
Genome-Wide Survey of the RWP-RK Gene Family in Cassava (Manihot esculenta Crantz) and Functional Analysis
 , ,
, ,
Abstract
:1. Introduction
2. Results
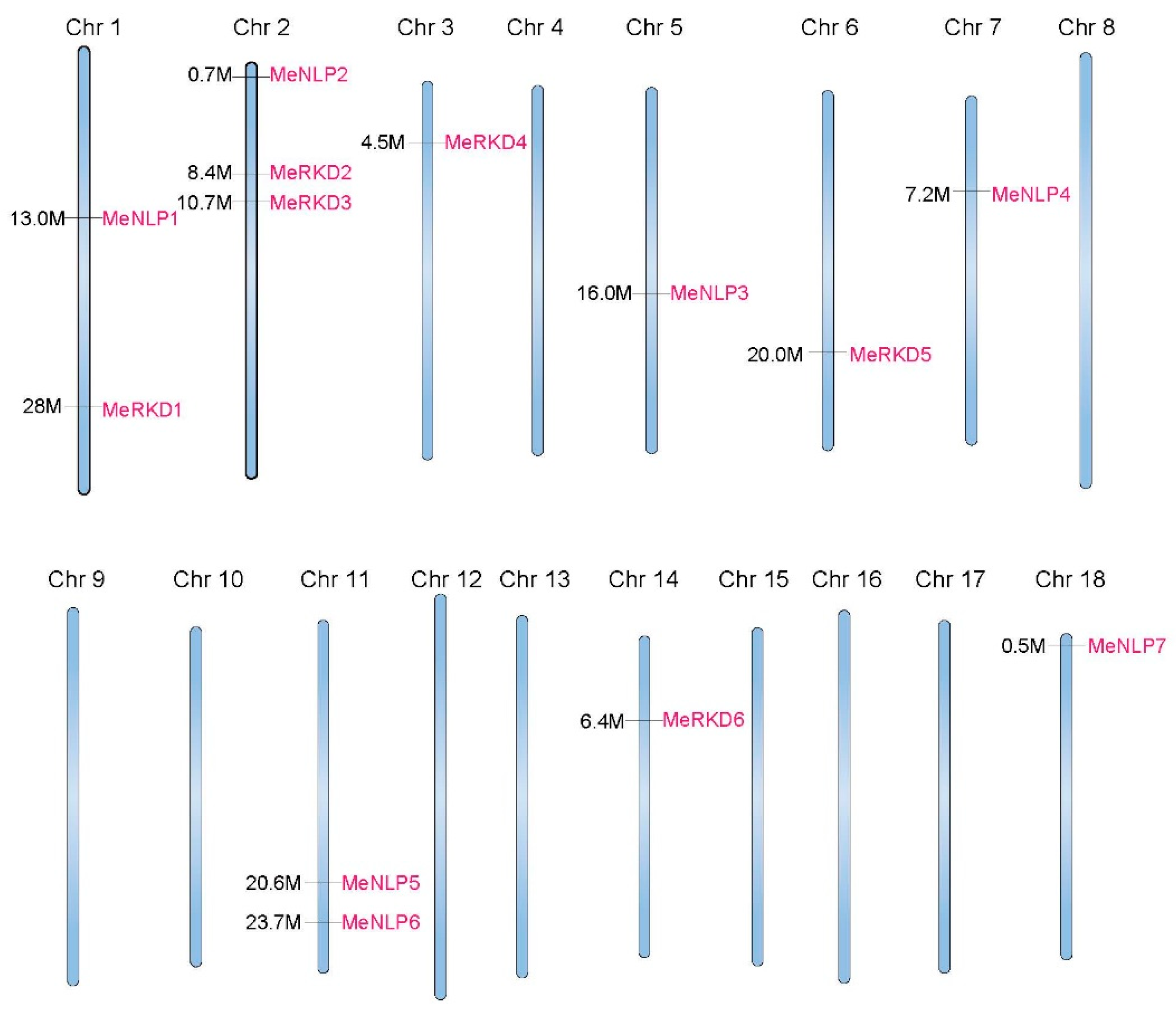
2.1. Cassava RWP-RK Protein Family
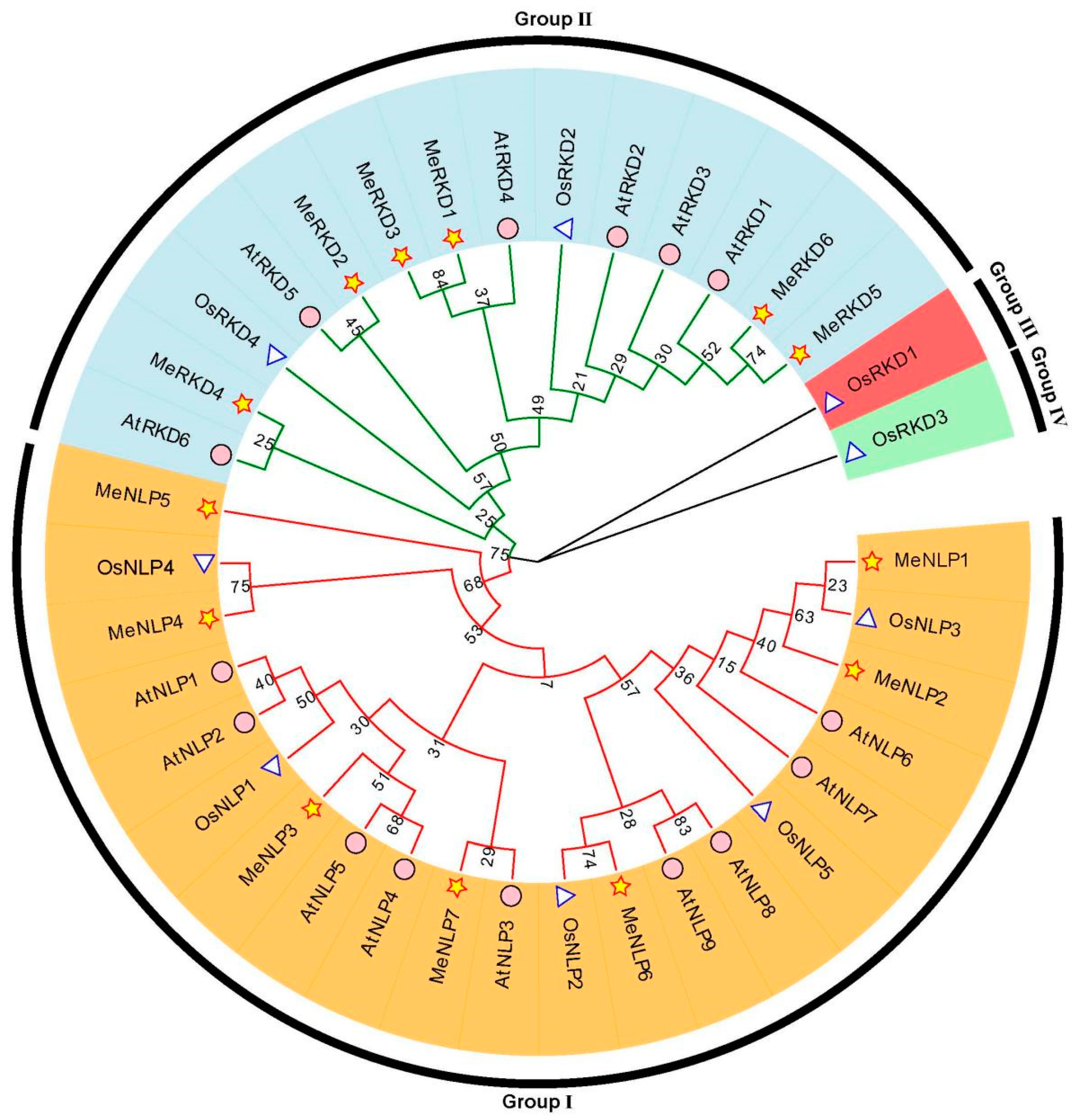
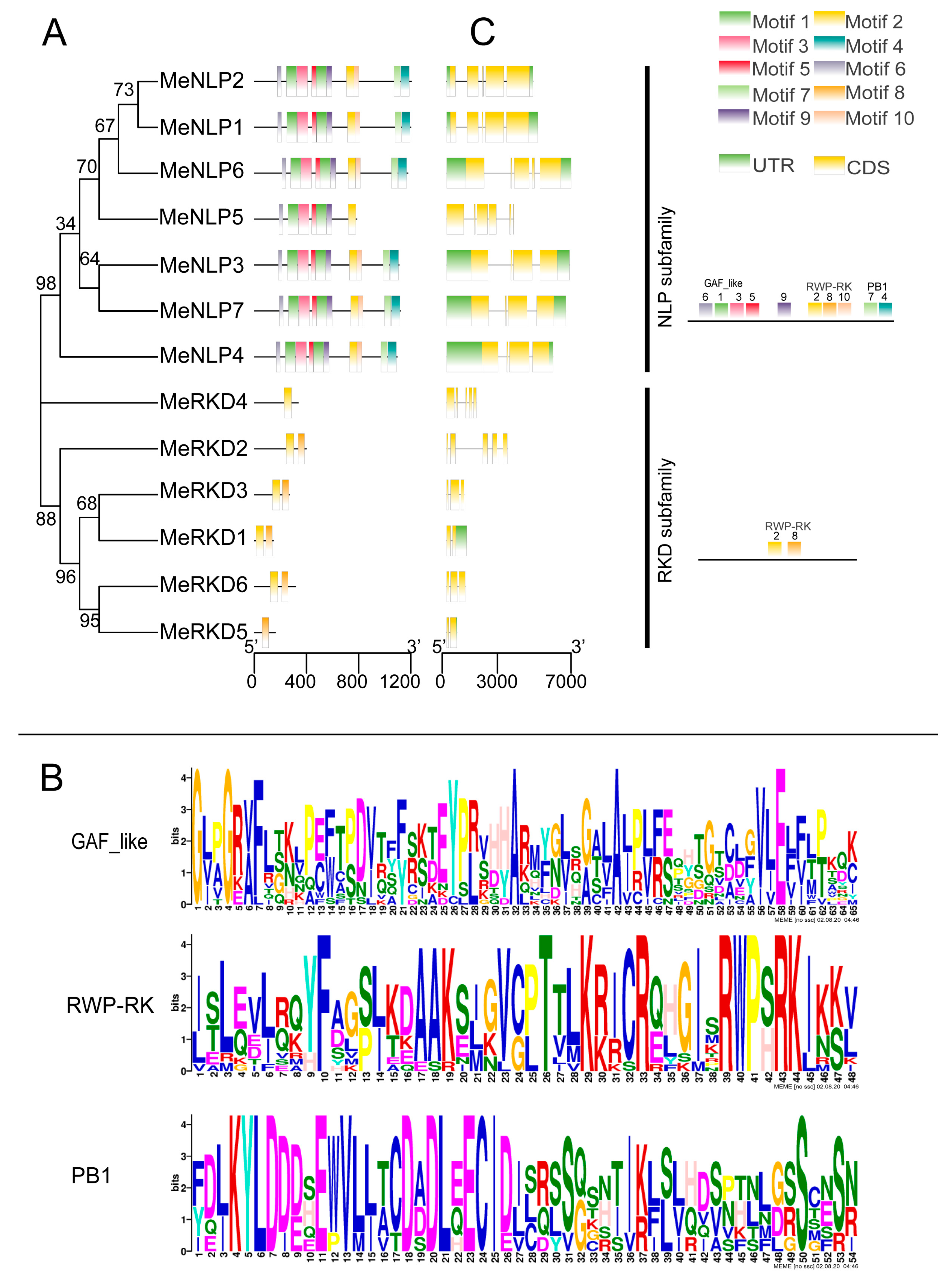
2.2. Phylogenetic Analysis of the RWP-RK Family
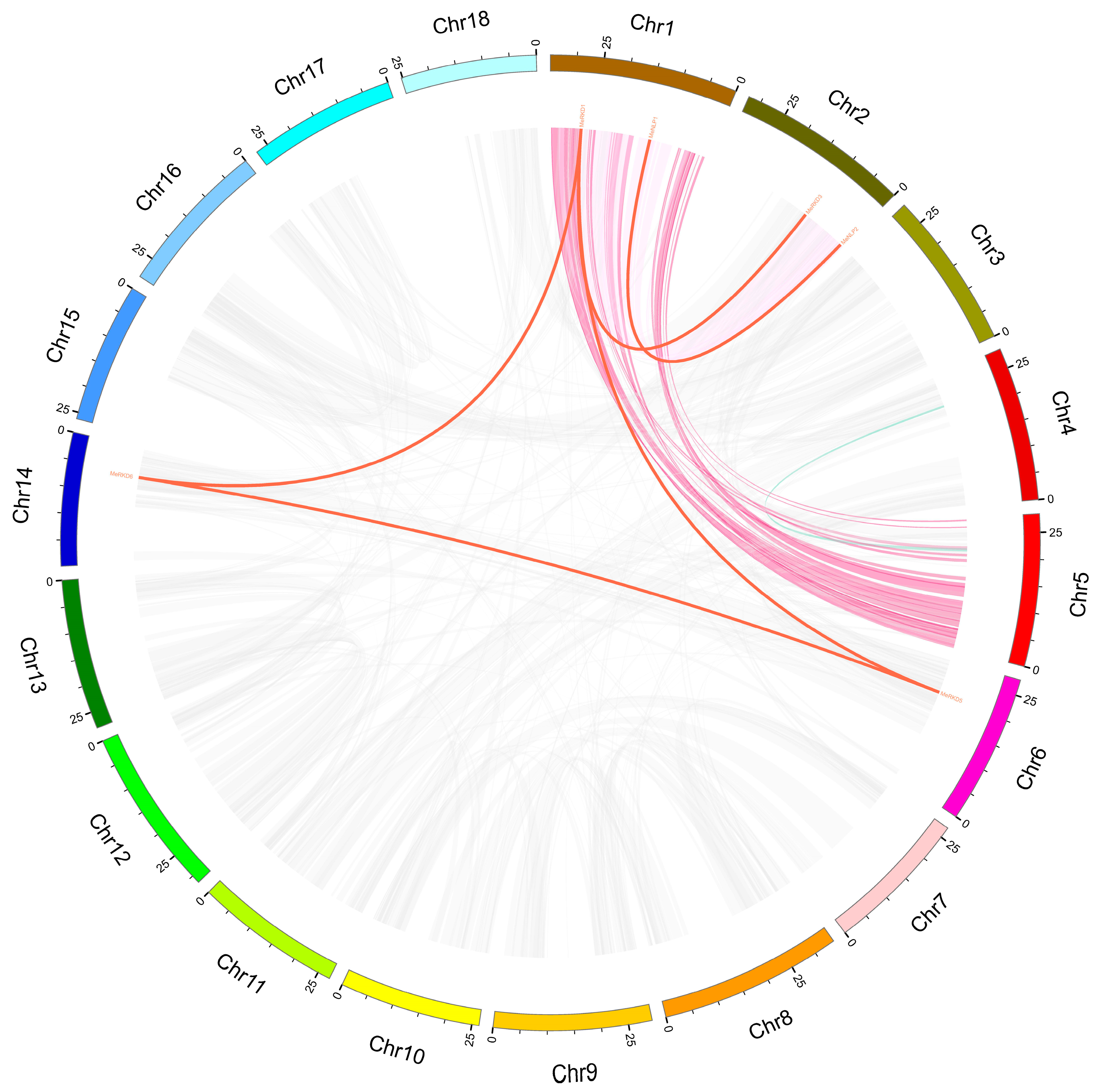
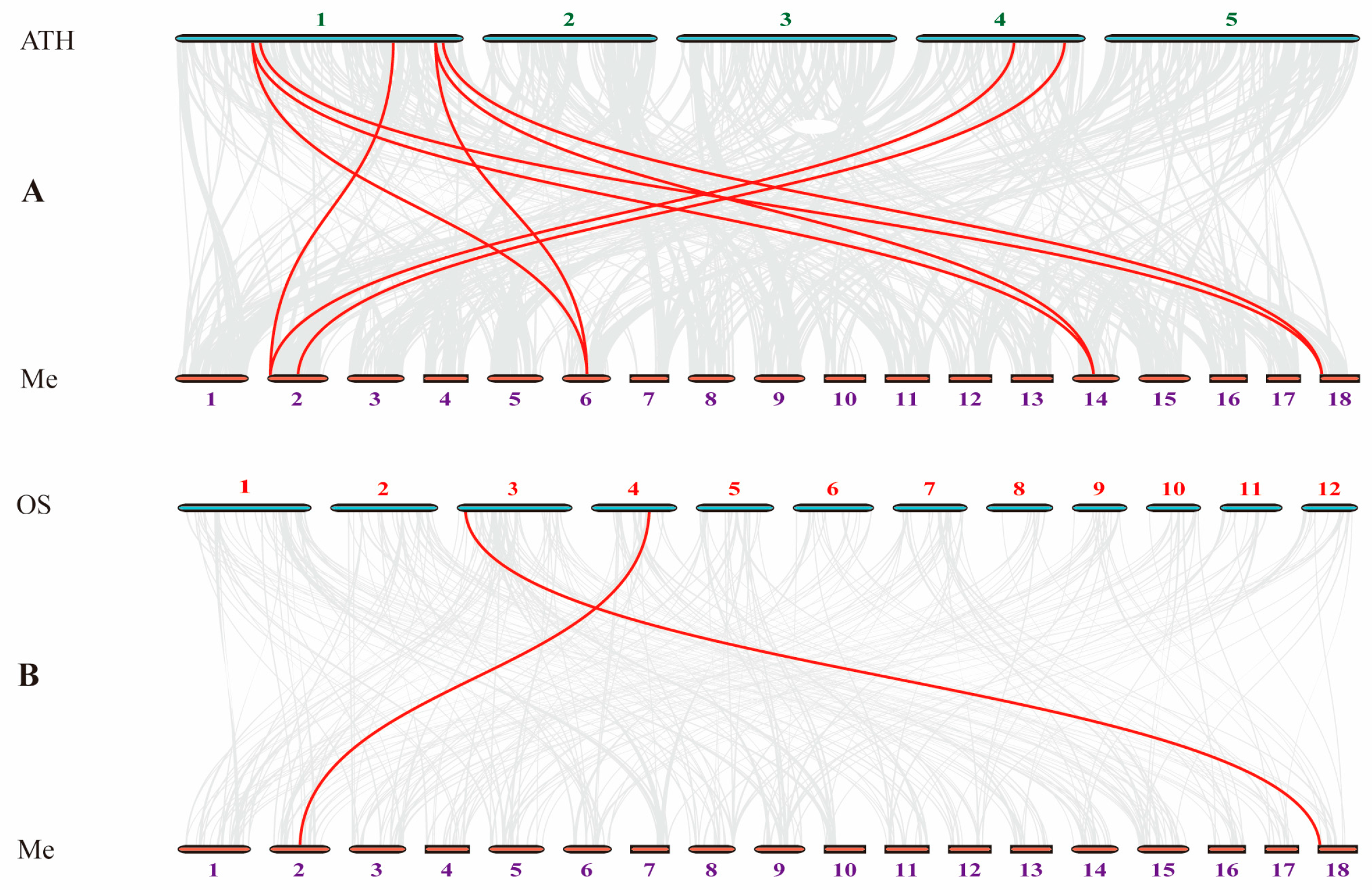
2.3. Synteny Analyses of RWP-RK Genes
2.4. RWP-RK Protein Structure and Conserved Motif Analysis
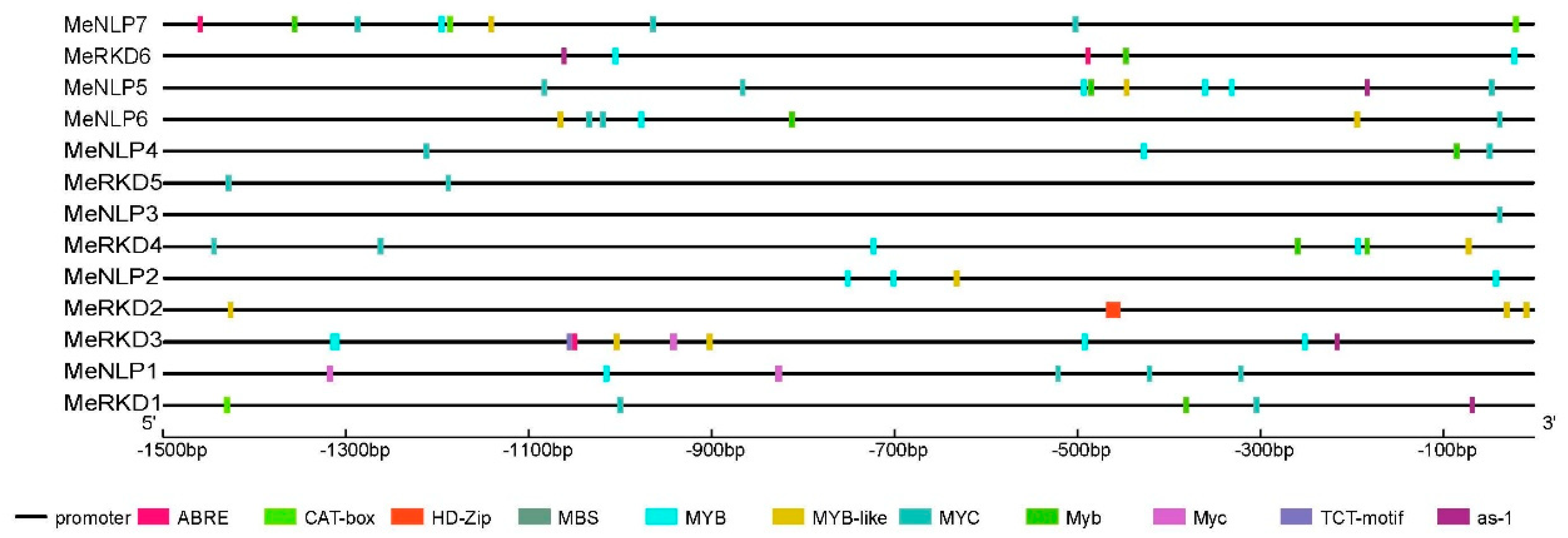
2.5. Analysis of Cassava RWP-RK Gene Promoters
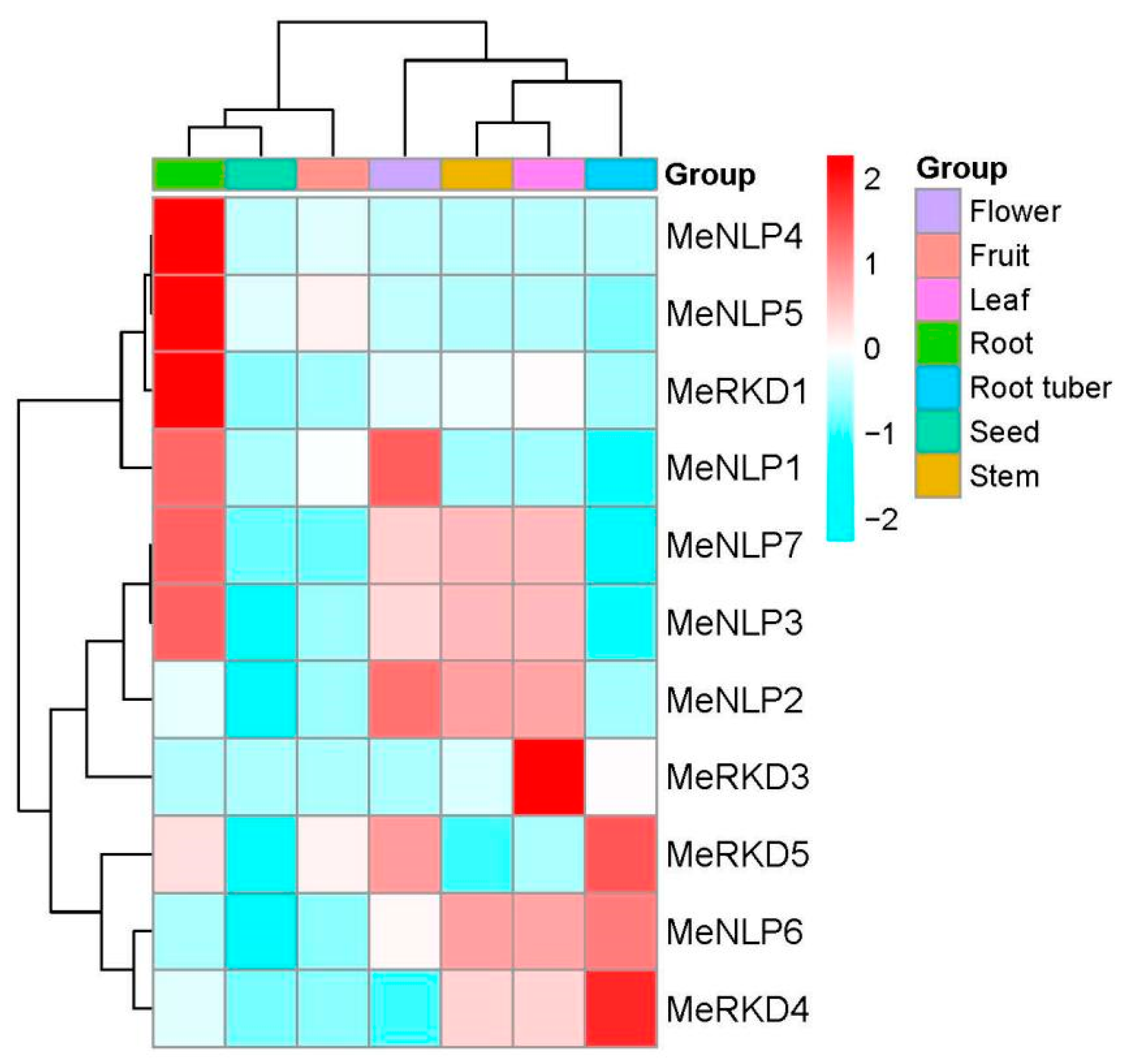
2.6. Transcription Profiles of RWP-RK Genes in Different Cassava Tissues
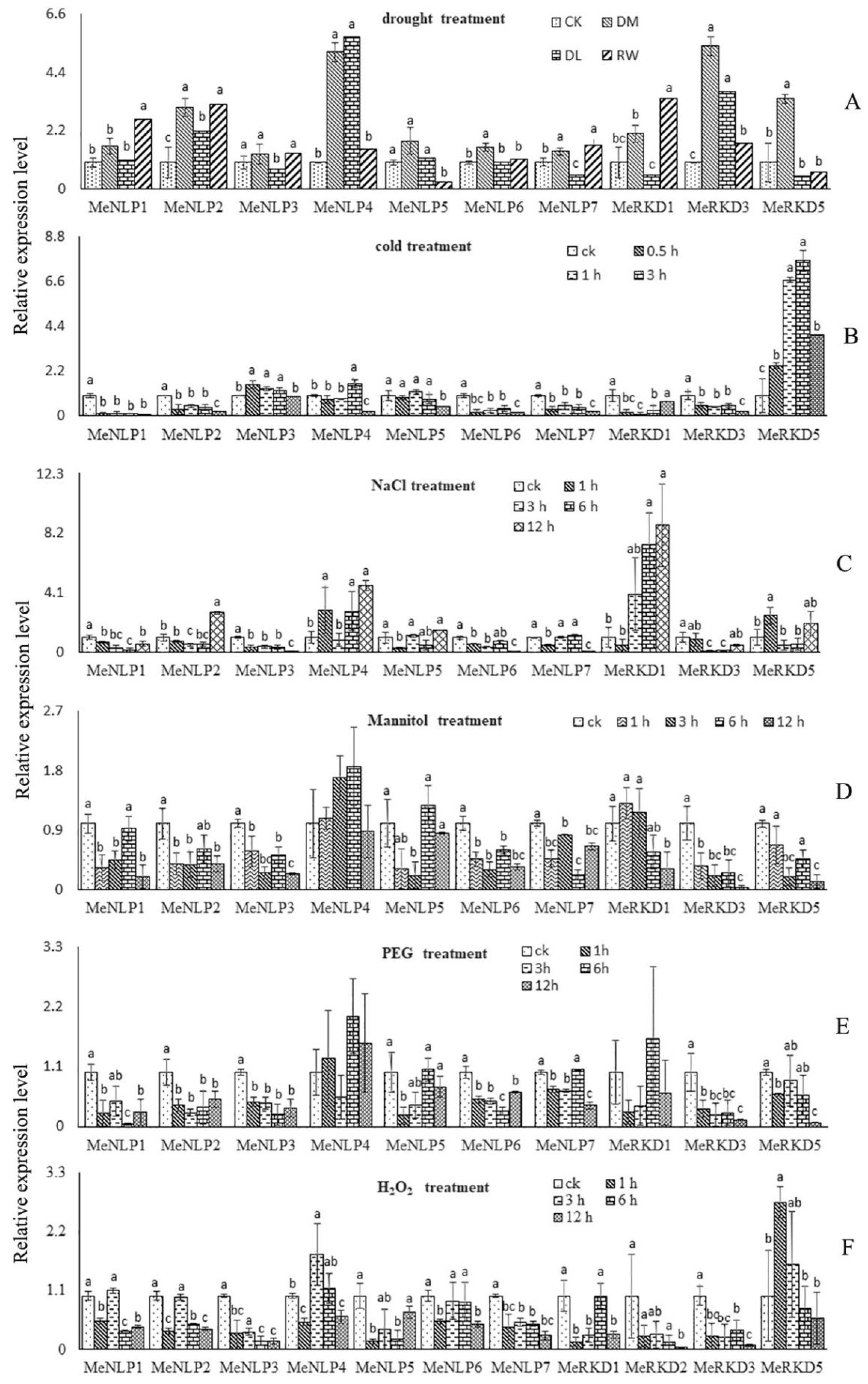
2.7. Expression Patterns of RWP-RK Genes in Response to Different Stresses in Cassava
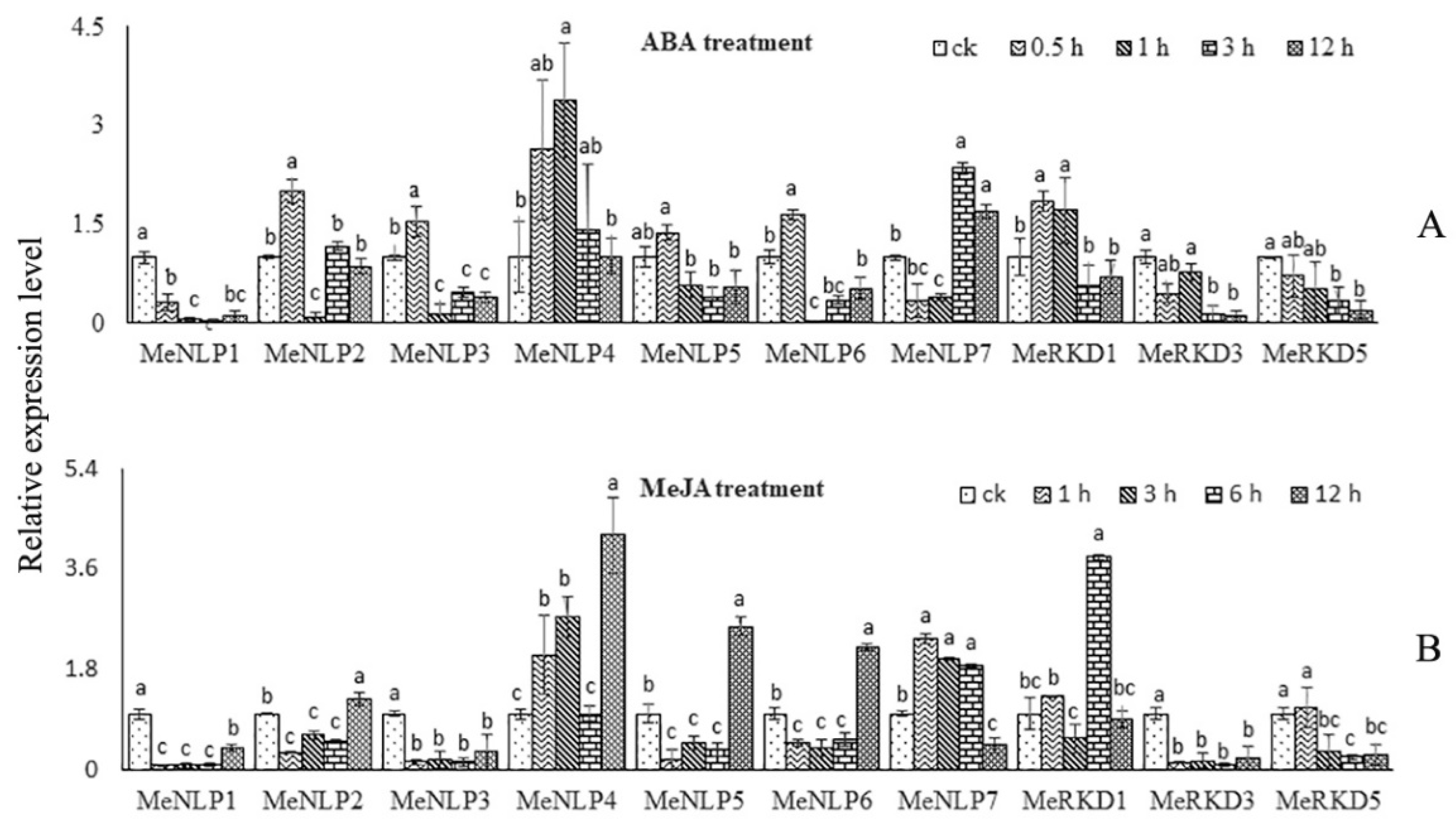
2.8. Expression Patterns of RWP-RK Genes in Response to Different Hormone Treatments in Cassava
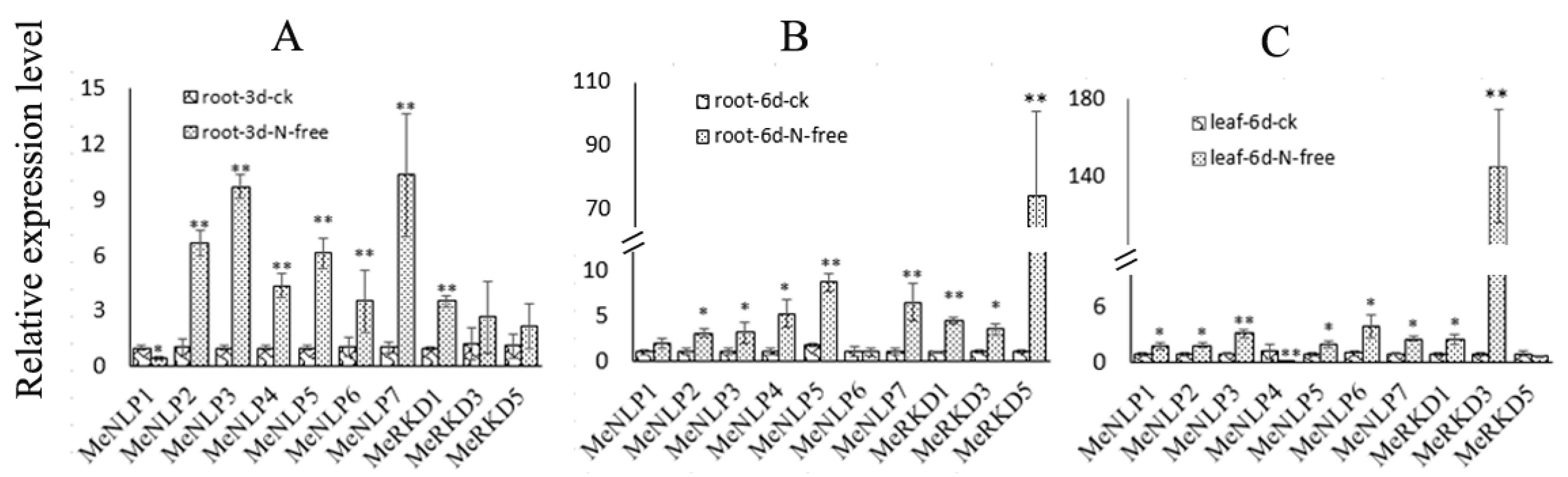
2.9. Expression Patterns of RWP-RK Genes in Response to Nitrogen Starvation Treatments in Cassava
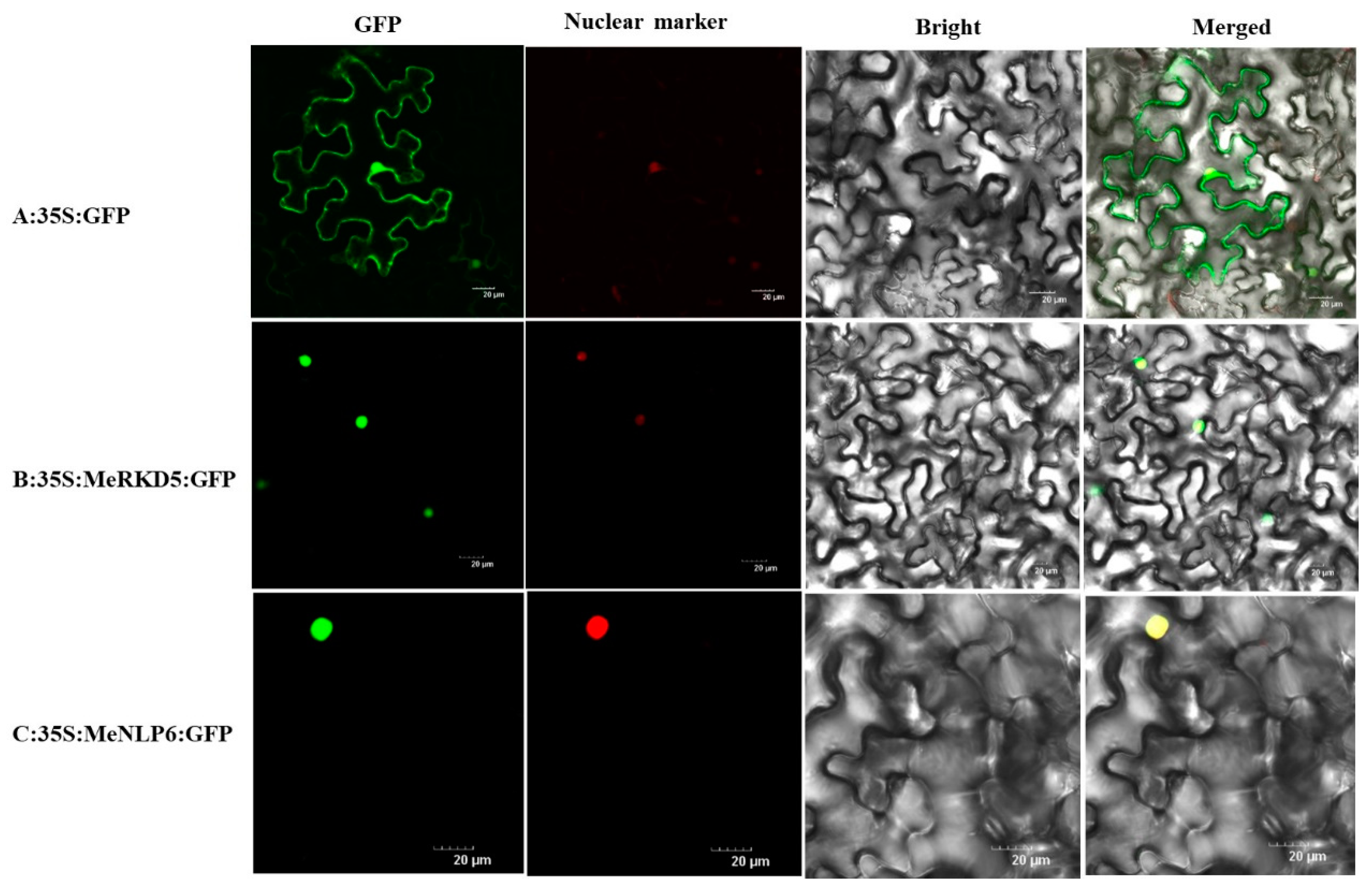
2.10. Subcellular Localization of MeNLP6 and MeRKD5
2.11. Proteins Interacting with NLP6/RKD5 by Yeast Two-Hybrid System
3. Discussion
4. Materials and Methods
4.1. Identification of RWP-RK Family Members in Cassava
4.2. Phylogenetic Analysis of RWP-RK Family Members
4.3. Cassava RWP-RK Gene Duplication
4.4. Cassava RWP-RK Gene Structure and RWP-RK Protein Motif Analysis
4.5. Cassava RWP-RK Gene Promoter Analysis
4.6. Transcription Profile of Cassava RWP-RK Genes in Different Tissues
4.7. Drought and Cold Treatment
4.8. Different Salt, Osmotic, Oxidation and Hormonal Treatments
4.9. Nitrogen Starvation Treatments
4.10. RNA Extraction and qRT-PCR Analysis
4.11. Subcellular Localization of MeNLP6 and MeRKD5
4.12. Yeast One-Hybrid Assay (Y2H)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Khumaida, N.; Ardie, S.; Dianasari, M.; Syukur, M. Cassava (Manihot esculenta Crantz.) Improvement through Gamma Irradiation. Procedia Food Sci. 2015, 3, 27–34. [Google Scholar] [CrossRef]
