
Differences in the Occurrence of Cell Wall Components between Distinct Cell Types in Glands of Drosophyllum lusitanicum

 , , and
, , and
Abstract
:1. Introduction
2. Results
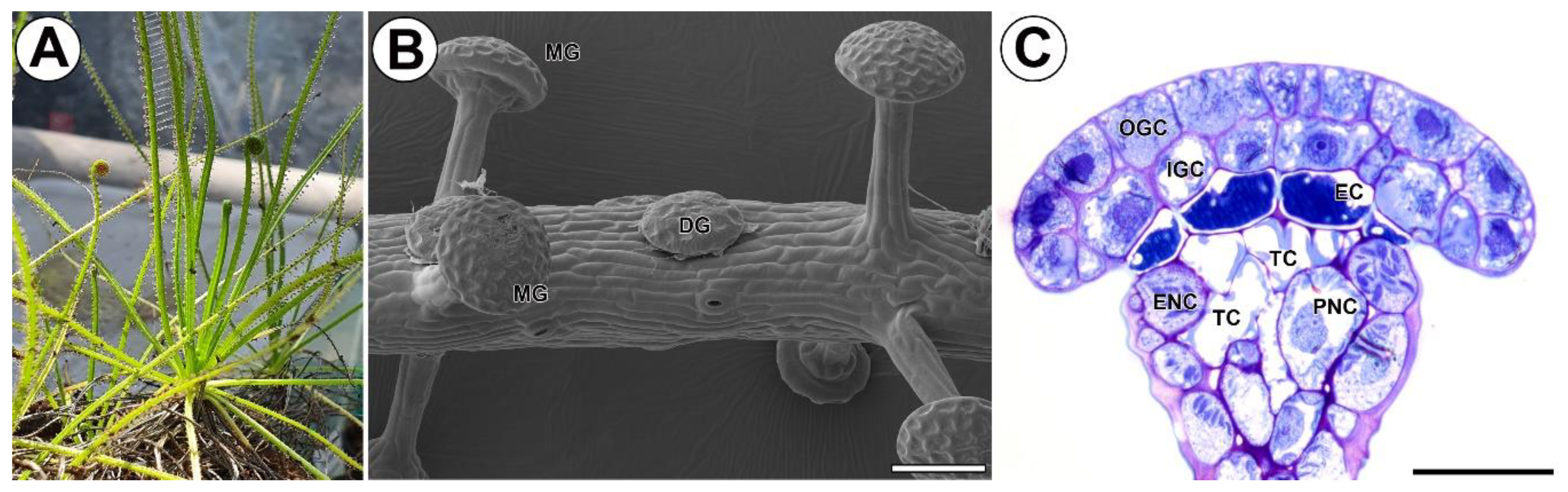
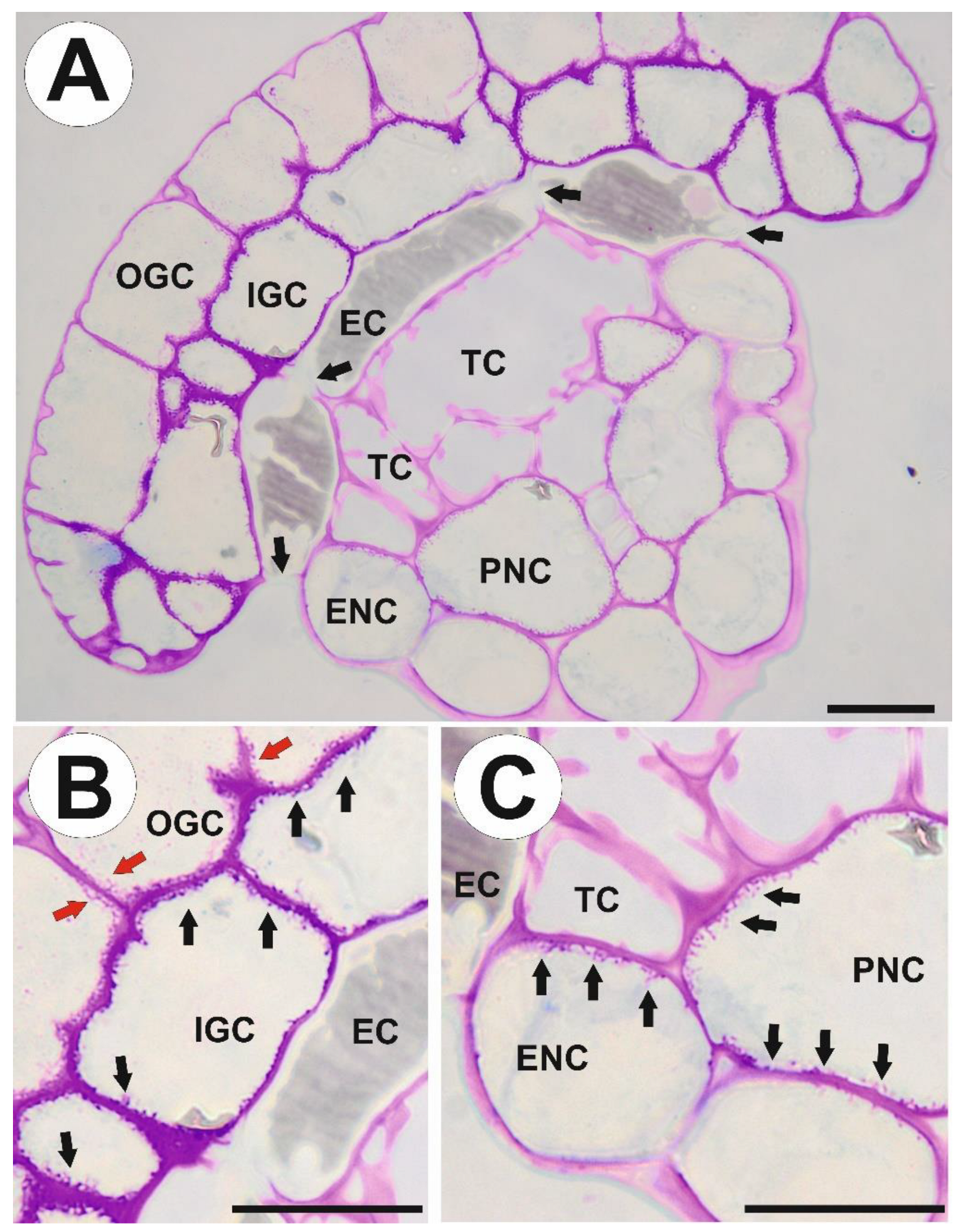
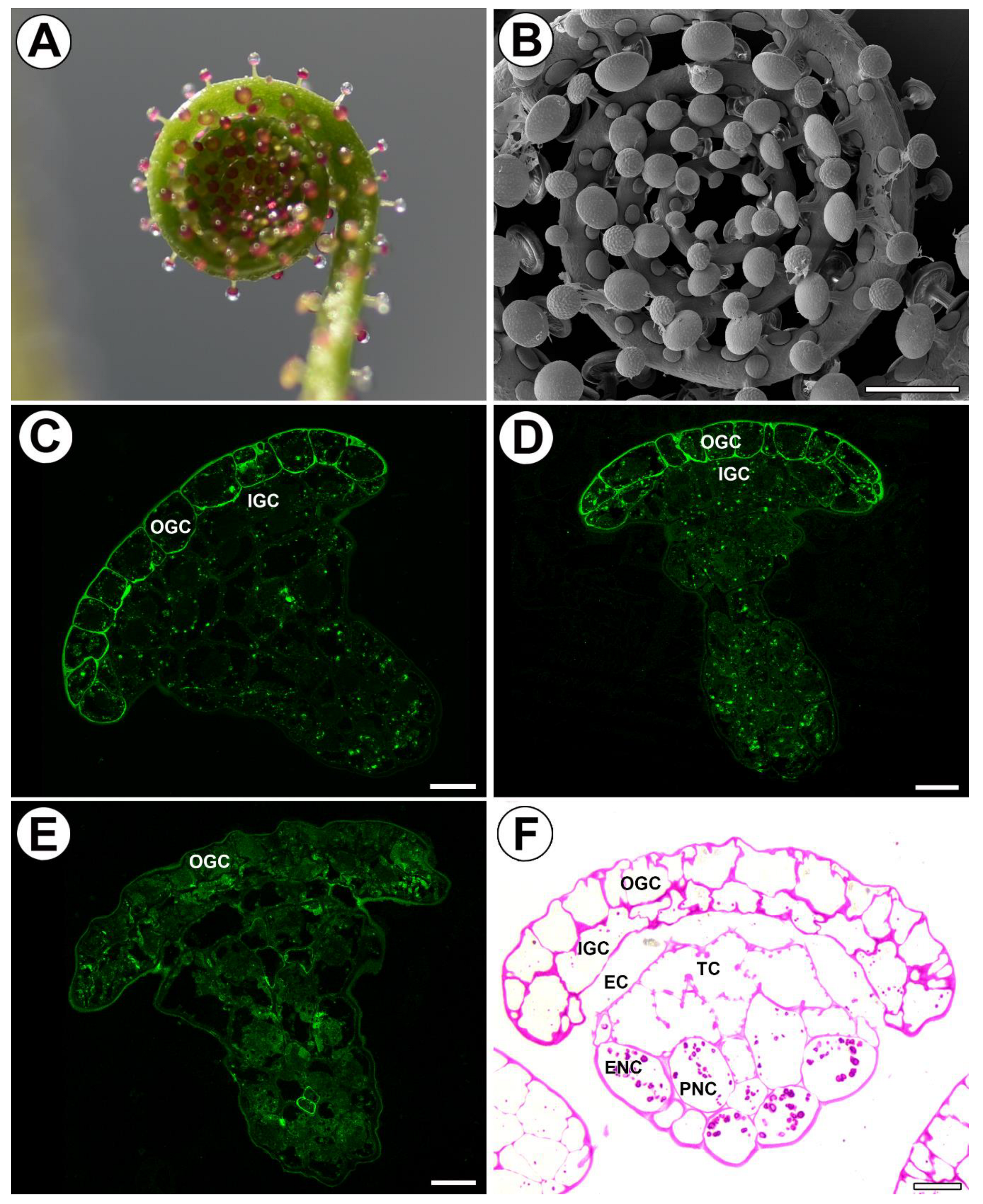
2.1. Structure of Stalked Mucilage Glands and Occurrence of Cell Wall Ingrowths
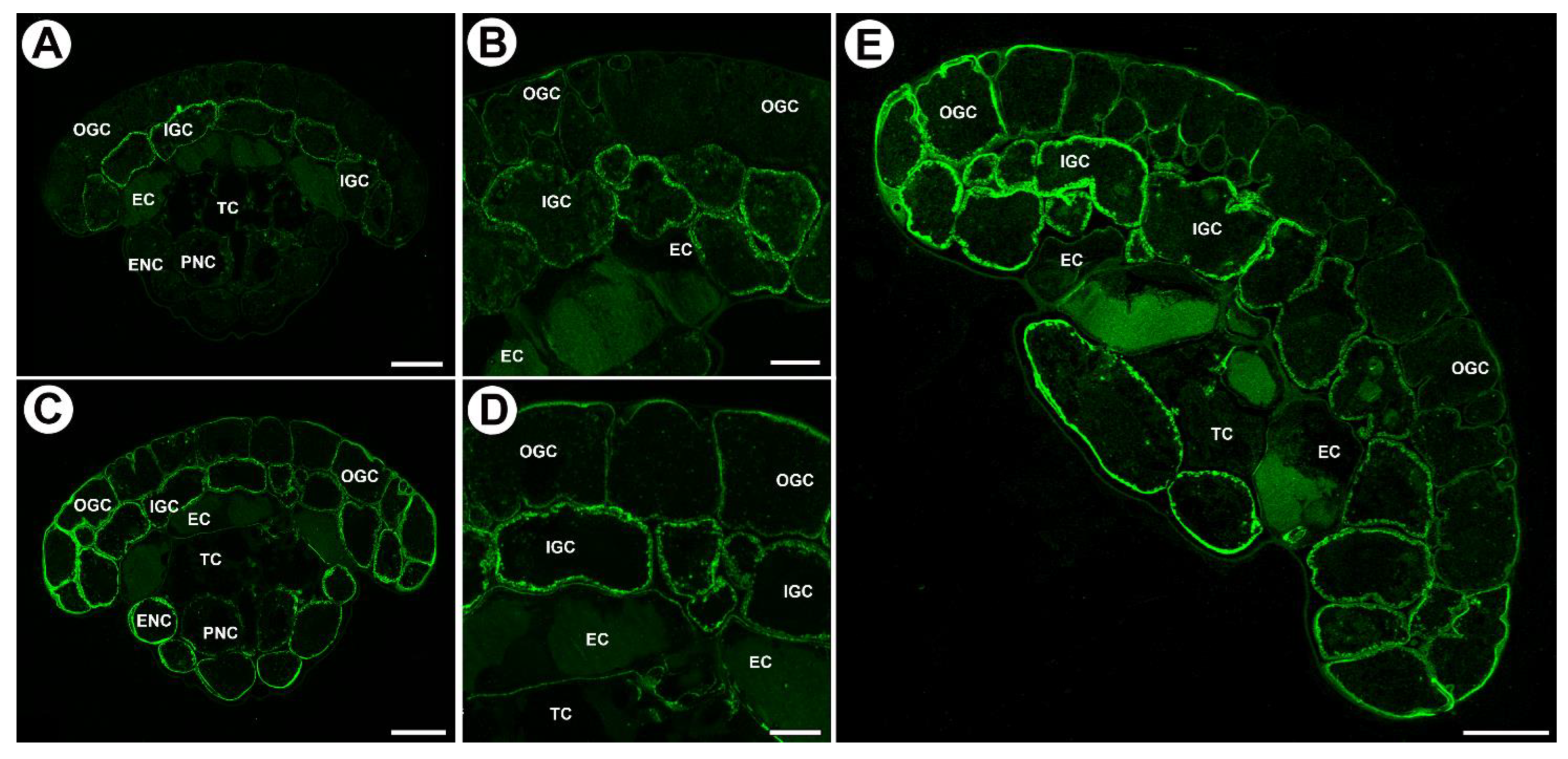
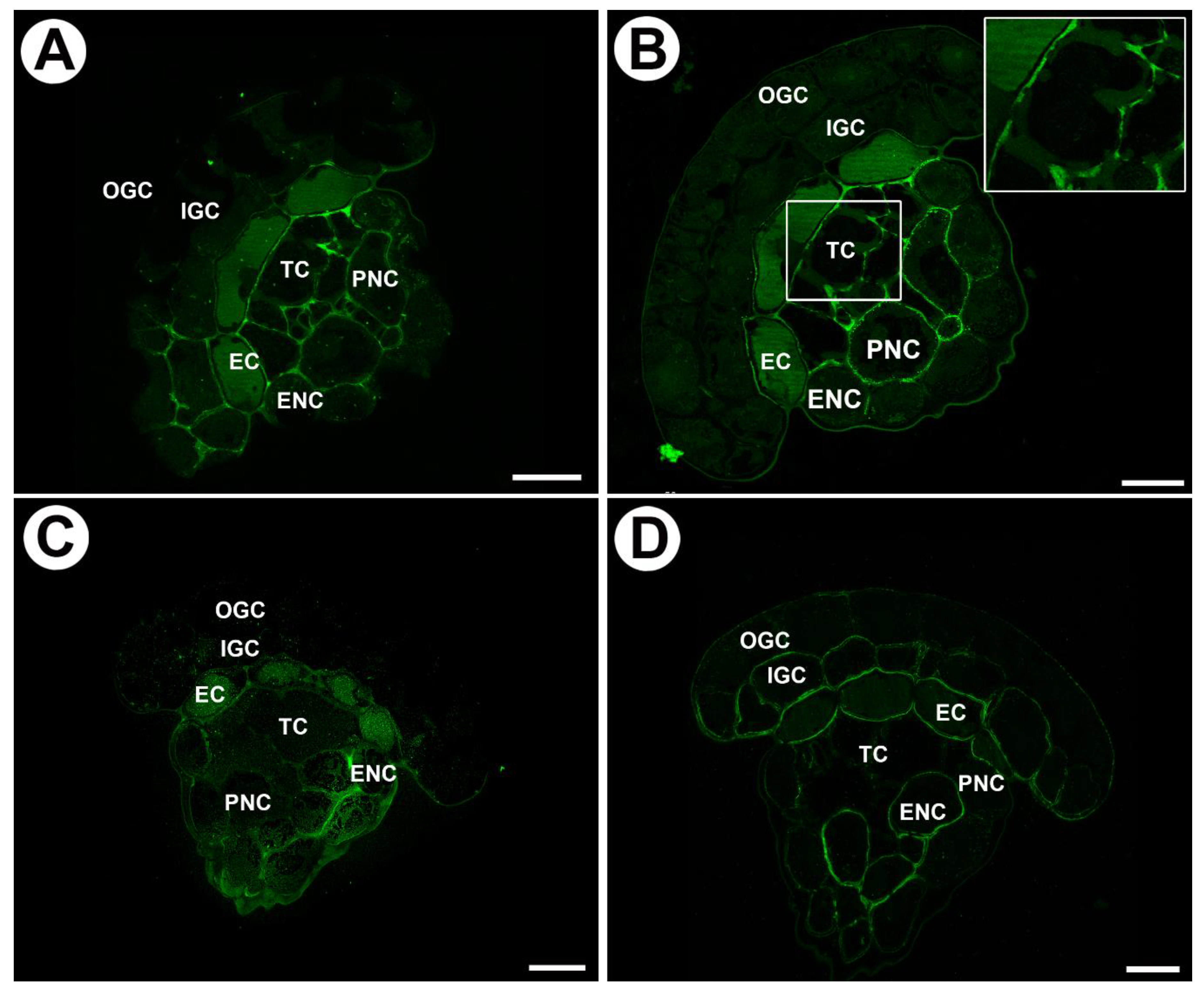
2.2. Distribution of Arabinogalactan Proteins (AGPs) in Mucilage Glands
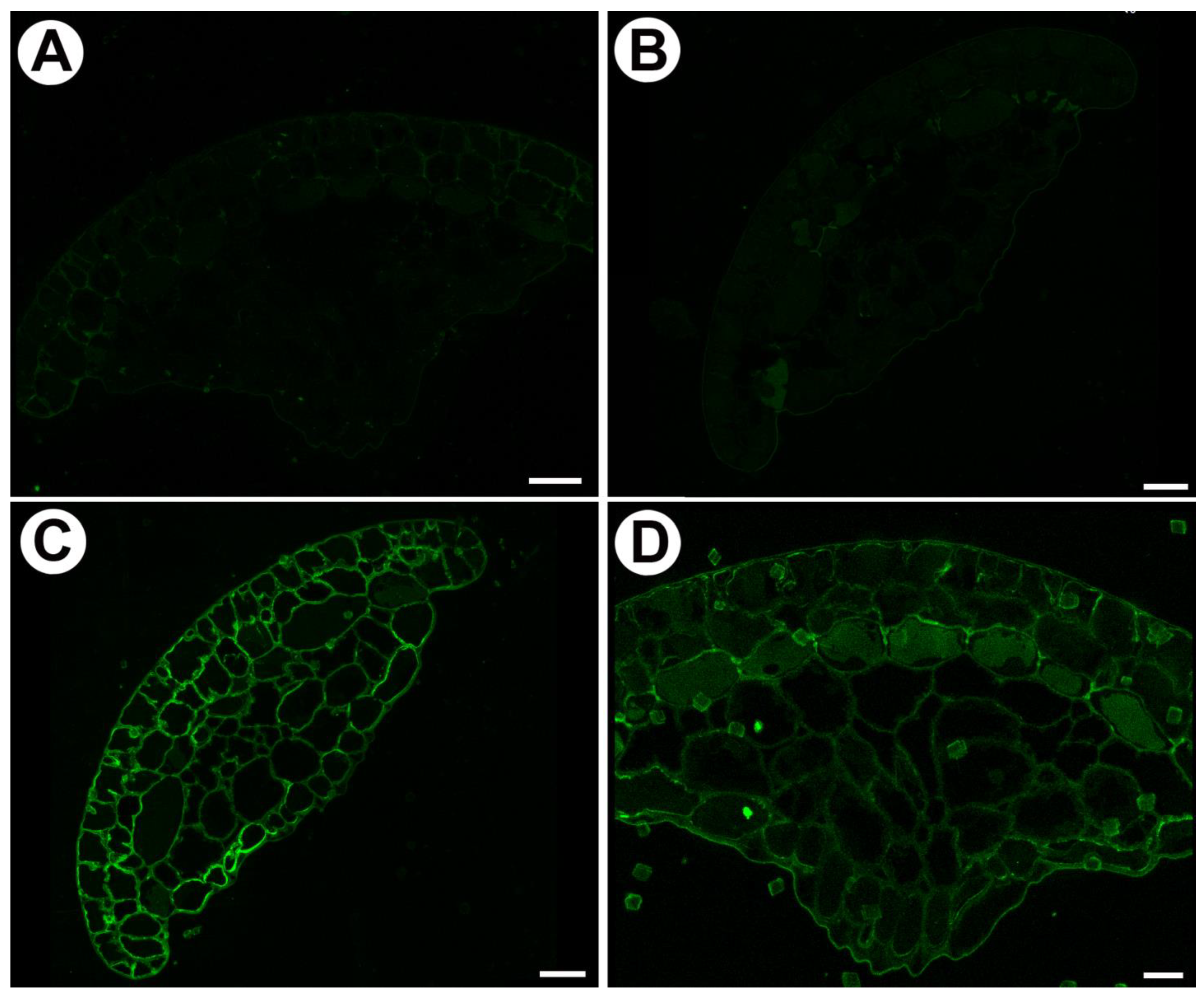
2.3. Distribution of Homogalacturonan in Mucilage Glands
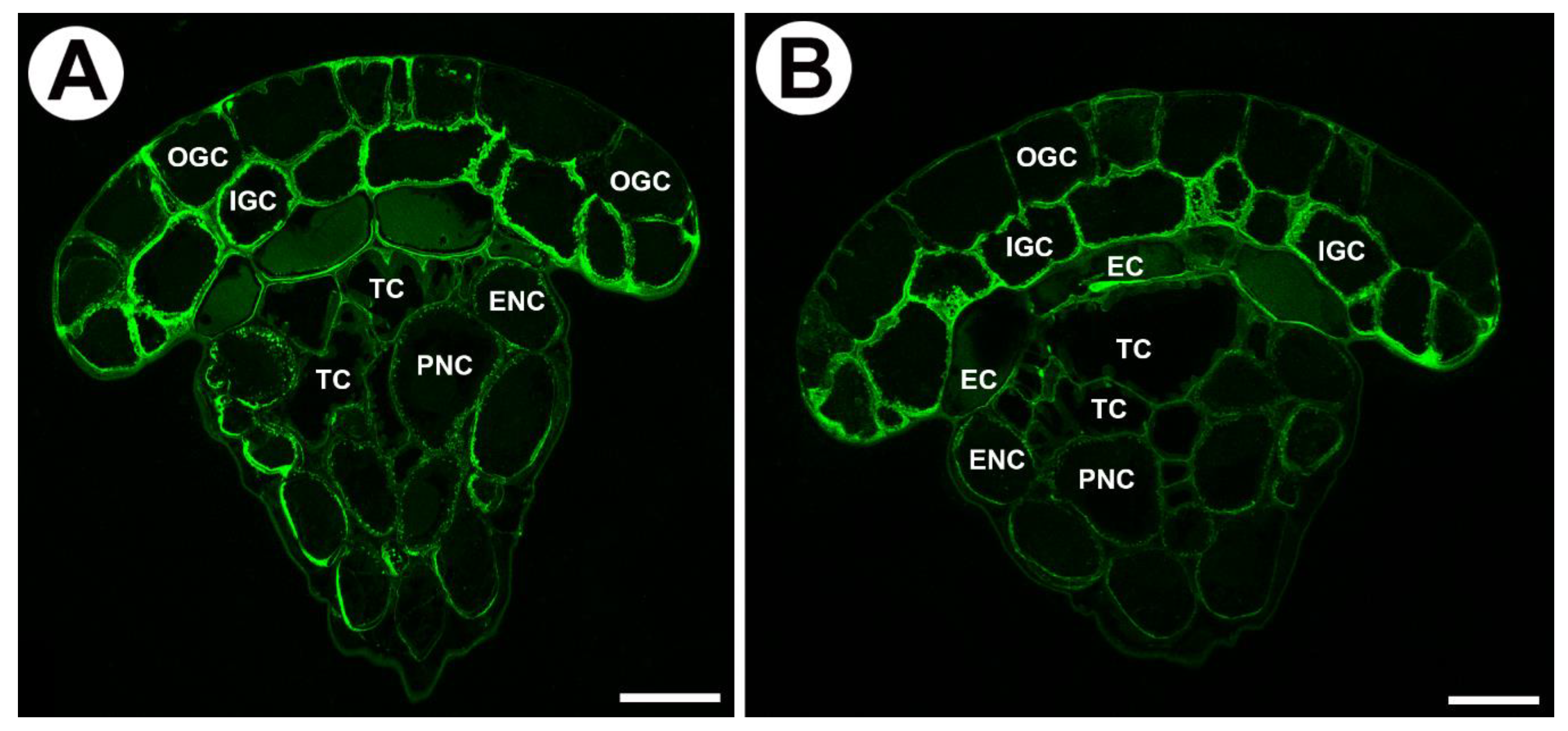
