1. Introduction
The reversibly glycosylated polypeptides (RGPs) were so named due to their ability to reversibly bind sugars in vitro by autoglycosylating a conserved arginine residue [
1,
2,
3,
4]; however, the function of these proteins in vivo remained unclear. Later, when UDP-Arap/UDP-Araf mutase activity was identified [
5], the RGPs were given a new name—UDP-Arap mutases (UAMs). Surprisingly, not all proteins in the family possess UAM activity. Therefore, based on this feature, they were divided into two classes: catalytically active RGPs were classified as class 1 RGPs, whereas the others, which lacked UAM activity, were classified as class 2. A range of 1 to 5 RGPs were identified in different species, both dicots and monocots. The majority of the known RGPs were expected to have UAM activity based on sequence similarity and were included in class 1. However, experimental evidence for UAM activity has only been obtained for
Arabidopsis thaliana RGPs 1–3 [
6],
Oryza sativa UAMs 1 and 3 [
5],
Hordeum vulgare UAMs 1–3 [
7], and
Nicotiana tabacum UAMs 1–4 [
8].
RGPs have also been demonstrated to play an important role in growth and development: they act cooperatively to provide sufficient UDP-Araf for polysaccharide synthesis of the cell wall and are highly expressed in actively growing tissues [
6,
9,
10]. Abiotic stresses such as wounding and dehydration induce
RGP expression, as it was shown in a study of an
RGP1 promoter from
Gossypium hirsutum in transgenic tobacco plants [
11]. Chromium (Cr) stress leads to the accumulation of OsRGP3 protein in rice roots and callose in cell walls, which, together with many other factors accompanying Cr stress, provides an enhanced immobilization of Cr ions and protects rice roots against Cr toxicity [
12]. In general, environmental stresses trigger a cascade of defense reactions and modulation of the intercellular transport in plant tissues via changes in the plasmodesmata (Pd) permeability [
13,
14]. Among multiple Pd affecting factors, RGPs are regarded as negative regulators [
15,
16,
17,
18].
Several studies indicate that RGPs contribute to plant–virus interactions: tobacco mosaic virus (TMV) infection spread was impeded in
AtRGP2:GFP transgenic tobacco [
15], whereas
RGP silencing in
N. benthamiana stimulated TMV intercellular spread [
16].
AtRGP2:GFP overexpression in
N. tabacum led to an increased callose deposition around plasmodesmata (Pd), which allowed the proposal of a putative mechanism for TMV intercellular movement restriction via callose-mediated decrease in Pd permeability [
15]. Another member of the RGP family—UDP-glucose:protein transglucosylase (UPTG) from
Solanum lycopersicum [
19,
20]—was shown to interact with tomato leaf curl virus (TLCV) V1 capsid protein; expression of SlUPTG1 in
