
Age-Related Cognitive Decline, Focus on Microbiome: A Systematic Review and Meta-Analysis
 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Methods
2.1. Search Strategy
2.2. Inclusion and Exclusion Criteria
2.3. Study Selection and Data Extraction
2.4. Outcome Measures
2.5. Statistical Analysis
2.6. Risk of Bias and Quality Assessment
3. Results
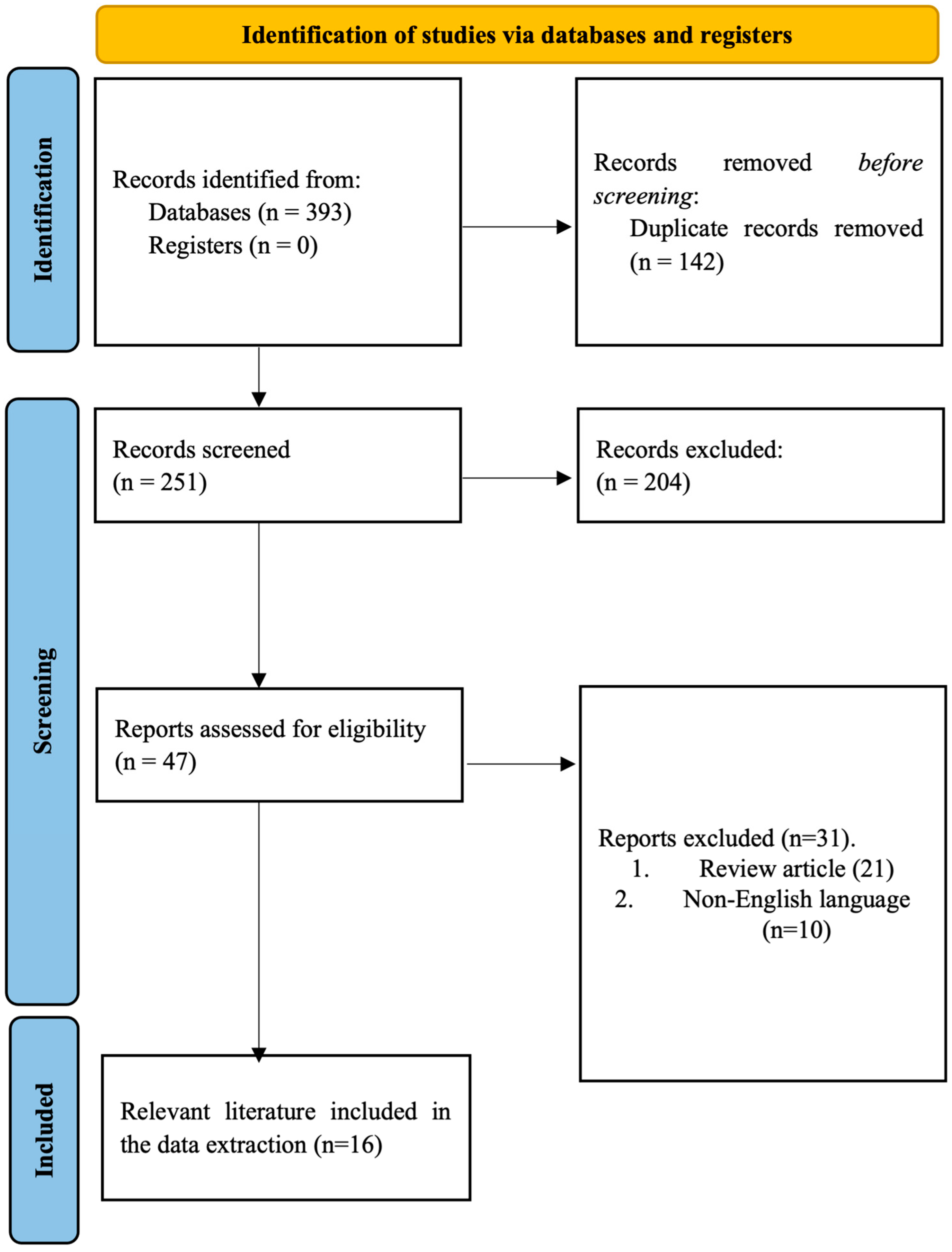
3.1. Identification of the Literature
3.2. Study Characteristics
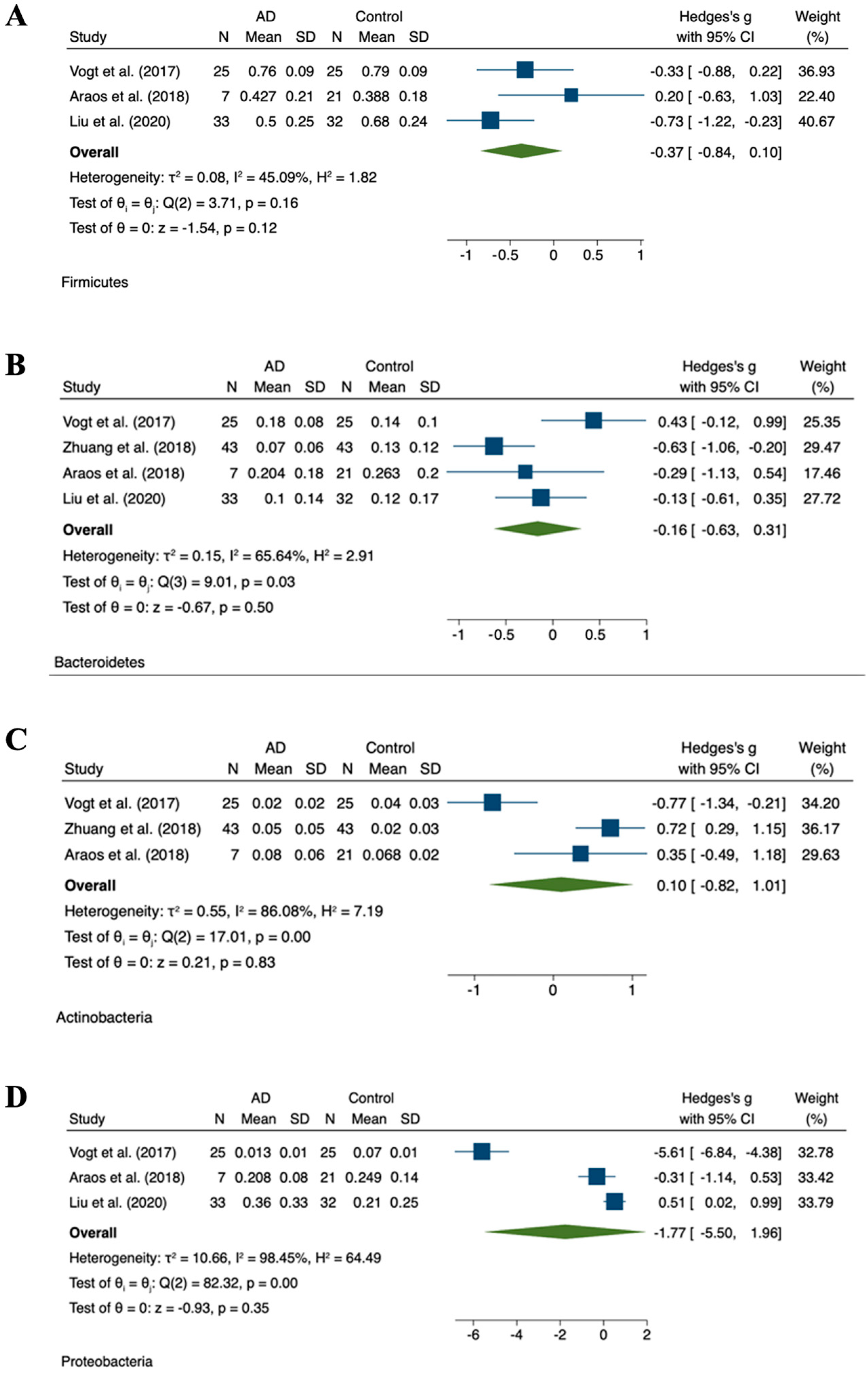
3.3. Relative Abundance


{kind=link}
{kind=link}
| Author | Study Design | Study Subjects | Biospecimen | Microbiological Analysis | Findings |
|---|---|---|---|---|---|
| Cattaneo et al. [51] | n = 73 cognitively impaired patients (n = 40 with and n = 33 without brain amyloidosis), n = 10 HC | Fecal samples | Cognitively impaired patients with brain amyloidosis showed lower abundance of Eubacterium rectale and higher abundance of Eschierichia/Shigella as compared to both HCs and cognitively impaired patients without brain amyloidosis. | ||
| Minato et al. [52] | Prospective cohort | n = 36 PD (n = 18 deteriorated and n = 18 stable groups based on the degree of worsening of total UPDRS scores in 2 years) | Fecal samples | qRT-PCR of bacterial 16S or 23S rRNA | Low Bifidobacterium and Bacteroides fragilis at year 0 were associated with worsening of UPDRS scores in 2 years. Low Bifidobacterium at year 0 was associated with worsening of hallucinations/delusions in 2 years. Low B. fragilis at year 0 was associated with worsening of motivation/initiative in 2 years. The deteriorated group had lower Bifidobacterium, B. fragilis, and Clostridium leptium than the stable group at year 0. |
| Vogt et al. [41] | n = 25 AD, n = 25 HC | Fecal samples | 16SrRNA sequencing | A decreased level of Firmicutes and Bifidobacteria and an increased level of Bacteroidetes in the fecal microbiota of dementia patients as compared to the controls. | |
| Araos et al. [48] | Retrospective analysis | n = 362 advanced dementia | Fecal samples | 16SrRNA sequencing | Among this patient population, the overall microbiome diversity was substantially lower than reported values of among HCs. |
| Nguyen et al. [53] | n = 4 AD | Fecal samples | 16SrRNA genes sequencing | A remarkable variety amongst the small group of butyrate-producing bacteria was observed in the gut of elderly Japanese patients diagnosed with AD. | |
| Saji et al. [54] | Cross-sectional | n = 34 dementia, n = 94 without dementia | Fecal samples | T-RFLP analysis | The number of Bacteroides (enterotype I) was lower and the number of “other” bacteria (enterotype III) was higher in dementia as compared to non-dementia patients. Multivariable analyses showed that the populations of enterotype I and enterotype III bacteria were strongly associated with dementia, independent of the traditional dementia biomarkers. |