2.4. Distribution of Hemicellulose in Mucilage Glands
2.5. Enzymatic Treatments
3. Discussion
3.1. Cell Wall Polymers and Gland Cell Types
3.2. Cell Wall Polymers and Cell Wall Ingrowths
3.3. Cell Wall Polymers and Gland Activity
4. Materials and Methods
4.1. Plant Material
4.2. Histological and Immunochemical Analysis
4.3. Light Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Juniper, B.E.; Robbins, R.J.; Joel, D.M. The Carnivorous Plants; Academic Press: London, UK, 1989. [Google Scholar]
- Król, E.; Płachno, B.J.; Adamec, L.; Stolarz, M.; Dziubińska, H.; Trębacz, K. Quite a few reasons for calling carnivores ‘the most wonderful plants in the world’. Ann. Bot. 2012, 109, 47–64. [Google Scholar] [CrossRef] [PubMed]
- Ellison, A.M.; Adamec, L. Carnivorous Plants: Physiology, Ecology, and Evolution; Oxford University Press: Oxford, UK, 2018; 510p. [Google Scholar]
- Adlassnig, W.; Lendl, T.; Peroutka, M.; Lichtscheidl, I.K. “Insektenfressende Pflanzen”—Darwin und die Anfänge der Karnivorenforschung. In Darwin und die Botanik; Stöcklin, J., Höxtermann, E., Eds.; Basilisken-Presse: Rangsdorf, Germany, 2009; pp. 102–130. [Google Scholar]
- Gibson, T.C.; Waller, D.M. Evolving Darwin’s ‘most wonderful’ plant: Ecological steps to a snap-trap. New Phytol. 2009, 183, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Płachno, B.J. Rośliny zjadające zwierzęta a Karol Darwin—Opowieść o pięknej fascynacji/The carnivorous plants and Charles Darwin—A story about beautiful fascination. In Karol Darwin w Oczach Polskich Botaników XIX-XXI; Zarzycki, K., Mirek, Z., Korzeniak, U., Eds.; Instytut Botaniki im. W. Szafera, PAN IB: Kraków, Poland, 2010; pp. 59–75. [Google Scholar]
- Darwin, C. Insectivorous Plants, 1st ed.; John Murray: London, UK, 1875. [Google Scholar]
- Darwin Correspondence Project. Letter No. 2996. Available online: https://www.darwinproject.ac.uk/letter/?docId=letters/DCP-LETT-2996.xml (accessed on 27 February 2023).
