
The Effect of Skeletal Muscle-Specific Creatine Treatment on ALS NMJ Integrity and Function
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
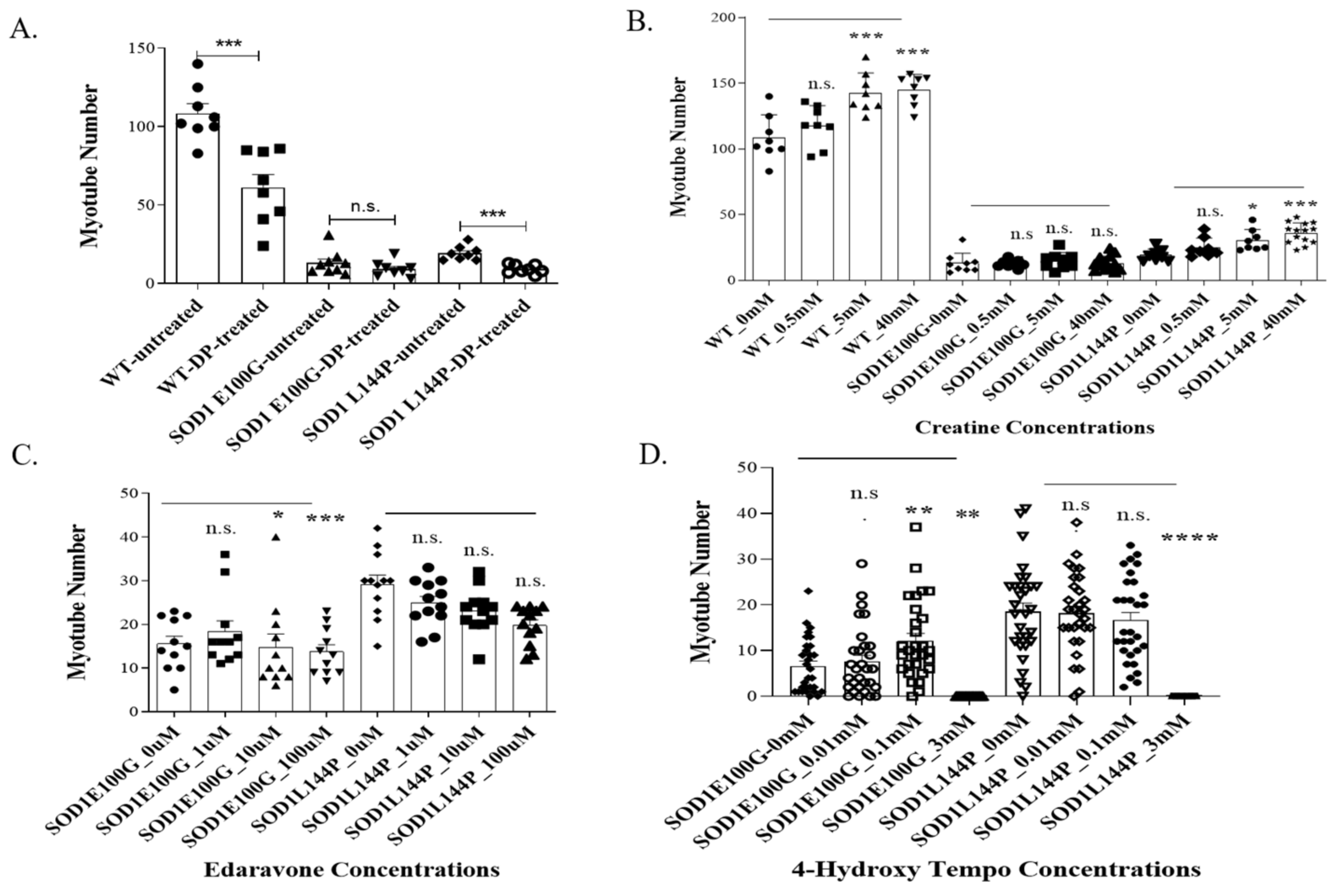
2.1. Effect of Therapeutics on ALS Myotubes
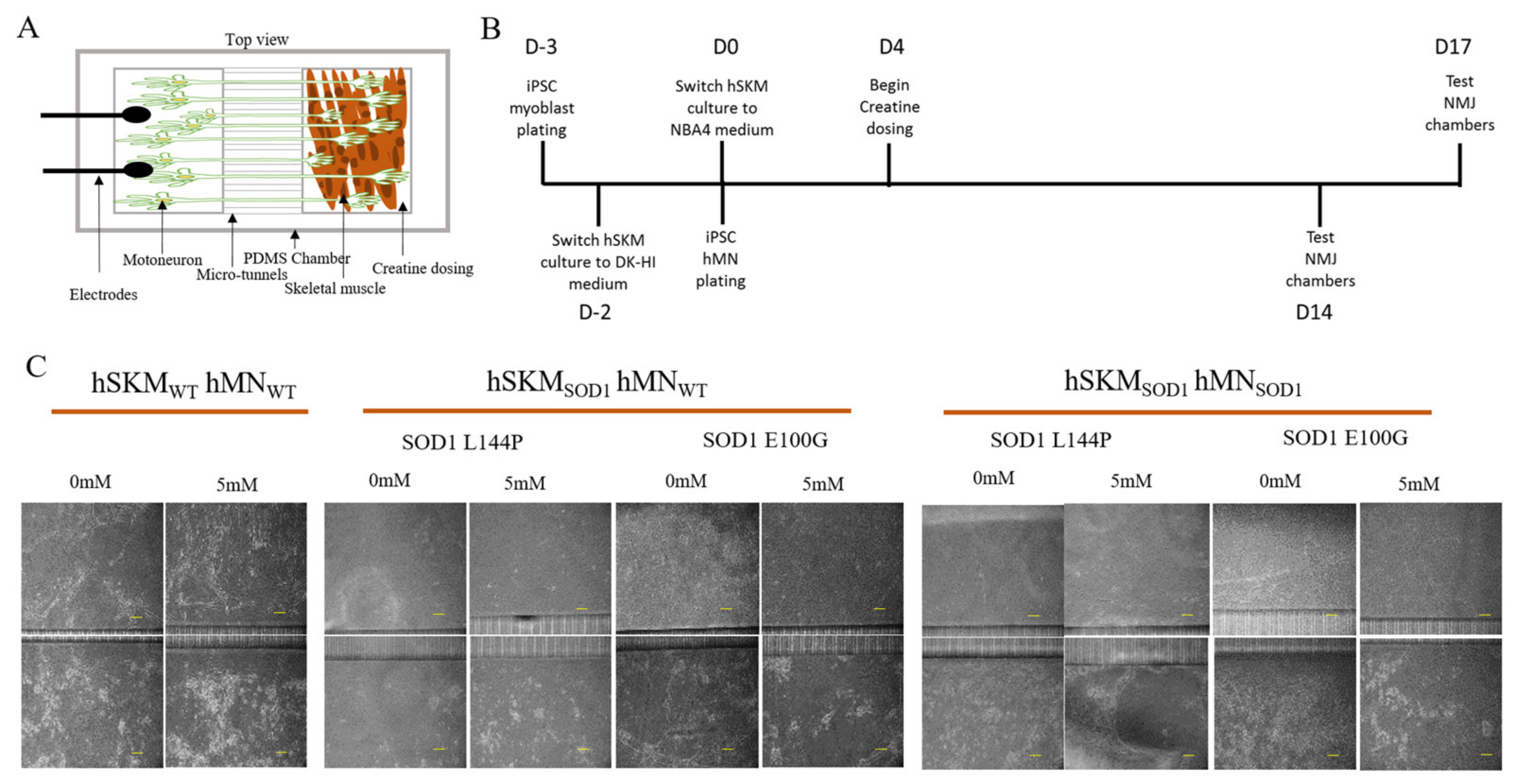
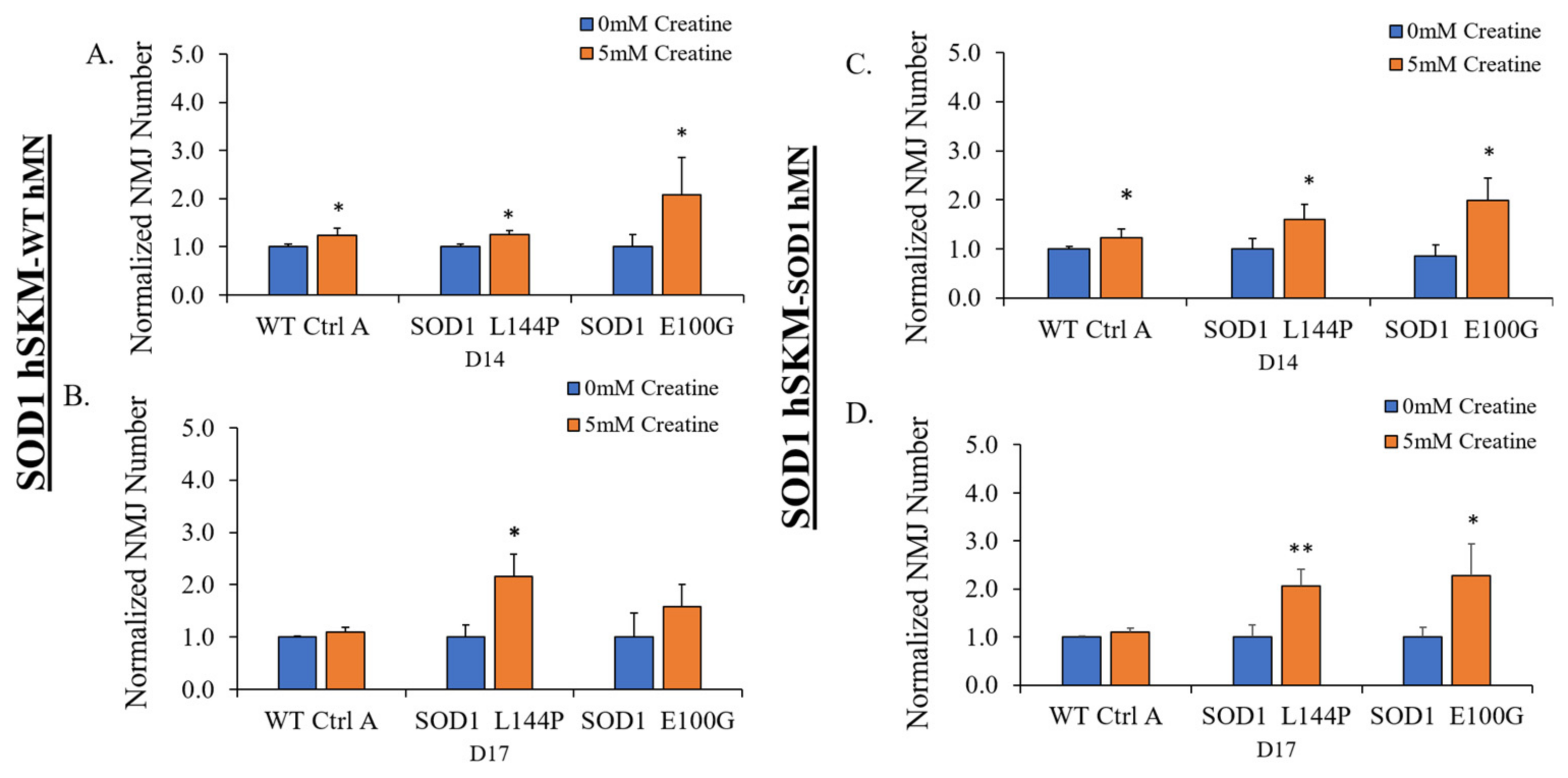
2.2. Effect of ALS hSKM-Specific Creatine Treatment on NMJ Numbers
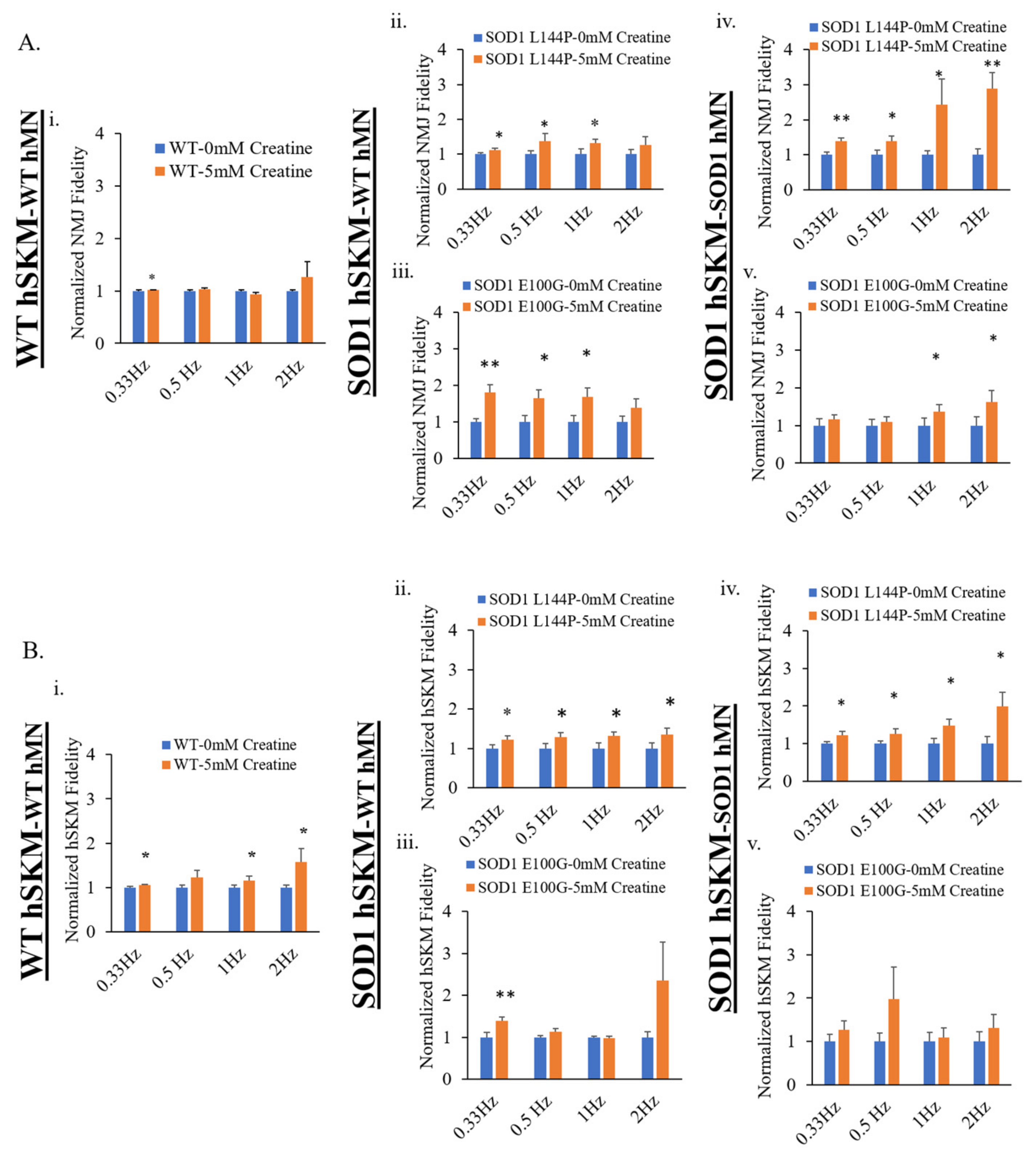
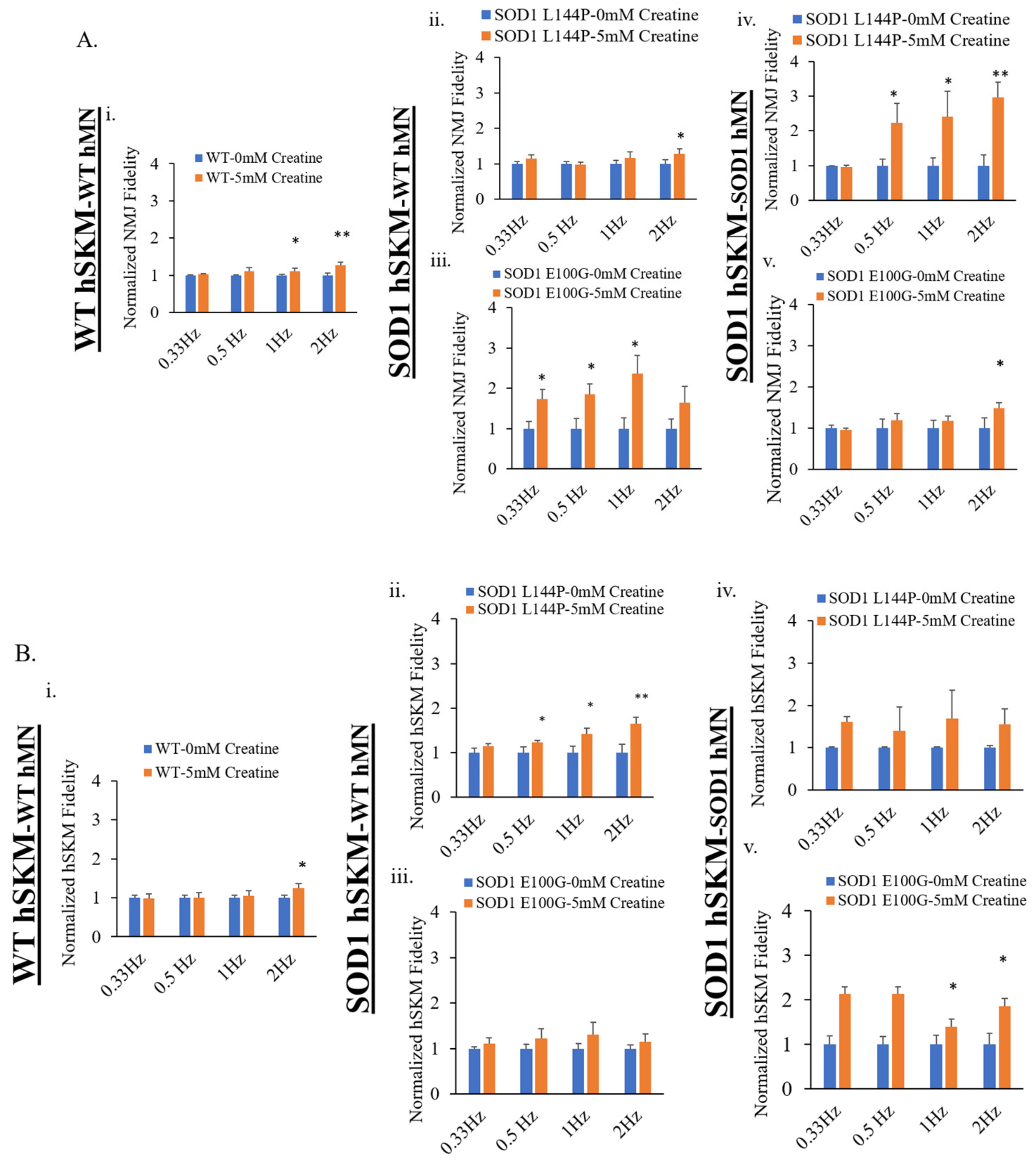
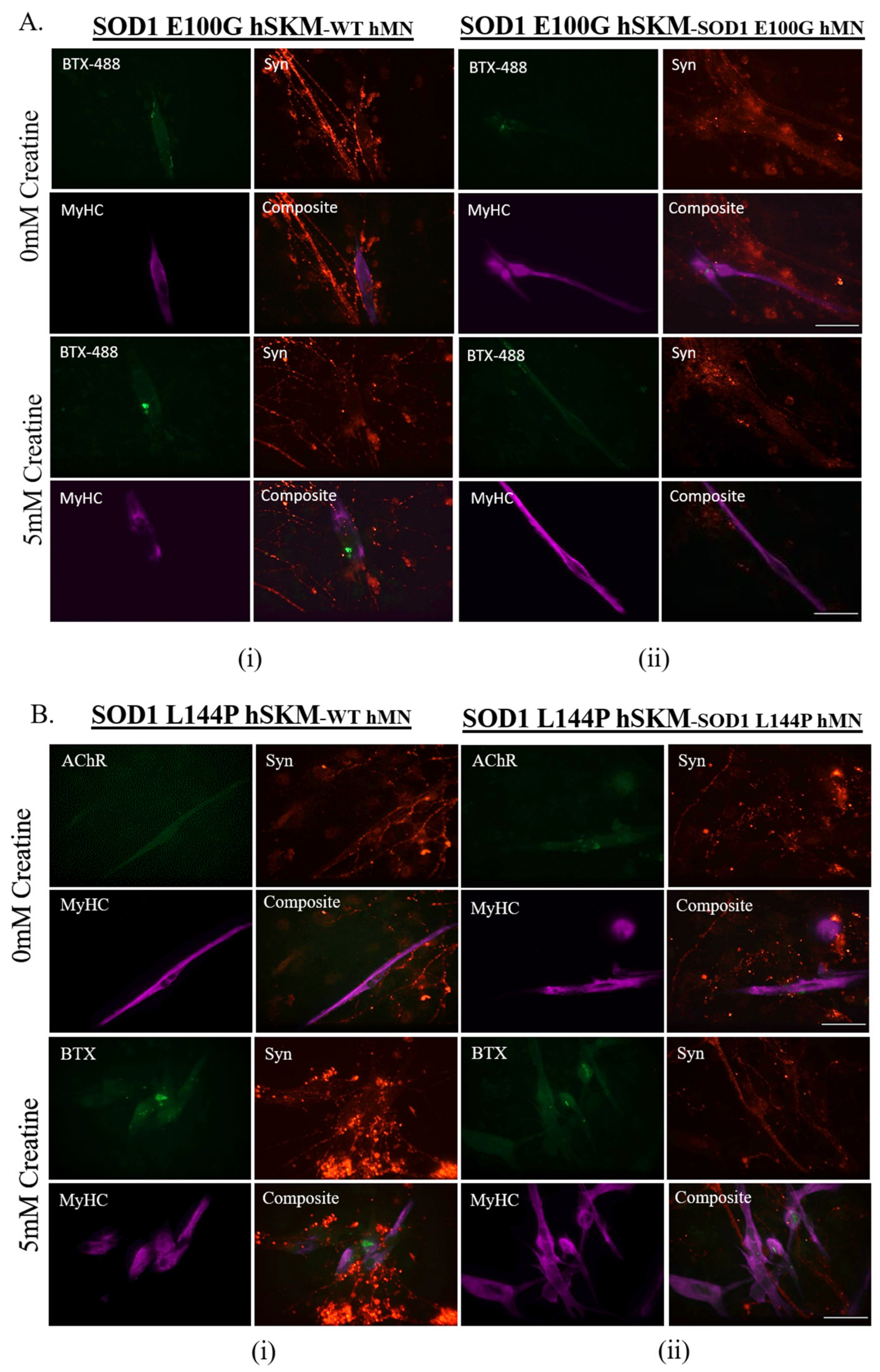
2.3. Effect of ALS hSKM-Specific Creatine Treatment on NMJ Fidelity
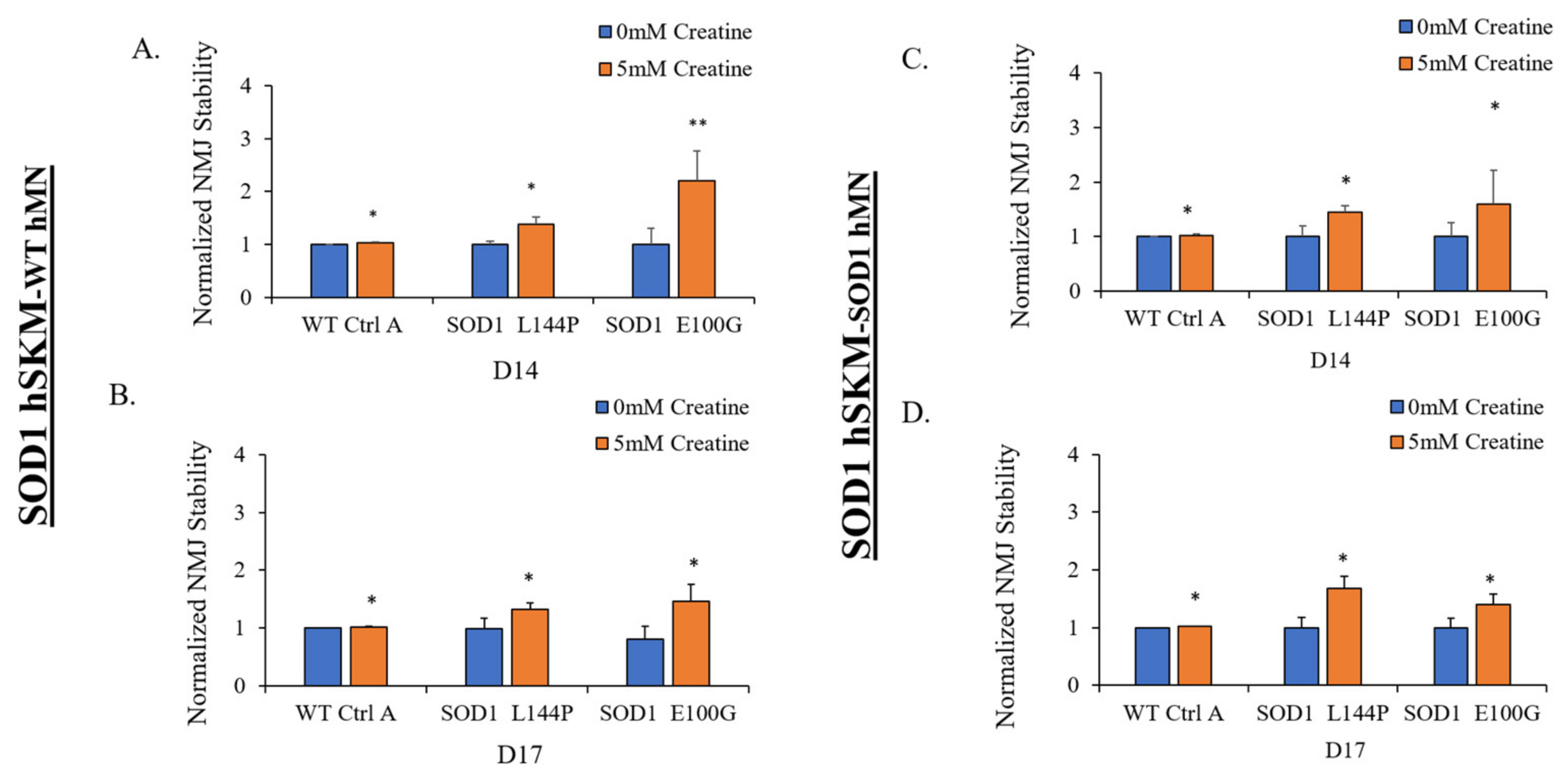
2.4. Effect of ALS hSKM-Specific Creatine Treatment on NMJ Stability
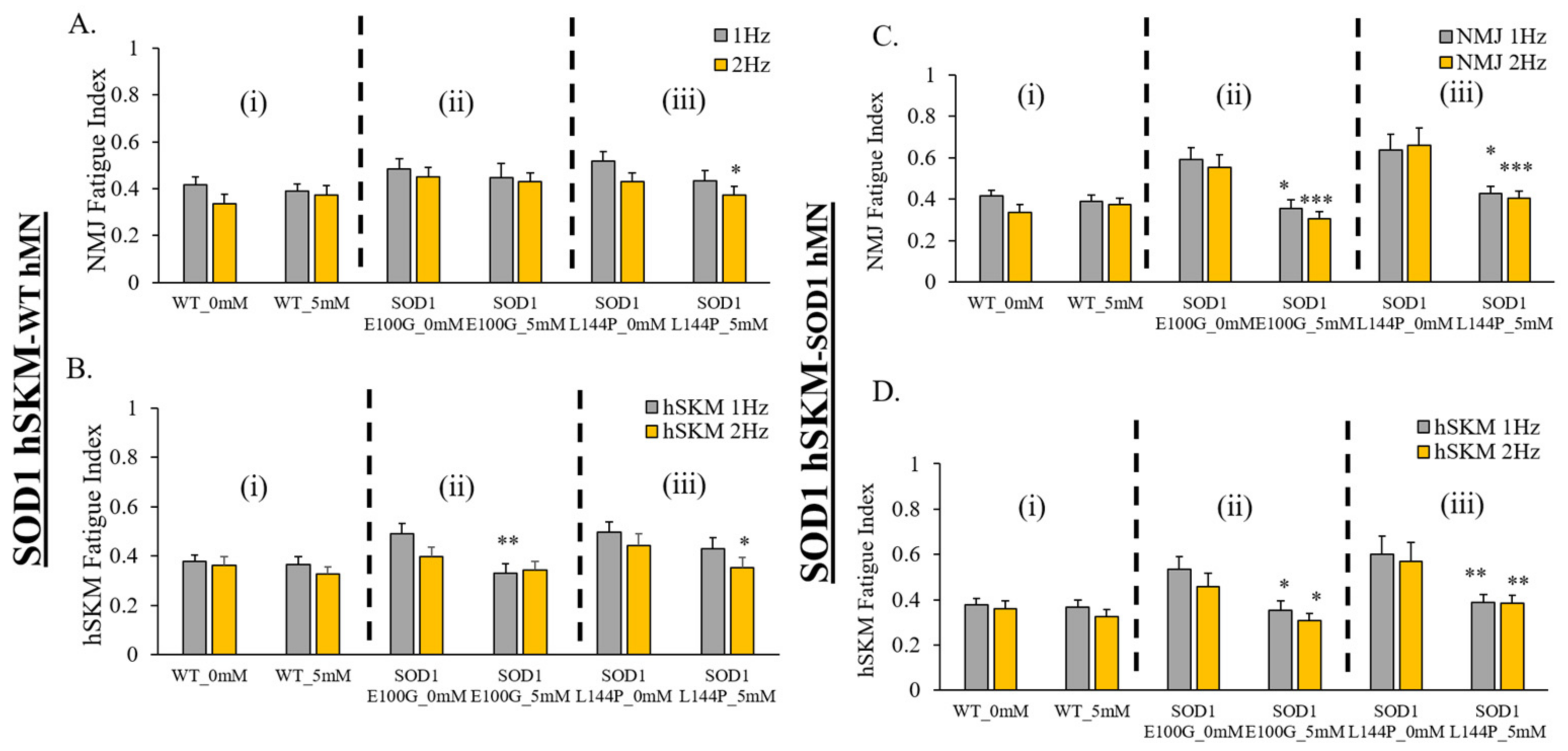
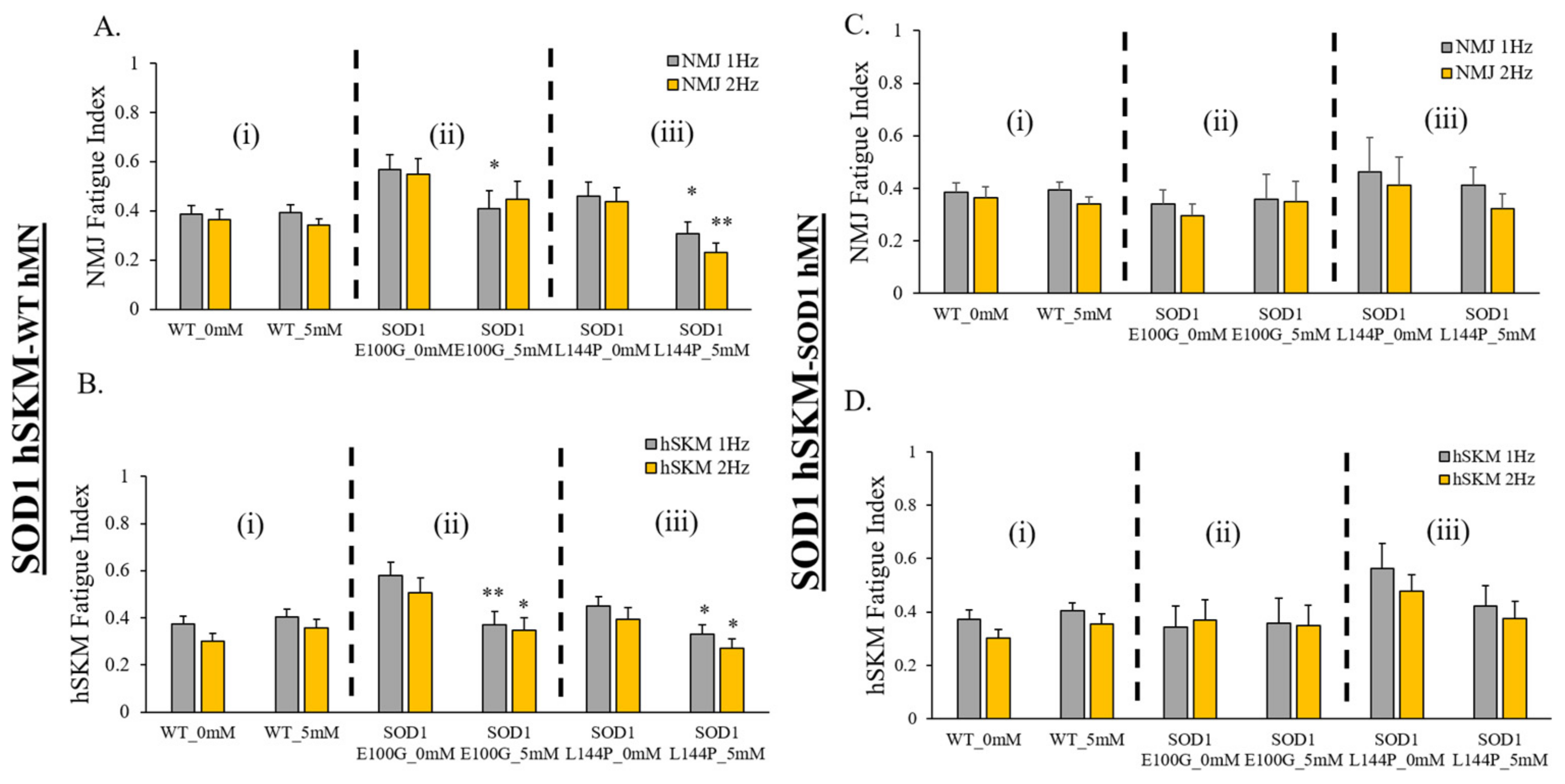
2.5. Effect of ALS hSKM-Specific Creatine Treatment on NMJ Fatigability
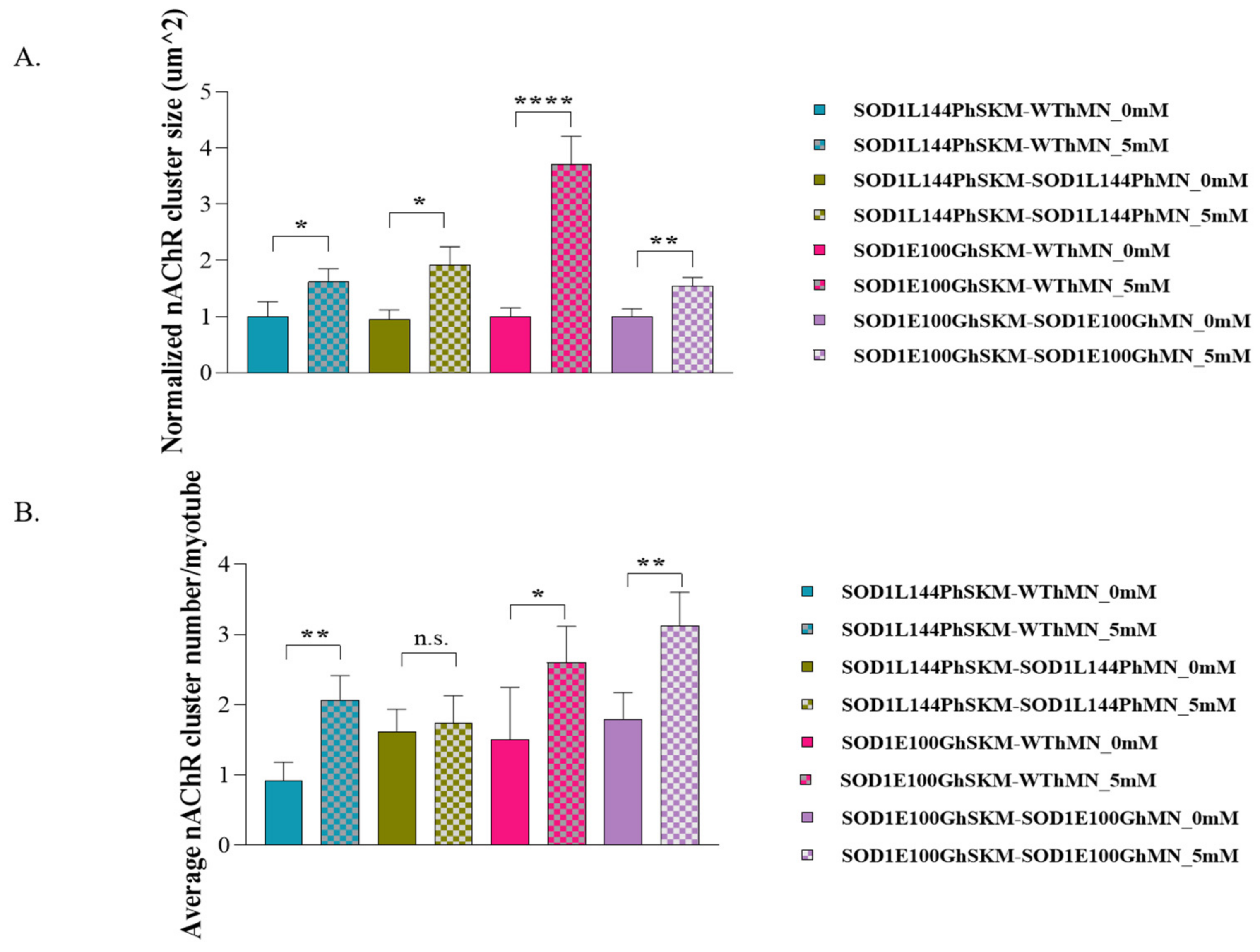
2.6. Effect of ALS hSKM-Specific Creatine Treatment on AChR Cluster Number per Myotube and Innervated AChR Cluster Size
3. Discussion
4. Materials and Methods
4.1. Tissue Derivation of IPSC-Derived Cell Source
4.2. BioMEMs NMJ Chamber Fabrication and Assembly
4.3. Cell Plating and Maintenance
4.4. ALS Skeletal Muscle Drug and Supplement Treatment
4.5. NMJ Testing and Quantification
4.6. Immunocytochemistry
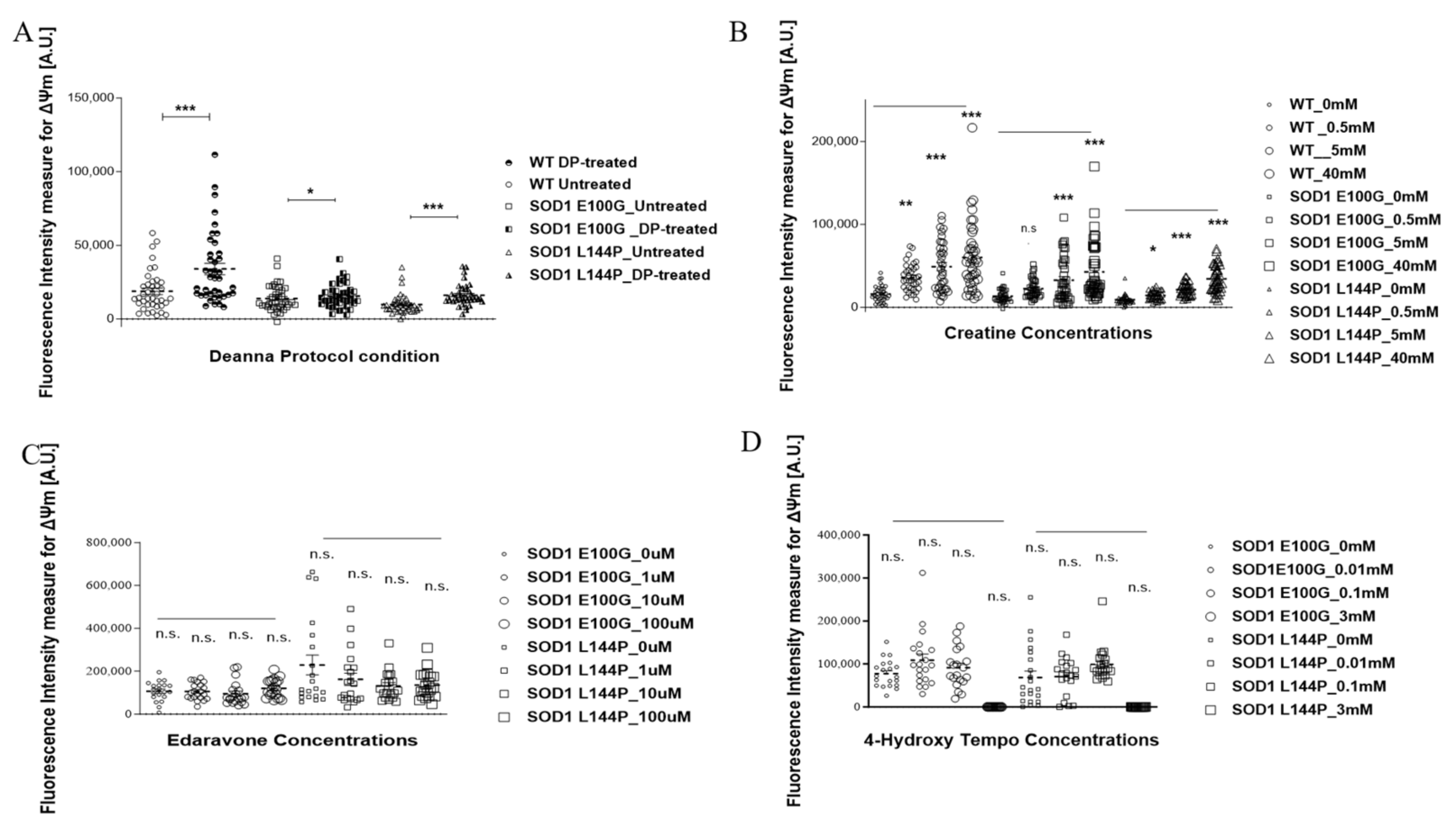
4.7. Mitochondrial Function Assessment
4.8. Acetylcholine Receptor Number and Size Quantification
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tsitkanou, S.; Della Gatta, P.A.; Russell, A.P. Skeletal muscle satellite cells, mitochondria, and micrornas: Their involvement in the pathogenesis of als. Front. Physiol. 2016, 7, 403. [Google Scholar] [CrossRef] [PubMed]
- Philips, T.; Rothstein, J.D. Glial cells in amyotrophic lateral sclerosis. Exp. Neurol. 2014, 262, 111–120. [Google Scholar] [CrossRef]