N. benthamiana plants in a transient TLCV replication assay increased the accumulation of viral DNA [
20].
One of the first discovered members of the RGP family, salt-extractable wall-associated 41 kDa protein (SeWAP41), was isolated from the cell wall fraction of maize seedling mesocotyls. Immunogold staining revealed punctate localization of the protein in association with Pd and Golgi membranes [
21]. Co-localization with the Golgi membranes was also demonstrated by Dhugga et al. [
2] for
Pisum sativum RGP. Later, numerous studies confirmed the association of RGPs with the Golgi apparatus in many different species [
4,
5,
6,
9,
22]. With regard to plasmodesmal localization, however, the data are more controversial. Besides SeWAP41, Pd localization was demonstrated and confirmed by plasmolysis for SlUPTG:GFP in
Allium cepa epidermal cells after biolistic transformation [
20] and AtRGP2:GFP in transgenic tobacco plants [
9,
15]. However, AtRGP:YFP translational fusions gave no Pd fluorescence in transgenic
A. thaliana plants [
6,
22]. This discrepancy between the observations for intracellular localization could be explained by the ectopic expression of the studied RGPs in a foreign plant as compared with the plant of origin and probably by the difference in the intracellular traffic of these proteins depending on the species.
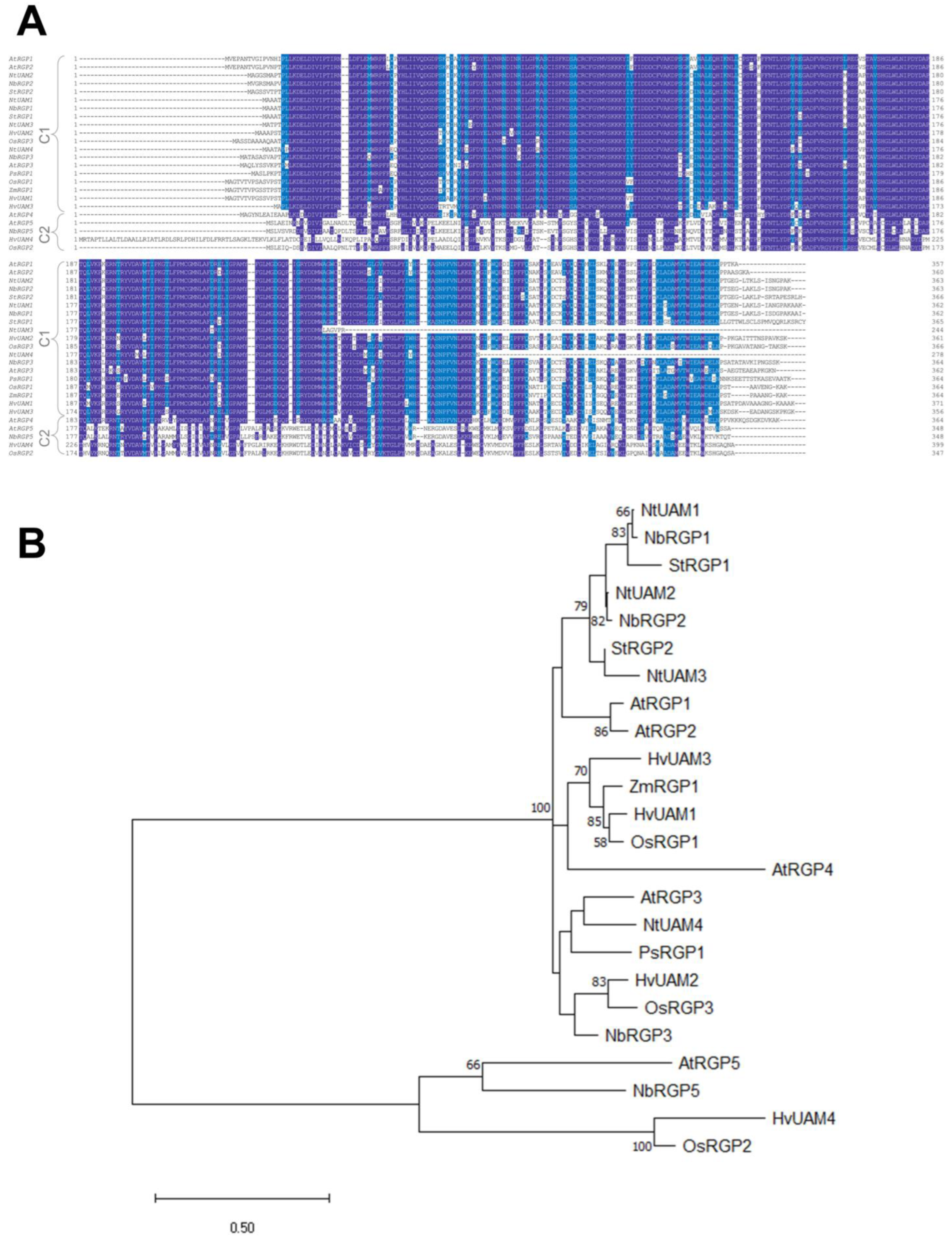
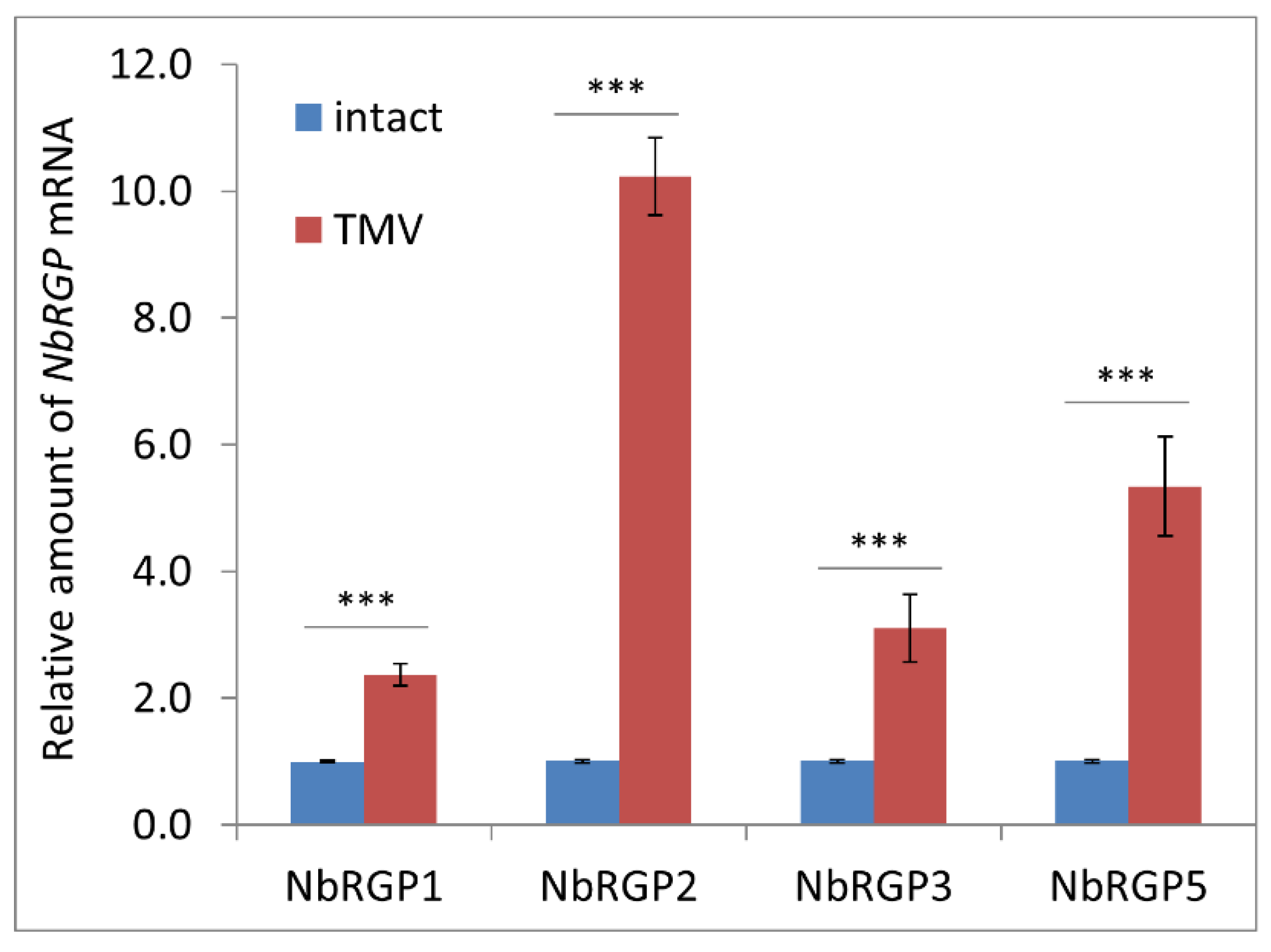
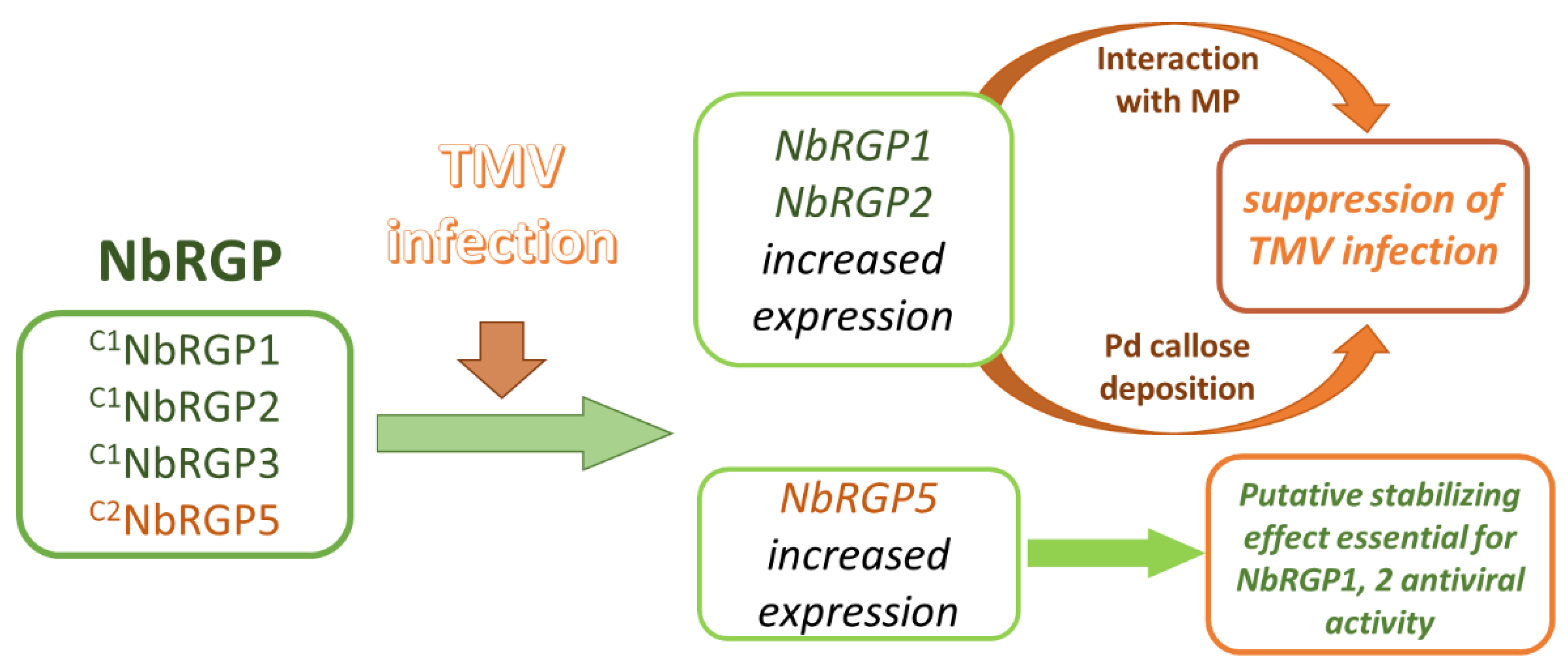
To understand whether N. benthamiana RGPs share features with the described RGPs, and reveal the role of NbRGPs in TMV infection, we used transient expression in N. benthamiana as an alternative to stably transformed plants. We have identified at least four members of the RGP family in the N. benthamiana genome and designated them as NbRGPs 1, 2, 3, and 5 based on their similarity to the known RGPs from A. thaliana and N. tabacum. Based on a homology search, NbRGP5 belongs to class 2, whereas the other three, NbRGP1–3, belong to class 1. We revealed an increased accumulation of all four NbRGP mRNAs in N. benthamiana leaves with systemic TMV infection. Moreover, class 1 NbRGP expression was demonstrated to suppress viral reproduction and local spread. To elucidate the mechanism underlying this effect, we have shown that (i) NbRGP1 and 2 stimulate Pd callose deposition and (ii) NbRGP1 directly interacts with TMV movement protein (MP) in vitro and in vivo. Based on the obtained results we hypothesize that class 1 NbRGP1 and 2 have a dual-mode action against the virus leading to the limitation of TMV local spread, having both general (via a callose-dependent mechanism) and specific (impediment to MP functioning) actions. Therefore, they play the role of antiviral cellular factors.