| Qian et al. [55] | n = 45 PD, n = 45 HC | Fecal samples | High-throughput iluminia miseq sequencing targeting the V3-V4 region of 16S rRNA gene | Abundance of Bifidobacterium, Butyricicoccus, and Clostridium XIVb in gut microbiota was negatively correlated with the presence of cognitive impairment in patients with PD. | |
| Zhuang et al. [50] | n = 43 AD, n = 43 HC | Fecal samples | 16S rRNA sequencing | Several bacteria taxa in AD patients were different from those in the controls at taxonomic levels, such as Bacteroides, Actinobacteria, Ruminococcus, Lachnospiraceae, and Selenomonadales. | |
| Haran et al. [56] | Prospective cohort | n = 51 no dementia n = 24 AD n = 33 other dementia types | Fecal samples | Shotgun metagenomics and mixed modeling rather than 16S rRNA | Increased proportions of Bacteroides spp., Alistipes spp., Odoribacter spp., and Barnesiella spp. and decreased proportions of Lachnoclostridium spp. were present in AD elders, while increased proportions of Odoribacter spp. and Barnesiella spp. and decreased proportions of Eubacterium spp., Roseburia spp., Lachnoclostridium spp., and Collinsella spp. were seen in elders with other dementia types. |
| Liu et al. [49] | Prospective and cross-sectional | n = 33 AD n = 32 pre-onset stage aMCI. n = 32 HC | Fecal samples | 16S rRNA miseq sequencing | Proportion of phylum firmicutes was significantly reduced, whereas Proteobacteria was highly enriched in the AD as compared to HC. Similar alterations were observed at the order, class, and family levels of these two phyla. Gammaproteobacteria, Enterobacteriales, and Enterobacteriaceae showed a progressive enriched prevalence from HC to aMCI and AD patients. |
| Saji et al. [57] | Cross-sectional | n = 61 MCI without dementia n = 21 NC function | Fecal samples | T-RFLP analysis | Patients with MCI had a higher prevalence of Bacteroides and more likely to present with white matter hyperintensity and high voxel- based specific regional analysis system for AD. A multivariable logistic regression analysis revealed that a greater prevalence of Bacteroides was independently associated with MCI. |
| Ren et al. [58] | Cross-sectional | n = 13 PD with MCI (PD-MCI) n = 14 PD with NC (PD-NC) n = 13 HC | Fecal samples | 16S rRNA sequencing and gas chromatography-mass spectrometry | Compared with HC and patients with PD-NC, the gut microbiota of patients with PD-MCI was significantly altered, particularly manifesting in enriched genera from the Porphyromonadaceae family and decreased the abundance of the genera Blautia and Ruminococcus. |
| Liu et al. [59] | Longitudinal | n = 30 PSCI n = 35 non-PSCI | Fecal samples | 16SrRNA gene sequencing and gas chromatography- mass spectrometry | PSCI patients had disturbed microbial composition, and corresponding metabolites compared with non-PSCI patients. Increased Fusobacterium and deficiency of microbial metabolized SCFAs were significantly associated with PSCI. |
| Scheperjans et al. [60] | Cross-sectional | n = 72 PD n = 72 HC | Fecal samples | 16S rRNA miseq sequencing | On average, the abundance of Prevotellaceae in the feces of PD patients was reduced by 77.6% as compared to the controls. Relative abundance of Prevotellaceae of 6.5% or less had 86.1% sensitivity and 38.9% specificity for PD. |