- Howeler, R.H. Cassava Mineral Nutrition and Fertilization; Centro Internacional de Agricultura Tropical CIAT: Cali, Colombia, 1981. [Google Scholar]
- Zhao, L.; Liu, F.; Crawford, N.M.; Wang, Y. Molecular Regulation of Nitrate Responses in Plants. Int. J. Mol. Sci. 2018, 19, 2039. [Google Scholar] [CrossRef] [PubMed]
- Chardin, C.; Girin, T.; Roudier, F.; Meyer, C.; Krapp, A. The plant RWP-RK transcription factors: Key regulators of nitrogen responses and of gametophyte development. J. Exp. Bot. 2014, 65, 5577–5587. [Google Scholar] [CrossRef] [PubMed]
- Ferris, P.J.; Goodenough, U.W. Mating Type in Chlamydomonas Is Specified by mid, the Minus-Dominance Gene. Genetics. 1997, 146, 859–869. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Goodenough, U.W. Gametogenesis in the Chlamydomonas reinhardtii minus mating type is controlled by two genes, MID and MTD1. Genetics. 2007, 176, 913–925. [Google Scholar] [CrossRef] [PubMed]
- Schauser, L.; Roussis, A.; Stiller, J.; Stougaard, J. A plant regulator controlling development of symbiotic root nodules. Nature 1999, 402, 191–195. [Google Scholar] [CrossRef] [PubMed]
- Borisov, A.Y.; Madsen, L.H.; Tsyganov, V.E.; Umehara, Y.; Voroshilova, V.A.; Batagov, A.O.; Sandal, N.; Mortensen, A.; Schauser, L.; Ellis, N.; et al. The Sym35 gene required for root nodule development in pea is an ortholog of Nin from Lotus japonicus. Plant Physiol. 2003, 131, 1009–1017. [Google Scholar] [CrossRef]
- Camargo, A.; Llamas, A.; Schnell, R.A.; Higuera, J.J.; Gonzalez-Ballester, D.; Lefebvre, P.A.; Fernandez, E.; Galvan, A. Nitrate signaling by the regulatory gene NIT2 in Chlamydomonas. Plant Cell. 2007, 19, 3491–3503. [Google Scholar] [CrossRef]
- Yu, L.H.; Wu, J.; Tang, H.; Yuan, Y.; Wang, S.M.; Wang, Y.P.; Zhu, Q.S.; Li, S.G.; Xiang, C.B. Overexpression of Arabidopsis NLP7 improves plant growth under both nitrogen-limiting and -sufficient conditions by enhancing nitrogen and carbon assimilation. Sci. Rep. 2016, 6, 27795. [Google Scholar] [CrossRef]
- Kumar, A.; Batra, R.; Gahlaut, V.; Gautam, T.; Kumar, S.; Sharma, M.; Tyagi, S.; Singh, K.P.; Balyan, H.S.; Pandey, R.; et al. Genome-wide identification and characterization of gene family for RWP-RK transcription factors in wheat (Triticum aestivum L.). PLoS ONE 2018, 13, e0208409. [Google Scholar] [CrossRef]
