
Properties of Plant Virus Protein Encoded by the 5′-Proximal Gene of Tetra-Cistron Movement Block
 , , , , and
, , , , and 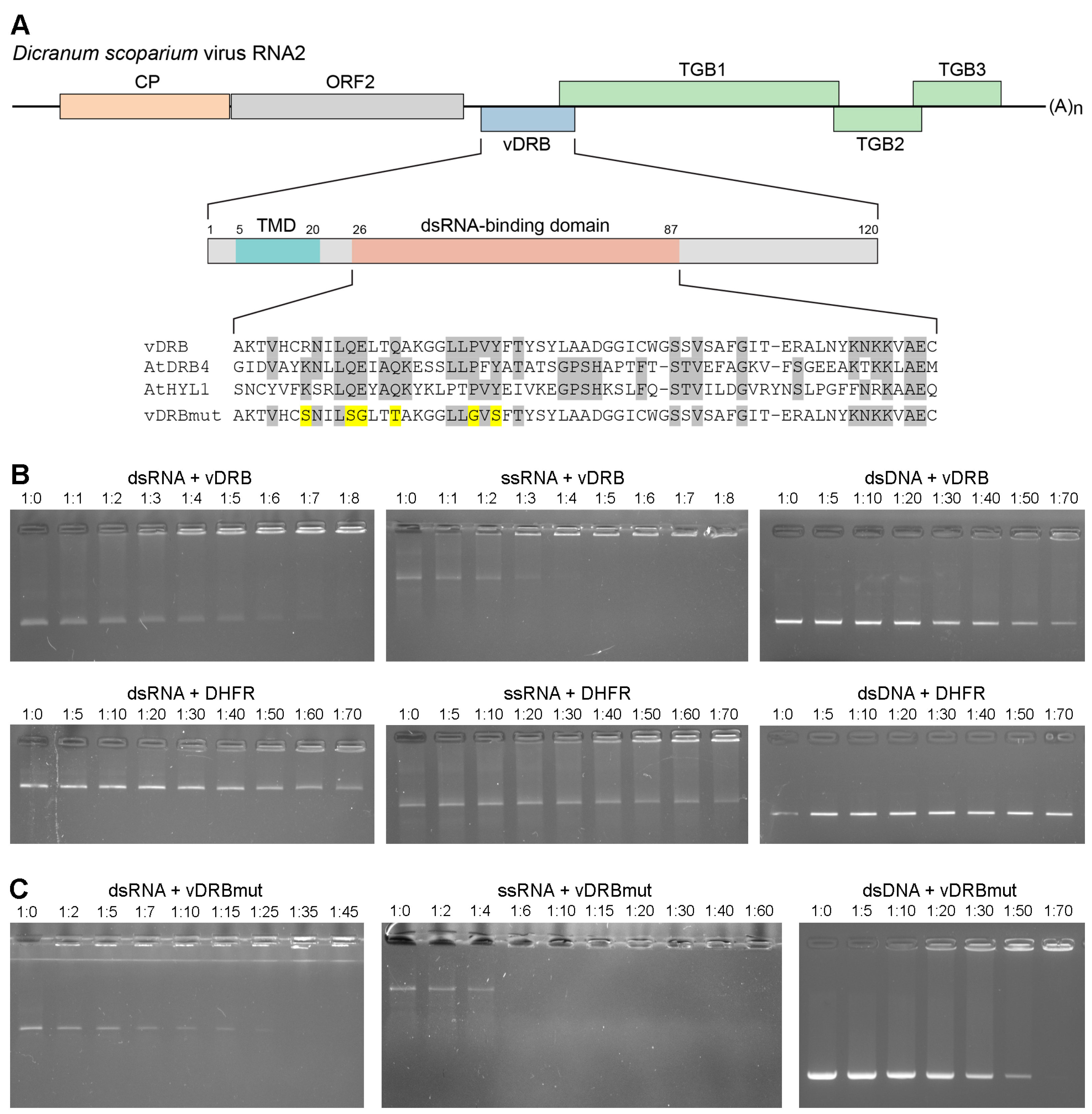{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
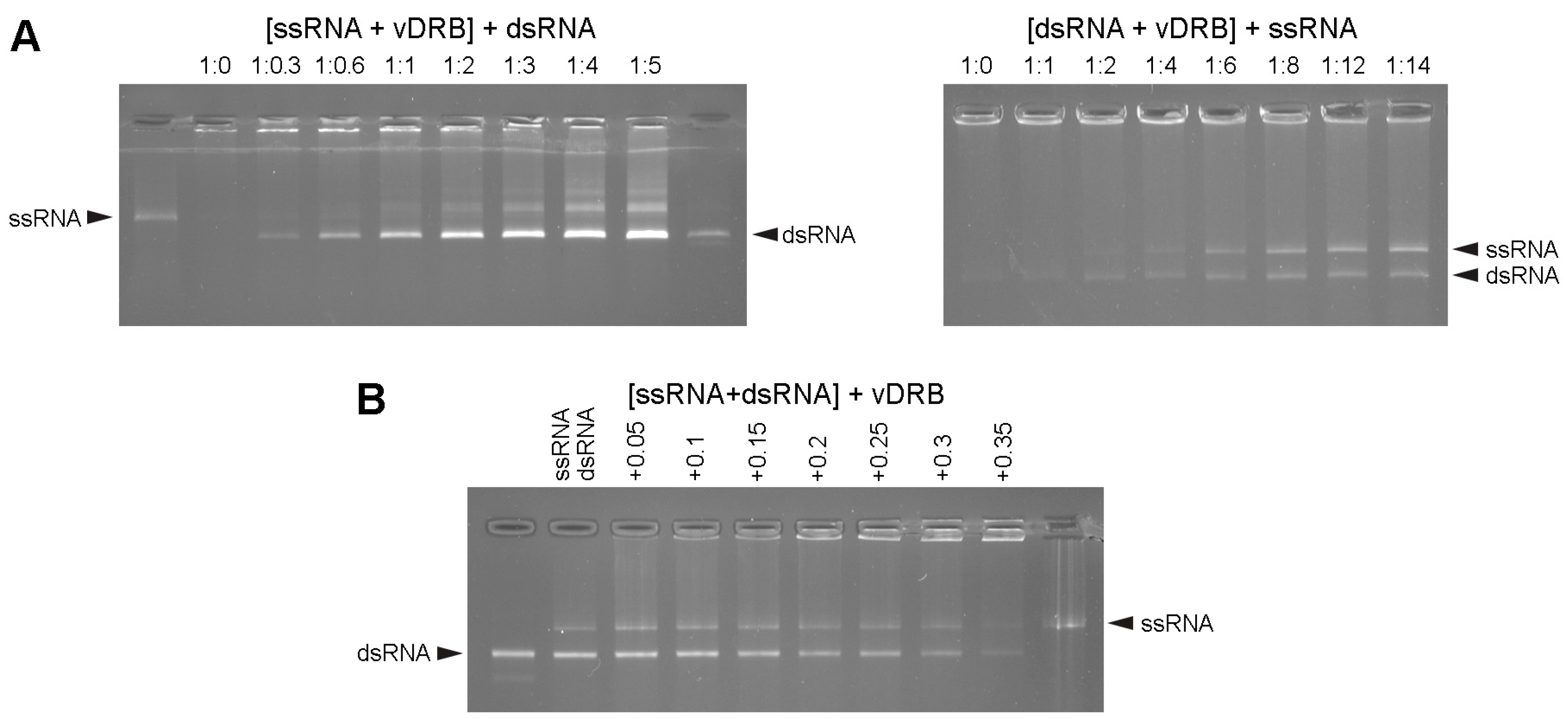
2.1. RNA-Binding Properties of vDRB
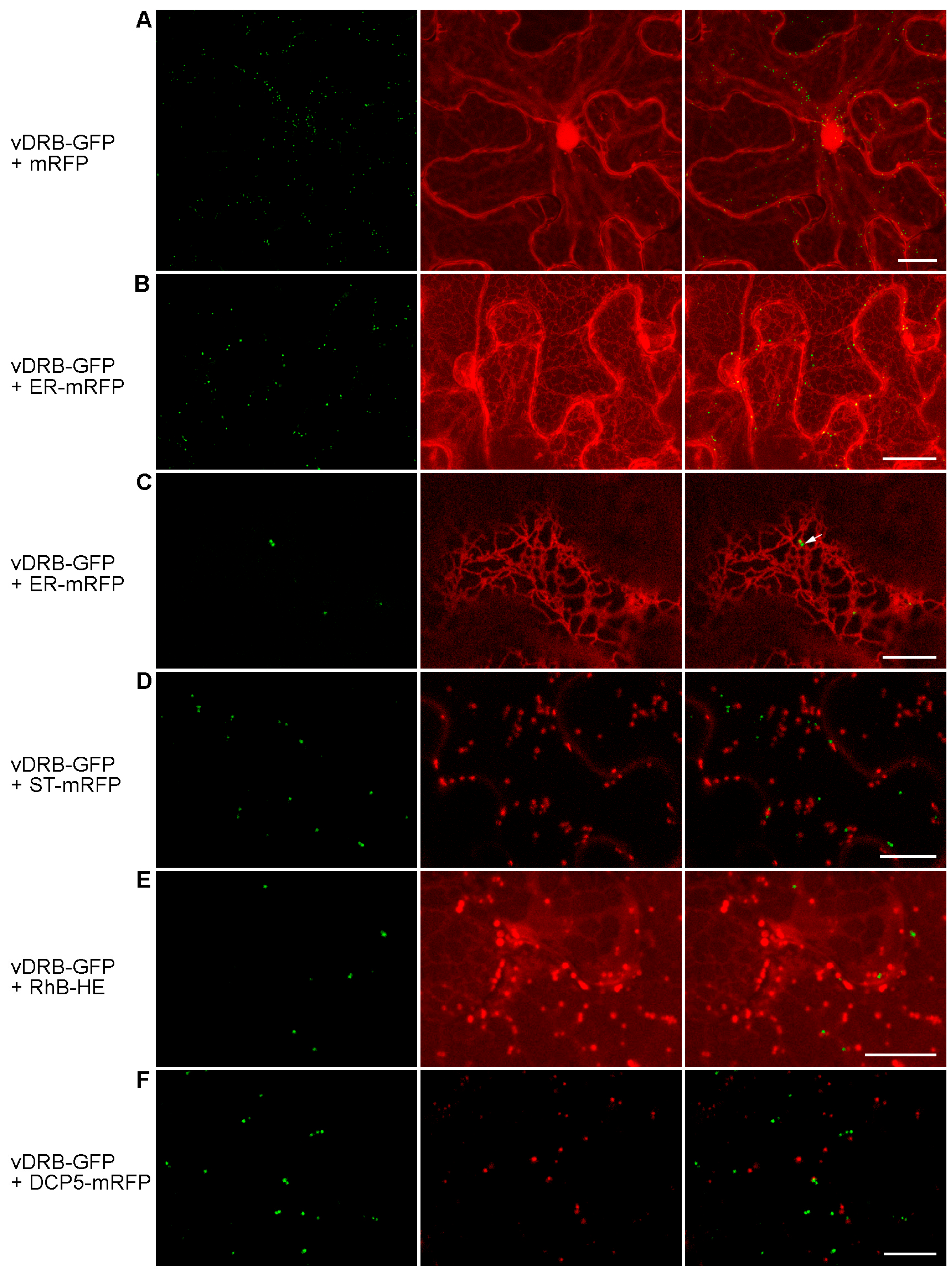
2.2. Subcellular Localization of vDRB
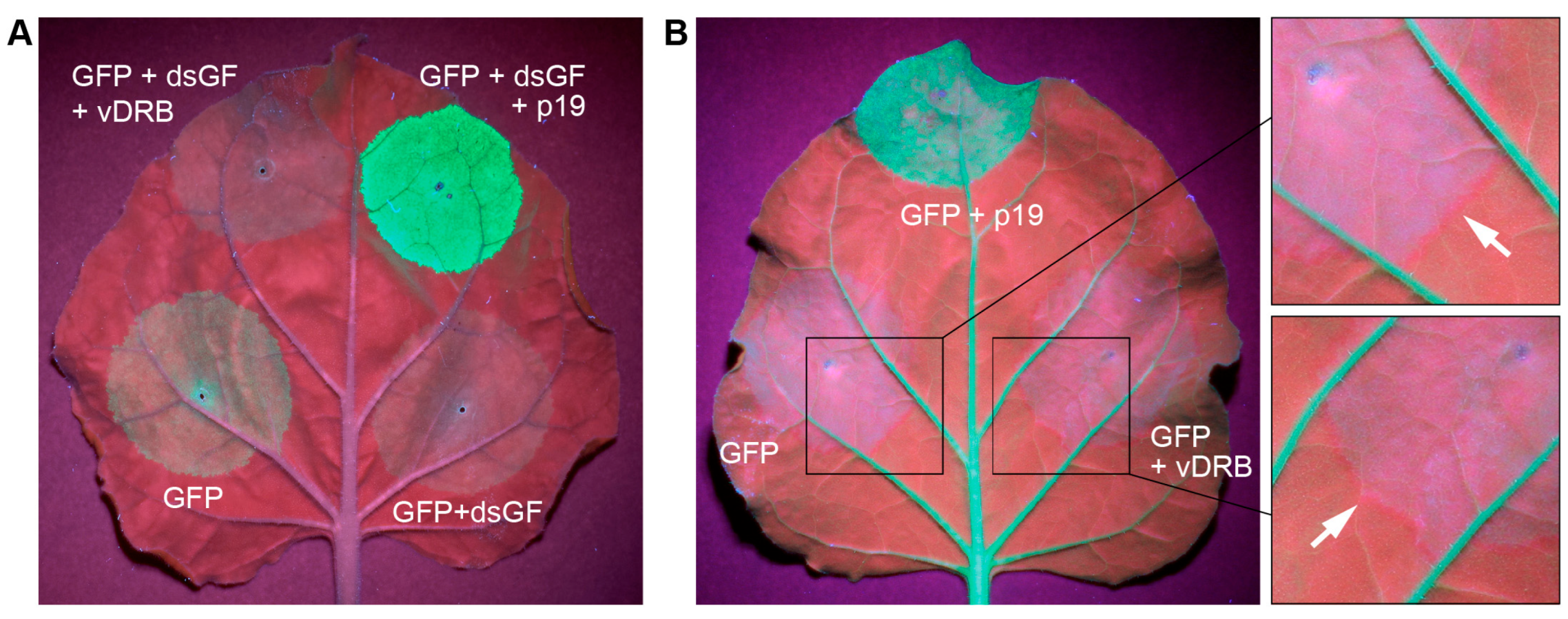
2.3. Suppression of RNA Silencing by vDRB in Non-Viral Experimental Systems
2.4. Suppression of RNA Silencing by vDRB in TCV-Based Assay
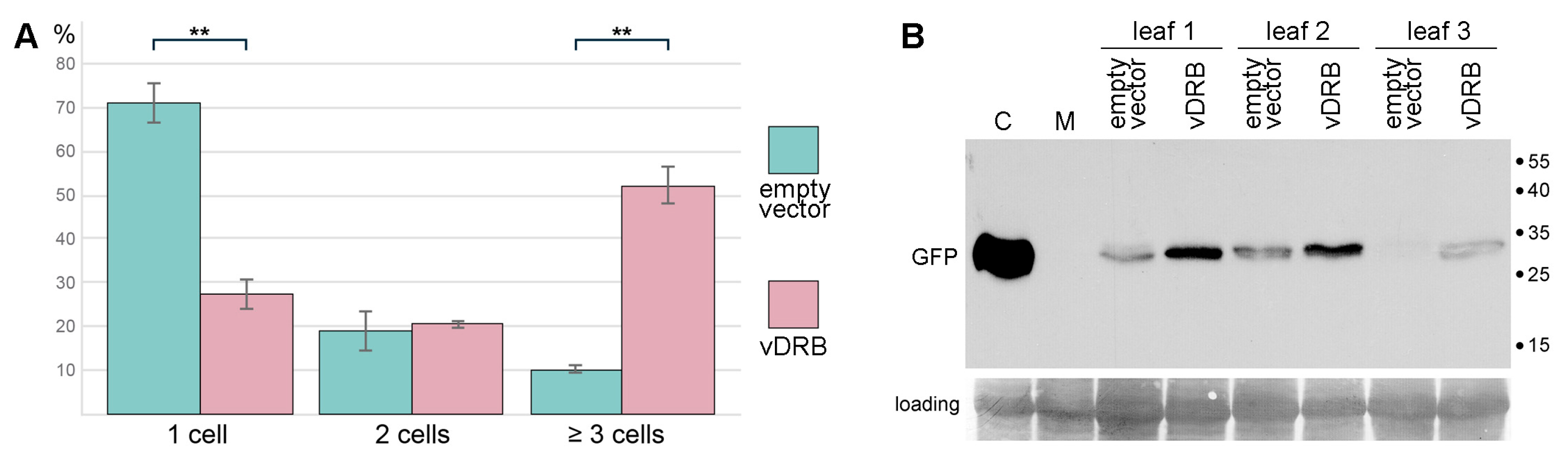
2.5. Influence of vDRB on PVX Infection
3. Discussion
4. Materials and Methods
4.1. Recombinant Constructs
4.2. Protein Expression in E. coli
4.3. Gel-Shift Experiments
4.4. Plant Material
4.5. Plant Agroinfiltration and Inoculation
4.6. Western Blot Analysis
4.7. Quantitative PCR
4.8. Microscopy and Virus Movement Visualization
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Z.P.; Paterlini, A.; Glavier, M.; Bayer, E.M. Intercellular Trafficking via Plasmodesmata: Molecular Layers of Complexity. Cell. Mol. Life Sci. 2021, 78, 799–816. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.G.; Oparka, K.J. Plasmodesmata and the Control of Symplastic Transport. Plant Cell Environ. 2003, 26, 103–124. [Google Scholar] [CrossRef]
- Lucas, W.J. Plant Viral Movement Proteins: Agents for Cell-to-Cell Trafficking of Viral Genomes. Virology 2006, 344, 169–184. [Google Scholar] [CrossRef]