3. Discussion
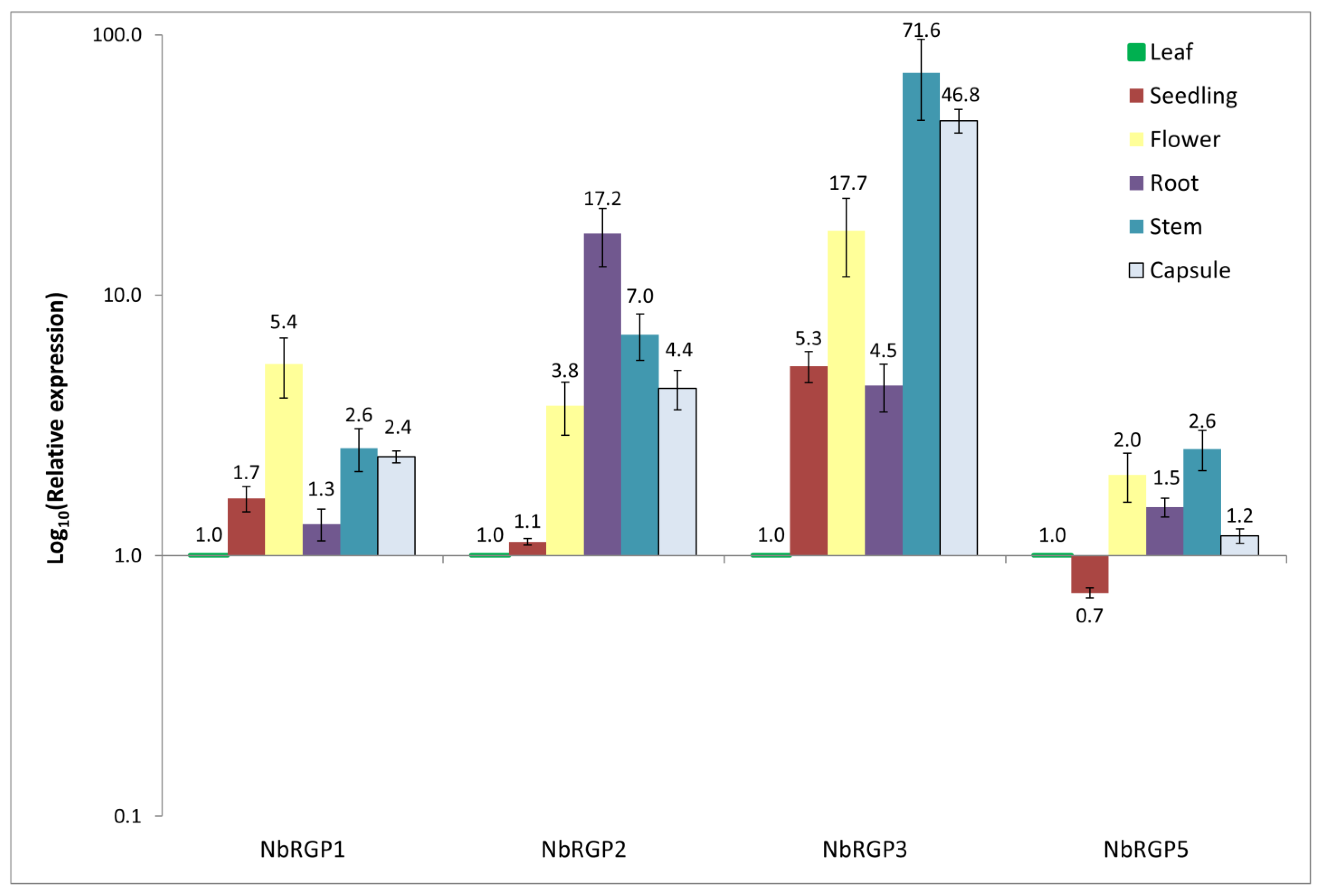
Reversibly glycosylated polypeptides are found in numerous plant species, both dicots and monocots, as well as in algae, mosses, spore plants, etc. [
6,
10,
37]. They play an important role at various stages of plant growth and development, contributing to cell wall formation, pollen development, response to different stresses, plant–pathogen interactions, and intercellular transport regulation. Attention to this protein family is related mainly to their UDP-Arap mutase activity, as Arap–Araf interconversion is indispensable for plant development [
6,
10,
37,
38].
We identified four
RGP genes in the
N. benthamiana genome assembly. As alignment with other known RGPs and a constructed phylogenetic tree show, NbRGPs 1–3 and NbRGP5 can be classified as class 1 and class 2 RGPs, respectively. We obtained the expression pattern of identified
NbRGPs using qRT-PCR (
Figure 2). According to our results,
NbRGP5 expression is evenly distributed in all analyzed organs and is lower in seedlings, which is consistent with the results from the Version 6 Gene expression Atlas (
https://sefapps02.qut.edu.au/atlas/tREX6.php (accessed on 8 August 2021)) based on the microarray data (
Figure S6). However, for
NbRGP1-3, the Gene Expression Atlas demonstrates prevailing root expression, whereas our data does not fully confirm such distribution: high root expression is detected for
NbRGP2–3 rather than
NbRGP1. This discrepancy between the qRT-PCR result and microarray data for
NbRGP1 and
2 could be explained by the high sequence similarity of these two genes, due to which the assembly and distribution of the microarray-based data might be not fully correct. In leaves, all
NbRGPs are fairly weakly expressed, and
NbRGP1 and
5 are represented mainly in leaves compared with other organs. The observed expression pattern of
NbRGPs (
Figure 2) resembles the previously obtained one for
AtRGPs [
22].
NbRGPs 1–3 are highly similar to the recently identified UAMs from
N. tabacum (
Figure 1), for which UAM activity was confirmed in vitro [
8]. This suggests that NbRGPs 1–3 could also be classified as UAMs. However, there is another equally important aspect of RGP function, namely their role in the development of viral infection [
9,
15,
16,
20]. We analyzed
NbRGP mRNA accumulation levels in
N. benthamiana plants with systemic TMV infection and revealed that viral infection induces a significant increase in the expression of all four
NbRGPs (
Figure 3).
