
Discovering Loci for Breeding Prospective and Phenology in Wheat Mediterranean Landraces by Environmental and eigenGWAS

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
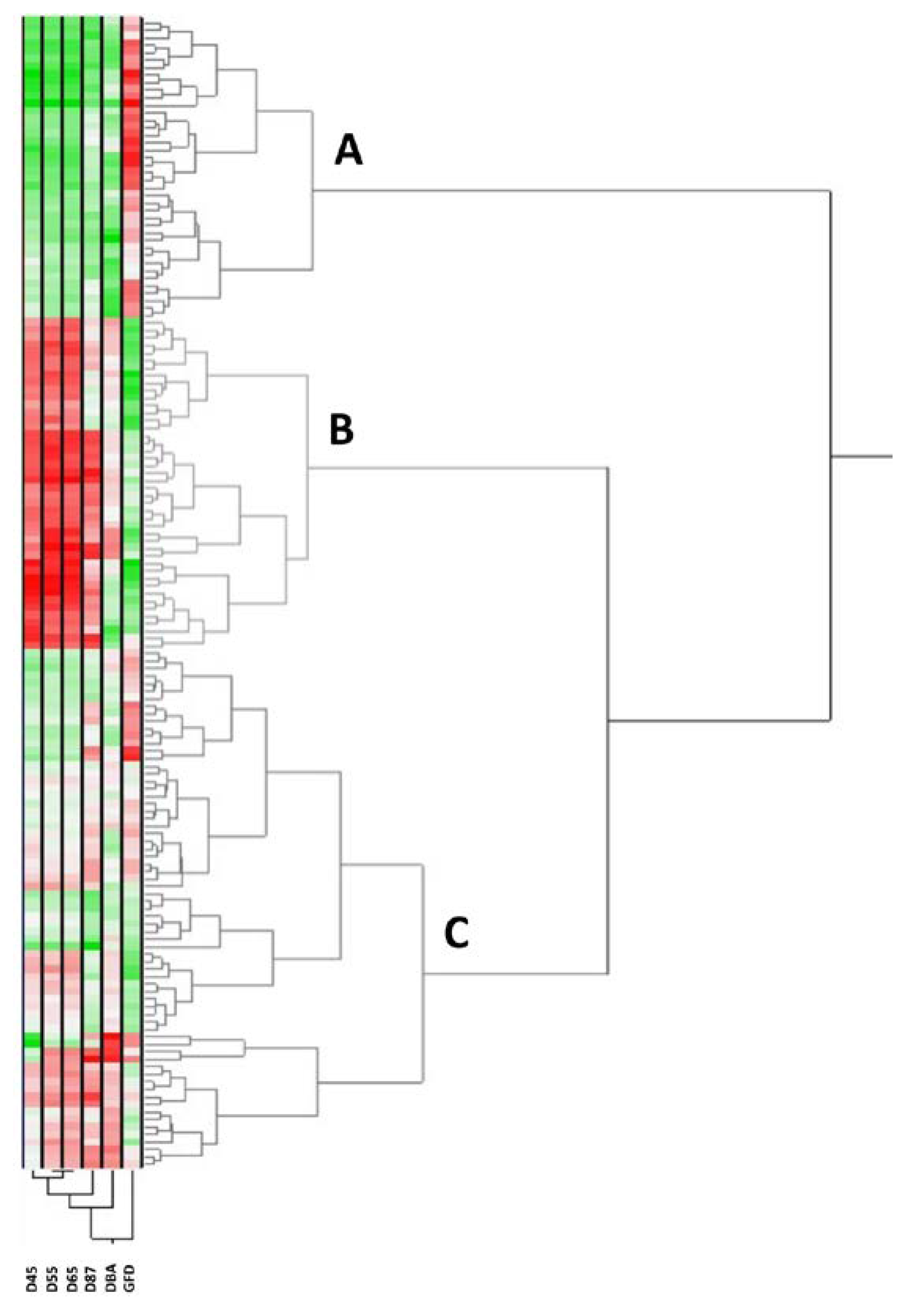
2.1. Climatic Data and Phenology Assessment
2.2. Detection of Loci for Phenology Adjustment
2.3. Detection of Loci for Climatic Variables by envGWAS
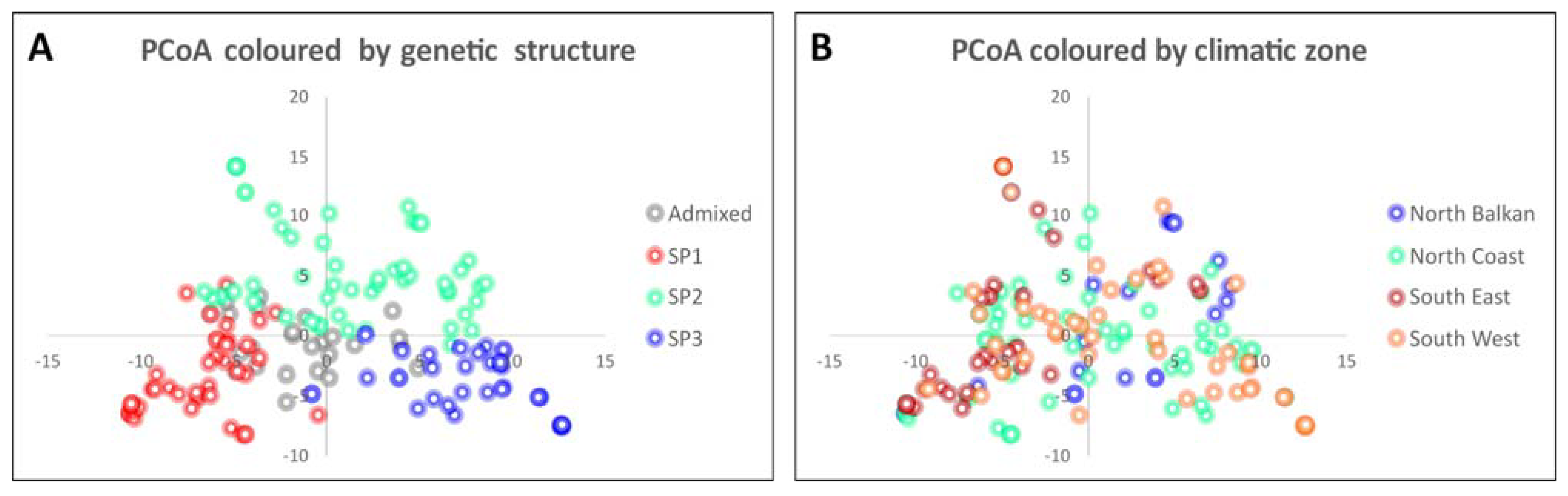
2.4. Detection of Loci for Genetic Differentiation by eigenGWAS
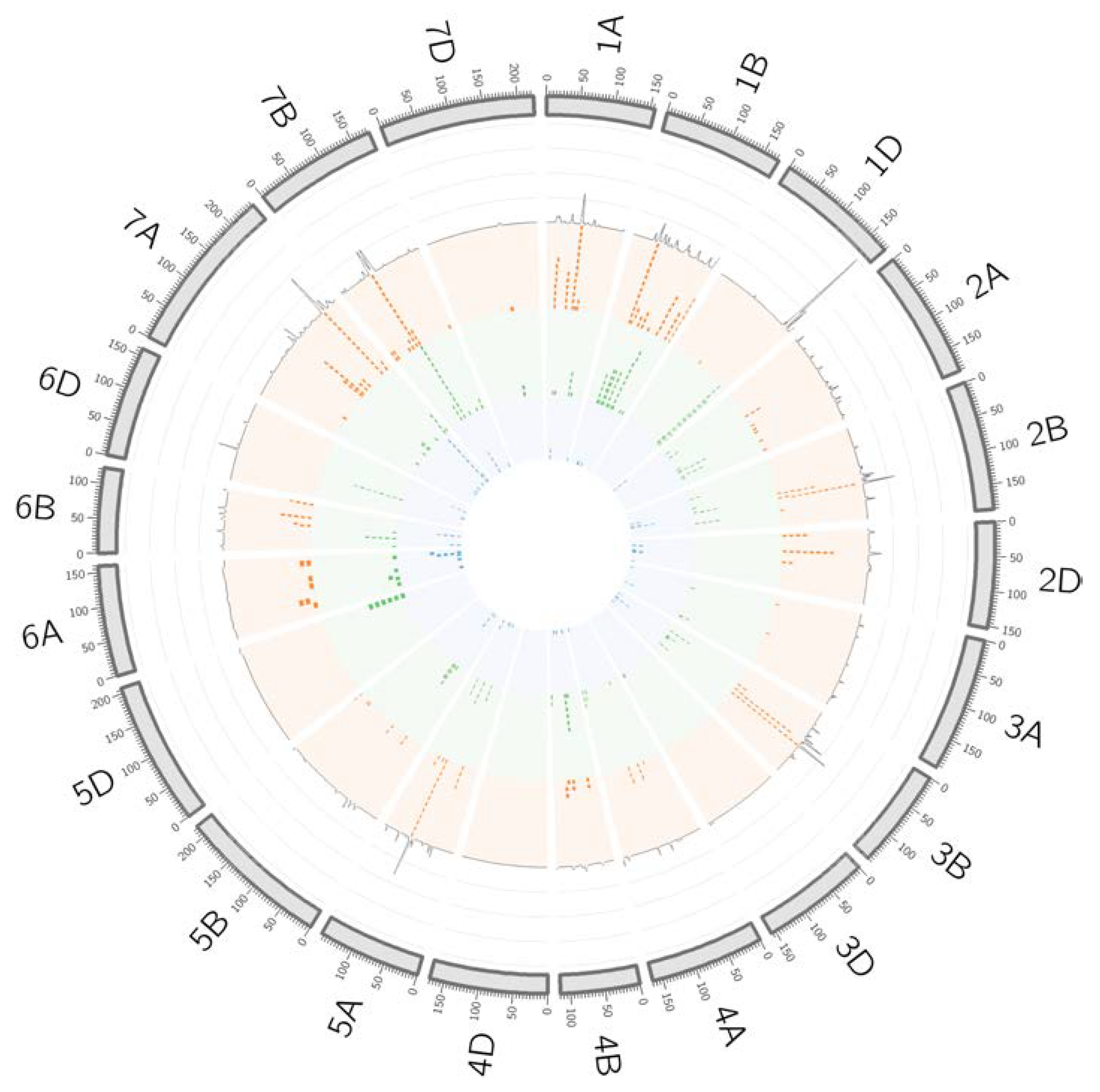
2.5. Identification of QTL Hotspots
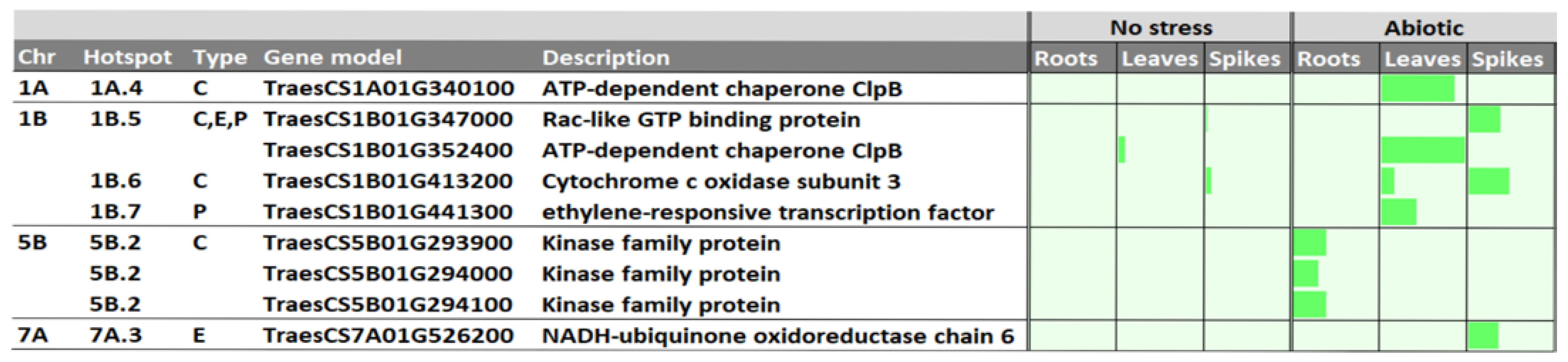
2.6. In Silico Analysis of Candidate Genes
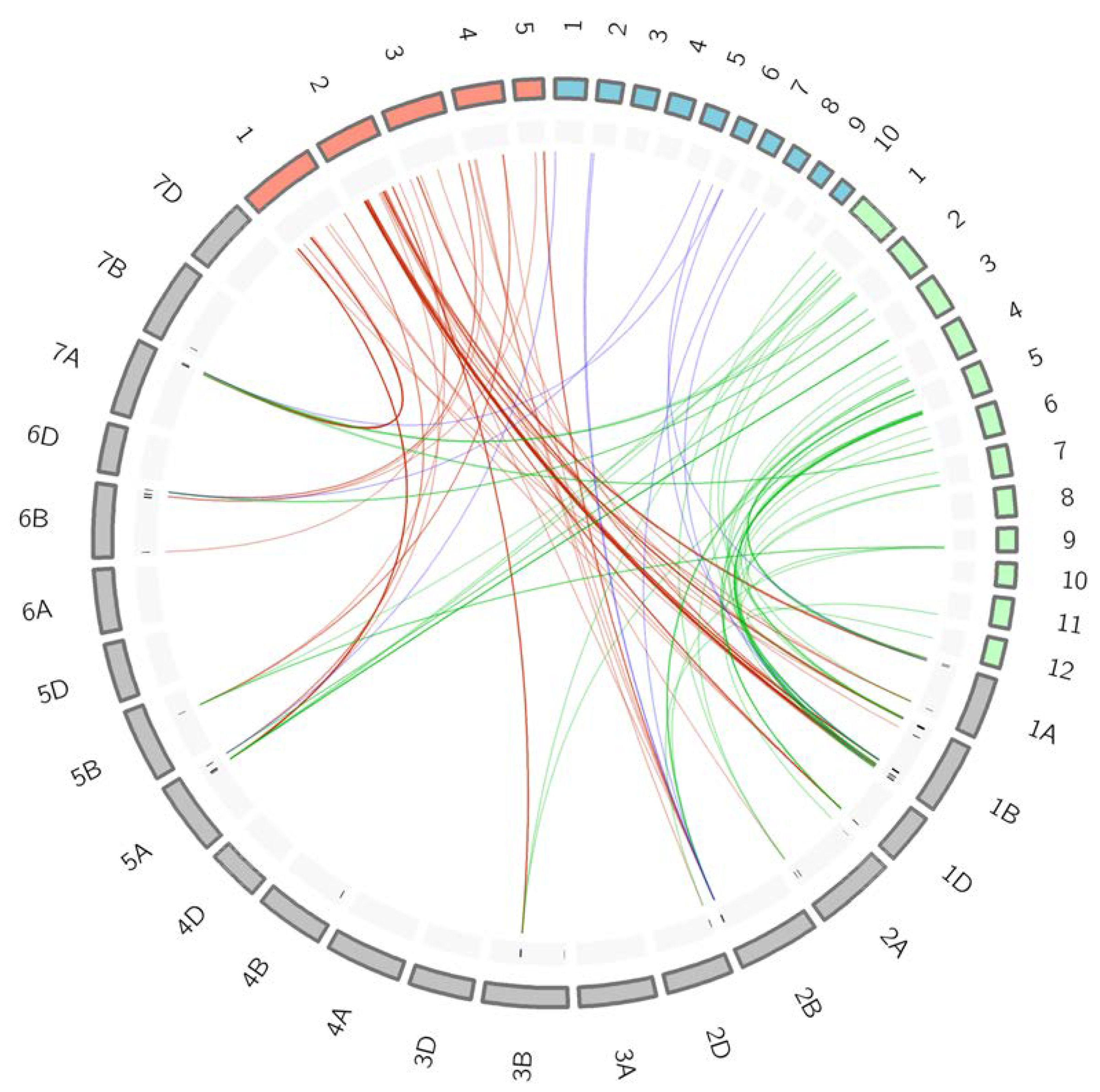
2.7. Synteny Analysis within Cereal Species
3. Discussion
3.1. Phenology Fitting
3.2. Genetic Control of Environmental Conditions
3.3. Genetic Loci for Differentiation Patterns among Mediterranean Landraces
3.4. Candidate Genes
3.5. Synteny among Cereal Species
4. Materials and Methods
4.1. Plant Material
4.2. Phenology Assessment
4.3. Environmental Variables
4.4. Genome-Wide Association Analyses
4.5. In Silico Gene Expression Analyses and Synteny against Cereal Genomes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Royo, C.; Soriano, J.M.; Alvaro, F. Wheat: A Crop in the Bottom of the Mediterranean Diet Pyramid. In Mediterranean Identities—Environment, Society, Culture; Fuerst-Bjelis, B., Ed.; IntechOpen: London, UK, 2017; pp. 381–399. [Google Scholar]
- Lopes, M.S. Will temperature and rainfall changes prevent yield progress in Europe. Food Energy Secur. 2022, 11, e372. [Google Scholar] [CrossRef]
- Soriano, J.M.; Villegas, D.; Sorrells, M.E.; Royo, C. Durum wheat landraces from east and west regions of the mediterranean basin are genetically distinct for yield components and phenology. Front. Plant Sci. 2018, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Araus, J.L.; Slafer, G.A.; Royo, C.; Serret, M.D. Breeding for yield potential and stress adaptation in cereals. Crit. Rev. Plant Sci. 2008, 27, 377–412. [Google Scholar] [CrossRef]
- Altenbach, S.B. New insights into the effects of high temperature.; drought and post-anthesis fertilizer on wheat grain development. J. Cereal Sci. 2012, 56, 39–50. [Google Scholar] [CrossRef]
- Rufo, R.; Soriano, J.M.; Villegas, D.; Royo, C.; Bellvert, J. Using Unmanned Aerial Vehicle and Ground-Based RGB Indices to Assess Agronomic Performance of Wheat Landraces and Cultivars in a Mediterranean-Type Environment. Remote Sens. 2021, 13, 1187. [Google Scholar] [CrossRef]
- Iglesias, A.; Garrote, L. Adaptation strategies for agricultural water management under climate change in Europe. Agric. Water Manag. 2015, 155, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Skovmand, B.; Warburton, M.L.; Sullivan, S.N.; Lage, J. Managing and collecting genetic resources. In Durum Wheat Breeding: Current Approaches and Future Strategies; Royo, C., Nachit, M.N., Di Fonzo, N., Araus, J.L., Pfeiffer, W.H., Slafer, G.A., Eds.; Haworth Press: New York, NY, USA, 2005; pp. 142–163. [Google Scholar]
- Lopes, M.S.; El-Basyoni, I.; Baenziger, P.S.; Singh, S.; Royo, C.; Ozbek, K.; Aktas, H.; Ozer, E.; Ozdemir, F.; Manickavelu, A.; et al. Exploiting genetic diversity from landraces in wheat breeding for adaptation to climate change. J. Exp. Bot. 2015, 66, 3477–3486. [Google Scholar] [CrossRef]