- Pitzalis, N.; Heinlein, M. The Roles of Membranes and Associated Cytoskeleton in Plant Virus Replication and Cell-to-Cell Movement. J. Exp. Bot. 2017, 69, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Heinlein, M. Plant Virus Replication and Movement. Virology 2015, 479–480, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Morozov, S.Y.; Solovyev, A.G. Triple Gene Block: Modular Design of a Multifunctional Machine for Plant Virus Movement. J. Gen. Virol. 2003, 84, 1351–1366. [Google Scholar] [CrossRef] [PubMed]
- Tilsner, J.; Linnik, O.; Louveaux, M.; Roberts, I.M.; Chapman, S.N.; Oparka, K.J. Replication and Trafficking of a Plant Virus Are Coupled at the Entrances of Plasmodesmata. J. Cell Biol. 2013, 201, 981–995. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Liu, J.; Chai, M.; Wang, J.; Li, D.; Wang, A.; Cheng, X. The Potato Virus X TGBp2 Protein Plays Dual Functional Roles in Viral Replication and Movement. J. Virol. 2019, 93, e01635-18. [Google Scholar] [CrossRef]
- Bayne, E.H.; Rakitina, D.V.; Morozov, S.Y.; Baulcombe, D.C. Cell-to-Cell Movement of Potato Potexvirus X Is Dependent on Suppression of RNA Silencing. Plant J. 2005, 44, 471–482. [Google Scholar] [CrossRef]
- Senshu, H.; Ozeki, J.; Komatsu, K.; Hashimoto, M.; Hatada, K.; Aoyama, M.; Kagiwada, S.; Yamaji, Y.; Namba, S. Variability in the Level of RNA Silencing Suppression Caused by Triple Gene Block Protein 1 (TGBp1) from Various Potexviruses during Infection. J. Gen. Virol. 2009, 90, 1014–1024. [Google Scholar] [CrossRef]
- Melzer, M.J.; Sether, D.M.; Borth, W.B.; Hu, J.S. Characterization of a Virus Infecting Citrus Volkameriana with Citrus Leprosis-like Symptoms. Phytopathology 2012, 102, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Lazareva, E.A.; Lezzhov, A.A.; Chergintsev, D.A.; Golyshev, S.A.; Dolja, V.V.; Morozov, S.Y.; Heinlein, M.; Solovyev, A.G. Reticulon-like Properties of a Plant Virus-Encoded Movement Protein. New Phytol. 2021, 229, 1052–1066. [Google Scholar] [CrossRef] [PubMed]