| Cerroni et al. [61] | Longitudinal | N = 18 PD N = 18 HC | Fecal samples | 16S rRNA amplicons analysis | No differences in the gut microbiota (beta diversity) structure at the follow-up in both PD patients and HC were observed, which remained stable for both patients and the controls. These results suggest that the gut microbiota may remain stable over a period of 14 months. |
| Unger et al. [62] | Longitudinal | n = 34 PD n = 34 HC | Fecal samples | Quantitative PCR | Fecal SCFA concentrations were significantly reduced in PD patients as compared to the controls. The bacterial phylum Bacteroidetes and the bacterial family Prevotellaceae were reduced, Enterobacteriaceae were more abundant in fecal samples from PD patients as compared to matched controls. |
3.4. Quality Appraisal
4. Discussion
4.1. Impact of the Microbiome on Parkinson’s Disease
4.2. Influence of the Microbiome on Alzheimer’s Disease and Dementia
4.3. Additional Considerations and Future Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coradduzza, D.; Congiargiu, A.; Chen, Z.; Cruciani, S.; Zinellu, A.; Carru, C.; Medici, S. Humanin and its pathophysiological roles in aging: A systematic review. Biology 2023, 12, 558. [Google Scholar] [CrossRef] [PubMed]
- Shabir, S.; Singh, M.P. The Aging: Introduction, Theories, Principles, and Future Prospective. Anti-Aging Drug Discovery on the Basis of Hallmarks of Aging; Elsevier: Amsterdam, The Netherlands, 2022; pp. 1–17. [Google Scholar]
- Coradduzza, D.; Congiargiu, A.; Chen, Z.; Zinellu, A.; Carru, C.; Medici, S. Ferroptosis and senescence: A systematic review. Int. J. Mol. Sci. 2023, 24, 3658. [Google Scholar] [CrossRef] [PubMed]
- Badal, V.D.; Vaccariello, E.D.; Murray, E.R.; Yu, K.E.; Knight, R.; Jeste, D.V.; Nguyen, T.T. The gut microbiome, aging, and longevity: A systematic review. Nutrients 2020, 12, 3759. [Google Scholar] [CrossRef] [PubMed]
- Husted, K.L.S.; Brink-Kjær, A.; Fogelstrøm, M.; Hulst, P.; Bleibach, A.; Henneberg, K.-Å.; Sørensen, H.B.D.; Dela, F.; Jacobsen, J.C.B.; Helge, J.W. A model for estimating biological age from physiological biomarkers of healthy aging: Cross-sectional study. JMIR Aging 2022, 5, e35696. [Google Scholar] [CrossRef] [PubMed]
- Islam, N.; Jdanov, D.A. Age and Sex Adjustments are Critical when Comparing Death Rates; British Medical Journal Publishing Group: London, UK, 2023. [Google Scholar]
- Quirós, P.; Fernando, S.L. Methodology for the projection of population pyramids based on monte Carlo simulation and genetic algorithms. Appl. Intell. 2023, 53, 18989–19006. [Google Scholar] [CrossRef]
- Azamatova, T. Inclusion of senior citizens in lifelong learning to reduce the negative impact of population aging in Latvia society. In Society Integration Education Proceedings of the International Scientific Conference, Novi Sad, Serbia, 16–17 November 2023; Tālrunis: Rezekne, Latvija; pp. 315–505.