- Soriano, J.M.; Villegas, D.; Aranzana, M.J.; García del Moral, L.F.; Royo, C. Genetic structure of modern durum wheat cultivars and mediterranean landraces matches with their agronomic performance. PLoS ONE 2016, 11, e0160983. [Google Scholar] [CrossRef] [Green Version]
- Pascual, L.; Ruiz, M.; López-Fernández, M.; Pérez-Peña, H.; Benavente, E.; Vázquez, J.F.; Sansaloni, C.; Giraldo, P. Genomic analysis of Spanish wheat landraces reveals their variability and potential for breeding. BMC Genom. 2020, 21, 122. [Google Scholar] [CrossRef] [Green Version]
- Rufo, R.; Alvaro, F.; Royo, C.; Soriano, J.M. From landraces to improved cultivars: Assessment of genetic diversity and population structure of Mediterranean wheat using SNP markers. PLoS ONE 2019, 14, e0219867. [Google Scholar] [CrossRef]
- Soriano, J.M.; Sansaloni, C.; Ammar, K.; Royo, C. Labelling Selective Sweeps Used in Durum Wheat Breeding from a Diverse and Structured Panel of Landraces and Cultivars. Biology 2021, 10, 258. [Google Scholar] [CrossRef] [PubMed]
- Sharma, R.; Cockram, J.; Gardenr, K.A.; Russell, J.; Ramsay, L.; Thomas, W.T.B.; O’Sullivan, D.M.; Powell, W.; Mackay, I. J Trends of genetic changes uncovered by Env- and Eigen-GWAS in wheat and barley. Theor. Appl. Genet. 2022, 135, 667–678. [Google Scholar] [CrossRef] [PubMed]
- Crespo-Herrera, L.A.; Crossa, J.; Huerta-Espino, J.; Vargas, M.; Mondal, S.; Velu, G.; Payne, T.S.; Braun, H.; Singh, R.P. Genetic Gains for Grain Yield in CIMMYT’s Semi-Arid Wheat Yield Trials Grown in Suboptimal Environments. Crop Sci. 2018, 58, 1890–1898. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumudini, S.; Andrade, F.H.; Boote, K.J.; Brown, G.A.; Dzosti, K.A.; Edmeades, G.O.; Gocken, T.; Goodwin, M.; Halter, A.L.; Hammer, G.L.; et al. Predicting maize phenology: Intercomparison of functions for developmental response to temperature. Agron. J. 2014, 106, 2087–2097. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Kabbaj, H.; El Hassouni, K.; Maccaferri, M.; Sanchez-Garcia, M.; Tuberosa, R.; Bassi, F.M. Genomic Regions Associated with the Control of Flowering Time in Durum Wheat. Plants 2020, 9, 1628. [Google Scholar] [CrossRef]
- Royo, C.; Dreisigacker, S.; Ammar, K.; Villegas, D. Agronomic performance of durum wheat landraces and modern cultivars and its association with genotypic variation in vernalization response (Vrn-1) and photoperiod sensitivity (Ppd-1) genes. Eur. J. Agron. 2020, 120, 126129. [Google Scholar] [CrossRef]
- Bosse, M.; Spurgin, L.G.; Laine, V.N.; Cole, E.F.; Firth, J.A.; Gienapp, P.; Gosler, A.G.; McMahon, K.; Poissant, J.; Verhagen, I.; et al. Recent natural selection causes adaptive evolution of an avian polygenic trait. Science 2017, 6361, 365–368. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Chen, G.B.; Rasheed, A.; Li, D.; Sonder, K.; Zavala Espinosa, C.; Wang, J.; Costich, D.E.; Schnable, P.S.; Hearne, S.J.; et al. Identifying loci with breeding potential across temperate and tropical adaptation via EigenGWAS and EnvGWAS. Mol. Ecol. 2019, 28, 3544–3560. [Google Scholar] [CrossRef] [Green Version]
- Royo, C.; Soriano, J.M.; Rufo, R.; Guzmán, C. Are the agronomic performance and grain quality characteristics of bread wheat Mediterranean landraces related to the climate prevalent in their area of origin? J. Cereal Sci. 2022, 105, 103478. [Google Scholar] [CrossRef]