- Lazareva, E.A.; Lezzhov, A.A.; Komarova, T.V.; Morozov, S.Y.; Heinlein, M.; Solovyev, A.G. A Novel Block of Plant Virus Movement Genes. Mol. Plant Pathol. 2017, 18, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Solovyev, A.G.; Morozov, S.Y. Non-Replicative Integral Membrane Proteins Encoded by Plant Alpha-like Viruses: Emergence of Diverse Orphan ORFs and Movement Protein Genes. Front. Plant Sci. 2017, 8, 1820. [Google Scholar] [CrossRef] [PubMed]
- Mifsud, J.C.O.; Gallagher, R.V.; Holmes, E.C.; Geoghegan, J.L. Transcriptome Mining Expands Knowledge of RNA Viruses across the Plant Kingdom. J. Virol. 2022, 96, e0026022. [Google Scholar] [CrossRef] [PubMed]
- Morozov, S.Y.; Solovyev, A.G. Bioinformatic Analysis Predicts a Novel Genetic Module Related to Triple Gene and Binary Movement Blocks of Plant Viruses: Tetra-Cistron Movement Block. Biomolecules 2022, 12, 861. [Google Scholar] [CrossRef] [PubMed]
- Eamens, A.L.; Wook Kim, K.; Waterhouse, P.M. DRB2, DRB3 and DRB5 Function in a Non-Canonical MicroRNA Pathway in Arabidopsis Thaliana. Plant Signal. Behav. 2012, 7, 1224–1229. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Sinha, K.V.; Sopory, S.K.; Sanan-Mishra, N. Influence of Virus–Host Interactions on Plant Response to Abiotic Stress. Plant Cell Rep. 2021, 40, 2225–2245. [Google Scholar] [CrossRef]
- Ryabov, E.V.; Oparka, K.J.; Santa Cruz, S.; Robinson, D.J.; Taliansky, M.E. Intracellular Location of Two Groundnut Rosette Umbravirus Proteins Delivered by PVX and TMV Vectors. Virology 1998, 242, 303–313. [Google Scholar] [CrossRef]
