

Molecular Mechanism of Cold Tolerance of Centipedegrass Based on the Transcriptome

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Physiological Changes in Centipedegrass under Low-Temperature Stress
2.2. RNA-Seq Results
2.3. Functional Annotation of Isoforms
2.4. Long Noncoding RNA Prediction and Transcription Factor Identification
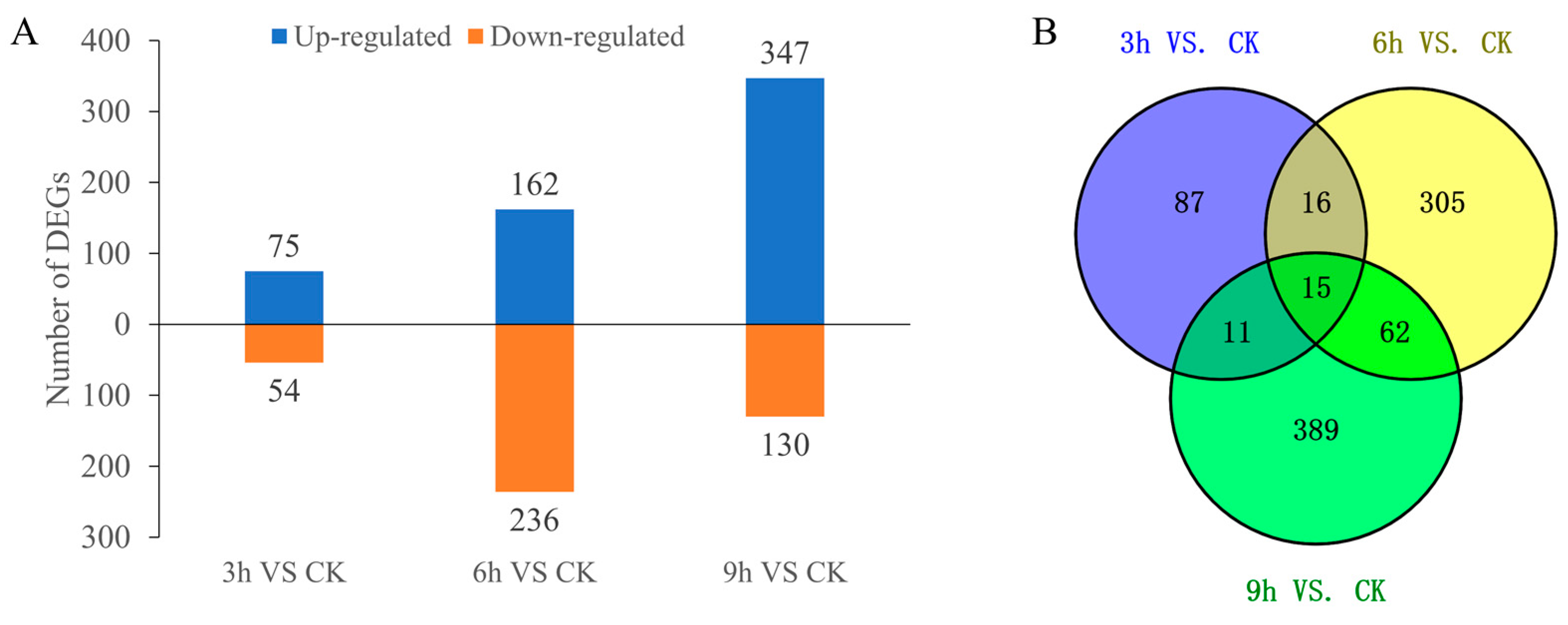
2.5. Differentially Expressed Genes (DEGs) in Response to Low-Temperature Stress
2.6. GO and KEGG Enrichment Analysis of DEGs
2.7. Arginine and Proline Metabolism
2.8. Flavonoid Biosynthesis
2.9. Plant Circadian Rhythm Pathway
2.10. Plant hormone Signal Transduction
2.11. Transcription Factors (TFs) and Transcriptional Regulation Factors (TRs)
2.12. Weighted Gene Coexpression Network Analysis (WGCNA)
2.13. qRT-PCR Validation of the DEGs
3. Discussion
3.1. Signal Transduction
3.2. Arginine Metabolism
3.3. Circadian Rhythm in Plants
3.4. Flavonoid Biosynthesis
3.5. Transcription Factors (TFs) and Transcriptional Regulation Factors (TRs)
4. Materials and Methods
4.1. Plant Materials and Low-Temperature Treatment
4.2. RNA Extraction, Library Construction and Sequencing
4.3. De Novo Assembly
4.4. Expression Calculation and Differential Expression Analysis
4.5. Functional Annotation of Transcripts
4.6. Quantitative Real-Time PCR (qRT–PCR) Analysis
4.7. WGCNA Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, J.; Guo, H.; Zong, J.; Chen, J.; Li, D.; Liu, J. Genetic diversity in centipedegrass [Eremochloa ophiuroides (Munro) Hack.]. Hortic. Res. 2020, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zi, H.; Wang, R.; Liu, J.; Wang, H.; Chen, R.; Li, L.; Guo, H.; Chen, J.; Li, J.; et al. A high-quality chromosome-scale assembly of the centipedegrass [Eremochloa ophiuroides (Munro) Hack.] genome provides insights into chromosomal structural evolution and prostrate growth habit. Hortic. Res. 2021, 8, 201. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Cui, R.; Xu, P.; Mao, J.; Chen, Y.; Zhou, C.; Yu, L.; Xiang, C. Arabidopsis ATHB17 Coordinates Nuclear and Plastidic Photosynthesis Gene Expression in Response to Abiotic Stress. bioRxiv 2016. bioRxiv 040501. [Google Scholar] [CrossRef] [Green Version]
- Zuther, E.; Lee, Y.P.; Erban, A.; Kopka, J.; Hincha, D.K. Natural Variation in Freezing Tolerance and Cold Acclimation Response in Arabidopsis thaliana and Related Species. In Survival Strategies in Extreme Cold and Desiccation; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1081, pp. 81–98. [Google Scholar] [CrossRef]
- Cretescu, I.; Caprita, R.; Velicevici, G.; Dorin, C.; Sirbu, C.; Buzamat, G.; Ropciuc, S. Researches regarding the influence of cold storage on the chlorophyll content in lettuce. Sci. Pap. Anim. Sci. Biotechnol. 2014, 47, 162–166. [Google Scholar]