- Chardon, F.; Virlon, B.; Moreau, L.; Falque, M.; Joets, J.; Decousset, L.; Murigneux, A.; Charcosset, A. Genetic architecture of flowering time in maize as inferred from quantitative trait loci meta-analysis and synteny conservation with the rice genome. Genetics 2004, 168, 2169–2185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wong, D.; Forrest, K.; Allen, A.; Chao, S.; Huang, B.E.; Maccaferri, M.; Salvi, S.; Milner, S.G.; Cattivelli, L.; et al. Characterization of polyploid wheat genomic diversity using a high-density 90,000 single nucleotide polymorphism array. Plant Biotechnol. J. 2014, 12, 787–796. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Rasheed, A.; He, Z.; Imtiaz, M.; Arif, A.; Mahmood, T.; Ghafoor, A.; Wen, W.; Gao, F.; Xie, C.; et al. Genome-wide variation patterns between landraces and cultivars uncover divergent selection during modern wheat breeding. Theor. Appl. Genet. 2019, 132, 2509–2523. [Google Scholar] [CrossRef] [PubMed]
- Marcotuli, I.; Soriano, J.M.; Gadaleta, A. A consensus map for quality traits in durum wheat based on genome-wide association studies and detection of ortho-meta QTL across cereal species. Front. Genet. 2022, 13, 982418. [Google Scholar] [CrossRef] [PubMed]
- Gooding, M.J.; Ellis, R.H.; Shewry, P.R.; Schofield, J.D. Effects of restricted water availability and increased temperature on the grain filling, drying and quality of winter wheat. J. Cereal. Sci. 2003, 37, 295–309. [Google Scholar] [CrossRef]
- Tewolde, H.; Fernandez, C.J.; Erickson, C.A. Wheat cultivars adapted to postheading high temperatures. J. Agron. Crop Sci. 2006, 192, 111–120. [Google Scholar] [CrossRef]
- Arriagada, O.; Gadaleta, A.; Marcotuli, I.; Maccaferri, M.; Campana, M.; Reveco, S.; Alfaro, C.; Matus, I.; Schwember, A.R. A comprehensive meta-QTL analysis for yield-related traits of durum wheat (Triticum turgidum L. var. durum) grown under different water regimes. Front. Plant Sci. 2022, 13, 984269. [Google Scholar]
- Patterson, N.; Price, A.L.; Reich, D. Population Structure and Eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef]
- McVean, G. A genealogical interpretation of principal compnents analysis. PLoS Genet. 2009, 5, e1000686. [Google Scholar] [CrossRef] [Green Version]
- Bryc, K.; Bryc, W.; Silverstein, J.W. Separation of the largest eigenvalues in eigenanalysis of genotype data from discrete subpopulations. Theor. Popul. Biol. 2013, 89, 34–43. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.B.; Lee, S.H.; Zhur, Z.X.; Benyamin, B.; Robinson, M.R. EigenGWAS: Finding loci under selection through genome-wide association studies of eigenvectors in structured populations. Heredity 2016, 117, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Chen, Z.; Cheng, M.; Chen, J.; Zhu, T.T.; Wang, R.; Liu, Y.; Qi, P.; Chen, G.; Jiang, Q.; et al. Uncovering the dispersion history, adaptive evolution and selection of wheat in China. Plant Biotechnol. J. 2018, 16, 280–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IWGSC. Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 2018, 361, eaar7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef]
- Soriano, J.M.; Colasuonno, P.; Marcotuli, I.; Gadaleta, A. Meta-QTL analysis and identification of candidate genes for quality, abiotic and biotic stress in durum wheat. Sci. Rep. 2021, 11, 11877. [Google Scholar] [CrossRef]