- Xu, J.; Chua, N.-H. Arabidopsis Decapping 5 Is Required for MRNA Decapping, P-Body Formation, and Translational Repression during Postembryonic Development. Plant Cell 2009, 21, 3270–3279. [Google Scholar] [CrossRef]
- Yelina, N.E.; Erokhina, T.N.; Lukhovitskaya, N.I.; Minina, E.A.; Schepetilnikov, M.V.; Lesemann, D.-E.; Schiemann, J.; Solovyev, A.G.; Morozov, S.Y. Localization of Poa Semilatent Virus Cysteine-Rich Protein in Peroxisomes Is Dispensable for Its Ability to Suppress RNA Silencing. J. Gen. Virol. 2005, 86, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Vargason, J.M.; Szittya, G.; Burgyán, J.; Hall, T.M.T. Size Selective Recognition of SiRNA by an RNA Silencing Suppressor. Cell 2003, 115, 799–811. [Google Scholar] [CrossRef] [PubMed]
- Ye, K.; Malinina, L.; Patel, D.J. Recognition of Small Interfering RNA by a Viral Suppressor of RNA Silencing. Nature 2003, 426, 874–878. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, M.T.; Voinnet, O.; Baulcombe, D.C. Initiation and Maintenance of Virus-Induced Gene Silencing. Plant Cell 1998, 10, 937–946. [Google Scholar] [CrossRef] [PubMed]
- Himber, C.; Dunoyer, P.; Moissiard, G.; Ritzenthaler, C.; Voinnet, O. Transitivity-Dependent and -Independent Cell-to-Cell Movement of RNA Silencing. EMBO J. 2003, 22, 4523–4533. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.G.; Sit, T.L.; Qu, F.; Morris, T.J.; Kim, K.-H.; Lommel, S.A. A Versatile Assay for the Identification of RNA Silencing Suppressors Based on Complementation of Viral Movement. Mol. Plant. Microbe. Interact. 2008, 21, 879–890. [Google Scholar] [CrossRef] [PubMed]