- World Health Organization. Regional Framework on Healthy Ageing (2018–2022); World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Coradduzza, D.; Cruciani, S.; Arru, C.; Garroni, G.; Pashchenko, A.; Jedea, M.; Zappavigna, S.; Caraglia, M.; Amler, E.; Carru, C. Role of miRNA-145, 148, and 185 and stem cells in prostate cancer. Int. J. Mol. Sci. 2022, 23, 1626. [Google Scholar] [CrossRef]
- Coradduzza, D.; Azara, E.; Medici, S.; Arru, C.; Solinas, T.; Madonia, M.; Zinellu, A.; Carru, C. A preliminary study procedure for detection of polyamines in plasma samples as a potential diagnostic tool in prostate cancer. J. Chromatogr. B 2021, 1162, 122468. [Google Scholar] [CrossRef]
- Angius, A.; Pira, G.; Cossu-Rocca, P.; Sotgiu, G.; Saderi, L.; Muroni, M.R. Deciphering clinical significance of BCL11A isoforms and protein expression roles in triple-negative breast cancer subtype. J. Cancer Res. Clin. Oncol. 2023, 149, 3951–3963. [Google Scholar] [CrossRef]
- Angulo, J.; El Assar, M.; Álvarez-Bustos, A.; Rodríguez-Mañas, L. Physical activity and exercise: Strategies to manage frailty. Redox Biol. 2020, 35, 101513. [Google Scholar] [CrossRef]
- Du, Y.; Mao, L.; Wang, Z.; Yan, K.; Zhang, L.; Zou, J. Osteopontin-The stirring multifunctional regulatory factor in multisystem aging. Front. Endocrinol. 2022, 13, 1014853. [Google Scholar] [CrossRef]
- Gallagher, A.; Bulteau, C.; Cohen, D.; Michaud, J.L. Neurocognitive Development: Disorders and Disabilities; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Park, D.C.; Polk, T.A.; Mikels, J.A.; Taylor, S.F.; Marshuetz, C. Cerebral aging: Integration of brain and behavioral models of cognitive function. Dialog. Clin. Neurosci. 2022. [Google Scholar] [CrossRef]
- Oosterhuis, E.J.; Slade, K.; May, P.J.C.; Nuttall, H.E. Toward an Understanding of Healthy Cognitive Aging: The Importance of Lifestyle in Cognitive Reserve and the Scaffolding Theory of Aging and Cognition. J. Gerontol. Ser. B 2023, 78, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Coradduzza, D.; Garroni, G.; Congiargiu, A.; Balzano, F.; Cruciani, S.; Sedda, S.; Nivoli, A.; Maioli, M. MicroRNAs, Stem Cells in Bipolar Disorder, and Lithium Therapeutic Approach. Int. J. Mol. Sci. 2022, 23, 10489. [Google Scholar] [CrossRef] [PubMed]
- Gareau, M.G. Chapter Eleven—Cognitive Function and the Microbiome. In International Review of Neurobiology; Cryan, J.F., Clarke, G., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 227–246. [Google Scholar]
- Gustavsson, A.; Norton, N.; Fast, T.; Frölich, L.; Georges, J.; Holzapfel, D.; Kirabali, T.; Krolak-Salmon, P.; Rossini, P.M.; Ferretti, M.T.; et al. Global estimates on the number of persons across the Alzheimer’s disease continuum. Alzheimer’s Dement. 2023, 19, 658–670. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, E.R.; Constantinescu, R.; Thompson, J.P.; Biglan, K.M.; Holloway, R.G.; Kieburtz, K.; Marshall, F.J.; Ravina, B.M.; Schifitto, G.; Siderowf, A.; et al. Projected number of people with Parkinson disease in the most populous nations, 2005 through 2030. Neurology 2007, 68, 384–386. [Google Scholar] [CrossRef] [PubMed]
- Strasser, B.; Ticinesi, A. Intestinal microbiome in normal ageing, frailty and cognition decline. Curr. Opin. Clin. Nutr. Metab. Care 2023, 26, 8–16. [Google Scholar] [CrossRef]
- Carter, C.S. A “gut feeling” to create a 10th hallmark of aging. J. Gerontol. Ser. A 2021, 76, 1891–1894. [Google Scholar] [CrossRef]
- Shi, X.; Ma, T.; Sakandar, H.A.; Menghe, B.; Sun, Z. Gut microbiome and aging nexus and underlying mechanism. Appl. Microbiol. Biotechnol. 2022, 106, 5349–5358. [Google Scholar] [CrossRef]
- Sharma, R. Emerging interrelationship between the gut microbiome and cellular senescence in the context of aging and disease: Perspectives and therapeutic opportunities. Probiotics Antimicrob. Proteins 2022, 14, 648–663. [Google Scholar] [CrossRef]
- Kim, M.; Benayoun, B.A. The microbiome: An emerging key player in aging and longevity. Transl. Med. Aging 2020, 4, 103–116. [Google Scholar] [CrossRef]