- Yoon, Y.-E.; Kuppusamy, S.; Cho, K.M.; Kim, P.J.; Kwack, Y.-B.; Lee, Y.B. Influence of cold stress on contents of soluble sugars, vitamin C and free amino acids including gamma-aminobutyric acid (GABA) in spinach (Spinacia oleracea). Food Chem. 2017, 215, 185–192. [Google Scholar] [CrossRef]
- Shi, H.; Jiang, C.; Ye, T.; Tan, D.; Reiter, R.; Zhang, H.; Liu, R.; Chan, Z. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass [Cynodon dactylon (L). Pers.] by exogenous melatonin. J. Exp. Bot. 2015, 66, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Kidokoro, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant cold-stress responses. Trends Plant Sci. 2022, 27, 922–935. [Google Scholar] [CrossRef]
- Chen, L.; Fan, J.; Hu, L.; Hu, Z.; Xie, Y.; Zhang, Y.; Lou, Y.; Nevo, E.; Fu, J. A transcriptomic analysis of bermudagrass (Cynodon dactylon) provides novel insights into the basis of low temperature tolerance. BMC Plant Biol. 2015, 15, 216. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Zhao, Y.; Gu, J.; Zhou, M.; Gao, L.; Sun, R.-X.; Wang, W.-W.; Zhang, S.-H.; Yang, X.-J. Proteomic analysis reveals the molecular mechanism underlying the cold acclimation and freezing tolerance of wheat (Triticum aestivum L.). Plant Sci. 2022, 318, 111242. [Google Scholar] [CrossRef]
- Yang, D.-G.; Yang, F.-P.; Chen, X.-Q.; Zhang, L.-Q.; Zhang, X.-D. Producing Transformed Maize with Dehydration-Responsive Transcription Factor CBF4 Gene. Acta Agron. Sin. 2009, 35, 1759–1763. [Google Scholar] [CrossRef]
- Wei, S.; Du, Z.; Gao, F.; Ke, X.; Li, J.; Liu, J.; Zhou, Y. Global transcriptome profiles of ‘Meyer’ Zoysiagrass in response to cold stress. PLoS ONE 2015, 10, e0131153. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.-F.; He, X.; Yuan, D.-J.; Xu, L.; Xu, L.; Tu, L.-L.; Shen, G.-X.; Zhang, H.; Zhang, X.-L. Genome-wide identification of genes responsive to ABA and cold/salt stresses in Gossypium hirsutum by data-mining and expression pattern analysis. Agric. Sci. China 2011, 10, 499–508. [Google Scholar] [CrossRef]
- Lu, T.; Lu, G.; Fan, D.; Zhu, C.; Li, W.; Zhao, Q.; Feng, Q.; Zhao, Y.; Guo, Y.; Li, W.; et al. Function annotation of the rice transcriptome at single-nucleotide resolution by RNA-seq. Genome Res. 2010, 20, 1238–1249. [Google Scholar] [CrossRef] [Green Version]
- Sadat-Hosseini, M.; Bakhtiarizadeh, M.R.; Boroomand, N.; Tohidfar, M.; Vahdati, K. Combining independent de novo assemblies to optimize leaf transcriptome of Persian walnut. PLoS ONE 2020, 15, e0232005. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.; Kang, H.; Park, M.; Jeong, H.; Sun, H.; Song, P.; Lee, H. Zoysia japonica MYC type transcription factor ZjICE1 regulates cold tolerance in transgenic Arabidopsis. Plant Sci. 2019, 289, 110254. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, J. Lanthanum Promotes Bahiagrass (Paspalum notatum) Roots Growth by Improving Root Activity, Photosynthesis and Respiration. Plants 2022, 11, 382. [Google Scholar] [CrossRef]