- Powers, J.G.; Sit, T.L.; Heinsohn, C.; George, C.G.; Kim, K.-H.; Lommel, S. a The Red Clover Necrotic Mosaic Virus RNA-2 Encoded Movement Protein Is a Second Suppressor of RNA Silencing. Virology 2008, 381, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Brigneti, G.; Voinnet, O.; Li, W.; Ji, L. Viral Pathogenicity Determinants Are Suppressors of Transgene Silencing in Nicotiana Benthamiana. EMBO J. 1998, 17, 6739–6746. [Google Scholar] [CrossRef]
- Scholthof, H.B.; Scholthof, K.B.; Jackson, A.O. Identification of Tomato Bushy Stunt Virus Host-Specific Symptom Determinants by Expression of Individual Genes from a Potato Virus X Vector. Plant Cell 1995, 7, 1157–1172. [Google Scholar] [CrossRef]
- Chen, L.-J.; Liu, J.; Zhao, F.-F.; Li, J.-Y.; Wang, S.-X.; Lin, H.-H.; Xi, D.-H. Characterisation of the Dark Green Islands of Cucumber Mosaic Virus Infected Nicotiana Tabacum. Plant Cell Rep. 2015, 34, 1225–1238. [Google Scholar] [CrossRef]
- Moore, C.J.; Sutherland, P.W.; Forster, R.L.S.; Gardner, R.C.; MacDiarmid, R.M. Dark Green Islands in Plant Virus Infection Are the Result of Posttranscriptional Gene Silencing. Mol. Plant-Microbe Interact. 2001, 14, 939–946. [Google Scholar] [CrossRef] [PubMed]
- Pouch-Pélissier, M.-N.; Pélissier, T.; Elmayan, T.; Vaucheret, H.; Boko, D.; Jantsch, M.F.; Deragon, J.-M. SINE RNA Induces Severe Developmental Defects in Arabidopsis Thaliana and Interacts with HYL1 (DRB1), a Key Member of the DCL1 Complex. PLoS Genet. 2008, 4, e1000096. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, E.; Almendral, D.; Allende, L.; Pacheco, R.; Chung, B.N.; Canto, T.; Tenllado, F. The P25 Protein of Potato Virus X (PVX) Is the Main Pathogenicity Determinant Responsible for Systemic Necrosis in PVX-Associated Synergisms. J. Virol. 2015, 89, 2090–2103. [Google Scholar] [CrossRef] [PubMed]