- Nagpal, R.; Mainali, R.; Ahmadi, S.; Wang, S.; Singh, R.; Kavanagh, K.; Kitzman, D.W.; Kushugulova, A.; Marotta, F.; Yadav, H. Gut microbiome and aging: Physiological and mechanistic insights. Nutr. Healthy Aging 2018, 4, 267–285. [Google Scholar] [PubMed]
- Wilmanski, T.; Diener, C.; Rappaport, N.; Patwardhan, S.; Wiedrick, J.; Lapidus, J.; Earls, J.C.; Zimmer, A.; Glusman, G.; Robinson, M. Gut microbiome pattern reflects healthy ageing and predicts survival in humans. Nat. Metabol. 2021, 3, 274–286. [Google Scholar] [CrossRef] [PubMed]
- Juárez-Fernández, M.; Porras, D.; García-Mediavilla, M.V.; Román-Sagüillo, S.; González-Gallego, J.; Nistal, E.; Sánchez-Campos, S. Aging, gut microbiota and metabolic diseases: Management through physical exercise and nutritional interventions. Nutrients 2020, 13, 16. [Google Scholar] [PubMed]
- Biagi, E.; Candela, M.; Turroni, S.; Garagnani, P.; Franceschi, C.; Brigidi, P. Ageing and gut microbes: Perspectives for health maintenance and longevity. Pharmacol. Res. 2013, 69, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, T.S.; Shanahan, F.; O’Toole, P.W. The gut microbiome as a modulator of healthy ageing. Nat. Revi. Gastroenterol. Hepatol. 2022, 19, 565–584. [Google Scholar]
- Li, Y.; Ning, L.; Yin, Y.; Wang, R.; Zhang, Z.; Hao, L.; Wang, B.; Zhao, X.; Yang, X.; Yin, L. Age-related shifts in gut microbiota contribute to cognitive decline in aged rats. Aging 2020, 12, 7801. [Google Scholar] [CrossRef]
- Sittipo, P.; Choi, J.; Lee, S.; Lee, Y.K. The function of gut microbiota in immune-related neurological disorders: A review. J. Neuroinflamm. 2022, 19, 1–17. [Google Scholar]
- Gruninger, T.R.; LeBoeuf, B.; Liu, Y.; Garcia, L.R. Molecular signaling involved in regulating feeding and other motivated behaviors. Mol. Neurobiol. 2007, 35, 1–20. [Google Scholar] [CrossRef]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Bienenstock, J.; Dinan, T.G. The probiotic Bifidobacteria infantis: An assessment of potential antidepressant properties in the rat. J. Psychiatr. Res. 2008, 43, 164–174. [Google Scholar] [CrossRef]
- Coradduzza, D.; Ghironi, A.; Azara, E.; Culeddu, N.; Cruciani, S.; Zinellu, A.; Maioli, M.; De Miglio, M.R.; Medici, S.; Fozza, C. Role of Polyamines as Biomarkers in Lymphoma Patients: A Pilot Study. Diagnostics 2022, 12, 2151. [Google Scholar] [CrossRef]
- Coradduzza, D.; Bellu, E.; Congiargiu, A.; Pashchenko, A.; Amler, E.; Necas, A.; Carru, C.; Medici, S.; Maioli, M. Role of Nano-miRNAs in Diagnostics and Therapeutics. Int. J. Mol. Sci. 2022, 23, 6836. [Google Scholar] [PubMed]
- Komanduri, M.; Gondalia, S.; Scholey, A.; Stough, C. The microbiome and cognitive aging: A review of mechanisms. Psychopharmacology 2019, 236, 1559–1571. [Google Scholar] [PubMed]
- Tooley, K.L. Effects of the Human Gut Microbiota on Cognitive Performance, Brain Structure and Function: A Narrative Review. Nutrients 2020, 12, 3009. [Google Scholar] [PubMed]
- Zhu, X.; Li, B.; Lou, P.; Dai, T.; Chen, Y.; Zhuge, A.; Yuan, Y.; Li, L. The relationship between the gut microbiome and neurodegenerative diseases. Neurosci. Bull. 2021, 37, 1510–1522. [Google Scholar] [PubMed]
- Vogt, N.M.; Kerby, R.L.; Dill-McFarland, K.A.; Harding, S.J.; Merluzzi, A.P.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Zetterberg, H.; Blennow, K.; et al. Gut microbiome alterations in Alzheimer’s disease. Sci. Rep. 2017, 7, 13537. [Google Scholar]
- Saji, N.; Niida, S.; Murotani, K.; Hisada, T.; Tsuduki, T.; Sugimoto, T.; Kimura, A.; Toba, K.; Sakurai, T. Analysis of the relationship between the gut microbiome and dementia: A cross-sectional study conducted in Japan. Sci. Rep. 2019, 9, 1008. [Google Scholar]
- Brożek, J.; Akl, E.A.; Alonso-Coello, P.; Lang, D.; Jaeschke, R.; Williams, J.W.; Phillips, B.; Lelgemann, M.; Lethaby, A.; Bousquet, J. Grading quality of evidence and strength of recommendations in clinical practice guidelines: Part 1 of 3. An overview of the GRADE approach and grading quality of evidence about interventions. Allergy 2009, 64, 669–677. [Google Scholar]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; the PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. Ann. Intern. Med. 2009, 151, 264–269. [Google Scholar]
