.; Bhardwaj, R.G.; Asikainen, S.E. Coaggregation and biofilm growth of Granulicatella spp. with Fusobacterium nucleatum and Aggregatibacter actinomycetemcomitans. BMC Microbiol. 2015, 15, 114. [Google Scholar] [CrossRef]
- Haffajee, A.D.; Socransky, S.S.; Smith, C.; Dibart, S. Relation of baseline microbial parameters to future periodontal attachment loss. J. Clin. Periodontol. 1991, 18, 744–750. [Google Scholar] [CrossRef]
- Haffajee, A.D.; Socransky, S.S.; Dibart, S.; Kent, R.L. Response to periodontal therapy in patients with high or low levels of P. gingivalis, P. intermedia, R nigrescens and B. forsythus. J. Clin. Periodontol. 1996, 23, 336–345. [Google Scholar] [CrossRef]
- Socransky, S.S.; Haffajee, A.D.; Cugini, M.A.; Smith, C.; Kent, R.L., Jr. Microbial complexes in subgingival plaque. J. Clin. Periodontol. 1998, 25, 134–144. [Google Scholar] [CrossRef]
- Oliveira, R.; Fermiano, D.; Feres, M.; Figueiredo, L.; Teles, F.; Soares, G.; Faveri, M. Levels of Candidate Periodontal Pathogens in Subgingival Biofilm. J. Dent. Res. 2016, 95, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Ghayoumi, N.; Chen, C.; Slots, J. Dialister pneumosintes, a new putative jreiodontal pathogen. J. Periodontal Res. 2002, 37, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Aruni, A.W.; Mishra, A.; Dou, Y.; Chioma, O.; Hamilton, B.N.; Fletcher, H.M. Filifactor alocis—A new emerging periodontal pathogen. Microbes Infect. 2015, 17, 517–530. [Google Scholar] [CrossRef]
- Ozuna, H.; Snider, I.; Belibasakis, G.N.; Oscarsson, J.; Johansson, A.; Uriarte, S.M. Aggregatibacter actinomycetemcomitans and Filifactor alocis: Two exotoxin-producing oral pathogens. Front. Oral Health 2022, 3, 981343. [Google Scholar] [CrossRef]
- Razooqi, Z.; Åberg, C.H.; Kwamin, F.; Claesson, R.; Haubek, D.; Oscarsson, J.; Johansson, A. Aggregatibacter actinomycetemcomitans and Filifactor alocis as Associated with Periodontal Attachment Loss in a Cohort of Ghanaian Adolescents. Microorganisms 2022, 10, 2511. [Google Scholar] [CrossRef]
- Peña-Chilet, M.; Esteban-Medina, M.; Falco, M.M.; Rian, K.; Hidalgo, M.R.; Loucera, C.; Dopazo, J. Using mechanistic models for the clinical interpretation of complex genomic variation. Sci. Rep. 2019, 9, 18937. [Google Scholar] [CrossRef] [PubMed]
- Ke, J.; Gao, W.; Wang, B.; Cao, W.; Lv, J.; Yu, C.; Huang, T.; Sun, D.; Liao, C.; Pang, Y.; et al. Exploring the Genetic Association between Obesity and Serum Lipid Levels Using Bivariate Methods. Twin Res. Hum. Genet. 2023, 25, 234–244. [Google Scholar] [CrossRef]
- de Vries, P.S.; van Herpt, T.T.W.; Ligthart, S.; Hofman, A.; Ikram, M.A.; van Hoek, M.; Sijbrands, E.J.G.; Franco, O.H.; de Maat, M.P.M.; Leebeek, F.W.G.; et al. ADAMTS13 activity as a novel risk factor for incident type 2 diabetes mellitus: A population-based cohort study. Diabetologia 2016, 60, 280–286. [Google Scholar] [CrossRef]
- Zillikens, M.C.; Demissie, S.; Hsu, Y.-H.; Yerges-Armstrong, L.M.; Chou, W.-C.; Stolk, L.; Livshits, G.; Broer, L.; Johnson, T.; Koller, D.L.; et al. Large meta-analysis of genome-wide association studies identifies five loci for lean body mass. Nat. Commun. 2017, 8, 80. [Google Scholar] [CrossRef]
- Zhu, Z.; Guo, Y.; Shi, H.; Liu, C.-L.; Panganiban, R.A.; Chung, W.; O’Connor, L.J.; Himes, B.E.; Gazal, S.; Hasegawa, K.; et al. Shared genetic and experimental links between obesity-related traits and asthma subtypes in UK Biobank. J. Allergy Clin. Immunol. 2019, 145, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Baca, P.; Barajas-Olmos, F.; Mirzaeicheshmeh, E.; Zerrweck, C.; Guilbert, L.; Sánchez, E.C.; Flores-Huacuja, M.; Villafán, R.; Martínez-Hernández, A.; García-Ortiz, H.; et al. DNA methylation and gene expression analysis in adipose tissue to identify new loci associated with T2D development in obesity. Nutr. Diabetes 2022, 12, 50. [Google Scholar] [CrossRef] [PubMed]
