
Identification of NCAPG as an Essential Gene for Neuroblastoma Employing CRISPR-Cas9 Screening Database and Experimental Verification
Abstract
:1. Introduction
2. Results
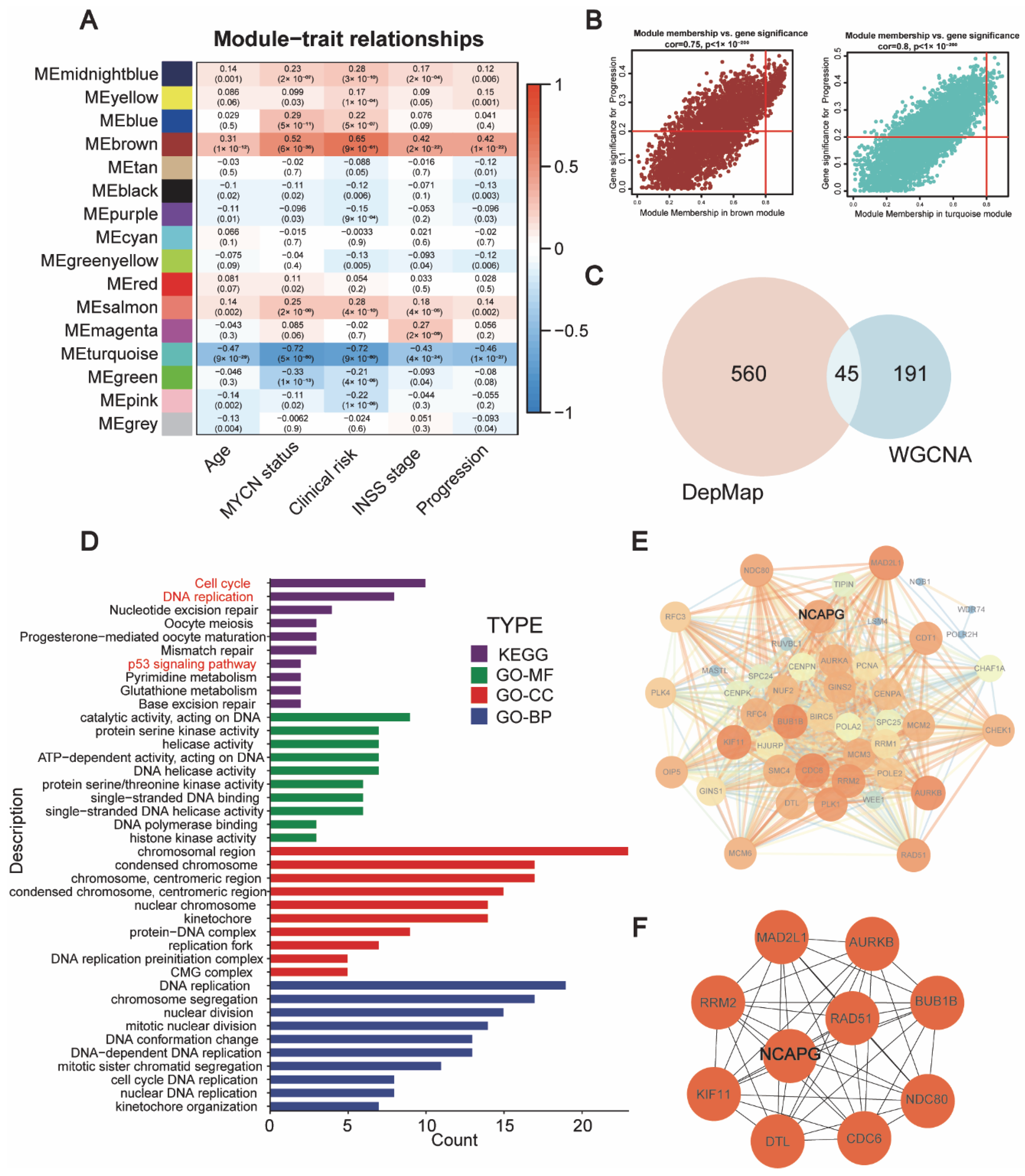
2.1. Identification of Candidate Genes That Were Crucial for Neuroblastoma Survival
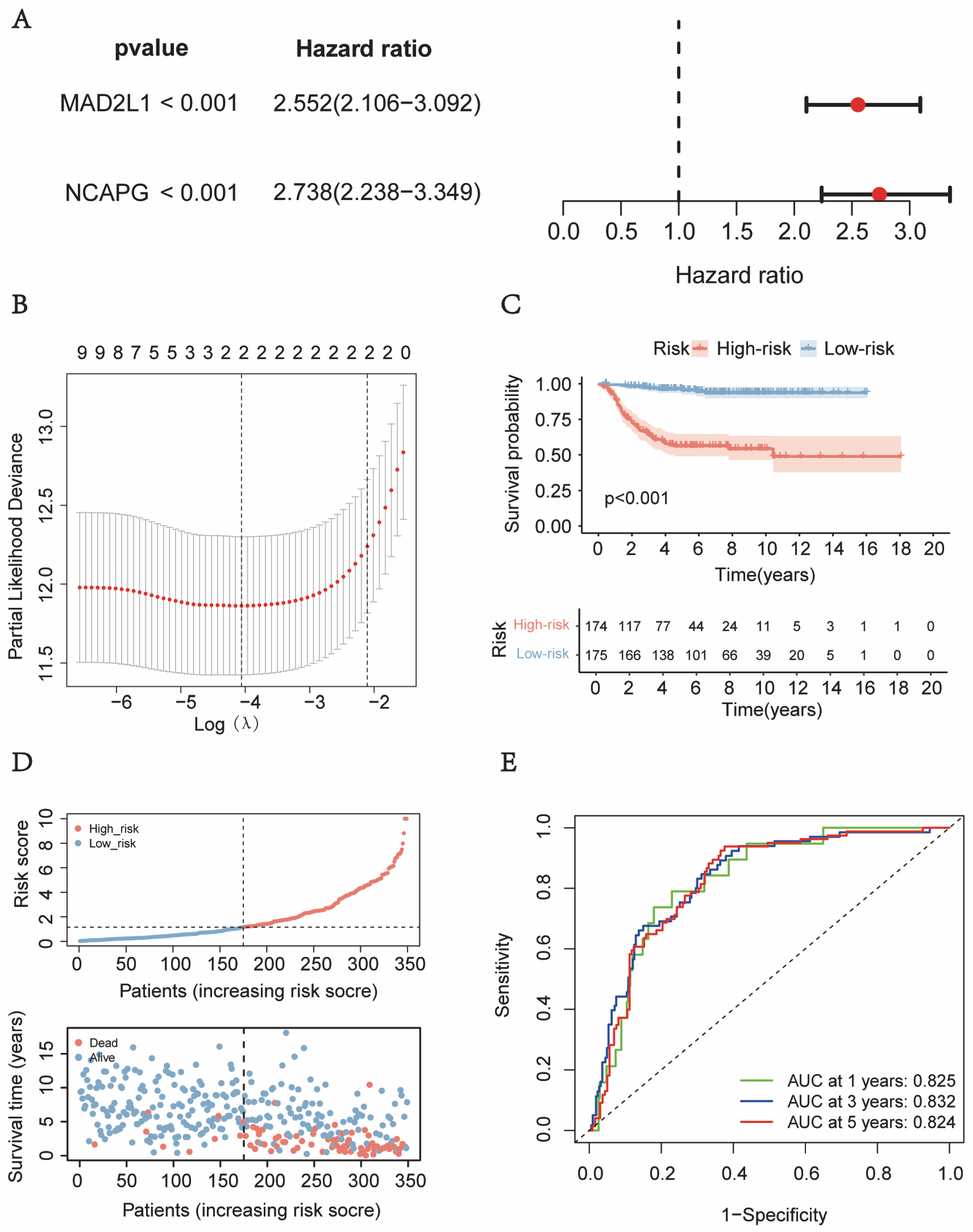
2.2. The Development and Verification of a Gene-Based Prognostic Signature
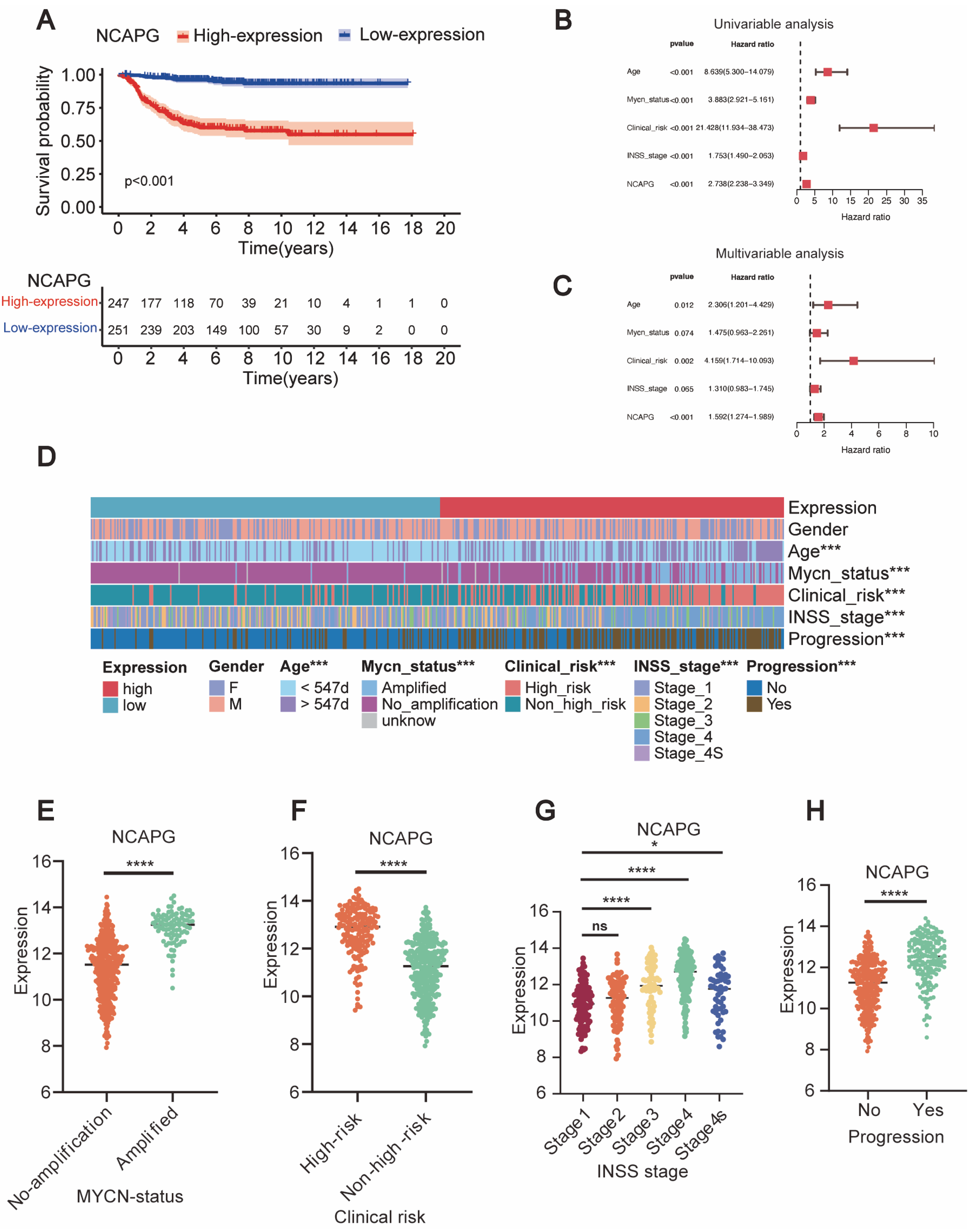
2.3. NCAPG Expression Correlated with Clinical Outcomes and Clinical Characteristics
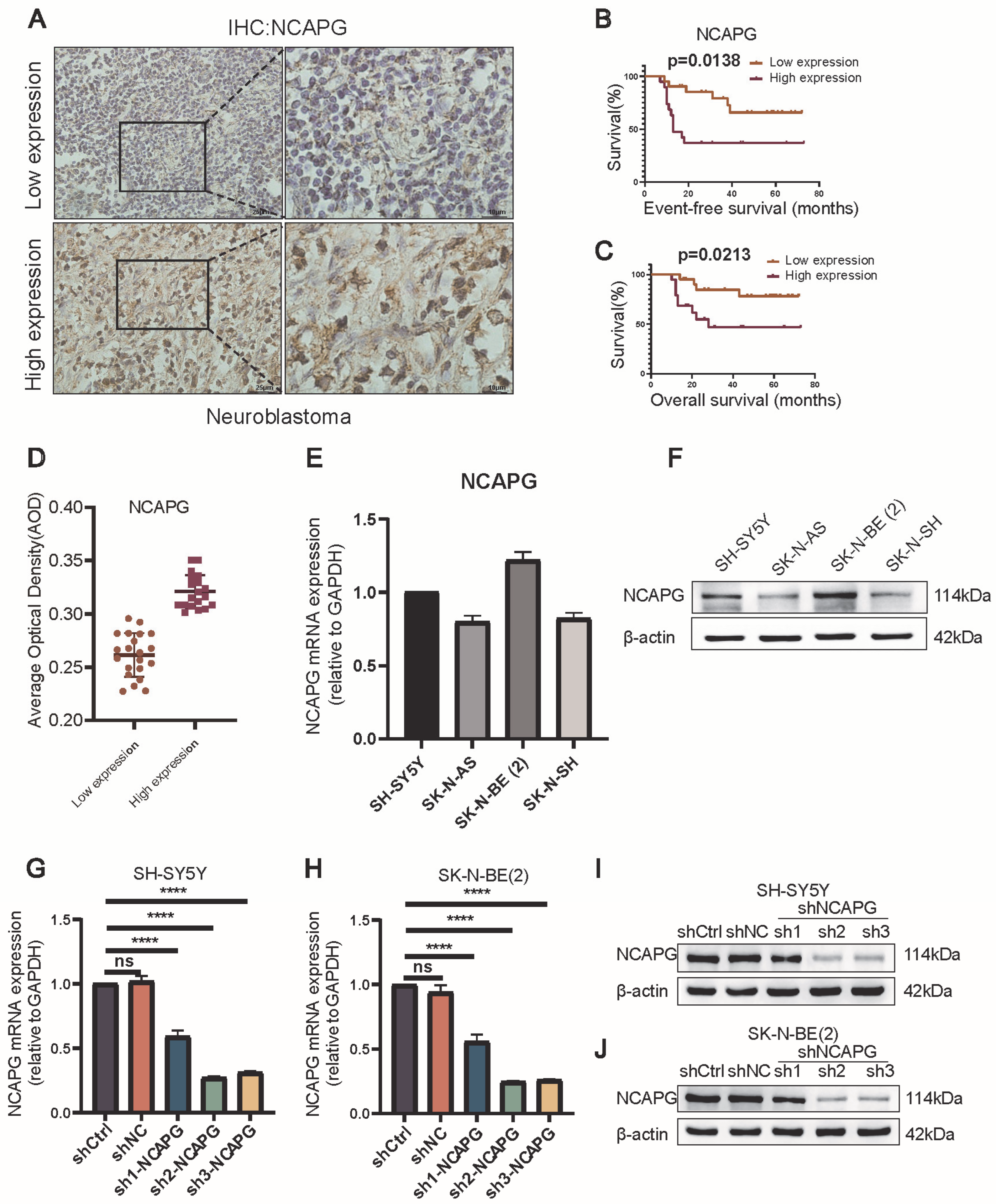
2.4. Neuroblastoma Tissues Expressed High Levels of NCAPG
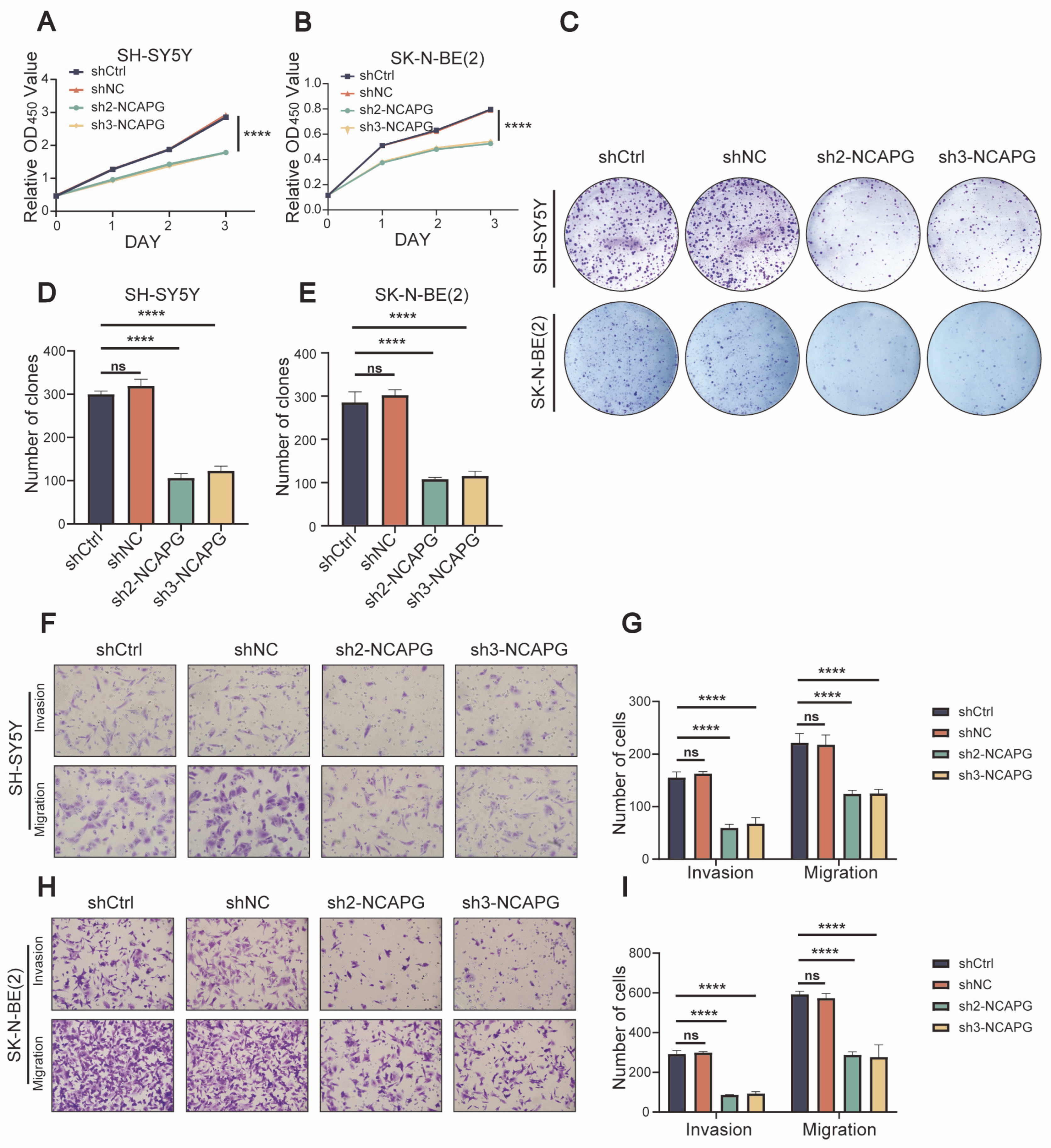
2.5. NCAPG Regulated Neuroblastoma Cells Proliferation, Migration, and Invasion
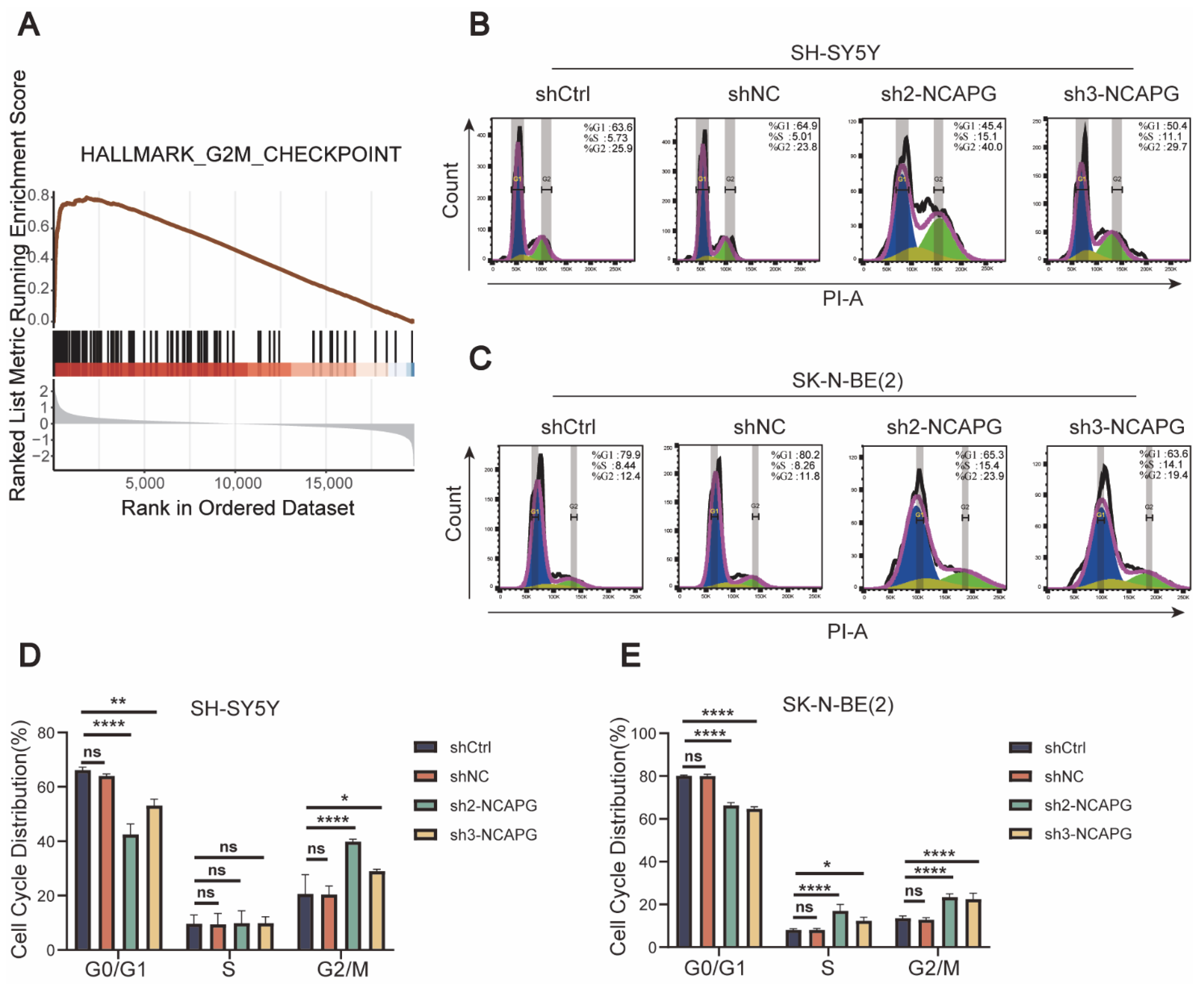
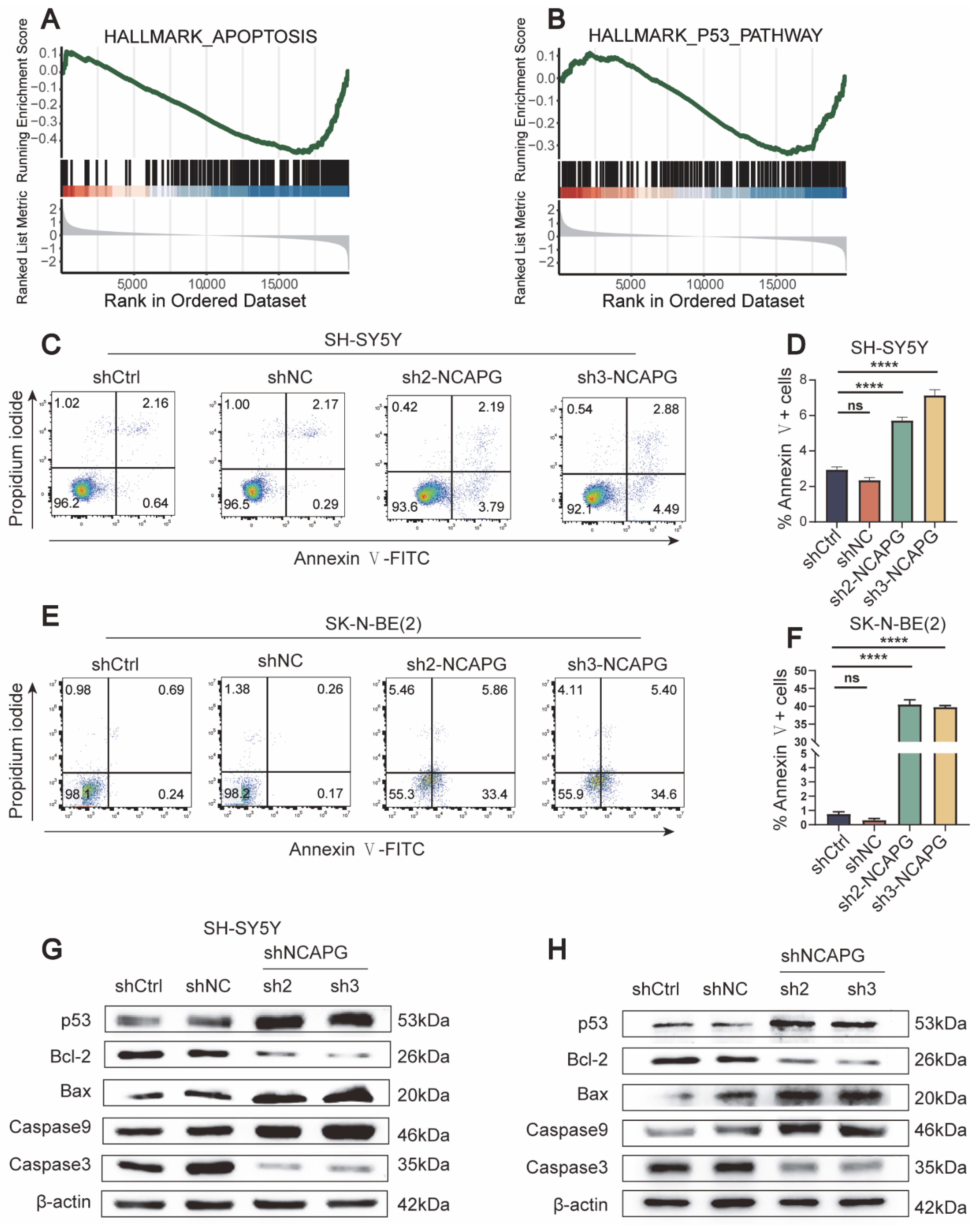
2.6. NCAPG Knockdown Caused Cell Cycle Arrest and Apoptosis in Neuroblastoma Cells
3. Discussion
4. Materials and Methods
4.1. The Identification of Essential Genes for Neuroblastoma Cell Survival Based on the Large-Scale CRISPR-Cas9 Screening Database
4.2. Acquisition of Data
4.3. Weight Gene Coexpression Network Analysis
4.4. Enrichment Analysis
4.5. Construction of the PPI Network and Hub Gene Identification
4.6. Development and Verification of the Prognostic Risk Signature
4.7. Clinical Samples
4.8. Immunohistochemistry (IHC)
4.9. Cell Lines and Cultures
4.10. Lentivirus Transduction
4.11. Western Blotting
4.12. Quantitative Real-Time PCR
4.13. Cell Counting Kit 8 Assay
4.14. Colony Formation Assay
4.15. Transwell Assay
4.16. Flow Cytometry