- Wan, X.; Wang, W.; Liu, J.; Tong, T. Estimating the sample mean and standard deviation from the sample size, median, range and/or interquartile range. BMC Med. Res. Methodol. 2014, 14, 135. [Google Scholar] [CrossRef]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar]
- Santos García, D.; García Roca, L.; de Deus Fonticoba, T.; Cores Bartolomé, C.; Naya Ríos, L.; Canfield, H.; Paz González, J.M.; Martínez Miró, C.; Jesús, S.; Aguilar, M. Constipation predicts cognitive decline in Parkinson’s disease: Results from the COPPADIS cohort at 2-year follow-up and comparison with a control group. J. Park. Dis. 2022, 12, 315–331. [Google Scholar]
- Araos, R.; Andreatos, N.; Ugalde, J.; Mitchell, S.; Mylonakis, E.; D’Agata, E.M.C. Fecal Microbiome Among Nursing Home Residents with Advanced Dementia and Clostridium difficile. Dig. Dis. Sci. 2018, 63, 1525–1531. [Google Scholar]
- Liu, S.; Gao, J.; Zhu, M.; Liu, K.; Zhang, H.L. Gut Microbiota and Dysbiosis in Alzheimer’s Disease: Implications for Pathogenesis and Treatment. Mol. Neurobiol. 2020, 57, 5026–5043. [Google Scholar] [PubMed]
- Zhuang, Z.Q.; Shen, L.L.; Li, W.W.; Fu, X.; Zeng, F.; Gui, L.; Lü, Y.; Cai, M.; Zhu, C.; Tan, Y.L.; et al. Gut Microbiota is Altered in Patients with Alzheimer’s Disease. J. Alzheimers Dis. 2018, 63, 1337–1346. [Google Scholar]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, S. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar]
- Minato, T.; Maeda, T.; Fujisawa, Y.; Tsuji, H.; Nomoto, K.; Ohno, K.; Hirayama, M. Progression of Parkinson’s disease is associated with gut dysbiosis: Two-year follow-up study. PLoS ONE 2017, 12, e0187307. [Google Scholar]
- Nguyen, T.T.T.; Fujimura, Y.; Mimura, I.; Fujii, Y.; Nguyen, N.L.; Arakawa, K.; Morita, H. Cultivable butyrate-producing bacteria of elderly Japanese diagnosed with Alzheimer’s disease. J. Microbiol. 2018, 56, 760–771. [Google Scholar] [PubMed]
- Saji, N.; Tsuduki, T.; Murotani, K.; Hisada, T.; Sugimoto, T.; Kimura, A.; Niida, S.; Toba, K.; Sakurai, T. Relationship between the Japanese-style diet, gut microbiota, and dementia: A cross-sectional study. Nutrition 2022, 94, 111524. [Google Scholar]
- Qian, Y.; Yang, X.; Xu, S.; Wu, C.; Song, Y.; Qin, N.; Chen, S.D.; Xiao, Q. Alteration of the fecal microbiota in Chinese patients with Parkinson’s disease. Brain Behav. Immun. 2018, 70, 194–202. [Google Scholar] [PubMed]
- Haran, J.P.; Bhattarai, S.K.; Foley, S.E.; Dutta, P.; Ward, D.V.; Bucci, V.; McCormick, B.A. Alzheimer’s Disease Microbiome Is Associated with Dysregulation of the Anti-Inflammatory P-Glycoprotein Pathway. mBio 2019, 10. [Google Scholar] [CrossRef]
- Saji, N.; Murotani, K.; Hisada, T.; Tsuduki, T.; Sugimoto, T.; Kimura, A.; Niida, S.; Toba, K.; Sakurai, T. The relationship between the gut microbiome and mild cognitive impairment in patients without dementia: A cross-sectional study conducted in Japan. Sci. Rep. 2019, 9, 19227. [Google Scholar] [CrossRef]
- Ren, T.; Gao, Y.; Qiu, Y.; Jiang, S.; Zhang, Q.; Zhang, J.; Wang, L.; Zhang, Y.; Wang, L.; Nie, K. Gut Microbiota Altered in Mild Cognitive Impairment Compared with Normal Cognition in Sporadic Parkinson’s Disease. Front. Neurol. 2020, 11, 137. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kong, C.; Gong, L.; Zhang, X.; Zhu, Y.; Wang, H.; Qu, X.; Gao, R.; Yin, F.; Liu, X.; et al. The Association of Post-Stroke Cognitive Impairment and Gut Microbiota and its Corresponding Metabolites. J. Alzheimers Dis. 2020, 73, 1455–1466. [Google Scholar] [CrossRef] [PubMed]
- Scheperjans, F.; Aho, V.; Pereira, P.A.; Koskinen, K.; Paulin, L.; Pekkonen, E.; Haapaniemi, E.; Kaakkola, S.; Eerola-Rautio, J.; Pohja, M.; et al. Gut microbiota are related to Parkinson’s disease and clinical phenotype. Mov. Disord. 2015, 30, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Cerroni, R.; Pietrucci, D.; Teofani, A.; Chillemi, G.; Liguori, C.; Pierantozzi, M.; Unida, V.; Selmani, S.; Mercuri, N.B.; Stefani, A. Not just a Snapshot: An Italian Longitudinal Evaluation of Stability of Gut Microbiota Findings in Parkinson’s Disease. Brain Sci. 2022, 12, 739. [Google Scholar] [CrossRef]