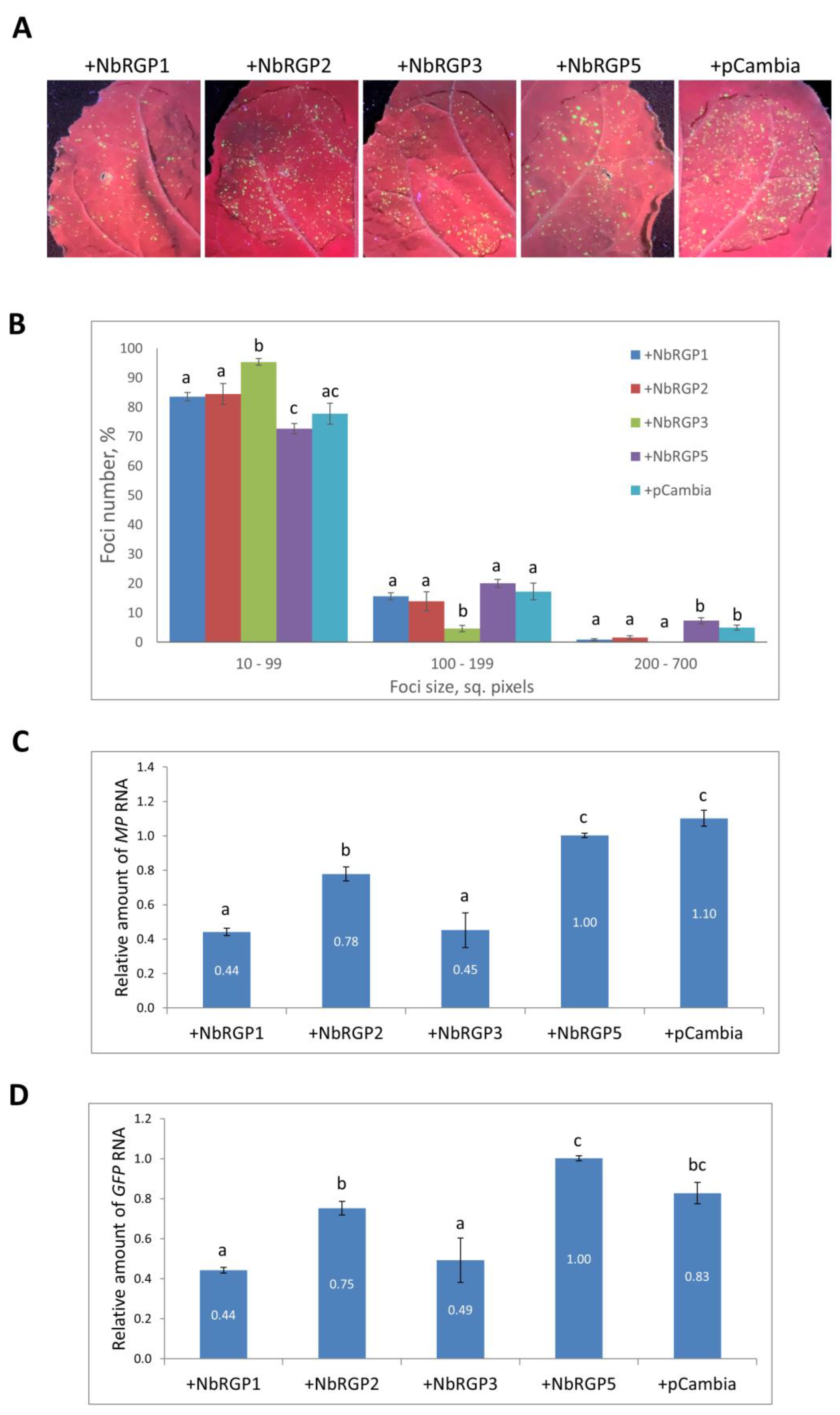
Previously, constitutively expressed
AtRGP2:GFP in
N. tabacum was demonstrated to limit TMV local and systemic spread [
15]. In this study, we assessed the role of NbRGPs in the development of TMV infection in
N. benthamiana plants. We created a model system using a TMV-based TMV:GFP viral vector to imitate infection and performed co-expression of TMV:GFP with each of the identified
NbRGPs in leaves. Visualizing infection foci under UV light, we assessed the efficiency of intercellular spread of the virus, measuring zones with GFP fluorescence. We found out that all three class 1 RGPs suppress intercellular transport of the virus as compared with NbRGP5 and control, an empty binary vector (pCambia1300). Viral replication manifesting itself in accumulation of the viral RNA encoding MP and GFP was also reduced when there was increased expression of class 1
NbRGPs rather than
NbRGP5. Therefore, we showed that the increased expression of class 1
NbRGPs provides protection from viral infection, which is consistent with the data obtained by Burch-Smith et al., who showed that class 1
NbRGP silencing accelerated the development of systemic TMV infection, whereas a TMV-based viral vector demonstrated more extensive spread in those plants [
16].
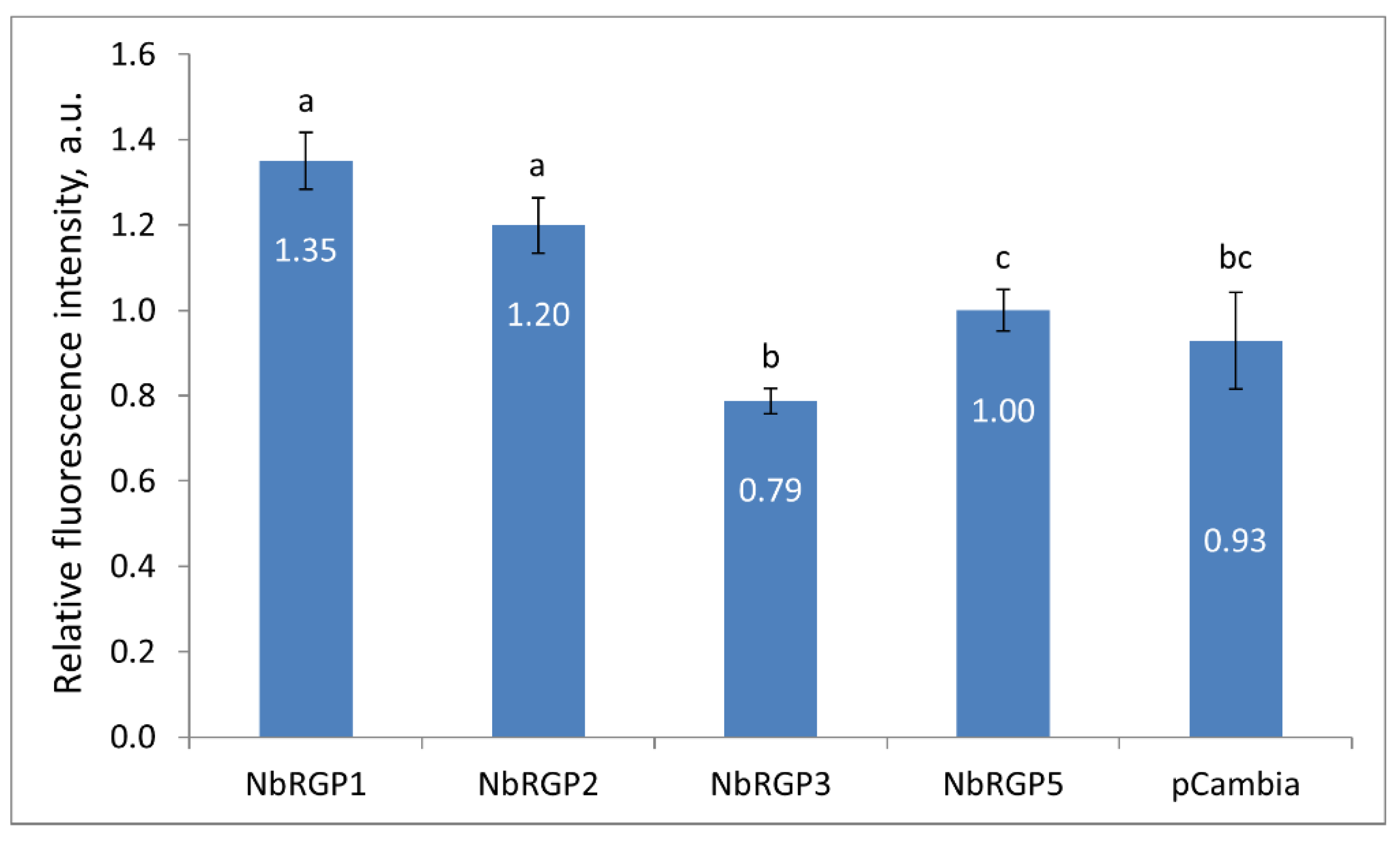
Our research into the mechanism of antiviral action of class 1 NbRGPs stemmed from the following: firstly, a well-known fact that RGPs are capable of increasing callose deposition around Pd [
15] and secondly that RGPs can be localized in Pd, creating a physical barrier for the intercellular transport of macromolecules [
9,
15,
21]. Thirdly, when RGPs are localized in Pd they can interact with TMV MP and inhibit its function, i.e., intercellular transport of viral genetic material. Indeed, according to our data, elevated
NbRGP1 and
2 expression leads to an increase in Pd callose levels by 35% and 20%, respectively, as compared with
NbRGP5 (