- Valenti, C.; Pagano, S.; Bozza, S.; Ciurnella, E.; Lomurno, G.; Capobianco, B.; Coniglio, M.; Cianetti, S.; Marinucci, L. Use of the Er:YAG Laser in Conservative Dentistry: Evaluation of the Microbial Population in Carious Lesions. Materials 2021, 14, 2387. [Google Scholar] [CrossRef] [PubMed]
- Nibali, L.; Stephen, A.; Hagi-Pavli, E.; Allaker, R.; Di Pino, A.; Terranova, V.; Pisano, M.; Di Marca, S.; Ferrara, V.; Scicali, R.; et al. Analysis of gingival crevicular fluid biomarkers in patients with metabolic syndrome. J. Dent. 2022, 118, 104065. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Brewer, H.B., Jr.; Cleeman, J.I.; Smith, S.C., Jr.; Lenfant, C. Definition of Metabolic Syndrome: Report of the National Heart, Lung, and Blood Institute/American Heart Association conference on scientific issues related to definition. Circulation 2004, 109, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Guerrero, A.; Griffiths, G.S.; Nibali, L.; Suvan, J.; Moles, D.R.; Laurell, L.; Tonetti, M.S. Adjunctive benefits of systemic amoxicillin and metronidazole in non-surgical treatment of generalized aggressive periodontitis: A randomized placebo-controlled clinical trial. J. Clin. Periodontol. 2005, 32, 1096–1107. [Google Scholar] [CrossRef]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. GigaScience 2015, 4, 7. [Google Scholar] [CrossRef]
- Loh, P.-R.; Danecek, P.; Palamara, P.F.; Fuchsberger, C.; Reshef, Y.A.; Finucane, H.K.; Schoenherr, S.; Forer, L.; McCarthy, S.; Abecasis, C.F.G.R.; et al. Reference-based phasing using the Haplotype Reference Consortium panel. Nat. Genet. 2016, 48, 1443–1448. [Google Scholar] [CrossRef]
- Durbin, R. Efficient haplotype matching and storage using the positional Burrows–Wheeler transform (PBWT). Bioinformatics 2014, 30, 1266–1272. [Google Scholar] [CrossRef]
- McCarthy, S.; Das, S.; Kretzschmar, W.; Delaneau, O.; Wood, A.R.; Teumer, A.; Kang, H.M.; Fuchsberger, C.; Danecek, P.; Sharp, K.; et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Zhang, H.; Tang, Z.; Xu, J.; Yin, D.; Zhang, Z.; Yuan, X.; Zhu, M.; Zhao, S.; Li, X.; et al. rMVP: A Memory-efficient, Visualization-enhanced, and Parallel-accelerated Tool for Genome-wide Association Study. Genom. Proteom. Bioinform. 2021, 19, 619–628. [Google Scholar] [CrossRef]
- Oscanoa, J.; Sivapalan, L.; Gadaleta, E.; Ullah, A.Z.D.; Lemoine, N.R.; Chelala, C. SNPnexus: A web server for functional annotation of human genome sequence variation (2020 update). Nucleic Acids Res. 2020, 48, W185–W192. [Google Scholar] [CrossRef] [PubMed]
- Eren, A.M.; Vineis, J.H.; Morrison, H.G.; Sogin, M.L. A Filtering Method to Generate High Quality Short Reads Using Illumina Paired-End Technology. PLoS ONE 2013, 8, e66643. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Eren, A.M.; Morrison, H.G.; Lescault, P.J.; Reveillaud, J.; Vineis, J.H.; Sogin, M.L. Minimum entropy decomposition: Unsupervised oligotyping for sensitive partitioning of high-throughput marker gene sequences. ISME J. 2014, 9, 968–979. [Google Scholar] [CrossRef]
- Chen, T.; Yu, W.-H.; Izard, J.; Baranova, O.V.; Lakshmanan, A.; Dewhirst, F.E. The Human Oral Microbiome Database: A web accessible resource for investigating oral microbe taxonomic and genomic information. Database 2010, 2010, baq013. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. Past: Paleontological statistics software package for educaton and data anlysis. Palaeontol. Electron. 2001, 4, 1. [Google Scholar]
- Cavalla, F.; Biguetti, C.C.; Melchiades, J.L.; Tabanez, A.P.; Azevedo, M.D.C.S.; Trombone, A.P.F.; Faveri, M.; Feres, M.; Garlet, G.P. Genetic Association with Subgingival Bacterial Colonization in Chronic Periodontitis. Genes 2018, 9, 271. [Google Scholar] [CrossRef] [PubMed]