4.17. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maris, J.M.; Hogarty, M.D.; Bagatell, R.; Cohn, S.L. Neuroblastoma. Lancet 2007, 369, 2106–2120. [Google Scholar] [CrossRef] [PubMed]
- Matthay, K.K.; Maris, J.M.; Schleiermacher, G.; Nakagawara, A.; Mackall, C.L.; Diller, L.; Weiss, W.A. Neuroblastoma. Nat. Rev. Dis. Prim. 2016, 2, 16078. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.S.; Naranjo, A.; Zhang, F.F.; Cohn, S.L.; London, W.B.; Gastier-Foster, J.M.; Ramirez, N.C.; Pfau, R.; Reshmi, S.; Wagner, E.; et al. Revised Neuroblastoma Risk Classification System: A Report From the Children’s Oncology Group. J. Clin. Oncol. 2021, 39, 3229–3241. [Google Scholar] [CrossRef]
- Qiu, B.; Matthay, K.K. Advancing therapy for neuroblastoma. Nat. Rev. Clin. Oncol. 2022, 19, 515–533. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.; Wang, W.; Liu, G.; Xian, W.; McKeon, F.; Zhou, J.; Zhang, R. Targeting the p53-MDM2 pathway for neuroblastoma therapy: Rays of hope. Cancer Lett. 2021, 496, 16–29. [Google Scholar] [CrossRef] [PubMed]
- Zafar, A.; Wang, W.; Liu, G.; Wang, X.; Xian, W.; McKeon, F.; Foster, J.; Zhou, J.; Zhang, R. Molecular targeting therapies for neuroblastoma: Progress and challenges. Med. Res. Rev. 2021, 41, 961–1021. [Google Scholar] [CrossRef]
- Doudna, J.A.; Charpentier, E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Kurata, M.; Yamamoto, K.; Moriarity, B.S.; Kitagawa, M.; Largaespada, D.A. CRISPR/Cas9 library screening for drug target discovery. J. Hum. Genet. 2018, 63, 179–186. [Google Scholar] [CrossRef]
- Tsherniak, A.; Vazquez, F.; Montgomery, P.G.; Weir, B.A.; Kryukov, G.; Cowley, G.S.; Gill, S.; Harrington, W.F.; Pantel, S.; Krill-Burger, J.M.; et al. Defining a Cancer Dependency Map. Cell 2017, 170, 564–576.e16. [Google Scholar] [CrossRef]