- Chen, D.; He, L.; Lin, M.; Jing, Y.; Liang, C.; Liu, H.; Gao, J.; Zhang, W.; Wang, M. A ras-related small GTP-binding protein, RabE1c, regulates stomatal movements and drought stress responses by mediating the interaction with ABA receptors. Plant Sci. 2021, 306, 110858. [Google Scholar] [CrossRef]
- Djemal, R.; Khoudi, H. The ethylene-responsive transcription factor of durum wheat, TdSHN1, confers cadmium, copper, and zinc tolerance to yeast and transgenic tobacco plants. Protoplasma 2022, 259, 19–31. [Google Scholar] [CrossRef]
- Gao, T.; Li, G.Z.; Wang, C.R.; Dong, J.; Yuan, S.S.; Wang, Y.H.; Kang, G.Z. Function of the ERFL1a Transcription Factor in Wheat Responses to Water Deficiency. Int. J. Mol. Sci. 2018, 19, 1465. [Google Scholar] [CrossRef] [Green Version]
- Rufo, R.; López, A.; Lopes, M.S.; Bellvert, J.; Soriano, J.M. Identification of Quantitative Trait Loci Hotspots Affecting Agronomic Traits and High-Throughput Vegetation Indices in Rainfed Wheat. Front. Plant Sci. 2021, 12, 735192. [Google Scholar] [CrossRef]
- Singh, R.; Saripalli, G.; Gautam, T.; Kumar, A.; Jan, I.; Batra, R.; Kumar, J.; Kumar, R.; Balyan, H.S.; Sharma, S.; et al. Meta-QTL, ortho-MetaQTL and candidate genes for grain Fe and Zn contents in wheat (Triticum aestivum L.). Physiol. Mol. Biol. Plants. 2022, 28, 637–650. [Google Scholar] [CrossRef]
- Chen, X.; Wang, H.; Li, X.; Ma, K.; Zhan, Y.; Zeng, F. Molecular cloning and functional analysis of 4-Coumarate:CoA ligase 4 (4CL-like 1) from Fraxinus mandshurica and its role in abiotic stress tolerance and cell wall synthesis. BMC Plant Biol. 2019, 19, 231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Guo, Z.; Gu, F.; Ke, S.; Sun, D.; Dong, S.; Liu, W.; Huang, M.; Xiao, W.; Yang, G.; et al. Chen, Z. 4-Coumarate-CoA Ligase-Like Gene OsAAE3 Negatively Mediates the Rice Blast Resistance, Floret Development and Lignin Biosynthesis. Front. Plant Sci. 2017, 7, 2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Ward, J.H. Hierarchical grouping to optimize an objective function. J. Am. Stat. Assoc. 1963, 58, 236–244. [Google Scholar] [CrossRef]
- Royo, C.; Nazco, R.; Villegas, D. The climate of the zone of origin of Mediterranean durum wheat (Triticum durum Desf.) landraces affects their agronomic performance. Genet. Resour. Crop Evol. 2014, 61, 1345–1358. [Google Scholar] [CrossRef] [Green Version]
- Bradbury, P.J.; Zhang, Z.; Kroon, D.E.; Casstevens, R.E.; Ramdoss, Y.; Buckler, E.S. TASSEL: Software for association mapping of complex traits in diverse samples. Bioinformatics 2007, 23, 2633–2635. [Google Scholar] [CrossRef] [PubMed]
- Condorelli, G.E.; Maccaferri, M.; Newcomb, M.; Andrade-Sanchez, P.; White, J.W.; French, A.N.; Sciara, G.; Ward, R.; Tuberosa, R. Comparative Aerial and Ground Based High Throughput Phenotyping for the Genetic Dissection of NDVI as a Proxy for Drought Adaptive Traits in Durum Wheat. Front. Plant Sci. 2018, 9, 893. [Google Scholar] [CrossRef] [Green Version]
- Mangini, G.; Gadaleta, A.; Colasuonno, P.; Marcotuli, I.; Signorile, A.M.; Simeone, R.; De Vita, P.; Mastrangelo, A.M.; Laidò, G.; Pecchioni, N.; et al. Genetic dissection of the relationships between grain yield components by genome-wide association mapping in a collection of tetraploid wheats. PLoS ONE 2018, 13, e0190162. [Google Scholar] [CrossRef] [Green Version]