- Unger, M.M.; Spiegel, J.; Dillmann, K.U.; Grundmann, D.; Philippeit, H.; Bürmann, J.; Faßbender, K.; Schwiertz, A.; Schäfer, K.H. Short chain fatty acids and gut microbiota differ between patients with Parkinson’s disease and age-matched controls. Park. Relat. Disord. 2016, 32, 66–72. [Google Scholar] [CrossRef]
- Van der Geest, S. Akan shit: Getting rid of dirt in Ghana. Anthropol. Today 1998, 14, 8–12. [Google Scholar] [CrossRef]
- Komanduri, M.; Savage, K.; Lea, A.; McPhee, G.; Nolidin, K.; Deleuil, S.; Stough, C.; Gondalia, S. The relationship between gut microbiome and cognition in older Australians. Nutrients 2021, 14, 64. [Google Scholar] [CrossRef]
- Li, Y.; Shi, L.; Yue, L.; Gao, R.; Yu, Z.; Yang, X.; Liu, L.; Zheng, H. Hippocampal gene expression profiling in a rat model of functional constipation reveals abnormal expression genes associated with cognitive function. Neurosci. Lett. 2018, 675, 103–109. [Google Scholar] [CrossRef]
- Jamshed, N.; Lee, Z.-E.; Olden, K.W. Diagnostic approach to chronic constipation in adults. Am. Fam. Phys. 2011, 84, 299–306. [Google Scholar]
- Aljumaah, M.R.; Bhatia, U.; Roach, J.; Gunstad, J.; Peril, M.A.A. The gut microbiome, mild cognitive impairment, and probiotics: A randomized clinical trial in middle-aged and older adults. Clin. Nutr. 2022, 41, 2565–2576. [Google Scholar] [CrossRef] [PubMed]
- Eastwood, J.; Walton, G.; Van Hemert, S.; Williams, C.; Lamport, D. The effect of probiotics on cognitive function across the human lifespan: A systematic review. Neurosci. Biobehav. Rev. 2021, 128, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.B.; Catchlove, S.; Tooley, K.L. Examining the influence of the human gut microbiota on cognition and stress: A systematic review of the literature. Nutrients 2022, 14, 4623. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s disease. Cell 2016, 167, 1469–1480.e12. [Google Scholar] [CrossRef]
- Toh, T.S.; Chong, C.W.; Lim, S.-Y.; Bowman, J.; Cirstea, M.; Lin, C.-H.; Chen, C.-C.; Appel-Cresswell, S.; Finlay, B.B.; Tan, A.H. Gut microbiome in Parkinson’s disease: New insights from meta-analysis. Park. Related Dis. 2022, 94, 1–9. [Google Scholar] [CrossRef]
- Wang, Y. Gut microbiota are related to Parkinson’s disease. J. Neurol. Sci. 2017, 381, 1047. [Google Scholar] [CrossRef]
- Hirayama, M.; Ohno, K. Parkinson’s disease and gut microbiota. Annals Nutr. Metabol. 2021, 77, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Mehanna, M.; AbuRaya, S.; Ahmed, S.M.; Ashmawy, G.; Ibrahim, A.; AbdelKhaliq, E. Study of the gut microbiome in Egyptian patients with Parkinson’s Disease. BMC Microbiol. 2023, 23, 196. [Google Scholar] [CrossRef]
- Chandra, S.; Sisodia, S.S.; Vassar, R.J. The gut microbiome in Alzheimer’s disease: What we know and what remains to be explored. Mol. Neurodegener. 2023, 18, 9. [Google Scholar] [CrossRef]
- Koblinsky, N.D.; Power, K.A.; Middleton, L.; Ferland, G.; Anderson, N.D. The role of the gut microbiome in diet and exercise effects on cognition: A review of the intervention literature. J. Gerontol. Ser. A 2023, 78, 195–205. [Google Scholar] [CrossRef]
- Pei, Y.; Lu, Y.; Li, H.; Jiang, C.; Wang, L. Gut microbiota and intestinal barrier function in subjects with cognitive impairments: A cross-sectional study. Front. Aging Neurosci. 2023, 15, 1174599. [Google Scholar] [CrossRef] [PubMed]
- Bendlin, B.B. Gut microbiome alterations in Alzheimer’s disease. Alzheimer’s Dement. 2022, 18, e069987. [Google Scholar] [CrossRef]
- Jemimah SChabib, C.M.M.; Hadjileontiadis, L.; AlShehhi, A. Gut microbiome dysbiosis in Alzheimer’s disease and mild cognitive impairment: A systematic review and meta-analysis. PLoS ONE 2023, 18, e0285346. [Google Scholar]
- Stanislawski, M.A.; Dabelea, D.; Lange, L.A.; Wagner, B.D.; Lozupone, C.A. Gut microbiota phenotypes of obesity. NPJ Biofilms Microbiomes 2019, 5, 18. [Google Scholar] [CrossRef]
- Aoun, A.; Darwish, F.; Hamod, N. The influence of the gut microbiome on obesity in adults and the role of probiotics, prebiotics, and synbiotics for weight loss. Prev. Nutr. Food Sci. 2020, 25, 113. [Google Scholar] [CrossRef]