Figure 5). As far as NbRGP3 is concerned, the results were contradictory. The increased
NbRGP3 expression suppressed TMV reproduction and intercellular transport; however, it caused a reduction in Pd callose level. This could be explained either by the fact that NbRGP3 needs to form complexes with other NbRGPs to fulfil its function or by the spatiotemporal features of its expression, i.e., the fact that normally in plants it is expressed and functions presumably not in leaves but other organs or at other stages of development. Thus, in our experimental system its expression in leaves is ectopic. For instance, research into
AtRGP3‘s expression pattern revealed its transient but strong expression during seed development [
6]. The authors suggest that AtRGP3 might be involved in Ara supply for the growing cell wall of the developing embryo, delivered via the endosperm. When we analyzed the expression of all four
NbRGPs in response to the systemic TMV infection, we revealed
NbRGP3 upregulation in leaves. Therefore, the mechanism for its suppressing impact on TMV reproduction and intercellular transport calls for further studies.
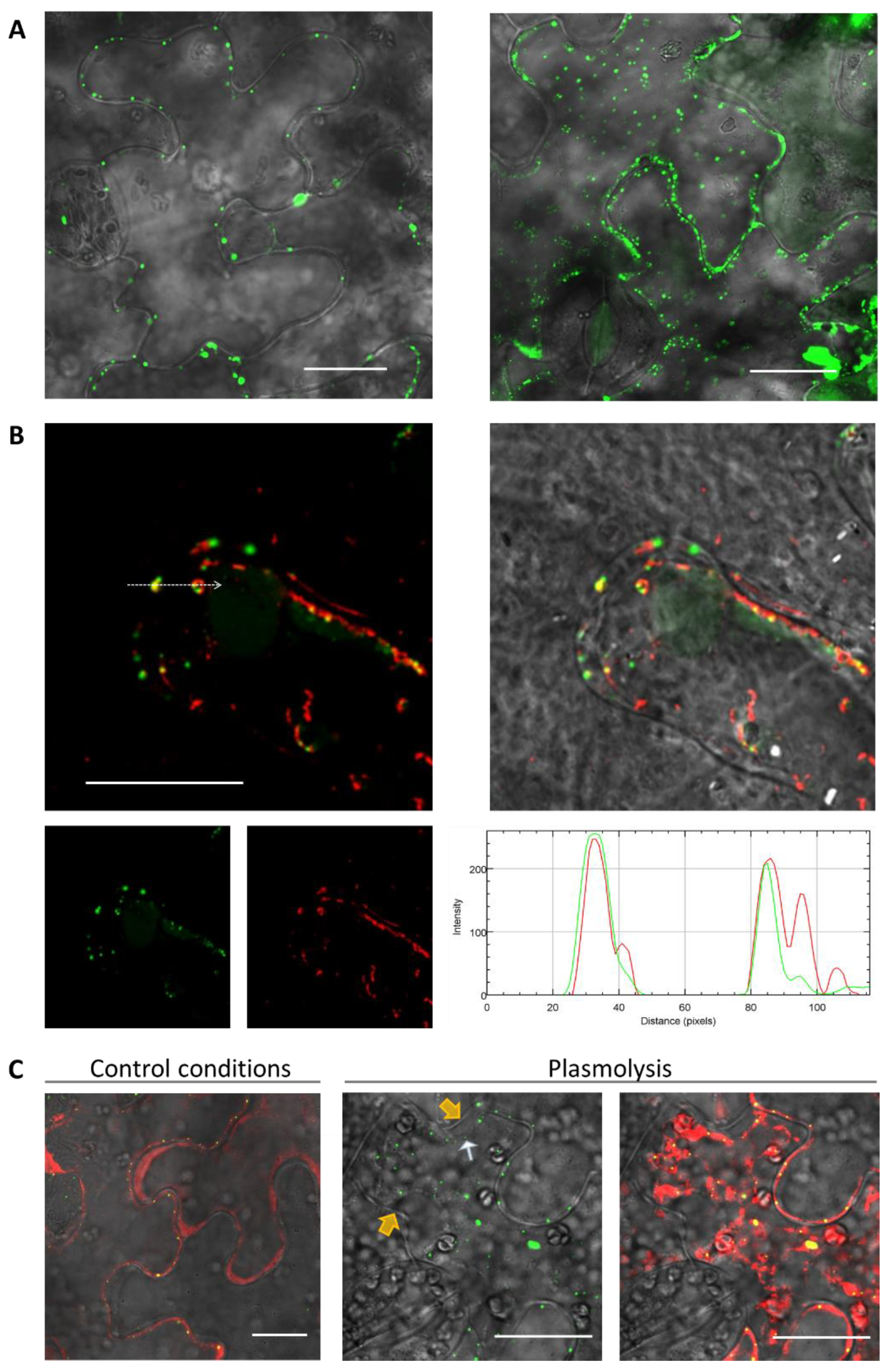
To study the intracellular localization, we chose NbRGP1, which had a high degree of amino acid sequence identity (94%) with NbRGP2 and with AtRGP1 and AtRGP2 (92% and 90% respectively). Additionally, the NbRGP1 nucleotide sequence has 97% similarity with
N. tabacum UAM1, whereas their amino acid sequences are almost identical (99%) [
8]. Our results revealed punctate distribution of NbRGP1:GFP at the cell periphery and partial co-localization with MP (
Figure 8A), which could be suggested to correspond to the Golgi apparatus that is characteristic of the RGPs of other species [
4,
5,
6,
9,
22]. We also detected aggregates of NbRGP1:GFP in the cytoplasm similar to those previously described [
4,
6,
22]. As regards to Pd localization, we did not find NbRGP1 in Pd of
N. benthamiana leaves (
Figure 7C). We have demonstrated that NbRGP1:GFP co-localized with the Golgi marker AtGONST4 (