- Mejzini, R.; Flynn, L.L.; Pitout, I.L.; Fletcher, S.; Wilton, S.D.; Akkari, P.A. Als genetics, mechanisms, and therapeutics: Where are we now? Front. Neurosci. 2019, 13, 1310. [Google Scholar] [CrossRef] [PubMed]
- Pikatza-Menoio, O.; Elicegui, A.; Bengoetxea, X.; Naldaiz-Gastesi, N.; López de Munain, A.; Gerenu, G.; Gil-Bea, F.J.; Alonso-Martín, S. The skeletal muscle emerges as a new disease target in amyotrophic lateral sclerosis. J. Pers. Med. 2021, 11, 671. [Google Scholar] [CrossRef]
- Petrov, D.; Mansfield, C.; Moussy, A.; Hermine, O. Als clinical trials review: 20 years of failure. Are we any closer to registering a new treatment? Front. Aging Neurosci. 2017, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Cappello, V.; Francolini, M. Neuromuscular junction dismantling in amyotrophic lateral sclerosis. Int. J. Mol. Sci. 2017, 18, 2092. [Google Scholar] [CrossRef]
- Ikenaka, K.; Katsuno, M.; Kawai, K.; Ishigaki, S.; Tanaka, F.; Sobue, G. Disruption of axonal transport in motor neuron diseases. Int. J. Mol. Sci. 2012, 13, 1225–1238. [Google Scholar] [CrossRef]
- Jiang, Y.M.; Yamamoto, M.; Kobayashi, Y.; Yoshihara, T.; Liang, Y.; Terao, S.; Takeuchi, H.; Ishigaki, S.; Katsuno, M.; Adachi, H.; et al. Gene expression profile of spinal motor neurons in sporadic amyotrophic lateral sclerosis. Ann. Neurol. 2005, 57, 236–251. [Google Scholar] [CrossRef]
- Badu-Mensah, A.; Guo, X.; McAleer, C.; Rumsey, J.; Hickman, J.J. Skeletal muscle model derived from familial als patient ipscs recapitulates hallmarks of disease progression. Sci. Rep. 2020, 10, 14302. [Google Scholar] [CrossRef]
- Picchiarelli, G.; Demestre, M.; Zuko, A.; Been, M.; Higelin, J.; Dieterlé, S.; Goy, M.A.; Mallik, M.; Sellier, C.; Scekic-Zahirovic, J.; et al. Fus-mediated regulation of acetylcholine receptor transcription at neuromuscular junctions is compromised in amyotrophic lateral sclerosis. Nat. Neurosci. 2019, 22, 1793–1805. [Google Scholar] [CrossRef]