- Ghandi, M.; Huang, F.W.; Jané-Valbuena, J.; Kryukov, G.V.; Lo, C.C.; McDonald, E.R., 3rd; Barretina, J.; Gelfand, E.T.; Bielski, C.M.; Li, H.; et al. Next-generation characterization of the Cancer Cell Line Encyclopedia. Nature 2019, 569, 503–508. [Google Scholar] [CrossRef]
- Corsello, S.; Nagari, R.; Spangler, R.; Rossen, J.; Kocak, M.; Bryan, J.; Humeidi, R.; Peck, D.; Wu, X.; Tang, A.; et al. Non-oncology drugs are a source of previously unappreciated anti-cancer activity. BioRxiv 2019. BioRxiv:730119. [Google Scholar]
- Meyers, R.M.; Bryan, J.G.; McFarland, J.M.; Weir, B.A.; Sizemore, A.E.; Xu, H.; Dharia, N.V.; Montgomery, P.G.; Cowley, G.S.; Pantel, S.; et al. Computational correction of copy number effect improves specificity of CRISPR-Cas9 essentiality screens in cancer cells. Nat. Genet. 2017, 49, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Sutani, T.; Sakata, T.; Nakato, R.; Masuda, K.; Ishibashi, M.; Yamashita, D.; Suzuki, Y.; Hirano, T.; Bando, M.; Shirahige, K. Condensin targets and reduces unwound DNA structures associated with transcription in mitotic chromosome condensation. Nat. Commun. 2015, 6, 7815. [Google Scholar] [CrossRef] [PubMed]
- Eberlein, A.; Takasuga, A.; Setoguchi, K.; Pfuhl, R.; Flisikowski, K.; Fries, R.; Klopp, N.; Fürbass, R.; Weikard, R.; Kühn, C. Dissection of genetic factors modulating fetal growth in cattle indicates a substantial role of the non-SMC condensin I complex, subunit G (NCAPG) gene. Genetics 2009, 183, 951–964. [Google Scholar] [CrossRef]