- Koszegi, D.; Johnston, A.J.; Rutten, T.; Czihal, A.; Altschmied, L.; Kumlehn, J.; Wust, S.E.; Kirioukhova, O.; Gheyselinck, J.; Grossniklaus, U.; et al. Members of the RKD transcription factor family induce an egg cell-like gene expression program. Plant J. 2011, 67, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Castaings, L.; Camargo, A.; Pocholle, D.; Gaudon, V.; Texier, Y.; Boutet-Mercey, S.; Taconnat, L.; Renou, J.P.; Daniel-Vedele, F.; Fernandez, E.; et al. The nodule inception-like protein 7 modulates nitrate sensing and metabolism in Arabidopsis. Plant J. 2009, 57, 426–435. [Google Scholar] [CrossRef] [PubMed]
- Schauser, L.; Wieloch, W.; Stougaard, J. Evolution of NIN-like proteins in Arabidopsis, rice, and Lotus japonicus. J. Mol. Evol. 2005, 60, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Konishi, M.; Yanagisawa, S. The role of protein-protein interactions mediated by the PB1 domain of NLP transcription factors in nitrate-inducible gene expression. BMC Plant Biol. 2019, 19, 90. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.H.; Niu, Y.; Konishi, M.; Wu, Y.; Du, H.; Sun Chung, H.; Li, L.; Boudsocq, M.; McCormack, M.; Maekawa, S.; et al. Discovery of nitrate-CPK-NLP signalling in central nutrient-growth networks. Nature 2017, 545, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Koi, S.; Hisanaga, T.; Sato, K.; Shimamura, M.; Yamato, K.T.; Ishizaki, K.; Kohchi, T.; Nakajima, K. An Evolutionarily Conserved Plant RKD Factor Controls Germ Cell Differentiation. Curr. Biol. 2016, 26, 1775–1781. [Google Scholar] [CrossRef]
- Tedeschi, F.; Rizzo, P.; Rutten, T.; Altschmied, L.; Baumlein, H. RWP-RK domain-containing transcription factors control cell differentiation during female gametophyte development in Arabidopsis. New Phytol. 2017, 213, 1909–1924. [Google Scholar] [CrossRef]
- Soyano, T.; Kouchi, H.; Hirota, A.; Hayashi, M. Nodule inception directly targets NF-Y subunit genes to regulate essential processes of root nodule development in Lotus japonicus. PLoS Genet. 2013, 9, e1003352. [Google Scholar] [CrossRef]
- Marchive, C.; Roudier, F.; Castaings, L.; Brehaut, V.; Blondet, E.; Colot, V.; Meyer, C.; Krapp, A. Nuclear retention of the transcription factor NLP7 orchestrates the early response to nitrate in plants. Nat. Commun. 2013, 4, 1713. [Google Scholar] [CrossRef]