- Pfeffer, S.; Dunoyer, P.; Heim, F.; Richards, K.E.; Jonard, G.; Ziegler-Graff, V. P0 of Beet Western Yellows Virus Is a Suppressor of Posttranscriptional Gene Silencing. J. Virol. 2002, 76, 6815–6824. [Google Scholar] [CrossRef] [PubMed]
- Luna, A.P.; Morilla, G.; Voinnet, O.; Bejarano, E.R. Functional Analysis of Gene-Silencing Suppressors from Tomato Yellow Leaf Curl Disease Viruses. Mol. Plant-Microbe Interact. 2012, 25, 1294–1306. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dong, J.; Xu, Y.; Wu, J. V2 Protein Encoded by Tomato Yellow Leaf Curl China Virus Is an RNA Silencing Suppressor. Virus Res. 2012, 163, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Burgyán, J.; Havelda, Z. Viral Suppressors of RNA Silencing. Trends Plant Sci. 2011, 16, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Incarbone, M.; Clavel, M.; Monsion, B.; Kuhn, L.; Scheer, H.; Vantard, É.; Poignavent, V.; Dunoyer, P.; Genschik, P.; Ritzenthaler, C. Immunocapture of DsRNA-Bound Proteins Provides Insight into Tobacco Rattle Virus Replication Complexes and Reveals Arabidopsis DRB2 to Be a Wide-Spectrum Antiviral Effector. Plant Cell 2021, 33, 3402–3420. [Google Scholar] [CrossRef]
- Huang, C.; Sede, A.R.; Elvira-González, L.; Yan, Y.; Rodriguez, M.; Mutterer, J.; Boutant, E.; Shan, L.; Heinlein, M. DsRNA-Induced Immunity Targets Plasmodesmata and Is Suppressed by Viral Movement Proteins. Plant Cell 2023. ahead of print. [Google Scholar] [CrossRef]
- Deng, X.; Kelloniemi, J.; Haikonen, T.; Vuorinen, A.L.; Elomaa, P.; Teeri, T.H.; Valkonen, J.P.T. Modification of Tobacco Rattle Virus RNA1 to Serve as a VIGS Vector Reveals That the 29K Movement Protein Is an RNA Silencing Suppressor of the Virus. Mol. Plant. Microbe. Interact. 2013, 26, 503–514. [Google Scholar] [CrossRef]