- Xiao, C.; Gong, J.; Jie, Y.; Cao, J.; Chen, Z.; Li, R.; Chong, Y.; Hu, B.; Zhang, Q. NCAPG Is a Promising Therapeutic Target Across Different Tumor Types. Front. Pharmacol. 2020, 11, 387. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, B.; Tan, P.Y.; Handoko, Y.A.; Sekar, K.; Deivasigamani, A.; Seshachalam, V.P.; OuYang, H.Y.; Shi, M.; Xie, C.; et al. Genome-wide CRISPR knockout screens identify NCAPG as an essential oncogene for hepatocellular carcinoma tumor growth. Faseb J. 2019, 33, 8759–8770. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Su, R.; Shan, C.; Gao, C.; Wu, P. Non-SMC Condensin I Complex, Subunit G (NCAPG) is a Novel Mitotic Gene Required for Hepatocellular Cancer Cell Proliferation and Migration. Oncol. Res. 2018, 26, 269–276. [Google Scholar] [CrossRef]
- Arai, T.; Okato, A.; Yamada, Y.; Sugawara, S.; Kurozumi, A.; Kojima, S.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Regulation of NCAPG by miR-99a-3p (passenger strand) inhibits cancer cell aggressiveness and is involved in CRPC. Cancer Med. 2018, 7, 1988–2002. [Google Scholar] [CrossRef]
- Jiang, L.; Ren, L.; Chen, H.; Pan, J.; Zhang, Z.; Kuang, X.; Chen, X.; Bao, W.; Lin, C.; Zhou, Z.; et al. NCAPG confers trastuzumab resistance via activating SRC/STAT3 signaling pathway in HER2-positive breast cancer. Cell Death Dis. 2020, 11, 547. [Google Scholar] [CrossRef]
- Sun, D.P.; Lin, C.C.; Hung, S.T.; Kuang, Y.Y.; Hseu, Y.C.; Fang, C.L.; Lin, K.Y. Aberrant Expression of NCAPG is Associated with Prognosis and Progression of Gastric Cancer. Cancer Manag. Res. 2020, 12, 7837–7846. [Google Scholar] [CrossRef]