- Badu-Mensah, A.; Guo, X.; Nimbalkar, S.; Cai, Y.; Hickman, J.J. Als mutations in both human skeletal muscle and motoneurons differentially affects neuromuscular junction integrity and function. Biomaterials 2022, 289, 121752. [Google Scholar] [CrossRef]
- Guo, X.; Smith, V.; Jackson, M.; Tran, M.; Thomas, M.; Patel, A.; Lorusso, E.; Nimbalkar, S.; Cai, Y.; McAleer, C.W.; et al. A human-based functional nmj system for personalized als modeling and drug testing. Adv. Ther. 2020, 3, 2000133. [Google Scholar] [CrossRef]
- Bastow, E.L.; Peswani, A.R.; Tarrant, D.S.; Pentland, D.R.; Chen, X.; Morgan, A.; Staniforth, G.L.; Tullet, J.M.; Rowe, M.L.; Howard, M.J.; et al. New links between sod1 and metabolic dysfunction from a yeast model of amyotrophic lateral sclerosis. J. Cell Sci. 2016, 129, 4118–4129. [Google Scholar] [CrossRef]
- Cozzolino, M.; Carri, M.T. Mitochondrial dysfunction in als. Prog. Neurobiol. 2012, 97, 54–66. [Google Scholar] [CrossRef]
- Luo, G.; Yi, J.; Ma, C.; Xiao, Y.; Yi, F.; Yu, T.; Zhou, J. Defective mitochondrial dynamics is an early event in skeletal muscle of an amyotrophic lateral sclerosis mouse model. PLoS ONE 2013, 8, e82112. [Google Scholar] [CrossRef]
- Wiedemann, F.R.; Winkler, K.; Kuznetsov, A.V.; Bartels, C.; Vielhaber, S.; Feistner, H.; Kunz, W.S. Impairment of mitochondrial function in skeletal muscle of patients with amyotrophic lateral sclerosis. J. Neurol. Sci. 1998, 156, 65–72. [Google Scholar] [CrossRef]
- Miller, R.G.; Mitchell, J.D.; Lyon, M.; Moore, D.H. Riluzole for amyotrophic lateral sclerosis (als)/motor neuron disease (mnd). Amyotroph. Lateral Scler. Other Mot. Neuron Disord. 2003, 4, 191–206. [Google Scholar]