- Fukushima, K.; Fang, X.; Alvarez-Ponce, D.; Cai, H.; Carretero-Paulet, L.; Chen, C.; Chang, T.H.; Farr, K.M.; Fujita, T.; Hiwatashi, Y.; et al. Genome of the pitcher plant Cephalotus reveals genetic changes associated with carnivory. Nat. Ecol. Evol. 2017, 6, 59. [Google Scholar] [CrossRef]
- Palfalvi, G.; Hackl, T.; Terhoeven, N.; Shibata, T.F.; Nishiyama, T.; Ankenbrand, M.; Becker, D.; Förster, F.; Freund, M.; Iosip, A.; et al. Genomes of the Venus Flytrap and Close Relatives Unveil the Roots of Plant Carnivory. Curr. Biol. 2020, 30, 12. [Google Scholar] [CrossRef]
- Adamec, L.; Matušíková, I.; Pavlovič, A. Recent ecophysiological, biochemical and evolutional insights into plant carnivory. Ann. Bot. 2021, 128, 241–259. [Google Scholar] [CrossRef] [PubMed]
- Durak, G.M.; Speck, T.; Poppinga, S. Shapeshifting in the Venus flytrap (Dionaea muscipula): Morphological and biomechanical adaptations and the potential costs of a failed hunting cycle. Front. Plant Sci. 2022, 2, 970320. [Google Scholar] [CrossRef]
- Nakamura, Y.; Reichelt, M.; Mayer, V.E.; Mithöfer, A. Jasmonates trigger prey-induced formation of ‘outer stomach’ in carnivorous sundew plants. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130228. [Google Scholar] [CrossRef] [PubMed]
- Pavlovič, A.; Mithöfer, A. Jasmonate signalling in carnivorous plants: Copycat of plant defence mechanisms. J. Exp. Bot. 2019, 70, 3379–3389. [Google Scholar] [CrossRef] [PubMed]
- Pavlovič, A.; Libiaková, M.; Bokor, B.; Jakšová, J.; Petřík, I.; Novák, O.; Baluška, F. Anaesthesia with diethyl ether impairs jasmonate signalling in the carnivorous plant Venus flytrap (Dionaea muscipula). Ann. Bot. 2020, 125, 173–183. [Google Scholar] [CrossRef]
- Hedrich, R.; Fukushima, K. On the Origin of Carnivory: Molecular physiology and evolution of plants on an animal diet. Annu. Rev. Plant Biol. 2021, 72, 133–153. [Google Scholar] [CrossRef]
- Fukushima, K.; Hasebe, M. Adaxial-abaxial polarity: The developmental basis of leaf shape diversity. Genesis 2014, 52, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Fukushima, K.; Fujita, H.; Yamaguchi, T.; Kawaguchi, M.; Tsukaya, H.; Hasebe, M. Oriented cell division shapes carnivorous pitcher leaves of Sarracenia purpurea. Nat. Commun. 2015, 6, 6450. [Google Scholar] [CrossRef] [PubMed]
- Whitewoods, C.D.; Gonçalves, B.; Cheng, J.; Cui, M.; Kennaway, R.; Lee, K.; Bushell, C.; Yu, M.; Piao, C.; Coen, E. Evolution of carnivorous traps from planar leaves through simple shifts in gene expression. Science 2020, 367, 91–96. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kozieradzka-Kiszkurno, M.; Świątek, P. Functional ultrastructure of Genlisea (Lentibulariaceae) digestive hairs. Ann. Bot. 2007, 100, 195–203. [Google Scholar] [CrossRef]
- Di Giusto, B.; Grosbois, V.; Fargeas, E.; Marshall, D.J.; Gaume, L. Contribution of pitcher fragrance and fluid viscosity to high prey diversity in a Nepenthes carnivorous plant from Borneo. J. Biosci. 2008, 33, 121–136. [Google Scholar] [CrossRef]
- Jürgens, A.; El-Sayed, A.M.; Suckling, D.M. Do carnivorous plants use volatiles for attracting prey insects? Funct. Ecol. 2009, 23, 875–887. [Google Scholar] [CrossRef]
- Kreuzwieser, J.; Scheerer, U.; Kruse, J.; Burzlaff, T.; Honsel, A.; Alfarraj, S.; Georgiev, P.; Schnitzler, J.P.; Ghirardo, A.; Kreuzer, I.; et al. The Venus flytrap attracts insects by the release of volatile organic compounds. J. Exp. Bot. 2014, 65, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Ojeda, F.; Carrera, C.; Paniw, M.; García-Moreno, L.; Barbero, G.F.; Palma, M. Volatile and semi-volatile organic compounds may help reduce pollinator-prey overlap in the carnivorous plant Drosophyllum lusitanicum (Drosophyllaceae). J. Chem. Ecol. 2021, 47, 73–86. [Google Scholar] [CrossRef]
- Adlassnig, W.; Lendl, T.; Peroutka, M.; Lang, I. Deadly glue—Adhesive traps of carnivorous plants. In Biological Adhesive Systems; Von Byern, J., Grunwald, I., Eds.; Springer: Vienna, NY, USA, 2010; pp. 15–28. [Google Scholar]
- Gaume, L.; Forterre, Y. A viscoelastic deadly fluid in carnivorous pitcher plants. PLoS ONE 2007, 2, e1185. [Google Scholar] [CrossRef] [PubMed]
- Kang, V.; Isermann, H.; Sharma, S.; Wilson, D.I.; Federle, W. How a sticky fluid facilitates prey retention in a carnivorous pitcher plant (Nepenthes rafflesiana). Acta Biomater. 2021, 128, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Adlassnig, W.; Koller-Peroutka, M.; Bauer, S.; Koshkin, E.; Lendl, T.; Lichtscheidl, I.K. Endocytotic uptake of nutrients in carnivorous plants. Plant J. 2012, 71, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Ivesic, C.; Krammer, S.; Koller-Peroutka, M.; Laarouchi, A.; Gruber, D.; Lang, I.; Lichtscheidl, I.K.; Adlassnig, W. Quantification of protein uptake by endocytosis in carnivorous Nepenthales. Plants 2023, 12, 341. [Google Scholar] [CrossRef] [PubMed]
- Freund, M.; Graus, D.; Fleischmann, A.; Gilbert, K.J.; Lin, Q.; Renner, T.; Stigloher, C.; Albert, V.A.; Hedrich, R.; Fukushima, K. The digestive systems of carnivorous plants. Plant Physiol. 2022, 190, 44–59. [Google Scholar] [CrossRef] [PubMed]
- Fineran, B.A.; Lee, M.S.L. Transfer cells in traps of the carnivorous plant Utricularia monanthos. J. Ultrastruct. Res. 1974, 48, 162–166. [Google Scholar] [CrossRef]
- Heslop-Harrison, Y. Enzyme release in carnivorous plants. Front. Biol. 1975, 43, 525–578. [Google Scholar]
- Gunning, B.E.S.; Pate, J.S. Transfer cells. In Dynamic Aspects of Plant Ultrastructure; Robards, A.W., Ed.; McGraw Hill: London, UK; New York, NY, USA, 1974; pp. 441–480. [Google Scholar]
- Offler, C.E.; Patrick, J.W. Transfer cells: What regulates the development of their intricate wall labyrinths? New Phytol. 2020, 228, 427–444. [Google Scholar] [CrossRef]