Figure 7B), which is in line with previously published results for other RGPs. Several research groups have showed that different RGPs are associated with Golgi membranes and distributed in the cytoplasm [
2,
4,
6,
19,
39]. However, it could not be excluded that NbRGP1 interacting with MP is potentially relocalized to Pd, serving there as a mechanical obstacle for intercellular transport. Sagi and colleagues [
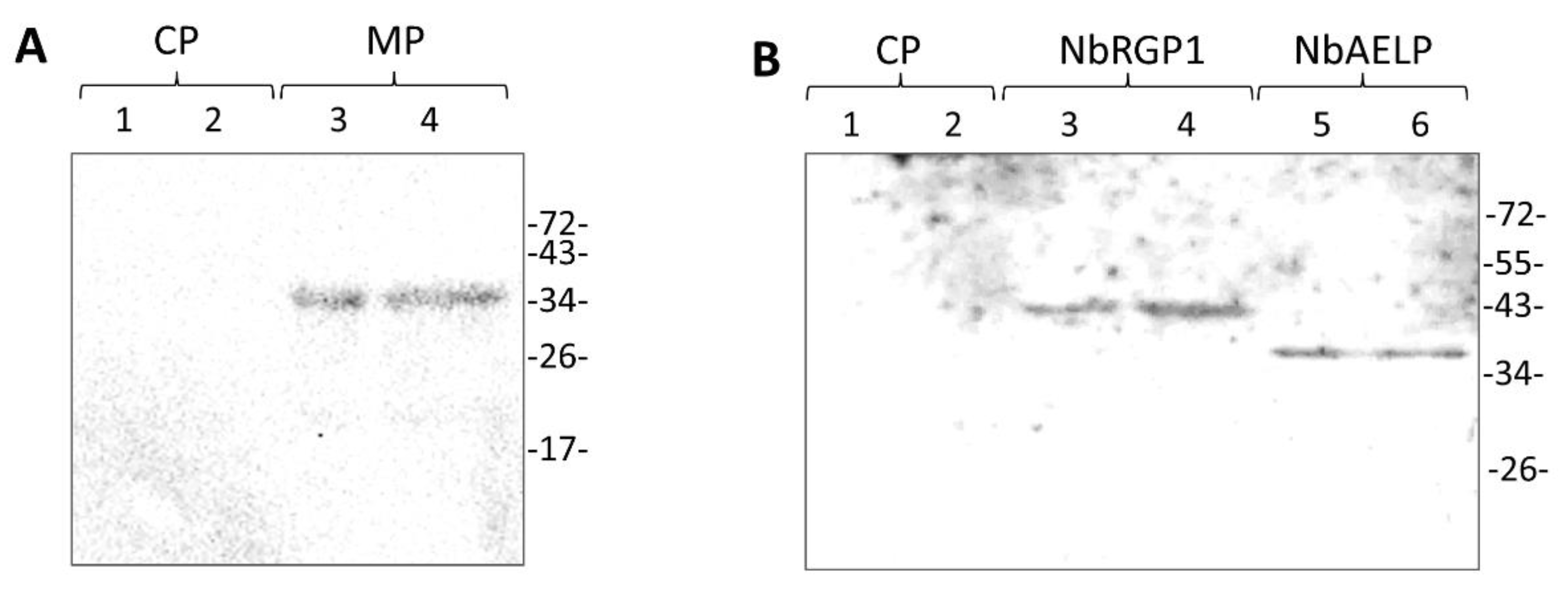
9] put forward such a hypothetical model for RGP-mediated Pd permeability reduction. We used two approaches that convincingly confirmed the interaction between these two proteins in vitro and in vivo. We demonstrated interaction between NbRGP1 and MP in vitro using the overlay assay technique (
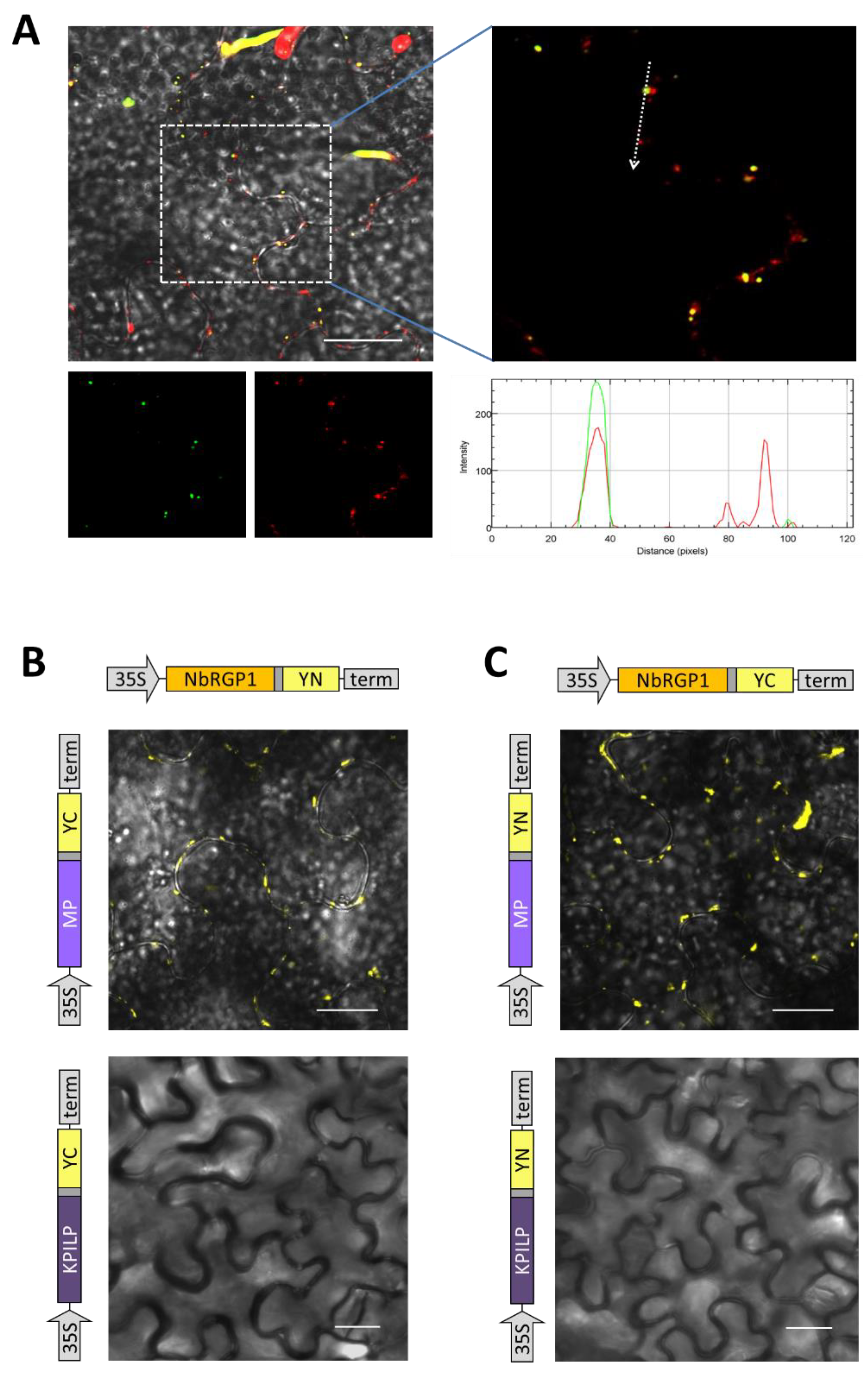
Figure 6). Furthermore, using the BiFC system, we demonstrated that both reciprocal pairs of NbRGP1 and TMV MP translational fuses formed a pair, with the restoration of YFP fluorescence in
N. benthamiana leaves, indicating NbRGP1/MP interactions in vivo (
Figure 8B,C). NbRGP1 and TMV MP likely interact on Golgi membranes. In this study, we did not investigate in detail the intracellular localization of NbRGP5 or its ability to interact with either TMV MP or other class 1 NbRGPs. However, we showed that NbRGP5 does not affect callose depositions and does not suppress TMV intercellular transport. Our phylogenetic analysis also demonstrated that NbRGP5 belongs to class 2, which lack mutase activity. Regarding the biological role of NbRGP5, we can assume that NbRGP5 participates in the formation of heteroprotein complexes with enzymatically active NbRGPs, as it was shown for RGPs of other species [
40], which may determine its stabilizing function in these interactions. This is why its expression increases in response to the viral infection without a direct negative impact on the virus. Therefore, the role of class 2 RGPs such as NbRGP5 in the development of viral infection is to be further investigated.