- Ohta, Y.; Nomura, E.; Shang, J.; Feng, T.; Huang, Y.; Liu, X.; Shi, X.; Nakano, Y.; Hishikawa, N.; Sato, K.; et al. Enhanced oxidative stress and the treatment by edaravone in mice model of amyotrophic lateral sclerosis. J. Neurosci. Res. 2019, 97, 607–619. [Google Scholar] [CrossRef]
- Samaiya, P.K.; Narayan, G.; Kumar, A.; Krishnamurthy, S. Tempol (4 hydroxy-tempo) inhibits anoxia-induced progression of mitochondrial dysfunction and associated neurobehavioral impairment in neonatal rats. J. Neurol. Sci. 2017, 375, 58–67. [Google Scholar] [CrossRef]
- Lavado, A.; Guo, X.; Smith, A.S.T.; Akanda, N.; Martin, C.; Cai, Y.; Elbrecht, D.; Tran, M.; Bryant, J.-P.; Colon, A.; et al. Evaluation of holistic treatment for als reveals possible mechanism and therapeutic potential. Int. J. Pharm. Pharm. Res. 2017, 11, 348–374. [Google Scholar]
- Farshidfar, F.; Pinder, M.A.; Myrie, S.B. Creatine supplementation and skeletal muscle metabolism for building muscle mass- review of the potential mechanisms of action. Curr. Protein Pept. Sci. 2017, 18, 1273–1287. [Google Scholar] [CrossRef]
- Wu, S.-H.; Chen, K.-L.; Hsu, C.; Chen, H.-C.; Chen, J.-Y.; Yu, S.-Y.; Shiu, Y.-J. Creatine supplementation for muscle growth: A scoping review of randomized clinical trials from 2012 to 2021. Nutrients 2022, 14, 1255. [Google Scholar] [CrossRef]
- Moloney, E.B.; de Winter, F.; Verhaagen, J. Als as a distal axonopathy: Molecular mechanisms affecting neuromuscular junction stability in the presymptomatic stages of the disease. Front. Neurosci. 2014, 8, 252. [Google Scholar] [CrossRef] [PubMed]
- Sanjak, M.; Konopacki, R.; Capasso, R.; Roelke, K.A.; Peper, S.M.; Houdek, A.M.; Waclawik, A.; Brooks, B.R. Dissociation between mechanical and myoelectrical manifestation of muscle fatigue in amyotrophic lateral sclerosis. Amyotroph. Lateral Scler. Other Mot. Neuron Disord. 2004, 5, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Lou, J.S.; Reeves, A.; Benice, T.; Sexton, G. Fatigue and depression are associated with poor quality of life in als. Neurology 2003, 60, 122–123. [Google Scholar] [CrossRef]