- Yan, D.; Easwaran, V.; Chau, V.; Okamoto, M.; Ierullo, M.; Kimura, M.; Endo, A.; Yano, R.; Pasha, A.; Gong, Y.; et al. NIN-like protein 8 is a master regulator of nitrate-promoted seed germination in Arabidopsis. Nat. Commun. 2016, 7, 13179. [Google Scholar] [CrossRef]
- Marsh, J.F.; Rakocevic, A.; Mitra, R.M.; Brocard, L.; Sun, J.; Eschstruth, A.; Long, S.R.; Schultze, M.; Ratet, P.; Oldroyd, G.E. Medicago truncatula NIN is essential for rhizobial-independent nodule organogenesis induced by autoactive calcium/calmodulin-dependent protein kinase. Plant Physiol. 2007, 144, 324–335. [Google Scholar] [CrossRef]
- Lin, J.S.; Li, X.; Luo, Z.; Mysore, K.S.; Wen, J.; Xie, F. NIN interacts with NLPs to mediate nitrate inhibition of nodulation in Medicago truncatula. Nat. Plants 2018, 4, 942–952. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xin, H.; Qu, L.; Ning, J.; Peng, X.; Yan, T.; Ma, L.; Li, S.; Sun, M.X. Dynamic changes of transcript profiles after fertilization are associated with de novo transcription and maternal elimination in tobacco zygote, and mark the onset of the maternal-to-zygotic transition. Plant J. 2011, 65, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Yokota, K.; Hayashi, M. Function and evolution of nodulation genes in legumes. Cell. Mol. Life Sci. 2011, 68, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.W.; Breakspear, A.; Guan, D.; Cerri, M.R.; Jackson, K.; Jiang, S.; Robson, F.; Radhakrishnan, G.V.; Roy, S.; Bone, C.; et al. NIN Acts as a Network Hub Controlling a Growth Module Required for Rhizobial Infection. Plant Physiol. 2019, 179, 1704–1722. [Google Scholar] [CrossRef]
- Mu, X.; Luo, J. Evolutionary analyses of NIN-like proteins in plants and their roles in nitrate signaling. Cell. Mol. Life Sci. 2019, 76, 3753–3764. [Google Scholar] [CrossRef]
- Wu, Z.; Liu, H.; Huang, W.; Yi, L.; Qin, E.; Yang, T.; Wang, J.; Qin, R. Genome-Wide Identification, Characterization, and Regulation of RWP-RK Gene Family in the Nitrogen-Fixing Clade. Plants 2020, 9, 1178. [Google Scholar] [CrossRef]