- Sukumaran, S.; Reynolds, M.P.; Sansaloni, C. Genome-Wide Association Analyses Identify QTL Hotspots for Yield and Component Traits in Durum Wheat Grown under Yield Potential, Drought, and Heat Stress Environments. Front. Plant Sci. 2018, 9, 81. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Guan, P.; Xin, M.; Wang, Y.; Chen, X.; Zhao, A.; Liu, M.; Li, H.; Zhang, M.; Lu, L.; et al. Genome wide association study identifies QTL for thousand grain weight in winter wheat under normal- and late-sown stressed environments. Theor. Appl. Genet. 2020, 134, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-González, R.H.; Borrill, P.; Lang, D.; Harrington, S.A.; Brinton, J.; Venturini, L.; Davey, M.; Jacobs, J.; van Ex, F.; Pasha, A.; et al. The transcriptional landscape of polyploid wheat. Science 2018, 361, aar6089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, W.H.; Tan, Y.C.; Yap, S.J.; Ng, K.P. ClicOFS: An interactive web based service of circos. Bioinformatics 2015, 31, 3685–3687. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | N | Tmax | Tmin | Tmean | Rh | Sunshine | Rad | ET0 | Rain | Tmax | Tmin | Tmean | Rh | Sunshine | Rad | ET0 | Rain |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sowing—Anthesis (SA) | Anthesis—Maturity (AM) | ||||||||||||||||
| Albania | 6 | 11.8 | 3.3 | 7.6 | 75.7 | 2.8 | 7.1 | 6.0 | 663.4 | 23.1 | 11.6 | 17.4 | 63.1 | 7.2 | 19.5 | 11.2 | 192.2 |
| Algeria | 26 | 16.1 | 6.1 | 11.1 | 75.4 | 5.4 | 11.0 | 200.0 | 193.4 | 26.1 | 13.6 | 19.9 | 60.7 | 8.6 | 22.1 | 385.2 | 78.2 |
| Bosnia & Herzegovina | 6 | 8.8 | 0.6 | 4.7 | 76.0 | 1.8 | 5.7 | 4.9 | 554.5 | 21.5 | 9.7 | 15.6 | 64.2 | 5.4 | 16.7 | 10.0 | 289.8 |
| Bulgaria | 6 | 7.5 | 0.5 | 4.0 | 81.7 | 3.0 | 6.9 | 111.2 | 180.8 | 20.6 | 11.0 | 15.8 | 74.0 | 7.2 | 19.3 | 290.3 | 162.8 |
| Croatia | 6 | 10.1 | 3.0 | 6.5 | 71.6 | 2.3 | 6.0 | 149.0 | 397.7 | 21.7 | 12.0 | 16.8 | 63.6 | 6.1 | 17.7 | 292.7 | 238.3 |
| Cyprus | 10 | 18.2 | 6.9 | 12.5 | 69.7 | 5.7 | 11.4 | 213.5 | 255.3 | 27.4 | 13.0 | 20.2 | 55.8 | 9.5 | 23.5 | 411.6 | 27.3 |
| Egypt | 14 | 21.9 | 9.8 | 15.8 | 67.5 | 7.1 | 14.3 | 333.7 | 30.6 | 31.0 | 16.5 | 23.8 | 49.9 | 9.9 | 24.4 | 541.0 | 2.2 |
| France | 24 | 11.4 | 3.5 | 7.4 | 78.3 | 3.3 | 6.9 | 162.0 | 255.9 | 20.8 | 10.4 | 15.6 | 66.8 | 7.1 | 18.9 | 335.6 | 161.8 |
| Greece | 12 | 14.4 | 6.0 | 10.2 | 74.2 | 3.2 | 8.0 | 196.1 | 307.8 | 24.0 | 13.4 | 18.7 | 64.0 | 6.1 | 16.5 | 313.8 | 82.5 |
| Israel | 10 | 18.7 | 8.4 | 13.5 | 72.0 | 6.8 | 13.2 | 292.1 | 442.9 | 27.8 | 14.1 | 20.9 | 56.2 | 10.9 | 25.7 | 542.4 | 31.2 |
| Italy | 24 | 14.0 | 6.8 | 10.4 | 75.7 | 3.6 | 7.9 | 180.2 | 282.7 | 22.5 | 13.2 | 17.9 | 68.6 | 7.8 | 20.3 | 335.6 | 104.2 |
| Jordan | 8 | 19.6 | 7.8 | 13.7 | 58.9 | 6.1 | 12.7 | 332.3 | 165.4 | 30.3 | 14.7 | 22.5 | 45.1 | 9.1 | 23.1 | 535.8 | 13.5 |
| Lebanon | 6 | 17.9 | 10.1 | 14.0 | 67.0 | 4.3 | 10.1 | 292.0 | 722.7 | 23.9 | 15.4 | 19.6 | 66.1 | 8.4 | 21.9 | 404.1 | 79.0 |
| Libya | 6 | 19.6 | 8.8 | 14.2 | 65.4 | 6.1 | 12.6 | 324.9 | 123.2 | 27.8 | 15.4 | 21.6 | 59.8 | 8.7 | 22.5 | 456.3 | 14.3 |
| Macedonia | 8 | 8.2 | −0.9 | 3.6 | 81.3 | 2.1 | 6.4 | 100.2 | 228.2 | 22.5 | 9.0 | 15.7 | 64.7 | 6.3 | 18.3 | 296.0 | 142.1 |