- McElhiney, M.; Rabkin, J.; Gordon, P.; Goetz, R.; Mitsumoto, H. Neurosurgery, psychiatry. Prevalence of fatigue and depression in als patients and change over time. J. Neurol. Neurosurg. Psychiatry 2009, 80, 1146–1149. [Google Scholar] [CrossRef]
- Argilés, J.M.; Campos, N.; Lopez-Pedrosa, J.M.; Rueda, R.; Rodriguez-Mañas, L. Skeletal muscle regulates metabolism via interorgan crosstalk: Roles in health and disease. J. Am. Med. Dir. Assoc. 2016, 17, 789–796. [Google Scholar] [CrossRef]
- Peter, R.S.; Rosenbohm, A.; Dupuis, L.; Brehme, T.; Kassubek, J.; Rothenbacher, D.; Nagel, G.; Ludolph, A.C. Life course body mass index and risk and prognosis of amyotrophic lateral sclerosis: Results from the als registry swabia. Eur. J. Epidemiol. 2017, 32, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Dobrowolny, G.; Aucello, M.; Rizzuto, E.; Beccafico, S.; Mammucari, C.; Boncompagni, S.; Belia, S.; Wannenes, F.; Nicoletti, C.; Del Prete, Z.; et al. Skeletal muscle is a primary target of sod1g93a-mediated toxicity. Cell Metab. 2008, 8, 425–436. [Google Scholar] [CrossRef]
- Quessada, C.; Bouscary, A.; René, F.; Valle, C.; Ferri, A.; Ngo, S.T.; Loeffler, J.P. Skeletal muscle metabolism: Origin or prognostic factor for amyotrophic lateral sclerosis (als) development? Cells 2021, 10, 1449. [Google Scholar] [CrossRef]
- Scaricamazza, S.; Salvatori, I.; Giacovazzo, G.; Loeffler, J.P.; Renè, F.; Rosina, M.; Quessada, C.; Proietti, D.; Heil, C.; Rossi, S.; et al. Skeletal-muscle metabolic reprogramming in als-sod1g93a mice predates disease onset and is a promising therapeutic target. iScience 2020, 23, 101087. [Google Scholar] [CrossRef] [PubMed]