- Clavijo, F.; Diedhiou, I.; Vaissayre, V.; Brottier, L.; Acolatse, J.; Moukouanga, D.; Crabos, A.; Auguy, F.; Franche, C.; Gherbi, H.; et al. The Casuarina NIN gene is transcriptionally activated throughout Frankia root infection as well as in response to bacterial diffusible signals. New Phytol. 2015, 208, 887–903. [Google Scholar] [CrossRef]
- Liu, M.; Soyano, T.; Yano, K.; Hayashi, M.; Kawaguchi, M. ERN1 and CYCLOPS coordinately activate NIN signaling to promote infection thread formation in Lotus japonicus. J. Plant Res. 2019, 132, 641–653. [Google Scholar] [CrossRef]
- Liu, J.; Bisseling, T. Evolution of NIN and NIN-like Genes in Relation to Nodule Symbiosis. Genes 2020, 11, 777. [Google Scholar] [CrossRef]
- Zhang, T.T.; Kang, H.; Fu, L.L.; Sun, W.J.; Gao, W.S.; You, C.X.; Wang, X.F.; Hao, Y.J. NIN-like protein 7 promotes nitrate-mediated lateral root development by activating transcription of TRYPTOPHAN AMINOTRANSFERASE RELATED 2. Plant Sci. 2021, 303, 110771. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Liu, Y.; Jiang, L.; Wang, Y.; Lv, Y.; Zhou, L.; Liang, S.; Bao, H.; Zhao, H. Genome-wide analysis of maize NLP transcription factor family revealed the roles in nitrogen response. Plant Growth Regul. 2017, 84, 95–105. [Google Scholar] [CrossRef]
- Liu, M.; Chang, W.; Fan, Y.; Sun, W.; Qu, C.; Zhang, K.; Liu, L.; Xu, X.; Tang, Z.; Li, J.; et al. Genome-Wide Identification and Characterization of NODULE-INCEPTION-Like Protein (NLP) Family Genes in Brassica napus. Int. J. Mol. Sci. 2018, 19, 2270. [Google Scholar] [CrossRef]
- Feng, Z.Q.; Li, T.; Wang, X.; Sun, W.J.; Zhang, T.T.; You, C.X.; Wang, X.F. Identification and characterization of apple MdNLP7 transcription factor in the nitrate response. Plant Sci. 2022, 316, 111158. [Google Scholar] [CrossRef]
- Reddy, A.S.; Marquez, Y.; Kalyna, M.; Barta, A. Complexity of the alternative splicing landscape in plants. Plant Cell. 2013, 25, 3657–3683. [Google Scholar] [CrossRef] [PubMed]
- Kuo, Y.T.; Chao, Y.T.; Chen, W.C.; Shih, M.C.; Chang, S.B. Segmental and tandem chromosome duplications led to divergent evolution of the chalcone synthase gene family in Phalaenopsis orchids. Ann. Bot. 2019, 123, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Holub, E.B. The arms race is ancient history in Arabidopsis, the wildflower. Nature 2001, 1, 516–527. [Google Scholar] [CrossRef]