- Fleischmann, A.; Schlauer, J.; Smith, S.A.; Givnish, T.J. Evolution of Carnivory in Angiosperms; Ellison, A.M., Adamec, L., Eds.; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Lloyd, F.E. The Carnivorous Plants; Chronica Botanica Company: Waltham, MA, USA, 1942. [Google Scholar]
- Adlassnig, W.; Peroutka, M.; Eder, G.; Pois, W.; Lichtscheidl, I.K. Ecophysiological observations on Drosophyllum lusitanicum. Ecol. Res. 2006, 21, 255–262. [Google Scholar] [CrossRef]
- Paniw, M.; Gil-Cabeza, E.; Ojeda, F. Plant carnivory beyond bogs: Reliance on prey feeding in Drosophyllum lusitanicum (Drosophyllaceae) in dry Mediterranean heathland habitats. Ann. Bot. 2017, 119, 1035–1041. [Google Scholar] [CrossRef]
- Bertol, N.; Paniw, M.; Ojeda, F. Effective prey attraction in the rare Drosophyllum lusitanicum, a flypaper-trap carnivorous plant. Am. J. Bot. 2015, 102, 689–694. [Google Scholar] [CrossRef]
- Fenner, C.A. Beiträge zur Kenntnis der Anatomie, Entwicklungsgeschichte und Biologie der Laubblätter und Drüsen Einiger Insektivoren; Universität Zürich: Zürich, Switzerland, 1904; p. 104. [Google Scholar]
- Schnepf, E. Zur Feinstruktur der Drüsen von Drosophyllum lusitanicum. Planta 1960, 54, 641–674. [Google Scholar] [CrossRef]
- Schnepf, E. Licht- und elektronenmikroskopische Beobachtungen an Insektivoren Drüsen über die Sekretion des Fangschleimes. Flora 1961, 151, 73–87. [Google Scholar]
- Green, S.; Green, T.L.; Heslop-Harrison, Y. Seasonal heterophylly and leaf gland features in Triphyophyllum (Dioncophyllaceae), a new carnivorous plant genus. Bot. J. Linn. Soc. 1979, 78, 99–116. [Google Scholar] [CrossRef]
- Goebel, K. Pflanzenbiologische Schilderungen; N.G. Elwert: Marburg, Germany, 1889. [Google Scholar]
- Adamec, L. Ecophysiological investigation on Drosophyllum lusitanicum: Why doesn’t the plant dry out? Carniv. Plant Newsl. 2009, 38, 71–74. [Google Scholar] [CrossRef]
- Vassilyev, A.E. Dynamics of ultrastructural characters of Drosophyllum lusitanicum Link (Droseraceae) digestive glands during maturation and after stimulation. Taiwania 2005, 50, 167–182. [Google Scholar]
- Baluška, F.; Hlavacka, A.; Šamaj, J.; Palme, K.; Robinson, D.G.; Matoh, T.; McCurdy, D.W.; Menzel, D.; Volkmann, D. F-actin-dependent endocytosis of cell wall pectins in meristematic root cells. Insights from brefeldin A-induced compartments. Plant Physiol. 2002, 130, 422–431. [Google Scholar] [CrossRef]
- Baluška, F.; Baroja-Fernandez, E.; Pozueta-Romero, J.; Hlavacka, A.; Etxeberria, E.; Šamaj, J. Endocytic Uptake of Nutrients, Cell Wall Molecules and Fluidized Cell Wall Portions into Heterotrophic Plant Cells. In Plant Endocytosis. Plant Cell Monographs; Šamaj, J., Baluška, F., Menzel, D., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; Volume 1. [Google Scholar]
- Baluška, F.; Liners, F.; Hlavačka, A.; Schlicht, M.; Van Cutsem, P.; McCurdy, D.W.; Menzel, D. Cell wall pectins and xyloglucans are internalized into dividing root cells and accumulate within cell plates during cytokinesis. Protoplasma 2005, 225, 141–155. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P.; Strzemski, M.; Miranda, V.F.O. Immunocytochemical analysis of the wall ingrowths in the digestive gland transfer cells in Aldrovanda vesiculosa L. (Droseraceae). Cells 2022, 11, 2218. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P. Arabinogalactan proteins in the digestive glands of Dionaea muscipula J. Ellis Traps. Cells 2022, 11, 586. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P. Stellate trichomes in Dionaea muscipula Ellis (Venus Flytrap) Traps, Structure and Functions. Int. J. Mol. Sci. 2023, 24, 553. [Google Scholar] [CrossRef]
- Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Wójciak, M.; Świątek, P. Immunocytochemical analysis of bifid trichomes in Aldrovanda vesiculosa L. Traps. Int. J. Mol. Sci. 2023, 24, 3358. [Google Scholar] [CrossRef]
- Offler, C.E.; McCurdy, D.W.; Patrick, J.W.; Talbot, M.J. Transfer cells: Cells specialized for a special purpose. Annu. Rev. Plant Biol. 2003, 54, 431–454. [Google Scholar] [CrossRef]
- Vaughn, K.C.; Talbot, M.J.; Offler, C.E.; McCurdy, D.W. Wall ingrowths in epidermal transfer cells of Vicia faba cotyledons are modified primary walls marked by localized accumulations of arabinogalactan proteins. Plant Cell Physiol. 2006, 48, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Ligrone, R.; Vaughn, K.C.; Rascio, N. A cytochemical and immunocytochemical analysis of the wall labyrinth apparatus in leaf transfer cells in Elodea canadensis. Ann. Bot. 2011, 107, 717–722. [Google Scholar] [CrossRef] [PubMed]
- Henry, J.S.; Lopez, R.A.; Renzaglia, K.S. Differential localization of cell wall polymers across generations in the placenta of Marchantia polymorpha. J. Plant Res. 2020, 133, 911–924. [Google Scholar] [CrossRef]
- Henry, J.S.; Ligrone, R.; Vaughn, K.C.; Lopez, R.A.; Renzaglia, K.S. Cell wall polymers in the Phaeoceros placenta reflect developmental and functional differences across generations. Bryophyt. Divers. Evol. 2021, 43, 265–283. [Google Scholar] [CrossRef]
- Henry, J.S.; Renzaglia, K.S. The placenta of Physcomitrium patens: Transfer cell wall polymers compared across the three Bryophyte groups. Diversity 2021, 13, 378. [Google Scholar] [CrossRef]
- Dahiya, P.; Brewin, N.J. Immunogold localization of callose and other cell wall components in pea nodule transfer cells. Protoplasma 2000, 214, 210–218. [Google Scholar] [CrossRef]
- Showalter, A. Arabinogalactan-proteins: Structure, expression and function. Cell. Mol. Life Sci. 2001, 58, 1399–1417. [Google Scholar] [CrossRef]
- Showalter, A.M.; Basu, D. Glycosylation of arabinogalactan-proteins essential for development in Arabidopsis. Commun. Integr. Biol. 2016, 9, e0125624. [Google Scholar] [CrossRef]