| Morocco | 40 | 18.9 | 7.2 | 13.0 | 69.8 | 5.9 | 12.1 | 262.7 | 227.6 | 25.1 | 12.2 | 18.6 | 66.1 | 8.7 | 22.4 | 364.9 | 84.0 |
| Portugal | 8 | 11.7 | 3.7 | 7.7 | 77.3 | 4.7 | 9.1 | 174.8 | 630.4 | 19.6 | 8.5 | 14.1 | 63.6 | 8.5 | 21.4 | 343.3 | 227.7 |
| Romania | 8 | 6.4 | −1.0 | 2.7 | 88.7 | 3.1 | 6.5 | 3.6 | 200.0 | 21.5 | 10.4 | 15.9 | 72.8 | 7.8 | 19.8 | 10.8 | 170.0 |
| Serbia | 8 | 7.7 | −0.1 | 3.8 | 79.7 | 1.8 | 5.8 | 96.0 | 215.2 | 21.8 | 10.3 | 16.1 | 65.7 | 5.7 | 17.2 | 278.1 | 216.0 |
| Spain | 22 | 13.0 | 4.0 | 8.5 | 77.5 | 3.9 | 8.4 | 163.3 | 226.4 | 23.0 | 10.5 | 16.8 | 58.5 | 7.8 | 20.4 | 357.9 | 119.0 |
| Syria | 22 | 15.4 | 4.4 | 9.9 | 71.7 | 5.1 | 10.6 | 212.8 | 178.8 | 29.7 | 14.7 | 22.2 | 44.1 | 9.2 | 23.0 | 580.2 | 38.8 |
| Tunisia | 10 | 17.9 | 8.3 | 13.1 | 72.7 | 5.6 | 11.1 | 244.6 | 182.5 | 26.0 | 14.7 | 20.3 | 67.5 | 8.5 | 21.9 | 393.9 | 45.6 |
| Turkey | 30 | 13.0 | 4.5 | 8.8 | 69.4 | 3.8 | 8.6 | 172.7 | 400.9 | 24.3 | 13.2 | 18.7 | 61.6 | 8.0 | 20.9 | 356.0 | 131.6 |
| Year (Y) | Climatic Zone (CZ) | Genetic Structure (GS) | ANOVA Interactions | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2016 | 2017 | 2018 | ANOVA | NB | NC | SW | SE | ANOVA | SP1 | SP2 | SP3 | ANOVA | Y x CZ | Y x GS | |
| D45 | 136.3 a | 145.0 b | 157.5 c | <0.0001 | 150.2 a | 147.7 a | 144.8 b | 143.0 b | <0.0001 | 144.8 b | 148.4 a | 146.2 ab | <0.0001 | 0.8669 | 0.8202 |
| D55 | 145.6 a | 153.8 b | 167.1 c | <0.0001 | 160.3 a | 157.7 a | 153.8 b | 150.3 b | <0.0001 | 153.9 b | 158.4 a | 154.3 b | <0.0001 | 0.9846 | 0.9617 |
| D65 | 151.8 a | 158.7 b | 172.2 c | <0.0001 | 165.2 a | 162.7 a | 159.4 b | 156.6 b | <0.0001 | 159.5 b | 163.4 a | 159.9 b | <0.0001 | 0.3626 | 0.5910 |
| D87 | 186.0 a | 187.8 b | 204.3 c | <0.0001 | 195.0 a | 194.0 a | 192.1 ab | 190.7 b | <0.0001 | 192.2 a | 194.3 a | 192.1 a | <0.0001 | <0.0001 | 0.0034 |
| DBA | 15.5 a | 13.4 b | 14.5 c | <0.0001 | 15.1 a | 14.8 a | 14.6 a | 13.6 b | 0.0001 | 14.6 a | 14.9 a | 13.8 b | 0.0001 | 0.1049 | 0.2895 |
| GFD | 34.0 a | 29.1 b | 32.1 c | <0.0001 | 34.0 a | 32.4 a | 30.7 b | 29.8 b | <0.0001 | 32.3 a | 30.5 b | 32.0 a | <0.0001 | <0.0001 | <0.0001 |
| Hotspot | Chr | CI (cM) | Trait | Left Marker | Position (bp) | Right Marker | Position (bp) | CI (Mb) | N Gene Models |
|---|---|---|---|---|---|---|---|---|---|
| hotspot 1A.1 | 1A | 21–27 | C,E,P | BS00023201_51 | 7643102 | Excalibur_c71158_54 | 8296998 | 0.65 | 19 |
| hotspot 1A.2 | 1A | 51–53 | P | Excalibur_c10689_254 | 27363007 | Kukri_c22508_119 | 28757949 | 1.39 | 9 |
| hotspot 1A.4 | 1A | 95–96 | C | BS00062876_51 | 529788778 | BobWhite_c96_170 | 531682571 | 1.89 | 34 |
| hotspot 1B.1 | 1B | 43–45 | C | BS00065053_51 | 38833829 | wsnp_Ex_c5780_10153638 | 26186242 | 12.65 | 89 |
| hotspot 1B.2 | 1B | 51–53 | P | Excalibur_c95656_129 | 44933589 | Tdurum_contig56188_569 | 28563880 | 16.37 | 114 |
| hotspot 1B.3 | 1B | 62–64 | C | wsnp_BE399980B_Ta_2_1 | 142523393 | BS00003575_51 | 148898343 | 6.37 | 27 |
| hotspot 1B.5 | 1B | 86–91 | C,E,P | Kukri_c25961_166 | 575863858 | BobWhite_c39656_106 | 589919646 | 14.06 | 147 |
| hotspot 1B.6 | 1B | 111–116 | C | BS00094237_51 | 638015155 | BS00084895_51 | 643101677 | 5.09 | 78 |
| hotspot 1B.7 | 1B | 1345–136 | P | GENE-0063_68 | 661515587 | Excalibur_rep_c71107_517 | 664599715 | 3.08 | 35 |