- Zhang, W.; Yu, Y.; Hertwig, F.; Thierry-Mieg, J.; Zhang, W.; Thierry-Mieg, D.; Wang, J.; Furlanello, C.; Devanarayan, V.; Cheng, J.; et al. Comparison of RNA-seq and microarray-based models for clinical endpoint prediction. Genome Biol. 2015, 16, 133. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Bader, G.D.; Hogue, C.W. An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinform. 2003, 4, 2. [Google Scholar] [CrossRef] [PubMed]
- Tibshirani, R. The lasso method for variable selection in the Cox model. Stat. Med. 1997, 16, 385–395. [Google Scholar] [CrossRef]
- Maris, J.M. Recent advances in neuroblastoma. N. Engl. J. Med. 2010, 362, 2202–2211. [Google Scholar] [CrossRef]
- Pugh, T.J.; Morozova, O.; Attiyeh, E.F.; Asgharzadeh, S.; Wei, J.S.; Auclair, D.; Carter, S.L.; Cibulskis, K.; Hanna, M.; Kiezun, A.; et al. The genetic landscape of high-risk neuroblastoma. Nat. Genet. 2013, 45, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Schleiermacher, G.; Janoueix-Lerosey, I.; Delattre, O. Recent insights into the biology of neuroblastoma. Int. J. Cancer. 2014, 135, 2249–2261. [Google Scholar] [CrossRef]
- Schulte, J.H.; Eggert, A. Neuroblastoma. Crit. Rev. Oncog. 2015, 20, 245–270. [Google Scholar] [CrossRef]
- Park, J.R.; Bagatell, R.; London, W.B.; Maris, J.M.; Cohn, S.L.; Mattay, K.K.; Hogarty, M. Children’s Oncology Group’s 2013 blueprint for research: Neuroblastoma. Pediatr. Blood Cancer 2013, 60, 985–993. [Google Scholar] [CrossRef]
- Ladenstein, R.; Pötschger, U.; Pearson, A.D.J.; Brock, P.; Luksch, R.; Castel, V.; Yaniv, I.; Papadakis, V.; Laureys, G.; Malis, J.; et al. Busulfan and melphalan versus carboplatin, etoposide, and melphalan as high-dose chemotherapy for high-risk neuroblastoma (HR-NBL1/SIOPEN): An international, randomised, multi-arm, open-label, phase 3 trial. Lancet Oncol. 2017, 18, 500–514. [Google Scholar] [CrossRef]