- Zhang, P.; Gu, Z.L.; Li, W.H. Different evolutionary patterns between young duplicate genes in the human genome. Genome Biol. 2003, 4, R56. [Google Scholar] [CrossRef]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef]
- Gomez-Porras, J.L.; Riano-Pachon, D.M.; Dreyer, I.; Mayer, J.E.; Mueller-Roeber, B. Genome-wide analysis of ABA-responsive elements ABRE and CE3 reveals divergent patterns in Arabidopsis and rice. BMC Genom. 2007, 8, 260. [Google Scholar] [CrossRef]
- He, G.; Liu, P.; Zhao, H.; Sun, J. The HD-ZIP II Transcription Factors Regulate Plant Architecture through the Auxin Pathway. Int. J. Mol. Sci. 2020, 21, 3250. [Google Scholar] [CrossRef] [PubMed]
- Garreton, V.; Carpinelli, J.; Jordana, X.; Holuigue, L. The as-1 promoter element is an oxidative stress-responsive element and salicylic acid activates it via oxidative species. Plant Physiol. 2002, 130, 1516–1526. [Google Scholar] [CrossRef] [PubMed]
- Ruan, M.B.; Guo, X.; Wang, B.; Yang, Y.L.; Li, W.Q.; Yu, X.L.; Zhang, P.; Peng, M. Genome-wide characterization and expression analysis enables identification of abiotic stress-responsive MYB transcription factors in cassava (Manihot esculenta). J. Exp. Bot. 2017, 68, 3657–3672. [Google Scholar] [CrossRef]
- Tran, L.S.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell. 2004, 16, 2481–2498. [Google Scholar] [CrossRef] [PubMed]
- Parry, T.J.; Theisen, J.W.; Hsu, J.Y.; Wang, Y.L.; Corcoran, D.L.; Eustice, M.; Ohler, U.; Kadonaga, J.T. The TCT motif, a key component of an RNA polymerase II transcription system for the translational machinery. Genes Dev. 2010, 24, 2013–2018. [Google Scholar] [CrossRef]
- Abe, H.; Yamaguchi-Shinozaki, K.; Urao, T.; Iwasaki, T.; Hosokawa, D.; Shinozaki, K. Role of Arabidopsis MYC and MYB Homologs in Drought- and Abscisic Acid-Regulated Gene Expression. Plant Cell. 1997, 9, 1859–1868. [Google Scholar] [CrossRef]
- Jagadhesan, B.; Sathee, L.; Meena, H.S.; Jha, S.K.; Chinnusamy, V.; Kumar, A.; Kumar, S. Genome wide analysis of NLP transcription factors reveals their role in nitrogen stress tolerance of rice. Sci. Rep. 2020, 10, 9368. [Google Scholar] [CrossRef]