- Rhodin, K.; Divaris, K.; North, K.; Barros, S.; Moss, K.; Beck, J.; Offenbacher, S. Chronic Periodontitis Genome-wide Association Studies. J. Dent. Res. 2014, 93, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Shusterman, A.; Durrant, C.; Mott, R.; Polak, D.; Schaefer, A.; Weiss, E.; Iraqi, F.; Houri-Haddad, Y. Host Susceptibility to Periodontitis. J. Dent. Res. 2013, 92, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Shusterman, A.; Munz, M.; Richter, G.; Jepsen, S.; Lieb, W.; Krone, B.; Hoffman, P.; Laudes, M.; Wellmann, J.; Berger, K.; et al. The PF4/PPBP/CXCL5 Gene Cluster Is Associated with Periodontitis. J. Dent. Res. 2017, 96, 945–952. [Google Scholar] [CrossRef]
- Munz, M.; Richter, G.M.; Loos, B.G.; Jepsen, S.; Divaris, K.; Offenbacher, S.; Teumer, A.; Holtfreter, B.; Kocher, T.; Bruckmann, C.; et al. Meta-analysis of genome-wide association studies of aggressive and chronic periodontitis identifies two novel risk loci. Eur. J. Hum. Genet. 2018, 27, 102–113. [Google Scholar] [CrossRef]
- Munz, M.; Willenborg, C.; Richter, G.M.; Jockel-Schneider, Y.; Graetz, C.; Staufenbiel, I.; Wellmann, J.; Berger, K.; Krone, B.; Hoffmann, P.; et al. A genome-wide association study identifies nucleotide variants at SIGLEC5 and DEFA1A3 as risk loci for periodontitis. Hum. Mol. Genet. 2017, 26, 2577–2588. [Google Scholar] [CrossRef]
- Divaris, K.; Monda, K.; North, K.; Olshan, A.; Lange, E.; Moss, K.; Barros, S.; Beck, J.; Offenbacher, S. Genome-wide Association Study of Periodontal Pathogen Colonization. J. Dent. Res. 2012, 91, S21–S28. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average | ||
| Age | 58.12 ± 9.89 | |
| BMI | 31.88 ± 4.37 | |
| Frequency | ||
| Gender | Male | 65 (63.1%) |
| Female | 48 (36.9%) | |
| Smoking status | Non smoker | 67 (65.0%) |
| Current smoker | 28 (27.2%) | |
| Former smoker | 8 (7.8%) | |
| Frequency of dental visits | Never been | 2 (1.9%) |
| Only in case of problems | 79 (76.7%) | |
| 1/year | 12 (11.7%) | |
| >1/year | 10 (9.7%) | |
| Previous periodontal treatment | Yes | 3 (2.9%) |
| No | 100 (97.1%) | |
| Last dental visit | Never been | 2 (1.9%) |
| >10 years ago | 7 (6.8%) | |
| 1–10 years ago | 60 (58.3%) | |
| <1 year ago | 34 (33.0%) | |
| Tooth-brushing frequency | <1/day | 6 (5.9%) |
| 1/day | 34 (33.0%) | |
| At least 2/day | 63 (61.1%) | |
| Type of toothbrush | None | 1 (1.0%) |
| Manual | 92 (89.3%) | |
| Electric | 10 (9.7%) | |
| Use of interdental cleaning tools | Daily–weekly | 14 (13.6%) |
| Never | 89 (86.4%) | |
| Average | Number | |
|---|---|---|
| Number of teeth (excluding third molars) | 22.77 ± 4.19 | 2345 |
| DMFT | 12.58 ± 6.07 | - |
| Decayed teeth | 0.63 ± 0.96 | 65 |
| Patients with caries detected | - | 38 (36.89%) |
| FMPS | 72.11 ± 22.51 | - |
| FMBS | 23.92 ± 19.62 | - |
| Average PPD | 2.44 ± 0.73 | - |
| Average CAL | 3.05 ± 1.12 | - |
| % PPDs 1–4 mm | 93.17 ± 0.88% | - |
| % PPDs 5–6 mm | 5.92 ± 0.75% | - |
| % of PPDs > 6 mm | 0.94 ± 0.23% | - |
| No–Mild periodontitis | - | 10 (9.7%) |
| Moderate periodontitis | - | 38 (36.9%) |
| Severe periodontitis | - | 55 (53.3%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nibali, L.; Stephen, A.S.; Allaker, R.P.; Di Pino, A.; Terranova, V.; Pisano, M.; Di Marca, S.; Ferrara, V.; Scicali, R.; Purrello, F.; et al. Associations between Host Genetic Variants and Subgingival Microbiota in Patients with the Metabolic Syndrome. Int. J. Mol. Sci. 2023, 24, 16649. https://doi.org/10.3390/ijms242316649
Nibali L, Stephen AS, Allaker RP, Di Pino A, Terranova V, Pisano M, Di Marca S, Ferrara V, Scicali R, Purrello F, et al. Associations between Host Genetic Variants and Subgingival Microbiota in Patients with the Metabolic Syndrome. International Journal of Molecular Sciences. 2023; 24(23):16649. https://doi.org/10.3390/ijms242316649
Chicago/Turabian StyleNibali, Luigi, Abish S. Stephen, Robert P. Allaker, Antonino Di Pino, Valentina Terranova, Marcella Pisano, Salvatore Di Marca, Viviana Ferrara, Roberto Scicali, Francesco Purrello, and et al. 2023. "Associations between Host Genetic Variants and Subgingival Microbiota in Patients with the Metabolic Syndrome" International Journal of Molecular Sciences 24, no. 23: 16649. https://doi.org/10.3390/ijms242316649