- Surjit, M.; Jameel, S.; Lal, S.K. Cytoplasmic Localization of the ORF2 Protein of Hepatitis E Virus Is Dependent on Its Ability To Undergo Retrotranslocation from the Endoplasmic Reticulum. J. Virol. 2007, 81, 3339–3345. [Google Scholar] [CrossRef]
- Duriez, M.; Rossignol, J.-M.; Sitterlin, D. The Hepatitis B Virus Precore Protein Is Retrotransported from Endoplasmic Reticulum (ER) to Cytosol through the ER-Associated Degradation Pathway. J. Biol. Chem. 2008, 283, 32352–32360. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Savenkov, E.I.; Valkonen, J.P.T. Complete Genome Sequence and Analyses of the Subgenomic RNAs of Sweet Potato Chlorotic Stunt Virus Reveal Several New Features for the Genus Crinivirus. J. Virol. 2002, 76, 9260–9270. [Google Scholar] [CrossRef] [PubMed]
- Chao, J.A.; Lee, J.H.; Chapados, B.R.; Debler, E.W.; Schneemann, A.; Williamson, J.R. Dual Modes of RNA-Silencing Suppression by Flock House Virus Protein B2. Nat. Struct. Mol. Biol. 2005, 12, 952–957. [Google Scholar] [CrossRef] [PubMed]
- Kreuze, J.F.; Savenkov, E.I.; Cuellar, W.; Li, X.; Valkonen, J.P.T. Viral Class 1 RNase III Involved in Suppression of RNA Silencing. J. Virol. 2005, 79, 7227–7238. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, W.X.; Ding, S.W. Induction and Suppression of RNA Silencing by an Animal Virus. Science 2002, 296, 1319–1321. [Google Scholar] [CrossRef] [PubMed]
- Santa Cruz, S.; Chapman, S.; Roberts, A.G.; Roberts, I.M.; Prior, D.A.; Oparka, K.J. Assembly and Movement of a Plant Virus Carrying a Green Fluorescent Protein Overcoat. Proc. Natl. Acad. Sci. USA 1996, 93, 6286–6290. [Google Scholar] [CrossRef] [PubMed]