- Wong, M.; Martin, L.J. Skeletal muscle-restricted expression of human sod1 causes motor neuron degeneration in transgenic mice. Hum. Mol. Genet. 2010, 19, 2284–2302. [Google Scholar] [CrossRef]
- Deldicque, L.; Theisen, D.; Bertrand, L.; Hespel, P.; Hue, L.; Francaux, M. Creatine enhances differentiation of myogenic C2C12 cells by activating both p38 and akt/pkb pathways. Am. J. Physiol. Cell Physiol. 2007, 293, C1263–C1271. [Google Scholar] [CrossRef]
- McAleer, C.W.; Smith, A.S.T.; Najjar, S.; Pirozzi, K.; Long, C.J.; Hickman, J.J. Mechanistic investigation of adult myotube response to exercise and drug treatment in vitro using a multiplexed functional assay system. J. Appl. Physiol. 2014, 117, 1398–1405. [Google Scholar] [CrossRef] [PubMed]
- Pulido, S.M.; Passaquin, A.C.; Leijendekker, W.J.; Challet, C.; Wallimann, T.; Rüegg, U.T. Creatine supplementation improves intracellular Ca2+ handling and survival in mdx skeletal muscle cells. FEBS Lett. 1998, 439, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Young, J.F.; Larsen, L.B.; Malmendal, A.; Nielsen, N.C.; Straadt, I.K.; Oksbjerg, N.; Bertram, H.C. Creatine-induced activation of antioxidative defence in myotube cultures revealed by explorative nmr-based metabonomics and proteomics. J. Int. Soc. Sports Nutr. 2010, 7, 9. [Google Scholar] [CrossRef]
- Deldicque, L.; Atherton, P.; Patel, R.; Theisen, D.; Nielens, H.; Rennie, M.J.; Francaux, M.; Samuelsson, H.; Moberg, M.; Apró, W.; et al. Effects of resistance exercise with and without creatine supplementation on gene expression and cell signaling in human skeletal muscle. J. Appl. Physiol. 2008, 104, 371–378. [Google Scholar] [CrossRef]
- Contreras, L.; Drago, I.; Zampese, E.; Pozzan, T. Mitochondria: The calcium connection. Biochim. Biophys. Acta 2010, 1797, 607–618. [Google Scholar] [CrossRef]