- Magadum, S.; Banerjee, U.; Murugan, P.; Gangapur, D.; Ravikesavan, R. Gene duplication as a major force in evolution. J. Genet. 2013, 92, 155–161. [Google Scholar] [CrossRef]
- Konishi, M.; Yanagisawa, S. Arabidopsis NIN-like transcription factors have a central role in nitrate signalling. Nat. Commun. 2013, 4, 1617. [Google Scholar] [CrossRef]
- Hruz, T.; Laule, O.; Szabo, G.; Wessendorp, F.; Bleuler, S.; Oertle, L.; Widmayer, P.; Gruissem, W.; Zimmermann, P. Genevestigator v3: A reference expression database for the meta-analysis of transcriptomes. Adv. Bioinform. 2008, 2008, 420747. [Google Scholar] [CrossRef]
- Wang, M.; Hasegawa, T.; Beier, M.; Hayashi, M.; Ohmori, Y.; Yano, K.; Teramoto, S.; Kamiya, T.; Fujiwara, T. Growth and Nitrate Reductase Activity Are Impaired in Rice Osnlp4 Mutants Supplied with Nitrate. Plant Cell Physiol. 2021, 62, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Ge, M.; Wang, Y.; Liu, Y.; Jiang, L.; He, B.; Ning, L.; Du, H.; Lv, Y.; Zhou, L.; Lin, F.; et al. The NIN-like protein 5 (ZmNLP5) transcription factor is involved in modulating the nitrogen response in maize. Plant J. 2020, 102, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, Z.S.; Xia, J.Q.; Alfatih, A.; Song, Y.; Huang, Y.J.; Wan, G.Y.; Sun, L.Q.; Tang, H.; Liu, Y.; et al. Rice NIN-LIKE PROTEIN 4 plays a pivotal role in nitrogen use efficiency. Plant Biotechnol. J. 2021, 19, 448–461. [Google Scholar] [CrossRef]
- Higuera, J.J.; Fernandez, E.; Galvan, A. Chlamydomonas NZF1, a tandem-repeated zinc finger factor involved in nitrate signalling by controlling the regulatory gene NIT2. Plant Cell Environ. 2014, 37, 2139–2150. [Google Scholar] [CrossRef]
- Mekawy, A.M.M.; Assaha, D.V.M.; Ueda, A. Constitutive overexpression of rice metallothionein-like gene OsMT-3a enhances growth and tolerance of Arabidopsis plants to a combination of various abiotic stresses. J. Plant Res. 2020, 133, 429–440. [Google Scholar] [CrossRef]
- Perez-Ruiz, R.V.; Garcia-Ponce, B.; Marsch-Martinez, N.; Ugartechea-Chirino, Y.; Villajuana-Bonequi, M.; de Folter, S.; Azpeitia, E.; Davila-Velderrain, J.; Cruz-Sanchez, D.; Garay-Arroyo, A.; et al. XAANTAL2 (AGL14) Is an Important Component of the Complex Gene Regulatory Network that Underlies Arabidopsis Shoot Apical Meristem Transitions. Mol. Plant. 