- Hackl, T.; Hedrich, R.; Schultz, J.; Förster, F. Proovread: Large-scale high-accuracy PacBio correction through iterative short read consensus. Bioinformatics 2014, 30, 3004–3011. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Li, X.; Li, T.; Yu, D.; Han, B. Genome-wide transcriptome profiling provides overwintering mechanism of Agropyron mongolicum. BMC Plant Biol. 2017, 17, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Chen, X.; Ding, Y.; Yang, Y.; Song, C.; Wang, B.; Yang, S.; Guo, Y.; Gong, Z. Protein kinases in plant responses to drought, salt, and cold stress. J. Integr. Plant Biol. 2021, 63, 53–78. [Google Scholar] [CrossRef] [PubMed]
- Perochon, A.; Dieterle, S.; Pouzet, C.; Aldon, D.; Galaud, J.; Ranty, B. Interaction of a plant pseudo-response regulator with a calmodulin-like protein. Biochem. Biophys. Res. Commun. 2010, 398, 747–751. [Google Scholar] [CrossRef] [PubMed]
- Almadanim, M.; Alexandre, B.; Rosa, M.; Sapeta, H.; Leitão, A.; Ramalho, J.; Lam, T.; Negrão, S.; Abreu, I.; Oliveira, M. Rice calcium-dependent protein kinase OsCPK17 targets plasma membrane intrinsic protein and sucrose-phosphate synthase and is required for a proper cold stress response. Plant Cell Environ. 2017, 40, 1197–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Zheng, T.; Li, L.; Zhuo, X.; Jiang, L.; Wang, J.; Cheng, T.; Zhang, Q. Identification and comparative analysis of the CIPK gene family and characterization of the cold stress response in the woody plant Prunus mume. PeerJ 2019, 7, e6847. [Google Scholar] [CrossRef] [Green Version]
- Kolukisaoglu, U.; Weinl, S.; Blazevic, D.; Batistic, O.; Kudla, J. Calcium sensors and their interacting protein kinases: Genomics of the Arabidopsis and rice CBL-CIPK signaling networks. Plant Physiol. 2004, 134, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Yang, B.; Liu, W.; Li, H.; Wang, L.; Wang, B.; Deng, M.; Liang, W.; Deyholos, M.; Jiang, Y. Identification and characterization of CBL and CIPK gene families in canola (Brassica napus L.). BMC Plant Biol. 2014, 14, 8. [Google Scholar] [CrossRef] [Green Version]
- Han, Q.Q.; Huang, H.J.; Wang, Y.L.; Yang, L.; Pilot, A.; Zhu, X.C.; Yu, R.; Wang, J.; Chen, X.R.; Liu, Q.; et al. Ghrelin exhibited antidepressant and anxiolytic effect via the p38-MAPK signaling pathway in hippocampus. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 93, 11–20. [Google Scholar] [CrossRef]
- Ning, J.; Li, X.; Hicks, L.M.; Xiong, L. A Raf-Like MAPKKK Gene DSM1 Mediates Drought Resistance through Reactive Oxygen Species Scavenging in Rice1[C][W][OA]. Plant Physiol. 2010, 152, 876–890. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Wang, P.; Si, T.; Hsu, C.-C.; Wang, L.; Zayed, O.; Yu, Z.; Zhu, Y.; Dong, J.; Tao, W.A.; et al. MAP Kinase Cascades Regulate the Cold Response by Modulating ICE1 Protein Stability. Dev. Cell 2017, 43, 618–629.e615. [Google Scholar] [CrossRef] [Green Version]