- Boevink, P.; Cruz, S.; Hawes, C.; Harris, N.; Oparka, K.J. Virus-Mediated Delivery of the Green Fluorescent Protein to the Endoplasmic Reticulum of Plant Cells. Plant J. 1996, 10, 935–941. [Google Scholar] [CrossRef]
- Makarova, S.S.; Solovyev, A.G.; Morozov, S.Y. RNA-Binding Properties of the Plant Protein Nt-4/1. Biochemistry 2014, 79, 717–726. [Google Scholar] [CrossRef]
- Solovyev, A.G.; Minina, E.A.; Makarova, S.S.; Erokhina, T.N.; Makarov, V.V.; Kaplan, I.B.; Kopertekh, L.; Schiemann, J.; Richert-Pöggeler, K.R.; Morozov, S.Y. Subcellular Localization and Self-Interaction of Plant-Specific Nt-4/1 Protein. Biochimie 2013, 95, 1360–1370. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chergintsev, D.A.; Solovieva, A.D.; Atabekova, A.K.; Lezzhov, A.A.; Golyshev, S.A.; Morozov, S.Y.; Solovyev, A.G. Properties of Plant Virus Protein Encoded by the 5′-Proximal Gene of Tetra-Cistron Movement Block. Int. J. Mol. Sci. 2023, 24, 14144. https://doi.org/10.3390/ijms241814144
Chergintsev DA, Solovieva AD, Atabekova AK, Lezzhov AA, Golyshev SA, Morozov SY, Solovyev AG. Properties of Plant Virus Protein Encoded by the 5′-Proximal Gene of Tetra-Cistron Movement Block. International Journal of Molecular Sciences. 2023; 24(18):14144. https://doi.org/10.3390/ijms241814144
Chicago/Turabian StyleChergintsev, Denis A., Anna D. Solovieva, Anastasia K. Atabekova, Alexander A. Lezzhov, Sergei A. Golyshev, Sergey Y. Morozov, and Andrey G. Solovyev. 2023. "Properties of Plant Virus Protein Encoded by the 5′-Proximal Gene of Tetra-Cistron Movement Block" International Journal of Molecular Sciences 24, no. 18: 14144. https://doi.org/10.3390/ijms241814144