All our results are in line with the hypothesis accounting for the protective role of NbRGPs in interaction between
N. benthamiana and TMV (
Figure 9) and are consistent with the previously reported data on RGP antiviral action [
15,
16]. The viral infection activates the expression of all four
NbRGPs. We assume that the antiviral mechanism consists of blocking intercellular transport of the virus: firstly, via the callose-mediated mechanism and secondly, via a direct interaction with MP, which probably interferes with MP transport function.
In this article, we presented convincing evidence that NbRGP1 and 2 stimulate callose deposition and NbRGP1 interacts with TMV MP. Being very similar in terms of amino acid and nucleotide sequences, NbRGP1 and 2 could have the same function in a cell and act in a similar way by stimulating callose deposition and potentially blocking MP function by interacting with it. Although we do not investigate the intracellular localization of NbRGP2 and its ability to interact with MP, its antiviral effect seems obvious and results are presented that demonstrate and confirm this function.
Therefore, based on the results of this study, we can conclude that class 1 NbRGP1 and NbRGP2, induced in response to TMV infection, suppress the intercellular transport of the virus and its reproduction. NbRGP1 stimulates the “closing” of plasmodesmata via the callose-mediated mechanism and interacting with TMV MP probably inhibits the intercellular transport of viral RNA. To summarize, we can state that NbRGPs play the role of antiviral cellular factors.
4. Materials and Methods
4.1. Plant Growth Conditions
Wild-type N. benthamiana plants were grown in soil in a controlled environment chamber under a 16 h/8 h day/night cycle.
4.2. Plasmid Constructs
The sequences encoding NbRGP1, 2 and 5 were amplified from N. benthamiana cDNA using the F1/R1, F2/R2, and F5/R5 primer pairs, respectively. Acc65I and SalI recognition sites at the 5′ and 3′ ends were introduced in the resulting fragment. NbRGP3 sequence was amplified with the F3/R3 primer pair. The resulting PCR product was flanked with NruI and SalI recognition sites. The PCR products were digested with the Acc65I-SalI or NruI-SalI restriction enzymes and cloned into the pCambia1300-based vector containing the 35S promoter and terminator (pCambia-35S) and digested with Acc65I/SalI or NruI/SalI. Therefore, we obtained 35S-NbRGP1, 35S-NbRGP2, 35S-NbRGP3, and 35S-NbRGP5 genetic constructs. The 35S-NbRGP1:GFP construct was obtained as follows. Using the F1/R4 primer pair, a fragment encoding NbRGP1 without a stop codon and flanked by Acc65I/BamHI recognition sites was obtained. The fragment encoding GFP without a start codon and flanked by BamHI/SalI recognition sites was produced using F9/R9 pair of primers. The NbRGP1 fragment without a stop codon, together with a GFP fragment, was ligated into a pCambia-35S vector digested with Acc65I/SalI enzymes.
To obtain plasmids for the BiFC system, the fragment encoding 154 YFP N-terminal amino acids (designated YN) and the fragment encoding 86-aa C-terminal part of YFP (YC) were amplified using the F4/R11 and F6/R12 primer pairs, respectively. Both fragments contained an ApaI recognition site instead of a start codon and SalI downstream of the stop codon. 35S-NbRGP1:YN and 35S-NbRGP1:YC were constructed by substitution of YN or YC fragments for GFP in 35S-NbRGP1:GFP plasmid using ApaI and SalI sites.
Sequences encoding AtGONST4 (NM_122005.4) without a stop codon were amplified from A. thaliana Col1 cDNA using the F11/R14 primer pairs. The fragment encoding RFP without a start codon and flanked by BamHI/SalI recognition sites was produced using F12/R15 pair of primers, respectively. The AtGONST4 PCR product was digested with Acc65I/BamHI and together with RFP fragment flanked with BamHI/SalI inserted into pCambia-35S via Acc65I and SalI sites resulting in 35S-AtGONST4:RFP.
Plasmids encoding MP translational fuses with RFP and YN or YC were obtained using the same algorithm. The TMV MP fragment was produced using a F7/R13 primer pair. The PCR product was digested with Acc65I/BamHI enzymes and cloned together with a fragment encoding the corresponding fluorescent tag into pCambia-35S via Acc65I and SalI sites resulting in the set of constructs: 35S-MP:YN/YC and 35S-MP:RFP.
Plasmids encoding KPILP translational fuses with YN or YC were obtained by amplification of the KPILP fragment using the F13/R16 pair of primers with Acc65I/BamHI flanking sites and 35S-NbKPILP(ACG) plasmid [
41] as a template. The PCR product was digested with the Acc65I-BamHI restriction enzymes and cloned into the 35S-NbRGP1:YN/YC plasmids replacing the NbRGP1 fragment.
To obtain a plasmid for NbRGP1 recombinant protein production in E.coli cells, a PCR fragment of NbRGP1 without a start codon was obtained with the F8/R1 primer pair and Acc65I and SalI recognition sites at the ends of the resulting fragment. The PCR product was digested with the Acc65I-SalI restriction enzymes and cloned into the pQE-30 (QIAGEN, Netherlands) vector containing the 6xHis-tag at the N-terminus and digested with Acc65I/SalI. Therefore, we obtained a 6xHis-NbRGP1 genetic construction.
GeneBank acc. numbers for NbRGPs: NbRGP1, LR961922; NbRGP2, OM056463; NbRGP3, OM056464; NbRGP5, OM056465.
The oligonucleotide sequences are listed in
Table S1.
4.3. Agroinfiltration Experiments
A. tumefaciens strain GV3101 was transformed with individual binary constructs and grown at 28 °C in LB medium supplemented with 50 mg/L rifampicin, 25 mg/L gentamycin, and 50 mg/L carbenicillin/kanamycin. An Agrobacterium overnight culture was diluted in an agrobuffer containing 10 mM MES (pH 5.5) and 10 mM MgSO4 and adjusted to a final OD600 of 0.1. In experiments with viral vector spread, the final OD600 for TMV:GFP was 0.005, which allowed to obtain individual transformed cells within the infiltrated area. Agroinjection was performed on almost fully expanded N. benthamiana leaves that were still attached to the intact plant. A bacterial suspension was infiltrated into the leaf tissue using a 2 mL syringe, after which the plants were grown under greenhouse conditions at 24 °C with a 16 h/8 h light/dark photoperiod.