- Jia, H.; Hao, L.; Guo, X.; Liu, S.; Yan, Y.; Guo, X. A Raf-like MAPKKK gene, GhRaf19, negatively regulates tolerance to drought and salt and positively regulates resistance to cold stress by modulating reactive oxygen species in cotton. Plant Sci. 2016, 252, 267–281. [Google Scholar] [CrossRef]
- Furihata, T.; Maruyama, K.; Fujita, Y.; Umezawa, T.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Abscisic acid-dependent multisite phosphorylation regulates the activity of a transcription activator AREB1. Proc. Natl. Acad. Sci. USA 2006, 103, 1988–1993. [Google Scholar] [CrossRef] [Green Version]
- Lin, Q.; Wu, F.; Sheng, P.; Zhang, Z.; Zhang, X.; Guo, X.; Wang, J.; Cheng, Z.; Wang, J.; Wang, H.; et al. The SnRK2-APC/C(TE) regulatory module mediates the antagonistic action of gibberellic acid and abscisic acid pathways. Nat. Commun. 2015, 6, 7981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslam, M.; Sugita, K.; Qin, Y.; Rahman, A. ArabidopsisAux/IAA14 Regulates microRNA-Mediated Cold Stress Response in Arabidopsis Roots. Int. J. Mol. Sci. 2020, 21, 8441. [Google Scholar] [CrossRef] [PubMed]
- Weijers, D.; Wagner, D. Transcriptional Responses to the Auxin Hormone. Annu. Rev. Plant Biol. 2016, 67, 539–574. [Google Scholar] [CrossRef] [PubMed]
- Alisoltani, A.; Karimi, M.; Ravash, R.; Fallahi, H.; Shiran, B. Molecular Responses to Cold Stress in Temperate Fruit Crops with Focus on Rosaceae Family. In Genomics Assisted Breeding of Crops for Abiotic Stress Tolerance; Springer: Berlin/Heidelberg, Germany, 2019; Volume 2, pp. 105–130. [Google Scholar] [CrossRef]
- Ritonga, F.N.; Chen, S. Physiological and molecular mechanism involved in cold stress tolerance in plants. Plants 2020, 9, 560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Shen, L.; Li, F.; Zhang, Y.; Meng, D.; Sheng, J. Up-regulating arginase contributes to amelioration of chilling stress and the antioxidant system in cherry tomato fruits. J. Sci. Food Agric. 2010, 90, 2195–2202. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; He, M.; Guo, S.; Zhong, M.; Shu, S.; Sun, J. NaCl stress induces CsSAMs gene expression in Cucumis sativus by mediating the binding of CsGT-3b to the GT-1 element within the CsSAMs promoter. Planta 2017, 245, 889–908. [Google Scholar] [CrossRef]
- Yu, Y.; Jin, C.; Sun, C.; Wang, J.; Ye, Y.; Lu, L.; Lin, X. Elevation of arginine decarboxylase-dependent putrescine production enhances aluminum tolerance by decreasing aluminum retention in root cell walls of wheat. J. Hazard. Mater. 2015, 299, 280–288. [Google Scholar] [CrossRef]