- Sheng, K.L.; Kang, L.; Pridham, K.J.; Dunkenberger, L.E.; Sheng, Z.; Varghese, R.T. An integrated approach to biomarker discovery reveals gene signatures highly predictive of cancer progression. Sci. Rep. 2020, 10, 21246. [Google Scholar] [CrossRef]
- Dempster, J.M.R.J.; Kazachkova, M.; Pan, J.; Kugener, G.; Root, D.E.; Tsherniak, A. Extracting Biological Insights from the Project Achilles Genome-Scale CRISPR Screens in Cancer Cell Lines. BioRxiv 2019. BioRxiv:720243. [Google Scholar]
- Kimura, K.; Cuvier, O.; Hirano, T. Chromosome condensation by a human condensin complex in Xenopus egg extracts. J. Biol. Chem. 2001, 276, 5417–5420. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Gao, J.; Shi, C.; Guo, W.Z.; Guo, D.; Zhang, S. The role of NCAPG in various of tumors. Biomed. Pharmacother. 2022, 155, 113635. [Google Scholar] [CrossRef] [PubMed]
- Ozaki, T.; Nakagawara, A. Role of p53 in Cell Death and Human Cancers. Cancers 2011, 3, 994–1013. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; White, E. p53-dependent apoptosis pathways. Adv. Cancer Res. 2001, 82, 55–84. [Google Scholar]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef]
- Roderwieser, A.; Sand, F.; Walter, E.; Fischer, J.; Gecht, J.; Bartenhagen, C.; Ackermann, S.; Otte, F.; Welte, A.; Kahlert, Y.; et al. Telomerase Is a Prognostic Marker of Poor Outcome and a Therapeutic Target in Neuroblastoma. JCO Precis. Oncol. 2019, 3, 1–20. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Tanabe, M.; Sato, Y.; Morishima, K. KEGG: New perspectives on genomes, pathways, diseases and drugs. Nucleic Acids Res. 2017, 45, D353–D361. [Google Scholar] [CrossRef]