- Rancour, D.M.; Marita, J.M.; Hatfield, R.D. Cell wall composition throughout development for the model grass Brachypodium distachyon. Front. Plant Sci. 2012, 3, 266. [Google Scholar] [CrossRef]
- Leszczuk, A.; Cybulska, J.; Skrzypek, T.; Zdunek, A. Properties of arabinogalactan proteins (AGPs) in apple (Malus × domestica) fruit at different stages of ripening. Biology 2020, 9, 225. [Google Scholar] [CrossRef]
- Leszczuk, A.; Kalaitzis, P.; Blazakis, K.N.; Zdunek, A. The role of arabinogalactan proteins (AGPs) in fruit ripening—A review. Hortic. Res. 2020, 7, 176. [Google Scholar] [CrossRef]
- Hromadová, D.; Soukup, A.; Tylová, E. Arabinogalactan proteins in plant roots—An update on possible functions. Front. Plant Sci. 2021, 12, 674010. [Google Scholar] [CrossRef]
- Jia, K.; Wang, W.; Zhang, Q.; Jia, W. Cell wall integrity signaling in fruit ripening. Int. J. Mol. Sci. 2023, 24, 4054. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Yan, Y.; Takác, T.; Li, X.; Chen, H.; Wang, Y.; Xu, E.; Xie, L.; Su, Z.; Šamaj, J.; Xu, C. Variable content and distribution of arabinogalactan proteins in banana (Musa spp.) under low temperature stress. Front. Plant Sci. 2015, 6, 353. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Bulgakov, V.P.; Jinn, T.L. Pectin methylesterases: Cell wall remodeling proteins are required for plant response to heat stress. Front. Plant Sci. 2018, 9, 1612. [Google Scholar] [CrossRef]
- Pinski, A.; Betekhtin, A.; Skupien-Rabian, B.; Jankowska, U.; Jamet, E.; Hasterok, R. Changes in the cell wall proteome of leaves in response to high temperature stress in Brachypodium distachyon. Int. J. Mol. Sci. 2021, 22, 6750. [Google Scholar] [CrossRef]
- Leszczuk, A.; Szczuka, E.; Lewtak, K.; Chudzik, B.; Zdunek, A. Effect of low temperature on changes in AGP distribution during development of Bellis perennis ovules and anthers. Cells 2021, 10, 1880. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S. Cell wall signaling in plant development and defense. Annu. Rev. Plant Biol. 2022, 73, 323–353. [Google Scholar] [CrossRef] [PubMed]
- Escalante-Perez, M.; Krol, E.; Stange, A.; Geiger, D.; Al-Rasheid, K.A.; Hause, B.; Neher, E.; Hedrich, R. A special pair of phytohormones controls excitability, slow closure, and external stomach formation in the Venus flytrap. Proc. Natl. Acad. Sci. USA 2011, 108, 15492–15497. [Google Scholar] [CrossRef]
- Schulze, W.X.; Sanggaard, K.W.; Kreuzer, I.; Knudsen, A.D.; Bemm, F.; Thøgersen, I.B.; Bräutigam, A.; Thomsen, L.R.; Schliesky, S.; Dyrlund, T.F.; et al. The protein composition of the digestive fluid from the Venus Flytrap sheds light on prey digestion mechanisms. Mol. Cell. Proteom. 2012, 1, 1306–1319. [Google Scholar] [CrossRef]
- Bemm, F.; Becker, D.; Larisch, C.; Kreuzer, I.; Escalante-Perez, M.; Schulze, W.X.; Ankenbrand, M.; Van de Weyer, A.L.; Krol, E.; Al-Rasheid, K.A.; et al. Venus flytrap carnivorous lifestyle builds on herbivore defense strategies. Genome Res. 2016, 26, 812–825. [Google Scholar] [CrossRef] [PubMed]
- Outenreath, R.; Dauwalder, M. Ultrastructural and radioautographic studies of the digestive gland cells of Drosera capensis: I. Development and mucilage secretion. J. Ultrastruct. Res. 1982, 80, 71–88. [Google Scholar] [CrossRef]
- Muravnik, L.E. The ultrastructure of the secretory cells of glandular hairs in two Drosera species as affected by chemical stimulation. Russ. J. Plant Physiol. 2000, 47, 540–548. [Google Scholar]
- Schwab, D.W.; Simmons, E.; Scala, J. Fine structure changes during function of the digestive gland of Venus’s-Flytrap. Am. J. Bot. 1969, 56, 88–100. [Google Scholar] [CrossRef]
- Fineran, B.A. Glandular trichomes in Utricularia: A review of their structure and function. Isr. J. Bot. 1985, 34, 295–330. [Google Scholar]
- Samaj, J.; Samajova, O.; Baluska, E.; Lichtscheidl, I.; Knox, J.P.; Volkmann, D. Immunolocalization of LM2 arabinogalactan protein epitope associated with endomembranes of plant cells. Protoplasma 2000, 212, 186–196. [Google Scholar] [CrossRef]
- Olmos, E.; García De La Garma, J.; Gomez-Jimenez, M.C.; Fernandez-Garcia, N. Arabinogalactan proteins are involved in salt-adaptation and vesicle trafficking in Tobacco BY-2 cell cultures. Front. Plant Sci. 2017, 8, 1092. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cheng, M.; Yang, Q.; Li, J.; Wang, X.; Zhou, Q.; Nagawa, S.; Xia, B.; Xu, T.; Huang, R.; et al. Arabinogalactan protein-rare earth element complexes activate plant endocytosis. Proc. Natl. Acad. Sci. USA 2019, 16, 14349–14357. [Google Scholar] [CrossRef]
- Akerman, A. Untersuchungen über die Aggregation in den Tentakeln von Drosera rotundifolia. Bot. Notiser 1917, 145, 192. [Google Scholar]
- Scherzer, S.; Shabala, L.; Hedrich, B.; Fromm, J.; Bauer, H.; Munz, E.; Jakob, P.; Al-Rascheid, K.A.S.; Kreuzer, I.; Becker, D.; et al. Insect haptoelectrical stimulation of Venus flytrap triggers exocytosis in gland cells. Proc. Natl. Acad. Sci. USA 2017, 114, 4822–4827. [Google Scholar] [CrossRef] [PubMed]
- Lichtscheidl, I.K.; Lancelle, S.A.; Hepler, P.K. Actin-endoplasmic reticulum complexes in Drosera—Their structural relationship with the plasmalemma, nucleus, and organelles in cells prepared by high pressure freezing. Protoplasma 1990, 155, 116–126. [Google Scholar] [CrossRef]
- Gergely, Z.R.; Martinez, D.E.; Donohoe, B.S.; Mogelsvang, S.; Herder, R.; Staehelin, L.A. 3D electron tomographic and biochemical analysis of ER, Golgi and trans Golgi network membrane systems in stimulated Venus flytrap (Dionaea muscipula) glandular cells. J. Biol. Res. 2018, 25, 15. [Google Scholar] [CrossRef] [PubMed]