- Espinoza, C.; Bieniawska, Z.; Hincha, D.; Hannah, M. Interactions between the circadian clock and cold-response in Arabidopsis. Plant Signal. Behav. 2008, 3, 593–594. [Google Scholar] [CrossRef]
- Fu, J.; Miao, Y.; Shao, L.; Hu, T.; Yang, P. De novo transcriptome sequencing and gene expression profiling of Elymus nutans under cold stress. BMC Genom. 2016, 17, 870. [Google Scholar] [CrossRef] [Green Version]
- Abeynayake, S.; Byrne, S.; Nagy, I.; Jonavičienė, K.; Etzerodt, T.; Boelt, B.; Asp, T. Changes in Lolium perenne transcriptome during cold acclimation in two genotypes adapted to different climatic conditions. BMC Plant Biol. 2015, 15, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.L.; Newton, L.; Liu, M.-J.; Shiu, S.-H.; Farre, E.M. A G-Box-Like Motif Is Necessary for Transcriptional Regulation by Circadian Pseudo-Response Regulators in Arabidopsis. Plant Physiol. 2016, 170, 528–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmer, S.; Hogenesch, J.; Straume, M.; Chang, H.; Han, B.; Zhu, T.; Wang, X.; Kreps, J.; Kay, S. Orchestrated transcription of key pathways in Arabidopsis by the circadian clock. Science 2000, 290, 2110–2113. [Google Scholar] [CrossRef]
- Wheeler, L.; Walker, J.; Ng, J.; Deanna, R.; Dunbar-Wallis, A.; Backes, A.; Pezzi, P.; Palchetti, M.; Robertson, H.; Monaghan, A.; et al. Transcription Factors Evolve Faster Than Their Structural Gene Targets in the Flavonoid Pigment Pathway. Mol. Biol. Evol. 2022, 39, msac044. [Google Scholar] [CrossRef] [PubMed]
- Kejík, Z.; Kaplánek, R.; Masařík, M.; Babula, P.; Matkowski, A.; Filipenský, P.; Veselá, K.; Gburek, J.; Sýkora, D.; Martásek, P.; et al. Iron Complexes of Flavonoids-Antioxidant Capacity and Beyond. Int. J. Mol. Sci. 2021, 22, 646. [Google Scholar] [CrossRef]
- Schulz, E.; Tohge, T.; Zuther, E.; Fernie, A.; Hincha, D. Natural variation in flavonol and anthocyanin metabolism during cold acclimation in Arabidopsis thaliana accessions. Plant Cell Environ. 2015, 38, 1658–1672. [Google Scholar] [CrossRef]
- Matus, J.; Poupin, M.; Cañón, P.; Bordeu, E.; Alcalde, J.; Arce-Johnson, P. Isolation of WDR and bHLH genes related to flavonoid synthesis in grapevine (Vitis vinifera L.). Plant Mol. Biol. 2010, 72, 607–620. [Google Scholar] [CrossRef]
- Yarra, R.; Wei, W. The NAC-type transcription factor GmNAC20 improves cold, salinity tolerance, and lateral root formation in transgenic rice plants. Funct. Integr. Genom. 2021, 21, 473–487. [Google Scholar] [CrossRef]
- Joshi, R.; Wani, S.H.; Singh, B.; Bohra, A.; Dar, Z.A.; Lone, A.A.; Pareek, A.; Singla-Pareek, S.L. Transcription factors and plants response to drought stress: Current understanding and future directions. Front. Plant Sci. 2016, 7, 1029. [Google Scholar] [CrossRef] [PubMed]