| hotspot 1B.8 | 1B | 159–161 | P | GENE-0543_201 | 681690469 | Excalibur_rep_c69522_83 | 685865389 | 4.17 | 41 |
| hotspot 1B.9 | 1B | 172–174 | P | wsnp_Ex_c1597_3045682 | 688283056 | wsnp_Ku_c13952_22097856 | 687413792 | 0.87 | 9 |
| hotspot 1D.1 | 1D | 161–172 | C | RAC875_c14613_68 | 485557589 | RFL_Contig3395_1575 | 487168787 | 1.61 | 34 |
| hotspot 1D.2 | 1D | 179–180 | C | BS00093275_51 | 486241852 | Tdurum_contig29915_167 | 491043383 | 4.80 | 75 |
| hotspot 2A.1 | 2A | 74–75 | P | Tdurum_contig11803_306 | 36041083 | Ku_c269_2643 | 36632073 | 0.59 | 15 |
| hotspot 2A.2 | 2A | 122–123 | C | BS00107804_51 | 707040172 | wsnp_Ex_rep_c66358_64543401 | 709701422 | 2.66 | 30 |
| hotspot 2A.3 | 2A | 143–144 | C | BS00062732_51 | 747090405 | Excalibur_c18514_238 | 750595232 | 3.50 | 90 |
| hotspot 2B.3 | 2B | 147–148 | C | BobWhite_c12911_788 | 780590397 | BS00100118_51 | 788524935 | 7.93 | 108 |
| hotspot 2D.1 | 2D | 8–9 | P | BS00067698_51 | 14860348 | BS00047901_51 | 15967448 | 1.11 | 30 |
| hotspot 3B.1 | 3B | 37–38 | C | Tdurum_contig43252_1762 | 23782080 | TA001028-0737 | 24007966 | 0.23 | 8 |
| hotspot 3B.3 | 3B | 67–69 | C,P | Ku_c27771_508 | 495471559 | BS00030430_51 | 503989868 | 8.52 | 52 |
| hotspot 4B.1 | 4B | 55–57 | C | Ra_c26080_461 | 36642697 | BS00095416_51 | 40233919 | 3.59 | 30 |
| hotspot 5A.2 | 5A | 84–86 | C | BS00073670_51 | 570716220 | Excalibur_c472_914 | 568272220 | 2.44 | 40 |
| hotspot 5A.3 | 5A | 90–92 | P | Kukri_c10033_724 | 584677742 | Excalibur_c26671_57 | 591319197 | 6.64 | 87 |
| hotspot 5A.4 | 5A | 115–116 | C | BS00076948_51 | 664273096 | Tdurum_contig11521_102 | 665779594 | 1.51 | 20 |
| hotspot 5B.2 | 5B | 60–61 | C | RAC875_c38511_91 | 476805518 | RAC875_c2437_1569 | 479025121 | 2.22 | 39 |
| hotspot 6B.1 | 6B | 39–40 | C | RAC875_c2291_123 | 41705928 | RAC875_c13920_836 | 42778537 | 1.07 | 23 |
| hotspot 6B.2 | 6B | 73–74 | E | BobWhite_c28409_462 | 635175311 | wsnp_Ex_c1276_2445537 | 642348416 | 7.17 | 39 |
| hotspot 6B.3 | 6B | 84–86 | P | Kukri_c58961_76 | 669019620 | Tdurum_contig68217_361 | 674946651 | 5.93 | 50 |
| hotspot 6B.4 | 6B | 120–121 | P | Kukri_c60966_261 | 719509426 | Tdurum_contig10729_989 | 720983865 | 1.47 | 25 |
| hotspot 7A.3 | 7A | 207–211 | E | BobWhite_c32347_219 | 708145137 | Ku_c19745_1093 | 712058458 | 3.91 | 74 |
| hotspot 7A.4 | 7A | 215–218 | E | BS00027226_51 | 717965474 | Kukri_c9728_1171 | 719567332 | 1.60 | 14 |
| hotspot 7A.5 | 7A | 227–229 | C,E | BS00020236_51 | 730426125 | Tdurum_contig46717_2021 | 731267973 | 0.84 | 18 |
| hotspot 7B.1 | 7B | 57–59 | C,P | GENE-4826_641 | 61557711 | BS00091302_51 | 64726430 | 3.17 | 31 |
| QTL Hotspot | Marker | Position (cM) | Position (bp) | Allele (Frecuency) | ||
|---|---|---|---|---|---|---|
| SP1 | SP2 | SP3 | ||||
| hotspot 2B.1 | Kukri_c35153_956 | 102.2 | 539,965,301 | A (54%) | A (25%) | A (91%) |
| G (46%) | G (75%) | G (9%) | ||||
| Kukri_c35153_145 | 104.4 | 539,964,058 | A (46%) | A (75%) | A (9%) | |
| G (54%) | G (25%) | G (91%) | ||||
| hotspot 5B.1 | wsnp_BE443187B_Ta_2_1 | 51.2 | 410,531,891 | A (4%) | A (25%) | A (77%) |
| C (96%) | C (75%) | C (23%) | ||||
| hotspot 5B.3 | RAC875_c19099_434 | 68.9 | 519,153,286 | T (92%) | T (67%) | T (18%) |
| C (8%) | C (29%) | C (82%) | ||||
| Ra_c73292_443 | 69.2 | 513,607,799 | A (8%) | A (46%) | A (55%) | |
| G (92%) | G (46%) | G (45%) | ||||