- Consortium, T.G.O. Gene Ontology Consortium: Going forward. Nucleic Acids Res. 2015, 43, D1049–D1056. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Zhou, Z.R.; Wang, W.W.; Li, Y.; Jin, K.R.; Wang, X.Y.; Wang, Z.W.; Chen, Y.S.; Wang, S.J.; Hu, J.; Zhang, H.N.; et al. In-depth mining of clinical data: The construction of clinical prediction model with R. Ann. Transl. Med. 2019, 7, 796. [Google Scholar] [CrossRef] [PubMed]
- Ternès, N.; Rotolo, F.; Michiels, S. Empirical extensions of the lasso penalty to reduce the false discovery rate in high-dimensional Cox regression models. Stat. Med. 2016, 35, 2561–2573. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic NCAPG Expression | Low Expression (N = 21), N (%) | High Expression (N = 19), N (%) | p-Value |
|---|---|---|---|
| Gender | 0.796 | ||
| Male | 8 (50) | 8 (50) | |
| Female | 13 (54) | 11 (46) | |
| Age | 0.115 | ||
| <18 | 8 (73) | 3 (27) | |
| ≥18 | 13 (45) | 16 (55) | |
| INSS stage | 0.004 | ||
| 1/2/4s | 14 (78) | 4 (22) | |
| 3/4 | 7 (32) | 15 (68) | |
| COG risk | 0.004 | ||
| LR/IR | 14 (78) | 4 (22) | |
| HR | 7 (32) | 15 (68) | |
| Bone marrow metastasis | 0.004 | ||
| No | 17 (71) | 7 (29) | |
| Yes | 4 (25) | 12 (75) | |
| Distant metastasis | 0.027 | ||
| No | 14 (70) | 6 (30) | |
| Yes | 7 (35) | 13 (65) | |
| Histological type | 0.062 | ||
| Ganglioneuroblastoma | 9 (75) | 3 (25) | |
| Neuroblastoma | 12 (43) | 16 (57) | |
| MYCN amplification | 0.112 | ||
| No | 17 (61) | 11 (39) | |
| Yes | 4 (33) | 8 (67) |
| Subtype | Univariate Analysis | Multivariate Analysis | ||
|---|---|---|---|---|
| Hazard Ratio (95%CI) | p-Value | Hazard Ratio (95%CI) | p-Value | |