- Han, D.; Du, M.; Zhou, Z.; Wang, S.; Li, T.; Han, J.; Xu, T.; Yang, G. An NAC transcription factor gene from Malus baccata, MbNAC29, increases cold and high salinity tolerance in Arabidopsis. In Vitr. Cell. Dev. Biol. Plant 2020, 56, 588–599. [Google Scholar] [CrossRef]
- Pandey, A.; Khan, M.K.; Hamurcu, M.; Brestic, M.; Topal, A.; Gezgin, S. Insight into the Root Transcriptome of a Boron-Tolerant Triticum zhukovskyi Genotype Grown under Boron Toxicity. Agronomy 2022, 12, 2421. [Google Scholar] [CrossRef]
- Sun, Q.; Wang, S.; Xu, G.; Kang, X.; Zhang, M.; Ni, M. SHB1 and CCA1 interaction desensitizes light responses and enhances thermomorphogenesis. Nat. Commun. 2019, 10, 3110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.-Y.; Qi, C.-H.; Jiang, H.; You, C.-X.; Guan, Q.-M.; Ma, F.-W.; Li, Y.-Y.; Hao, Y.-J. The MdWRKY31 transcription factor binds to the MdRAV1 promoter to mediate ABA sensitivity. Hortic. Res. 2019, 6, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Chung, W.S.; Lim, C.O. Overexpression of heat shock factor gene HsfA3 increases galactinol levels and oxidative stress tolerance in Arabidopsis. Mol. Cells 2016, 39, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [Green Version]
- Love, M.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Benaim, A.R. Controlling the False Discovery Rate for Multiple Testing Involved in the Statistical Analysis of Gene Expression Data; University of Tel-Aviv: Tel Aviv, Israel, 2006. [Google Scholar]
- Young, M.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Minoru, K.; Michihiro, A.; Susumu, G.; Masahiro, H.; Mika, H.; Masumi, I.; Toshiaki, K.; Shuichi, K.; Shujiro, O.; Toshiaki, T. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | TML |
|---|---|
| Subreads base (G) | 38.76 |
| Subreads number | 16,244,578 |
| Average subreads length | 2386 |
| CCS | 331,513 |
| FLNC | 281,066 |
| Average FLNC read length | 2652 |
| FLNC/CCS | 0.84 |
| Total nucleotides | 349,159,670 |
| Total number | 127,142 |
| Mean length | 2746 |
| Max length | 13,081 |
| N50 | 3089 |
| N90 | 1773 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Xiong, Y.; Zhao, J.; Bai, S.; Li, D.; Chen, L.; Feng, J.; Li, Y.; Ma, X.; Zhang, J. Molecular Mechanism of Cold Tolerance of Centipedegrass Based on the Transcriptome. Int. J. Mol. Sci. 2023, 24, 1265. https://doi.org/10.3390/ijms24021265
Liu Y, Xiong Y, Zhao J, Bai S, Li D, Chen L, Feng J, Li Y, Ma X, Zhang J. Molecular Mechanism of Cold Tolerance of Centipedegrass Based on the Transcriptome. International Journal of Molecular Sciences. 2023; 24(2):1265. https://doi.org/10.3390/ijms24021265
Chicago/Turabian StyleLiu, Yingjie, Yi Xiong, Junming Zhao, Shiqie Bai, Daxu Li, Limin Chen, Junjie Feng, Yingzhu Li, Xiao Ma, and Jianbo Zhang. 2023. "Molecular Mechanism of Cold Tolerance of Centipedegrass Based on the Transcriptome" International Journal of Molecular Sciences 24, no. 2: 1265. https://doi.org/10.3390/ijms24021265