- O’Connor, R.S.; Steeds, C.M.; Wiseman, R.W.; Pavlath, G.K. Phosphocreatine as an energy source for actin cytoskeletal rearrangements during myoblast fusion. J. Physiol. 2008, 586, 2841–2853. [Google Scholar] [CrossRef]
- Busanello, E.N.B.; Marques, A.C.; Lander, N.; de Oliveira, D.N.; Catharino, R.R.; Oliveira, H.C.F.; Vercesi, A.E. Pravastatin chronic treatment sensitizes hypercholesterolemic mice muscle to mitochondrial permeability transition: Protection by creatine or coenzyme q10. Front. Pharmacol. 2017, 8, 185. [Google Scholar] [CrossRef]
- Wallimann, T.; Dolder, M.; Schlattner, U.; Eder, M.; Hornemann, T.; O’Gorman, E.; Rück, A.; Brdiczka, D. Some new aspects of creatine kinase (CK): Compartmentation, structure, function and regulation for cellular and mitochondrial bioenergetics and physiology. Biofactors 1998, 8, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Tokarska-Schlattner, M.; Epand, R.F.; Meiler, F.; Zandomeneghi, G.; Neumann, D.; Widmer, H.R.; Meier, B.H.; Epand, R.M.; Saks, V.; Wallimann, T.; et al. Phosphocreatine interacts with phospholipids, affects membrane properties and exerts membrane-protective effects. PLoS ONE 2012, 7, e43178. [Google Scholar] [CrossRef]
- Handschin, C.; Kobayashi, Y.M.; Chin, S.; Seale, P.; Campbell, K.P.; Spiegelman, B.M. Pgc-1α regulates the neuromuscular junction program and ameliorates duchenne muscular dystrophy. Genes Dev. 2007, 21, 770–783. [Google Scholar] [CrossRef]
- Palomer, X.; Alvarez-Guardia, D.; Rodríguez-Calvo, R.; Coll, T.; Laguna, J.C.; Davidson, M.M.; Chan, T.O.; Feldman, A.M.; Vázquez-Carrera, M. Tnf-α reduces pgc-1 α expression through nf-κb and p38 mapk leading to increased glucose oxidation in a human cardiac cell model. Cardiovasc. Res. 2009, 81, 703–712. [Google Scholar] [CrossRef]