| Gender | ||||
| Male | 1 | |||
| Female | 1.628 (0.500–5.301) | 0.419 | ||
| Age | ||||
| <18 | 1 | |||
| ≥18 | 2.328 (0.514–10.542) | 0.273 | ||
| INSS stage | ||||
| 1/2/4s | 1 | 1 | ||
| 3/4 | 15.302 (1.974–118.632) | 0.009 | 3.379 (0.354–32.263) | 0.290 |
| Distant metastasis | ||||
| No | 1 | 1 | ||
| Yes | 21.972 (2.788–173.127) | 0.003 | 10.492 (1.049–104.894) | 0.045 |
| Histological type | ||||
| Ganglioneuroblastoma | 1 | |||
| Neuroblastoma | 1.137 (0.349–3.701) | 0.832 | ||
| MYCN amplification | ||||
| No | 1 | |||
| Yes | 2.073 (0.674–6.373) | 0.203 | ||
| NCAPG expression | ||||
| Low | 1 | 1 | ||
| High | 3.636 (1.110–11.913) | 0.033 | 2.639 (0.741–9.391) | 0.134 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Yang, J.; Chen, Y.; Liu, Y.; Jin, Y.; Wang, C.; Gong, B.; Zhao, Q. Identification of NCAPG as an Essential Gene for Neuroblastoma Employing CRISPR-Cas9 Screening Database and Experimental Verification. Int. J. Mol. Sci. 2023, 24, 14946. https://doi.org/10.3390/ijms241914946
Jia Y, Yang J, Chen Y, Liu Y, Jin Y, Wang C, Gong B, Zhao Q. Identification of NCAPG as an Essential Gene for Neuroblastoma Employing CRISPR-Cas9 Screening Database and Experimental Verification. International Journal of Molecular Sciences. 2023; 24(19):14946. https://doi.org/10.3390/ijms241914946
Chicago/Turabian StyleJia, Yubin, Jiaxing Yang, Yankun Chen, Yun Liu, Yan Jin, Chaoyu Wang, Baocheng Gong, and Qiang Zhao. 2023. "Identification of NCAPG as an Essential Gene for Neuroblastoma Employing CRISPR-Cas9 Screening Database and Experimental Verification" International Journal of Molecular Sciences 24, no. 19: 14946. https://doi.org/10.3390/ijms241914946