- Crovesy, L.; Masterson, D.; Rosado, E.L. Profile of the gut microbiota of adults with obesity: A systematic review. Eur. J. Clin. Nutr. 2020, 74, 1251–1262. [Google Scholar] [CrossRef]
- Zhang, F.; Fan, D.; Huang J-l Zuo, T. The gut microbiome: Linking dietary fiber to inflammatory diseases. Med. Microecol. 2022, 14, 100070. [Google Scholar] [CrossRef]
- Kuwahara, A.; Matsuda, K.; Kuwahara, Y.; Asano, S.; Inui, T.; Marunaka, Y. Microbiota-gut-brain axis: Enteroendocrine cells and the enteric nervous system form an interface between the microbiota and the central nervous system. Biomed. Res. 2020, 41, 199–216. [Google Scholar] [CrossRef]
- O’Riordan, K.J.; Collins, M.K.; Moloney, G.M.; Knox, E.G.; Aburto, M.R.; Fülling, C.; Morley, S.J.; Clarke, G.; Schellekens, H.; Cryan, J.F. Short chain fatty acids: Microbial metabolites for gut-brain axis signalling. Mol. Cell. Endocrinol. 2022, 546, 111572. [Google Scholar] [CrossRef]
- Ahmed, L.A.; Al-Massri, K.F. Gut Microbiota Modulation for Therapeutic Management of Various Diseases: A New Perspective Using Stem Cell Therapy. Curr. Mol. Pharmacol. 2023, 16, 43–59. [Google Scholar] [CrossRef]
- Leo, E.E.M.; Campos, M.R.S. Effect of ultra-processed diet on gut microbiota and thus its role in neurodegenerative diseases. Nutrition 2020, 71, 110609. [Google Scholar]
- Coradduzza, D.; Solinas, T.; Balzano, F.; Culeddu, N.; Rossi, N.; Cruciani, S.; Azara, E.; Maioli, M.; Zinellu, A.; De Miglio, M.R. miRNAs as Molecular Biomarkers for Prostate Cancer. J. Mol. Diag. 2022, 24, 1171–1180. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef] [PubMed]

| Phylum | Major Families | Major Genera |
|---|---|---|
| Firmicutes | Lachnospiraceae, Ruminococcaceae, Clostridiaceae | Faecalibacterium, Roseburia |
| Erysipelotrichaceae | Eubacterium, Clostridium | |
| Bacteroidetes | Bacteroidaceae, Prevotellaceae | Bacteroides, Prevotella |
| Rikenellaceae, Porphyromonadaceae | Parabacteroides, Alistipes | |
| Actinobacteria | Bifidobacteriaceae | Bifidobacterium |
| Coriobacteriaceae | Collinsella | |
| Proteobacteria | Enterobacteriaceae | Escherichia, Klebsiella |
| Desulfovibrionaceae | Desulfovibrio |
| Reference | Methodological Quality | Directness of Evidence | Heterogeneity | Precision of Effect Estimates | Publication Bias | Level of Evidence | Recommendation for Use |
|---|---|---|---|---|---|---|---|
| Cattaneo et al. [51] | Moderate | Moderate | |||||
| Mitano et al. [52] | Moderate | Moderate | |||||
| Vogt et al. [41] | Moderate | Moderate | |||||
| Araos et al. [48] | Low | Low | |||||
| Nguyen et al. [53] | Moderate | Moderate | |||||
| Saji et al. [54] | Moderate | Low | |||||
| Qian et al. [55] | Moderate | Moderate | |||||
| Zhuang et al. [50] | Moderate | Moderate | |||||
| Haran et al. [56] | Moderate | Low | |||||
| Liu et al. [49] | Moderate | Moderate | |||||
| Saji et al. [57] | Moderate | Low | |||||
| Ren et al. [58] | Moderate | Moderate | |||||
| Liu et al. [59] | Moderate | Moderate | |||||
| Scheperjans et al. [60] | Moderate | Moderate | |||||
| Cerroni et al. [61] | Moderate | Moderate | |||||
| Unger et al. [62] | Moderate | Moderate |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coradduzza, D.; Sedda, S.; Cruciani, S.; De Miglio, M.R.; Ventura, C.; Nivoli, A.; Maioli, M. Age-Related Cognitive Decline, Focus on Microbiome: A Systematic Review and Meta-Analysis. Int. J. Mol. Sci. 2023, 24, 13680. https://doi.org/10.3390/ijms241813680
Coradduzza D, Sedda S, Cruciani S, De Miglio MR, Ventura C, Nivoli A, Maioli M. Age-Related Cognitive Decline, Focus on Microbiome: A Systematic Review and Meta-Analysis. International Journal of Molecular Sciences. 2023; 24(18):13680. https://doi.org/10.3390/ijms241813680
Chicago/Turabian StyleCoradduzza, Donatella, Stefania Sedda, Sara Cruciani, Maria Rosaria De Miglio, Carlo Ventura, Alessandra Nivoli, and Margherita Maioli. 2023. "Age-Related Cognitive Decline, Focus on Microbiome: A Systematic Review and Meta-Analysis" International Journal of Molecular Sciences 24, no. 18: 13680. https://doi.org/10.3390/ijms241813680