- Safdar, A.; Yardley, N.J.; Snow, R.; Melov, S.; Tarnopolsky, M.A. Global and targeted gene expression and protein content in skeletal muscle of young men following short-term creatine monohydrate supplementation. Physiol. Genom. 2008, 32, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, N.; Kumanchik, L.; Guo, X.; Sommerhage, F.; Cai, Y.; Jackson, M.; Martin, C.; Saad, G.; McAleer, C.W.; Wang, Y.; et al. Stem cell derived phenotypic human neuromuscular junction model for dose response evaluation of therapeutics. Biomaterials 2018, 166, 64–78. [Google Scholar] [CrossRef]
- Guo, X.; Das, M.; Rumsey, J.W.; Gonzalez, M.; Stancescu, M.; Hickman, J.J. Neuromuscular junction formation between human stem cell derived motoneurons and rat skeletal muscle in a defined system. Tissue Eng. Part. C Methods 2010, 16, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Hammond, L. Measuring Cell Fluorescence Using Imagej. 2014. Available online: https://theolb.readthedocs.io/en/latest/imaging/measuring-cell-fluorescence-using-imagej.html (accessed on 1 June 2023).











Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Badu-Mensah, A.; Guo, X.; Mendez, R.; Parsaud, H.; Hickman, J.J. The Effect of Skeletal Muscle-Specific Creatine Treatment on ALS NMJ Integrity and Function. Int. J. Mol. Sci. 2023, 24, 13519. https://doi.org/10.3390/ijms241713519
Badu-Mensah A, Guo X, Mendez R, Parsaud H, Hickman JJ. The Effect of Skeletal Muscle-Specific Creatine Treatment on ALS NMJ Integrity and Function. International Journal of Molecular Sciences. 2023; 24(17):13519. https://doi.org/10.3390/ijms241713519
Chicago/Turabian StyleBadu-Mensah, Agnes, Xiufang Guo, Roxana Mendez, Hemant Parsaud, and James J. Hickman. 2023. "The Effect of Skeletal Muscle-Specific Creatine Treatment on ALS NMJ Integrity and Function" International Journal of Molecular Sciences 24, no. 17: 13519. https://doi.org/10.3390/ijms241713519