4.4. Plasmolysis
N. benthamiana leaves were infiltrated with a 0.25 M mannitol solution three days after agroinfiltration. About 5–10 min after mannitol solution injection, leaf sections were excised and examined using confocal microscopy.
4.5. Aniline Blue Callose Staining and Quantification
To visualize Pd-located callose,
N. benthamiana leaves transiently expressing 35S-NbRGPs, as well as control leaves, were infiltrated with aniline blue solution (0.1% aniline blue (Sigma Aldrich, Burlington, VT, USA) in 0.01 M K
3PO
4 at pH 12. Then, the leaves were incubated in the dark at room temperature for 15 min before imaging using a Nikon C2 laser scanning confocal microscope. Quantification of the stained callose fluorescence was performed as described by Zavaliev and Epel [
33].
4.6. GFP, YFP, and RFP Imaging
GFP fluorescence in the inoculated leaves was monitored by illumination with a handheld UV (366 nm) source. The TMV:GFP foci were analyzed at 3 dpi. YFP fluorescence was detected using an AxioVert 200M microscope (Carl Zeiss, Jena, Germany) equipped with AxioCam MRc digital camera. GFP- and RFP-containing fusion proteins were imaged with Nikon C2 confocal laser scanning microscope. The excitation and detection wavelengths for GFP and YFP were 487 nm and 525 nm, respectively; the excitation and emission wavelengths for RFP were 561 nm and 625 nm, respectively. The intracellular distribution of fluorescent proteins was imaged 72 h after infiltration.
4.7. Genomic DNA Extraction
N. benthamiana mature leaves were frozen in liquid nitrogen and ground into a fine powder. Genomic DNA was extracted using Diatom DNA Prep kit (Galart-Diagnosticum, Moscow, Russia) in accordance with the manufacturer’s protocol.
4.8. RNA Extraction and cDNA Synthesis
The total RNA was extracted from plant tissues using TriReagent (MRC, Houston, TX, USA) in accordance with the manufacturer’s instructions. The RNA concentration was determined using a Nanodrop ND-1000 spectrophotometer (Isogen Life Sciences, Utrecht, The Netherlands). For the synthesis of first-strand cDNA, 0.1 mg of random hexamers and 0.1 mg of oligo-dT primer were added to 2 µg of total RNA to obtain cDNA by reverse transcription performed using Superscript II reverse transcriptase (Invitrogen, Waltham, MA, USA) in accordance with the manufacturer’s protocol.
4.9. Quantitative Real-Time PCR (qRT-PCR)
Quantitative real-time PCR was carried out using the iCycler iQ real-time PCR detection system (Bio-Rad, Hercules, CA, USA). Target genes were detected using sequence-specific primers (
Table S2) and Eva Green master mix (Syntol, Moscow, Russia) in accordance with the manufacturer’s instructions. Reference genes were detected using the primers to the 18S rRNA gene and the protein phosphatase 2A gene (PP2A). Each sample was run in triplicate, and a non-template control was added to each run. A minimum of five biological replicates were performed. The qRT-PCR results were evaluated using the Pfaffl algorithm [
42].
4.10. Renatured Blot Overlay Binding Assay
Recombinant 6His-NbRGP1 protein was produced in Escherichia coli (strain SG13009) and purified via a Ni-NTA affinity chromatography. Briefly, the cells were transfected with the pQE30-based 6His-NbRGP1 construct. An E. coli overnight culture was diluted 10 times with liquid LB medium supplemented with kanamycin 25 mg/L and ampicillin 100 mg/L and grown at 37 °C to OD600 = 0.6. After addition of isopropyl-β-D-1-thiogalactopyranoside to a final concentration of 1 mM, the culture was incubated for four hours at 37 °C. E. coli cells were collected by centrifugation at 4000× g for 20 min at 4 °C. 6His-NbRGP1 recombinant protein was purified from the obtained cells by metal-chelate affinity chromatography on Ni-NTA agarose in accordance with “The QIAexpressionist™” handbook protocol.
The renatured blot overlay assay was performed in accordance with [
43], with modifications. Briefly, the analyzed recombinant 6His-NbRGP1 or 6His-MP were resolved in 10% SDS-PAAG and transferred onto a polyvinylidene difluoride membrane (GE Healthcare, Chicago, IL, USA). The membrane was incubated in buffer A (30 mM Tris–HCl pH 7.4, 0.05% Tween-20) for 15 min to remove residual SDS. For immobilized protein denaturation, the membrane was incubated in a denaturing buffer (7 M guanidine hydrochloride, 2 mM EDTA, 50 mM DTT, 50 mM Tris–HCl pH 8.0) for two hours. After the membrane was washed in tTBS (140 mM NaCl, 30 mM Tris–HCl pH 7.4, 0.1% Tween-20), it was incubated overnight at +4 C⁰ in the renaturing buffer (140 mM NaCl, 10 mM Tris–HCl pH 7.4, 2 mM EDTA, 0.05% milk, 0.1% Tween-20, 2 mm DTT). After renaturation of the immobilized protein, the membrane was incubated in the renaturing buffer, supplemented with the potential partner protein added to a concentration of 10 μg/mL for 90 min. Then, the membrane was rinsed in tTBS and incubated in the blocking solution (2.5% powdered skim milk in tTBS) for 1 h. Next, the membrane was incubated in tTBS supplemented with 0.5% skim milk and rabbit polyclonal antibodies against the protein that was in the solution in the previous step—either 6His-NbRGP1 or 6His-MP (Almabion, Voronezh, Russia). Anti-rabbit antibodies conjugated to horseradish peroxidase (Imtek, Moscow, Russia) were used as secondary antibodies. The bands were visualized using a chemiluminescence ECL kit (GE Healthcare, Chicago, IL, USA) and ChemiDoc XRS+ imaging system (Bio-Rad, Hercules, CA, USA).
4.11. Statistical Analysis
The data were analyzed either by Student’s t-test or by one-way ANOVA, as indicated in figure captions. The significance of difference between groups was assessed using Tukey’s honestly significant difference (HSD) test at p < 0.05 level or Student’s t-test. In all histograms, y-axis error bars represent the standard error of the mean values.
4.12. Homology Search and Phylogenetic Analysis
The homology search was conducted using the amino acid sequences for RGP proteins from the UniProt database (
https://www.uniprot.org/ (accessed on 9 July 2021)) for the following species:
A. thaliana (AtRGP1, Q9SRT9; AtRGP2, Q9LFW1; AtRGP3, O22666; AtRGP4, Q9LUE6; AtRGP5, Q9FFD2),
O. sativa (OsRGP1, Q8H8T0; OsRGP2, Q7FAY6; OsRGP3, Q6Z4G3),
H. vulgare (HvUAM1, A0A0U2GJJ3; HvUAM2, A0A0U2GJL1; HvUAM3, A0A0U2GJ84; HvUAM4, A0A0U2GJM5),
P. sativum (PsRGP1, O04300),
S. tuberosum (StRGP1, Q9SC19; StRGP2, Q8RU27), and
Z. mays (ZmRGP1, P80607). Multiple alignment was generated using ClustalW [
25]. The phylogenetic tree was obtained using the maximum likelihood method and the model of Lebre and Gascuel [
26]. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Bootstrap values are calculated for 10,000 replications. Evolutionary analyses were conducted in MEGA X [
27].

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