| hotspot 6B.2 | GENE-1074_108 | 73.4 | 633,906,287 | T (92%) | T (4%) | T (32%) |
| C (8%) | C (96%) | C (68%) | ||||
| GENE-4086_659 | 73.4 | - | A (4%) | A (50%) | A (36%) | |
| C (96%) | C (50%) | C (64%) | ||||
| GENE-4086_876 | 73.4 | 641,286,640 | A (4%) | A (46%) | A (36%) | |
| G (96%) | G (50%) | G (64%) | ||||
| Kukri_c59960_211 | 73.4 | 641,291,882 | T (4%) | T (46%) | T (36%) | |
| C (96%) | C (50%) | C (64%) | ||||
| hotspot 7A.4 | Excalibur_c46453_144 | 217.0 | 719,568,282 | A (19%) | A (50%) | A (59%) |
| G (81%) | G (46%) | G (41%) | ||||
| Kukri_c9728_1171 | 217.0 | 719,567,232 | A (81%) | A (46%) | A (41%) | |
| G (19%) | G (50%) | G (59%) | ||||
| hotspot 7B.2 | wsnp_Ex_c106_217340 | 76.0 | 538,868,953 | A (96%) | A (58%) | A (100%) |
| G (4%) | G (33%) | G (0%) | ||||
| Kukri_c51296_438 | 77.1 | 565,911,626 | A (15%) | A (96%) | A (91%) | |
| G (85%) | G (4%) | G (9%) | ||||
| hotspot 7B.2 | wsnp_Ex_c5270_9324025 | 77.1 | - | T (15%) | T (96%) | T (91%) |
| C (85%) | C (4%) | C (9%) | ||||
| hotspot 7B.2 | wsnp_RFL_Contig4753_5709032 | 77.1 | 566,481,254 | A (85%) | A (4%) | A (9%) |
| G (15%) | G (96%) | G (91%) | ||||
| Chromosome | Hotspot | Syntenic Chromosomes | ||||||
|---|---|---|---|---|---|---|---|---|
| 1A | hotspot 1A.1 | bd2 | os5 | |||||
| 1A | hotspot 1A.2 | bd2 | os5 | zm6 | ||||
| 1A | hotspot 1A.4 | bd2 | bd3 | os5 | ||||
| 1B | hotspot 1B.1-2 | bd1 | bd3 | os1 | os2 | os5 | os7 | os12 |
| 1B | hotspot 1B.3 | bd2 | ||||||
| 1B | hotspot 1B.5 | bd2 | bd3 | os1 | os5 | os6 | os7 | zm6 |
| 1B | hotspot 1B.6 | bd1 | bd2 | os5 | os6 | |||
| 1B | hotspot 1B.7 | bd2 | os5 | zm8 | ||||
| 1B | hotspot 1B.8 | bd2 | os4 | os5 | ||||
| 1B | hotspot 1B.9 | bd4 | os4 | os9 | ||||
| 1D | hotspot 1D.1-2 | bd1 | bd2 | os5 | ||||
| 2A | hotspot 2A.1 | os7 | ||||||
| 2A | hotspot 2A.3 | bd3 | os1 | os4 | ||||
| 2B | hotspot 2B.3 | bd2 | bd4 | bd5 | os4 | os9 | zm2 | zm8 |
| 2D | hotspot 2D.1 | bd4 | os11 | |||||
| 3B | hotspot 3B.3 | bd2 | os1 | os5 | ||||
| 5A | hotspot 5A.3 | bd1 | bd2 | os1 | os3 | |||
| 5A | hotspot 5A.4 | bd1 | os3 | zm1 | ||||
| 5B | hotspot 5B.2 | bd2 | bd4 | os1 | os9 | |||
| 6B | hotspot 6B.1 | bd3 | ||||||
| 6B | hotspot 6B.3 | bd4 | bd5 | |||||
| 6B | hotspot 6B.4 | bd3 | os2 | zm5 | ||||
| 7A | hotspot 7A.3 | bd1 | os2 | os6 | ||||
| 7A | hotspot 7A.5 | bd1 | os2 | os6 | zm6 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yannam, V.R.R.; Rufo, R.; Marcotuli, I.; Gadaleta, A.; Lopes, M.S.; Soriano, J.M. Discovering Loci for Breeding Prospective and Phenology in Wheat Mediterranean Landraces by Environmental and eigenGWAS. Int. J. Mol. Sci. 2023, 24, 1700. https://doi.org/10.3390/ijms24021700
Yannam VRR, Rufo R, Marcotuli I, Gadaleta A, Lopes MS, Soriano JM. Discovering Loci for Breeding Prospective and Phenology in Wheat Mediterranean Landraces by Environmental and eigenGWAS. International Journal of Molecular Sciences. 2023; 24(2):1700. https://doi.org/10.3390/ijms24021700
Chicago/Turabian StyleYannam, Venkata Rami Reddy, Rubén Rufo, Ilaria Marcotuli, Agata Gadaleta, Marta S. Lopes, and Jose Miguel Soriano. 2023. "Discovering Loci for Breeding Prospective and Phenology in Wheat Mediterranean Landraces by Environmental and eigenGWAS" International Journal of Molecular Sciences 24, no. 2: 1700. https://doi.org/10.3390/ijms24021700