- Boulogne, C.; Gillet, C.; Hughes, L.; LE Bars, R.; Canette, A.; Hawes, C.R.; Satiat-Jeunemaitre, B. Functional organisation of the endomembrane network in the digestive gland of the Venus flytrap: Revisiting an old story with a new microscopy toolbox. J. Microsc. 2020, 280, 86–103. [Google Scholar] [CrossRef] [PubMed]
- Lichtscheidl, I.; Lancelle, S.; Weidinger, M.; Adlassnig, W.; Koller-Peroutka, M.; Bauer, S.; Krammer, S.; Hepler, P.K. Gland cell responses to feeding in Drosera capensis, a carnivorous plant. Protoplasma 2021, 258, 1291–1306. [Google Scholar] [CrossRef]
- Dexheimer, J. Quelques aspects ultrastructuraux de la secretion de mucilage par les glandes digestives de Drosera rotundifolia L. Acad. Sci. Paris CR Ser. D 1972, 275, 1983–1986. [Google Scholar]
- Dexheimer, J. Etude de la secretion de mucilage par les cellules des glandes digestives de Drosera (D. rotundifolia L. et D. capensis L.) Application de quelques techniques cytochimiques. Cytobiologie 1976, 13, 307–321. [Google Scholar] [CrossRef]
- Dexheimer, J. Study of mucilage secretion by the cells of the digestive glands of Drosera capensis L. using staining of the plasmalemma and mucilage by Phosphotungstic Acid. Cytologia 1978, 43, 45–52. [Google Scholar] [CrossRef]
- Schnepf, E. Zur Cytologie und Physiologie pflanzlicher Drüsen: 1. Teil. Über den Fangschleim der Insectivoren. Flora 1963, 153, 1–22. [Google Scholar]
- McCurdy, D.W.; Patrick, J.W.; Offler, C.E. Wall ingrowth formation in transfer cells: Novel examples of localized wall deposition in plant cells. Curr. Opin. Plant Biol. 2008, 11, 653–661. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.P.; Day, S.; Roberts, K. A set of cell surface glycoproteins forms an early position, but not cell type, in the root apical meristem of Daucus carota L. Development 1989, 106, 47–56. [Google Scholar] [CrossRef]
- Pennell, R.I.; Knox, J.P.; Scofield, G.N.; Selvendran, R.R.; Roberts, K. A family of abundant plasma membrane-associated glycoproteins related to the arabinogalactan proteins is unique to flowering plants. J. Cell Biol. 1989, 108, 1967–1977. [Google Scholar] [CrossRef]
- Pennell, R.I.; Janniche, L.; Kjellbom, P.; Scofield, G.N.; Peart, J.M.; Roberts, K. Developmental regulation of a plasma membrane arabinogalactan protein epitope in oilseed rape flowers. Plant Cell 1991, 3, 1317–1326. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.P.; Linstead, P.J.; Cooper, J.P.C.; Roberts, K. Developmentally regulated epitopes of cell surface arabinogalactan proteins and their relation to root tissue pattern formation. Plant J. 1991, 1, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Marcus, S.E.; Verhertbruggen, Y.; Hervé, C.; Ordaz-Ortiz, J.J.; Farkas, V.; Pedersen, H.L.; Willats, W.G.; Knox, J.P. Pectic homogalacturonan masks abundant sets of xyloglucan epitopes in plant cell walls. BMC Plant Biol. 2008, 8, 60. [Google Scholar] [CrossRef]
- Verhertbruggen, Y.; Marcus, S.E.; Haeger, A.; Ordaz-Ortiz, J.J.; Knox, J.P. An extended set of monoclonal antibodies to pectic homogalacturonan. Carbohydr. Res. 2009, 344, 1858–1862. [Google Scholar] [CrossRef]
- McCartney, L.; Marcus, S.E.; Knox, J.P. Monoclonal antibodies to plant cell wall xylans and arabinoxylans. J. Histochem. Cytochem. 2005, 53, 543–546. [Google Scholar] [CrossRef]
- Humphrey, C.; Pittman, G. A simple methylene blue-azure II-basic fuchsin stain for epoxy-embedded tissue sections. Stain. Technol. 1974, 49, 9–14. [Google Scholar] [CrossRef]
- Wędzony, M. Fluorescence Microscopy for Botanists; Department of Plant Physiology Monographs 5: Kraków, Poland, 1996. (In Polish) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Wall Polysaccharides and Glycoproteins | Gland Cell Types | |||||
|---|---|---|---|---|---|---|
| Cell Walls of Outer Glandular Cell | Cell Walls of Inner Glandular Cell | Cell Walls of Endodermoid Cell | Cell Walls of Tracheid | Cell Walls of Epidermal Neck Cell | Cell Walls of Parenchymal Neck Cell | |
| Arabinogalactan proteins | Occurred especially in cell wall ingrowths; in case of JIM14, weaker signal in cell walls | Occurred especially in cell wall ingrowths | Lack (JIM14); occurred (JIM8, JIM13) | Occurred (JIM14); lack (JIM8, JIM13) | Occurred especially in cell wall ingrowths (LM19) | Occurred especially in cell wall ingrowths (LM19) |
| Low methyl esterified homogalacturonans | Lack or weak signal | Lack or weak signal | Occurred | Occurred | Occurred | Occurred |
| Highly esterified homogalacturonans | Lack | Lack or weak signal | Occurred | Occurred | Occurred | Occurred |
| Galactan | Weak signal | Occurred | Occurred | Lack | Weak signal | Occurred |
| Xyloglucan | Occurred; strong signal in cell wall ingrowths | Occurred; strong signal in cell wall ingrowths | Occurred | Occurred | Occurred | Occurred |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Płachno, B.J.; Kapusta, M.; Stolarczyk, P.; Świątek, P.; Lichtscheidl, I. Differences in the Occurrence of Cell Wall Components between Distinct Cell Types in Glands of Drosophyllum lusitanicum. Int. J. Mol. Sci. 2023, 24, 15045. https://doi.org/10.3390/ijms242015045
Płachno BJ, Kapusta M, Stolarczyk P, Świątek P, Lichtscheidl I. Differences in the Occurrence of Cell Wall Components between Distinct Cell Types in Glands of Drosophyllum lusitanicum. International Journal of Molecular Sciences. 2023; 24(20):15045. https://doi.org/10.3390/ijms242015045
Chicago/Turabian StylePłachno, Bartosz J., Małgorzata Kapusta, Piotr Stolarczyk, Piotr Świątek, and Irene Lichtscheidl. 2023. "Differences in the Occurrence of Cell Wall Components between Distinct Cell Types in Glands of Drosophyllum lusitanicum" International Journal of Molecular Sciences 24, no. 20: 15045. https://doi.org/10.3390/ijms242015045