2015, 8, 796–813. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Tian, S.; Lu, L.; Shohag, M.J.; Yang, X. Metallothionein 2 (SaMT2) from Sedum alfredii Hance confers increased Cd tolerance and accumulation in yeast and tobacco. PLoS ONE. 2014, 9, e102750. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Liu, P.; Shao, J.; Li, C.; Wang, B.; Guo, X.; Yan, B.; Xia, Y.; Peng, M. Analysis of different strategies adapted by two cassava cultivars in response to drought stress: Ensuring survival or continuing growth. J. Exp. Bot. 2015, 66, 1477–1488. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wei, W.; Li, F.; Zhang, L.; Deng, X.; Liu, Y.; Yang, S. The Plastidial Glyceraldehyde-3-Phosphate Dehydrogenase Is Critical for Abiotic Stress Response in Wheat. Int. J. Mol. Sci. 2019, 20, 1104. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name | Identify | Gene Annotation | Amino Acid |
|---|---|---|---|
| MeNLP6 | Manes.03G056700 | PPR repeat family (PPR_2) | 1498 |
| MeRKD5 | Manes.02G030100 | Agamous-like mads-box protein AGL14 | 216 |
| Manes.01G174400 | Metallothionein (Metallothio_2) | 79 | |
| Manes.08G023700 | Zine finger FYVE domain containing protein | 343 | |
| Manes.05G125200 | Glyceraldehyde-3-phosphate dehydrogenase | 453 | |
| Manes.15G059000 | E3 Ubiquitin-protein ligase HUWE1 | 3649 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, C.; Guo, X.; Yu, X.; Li, S.; Li, W.; Yu, X.; An, F.; Zhao, P.; Ruan, M. Genome-Wide Survey of the RWP-RK Gene Family in Cassava (Manihot esculenta Crantz) and Functional Analysis. Int. J. Mol. Sci. 2023, 24, 12925. https://doi.org/10.3390/ijms241612925
Lin C, Guo X, Yu X, Li S, Li W, Yu X, An F, Zhao P, Ruan M. Genome-Wide Survey of the RWP-RK Gene Family in Cassava (Manihot esculenta Crantz) and Functional Analysis. International Journal of Molecular Sciences. 2023; 24(16):12925. https://doi.org/10.3390/ijms241612925
Chicago/Turabian StyleLin, Chenyu, Xin Guo, Xiaohui Yu, Shuxia Li, Wenbin Li, Xiaoling Yu, Feng An, Pingjuan Zhao, and Mengbin Ruan. 2023. "Genome-Wide Survey of the RWP-RK Gene Family in Cassava (Manihot esculenta Crantz) and Functional Analysis" International Journal of Molecular Sciences 24, no. 16: 12925. https://doi.org/10.3390/ijms241612925