
EGF, TGF-α and Amphiregulin Differently Regulate Endometrium-Derived Mesenchymal Stromal/Stem Cells
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
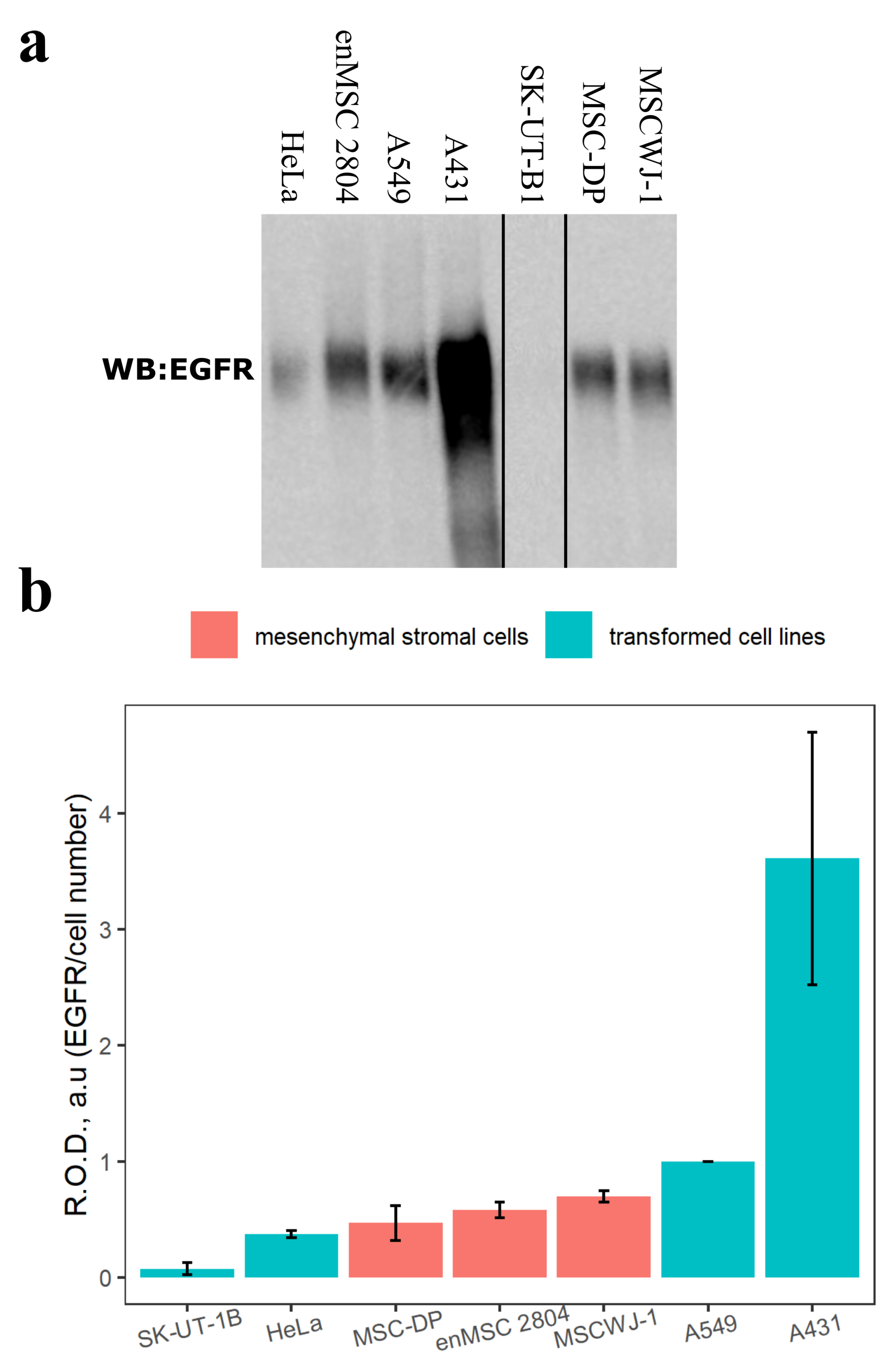
2.1. Estimation of the EGFR Protein Level in Different Mesenchymal Stromal and Transformed Cell Lines
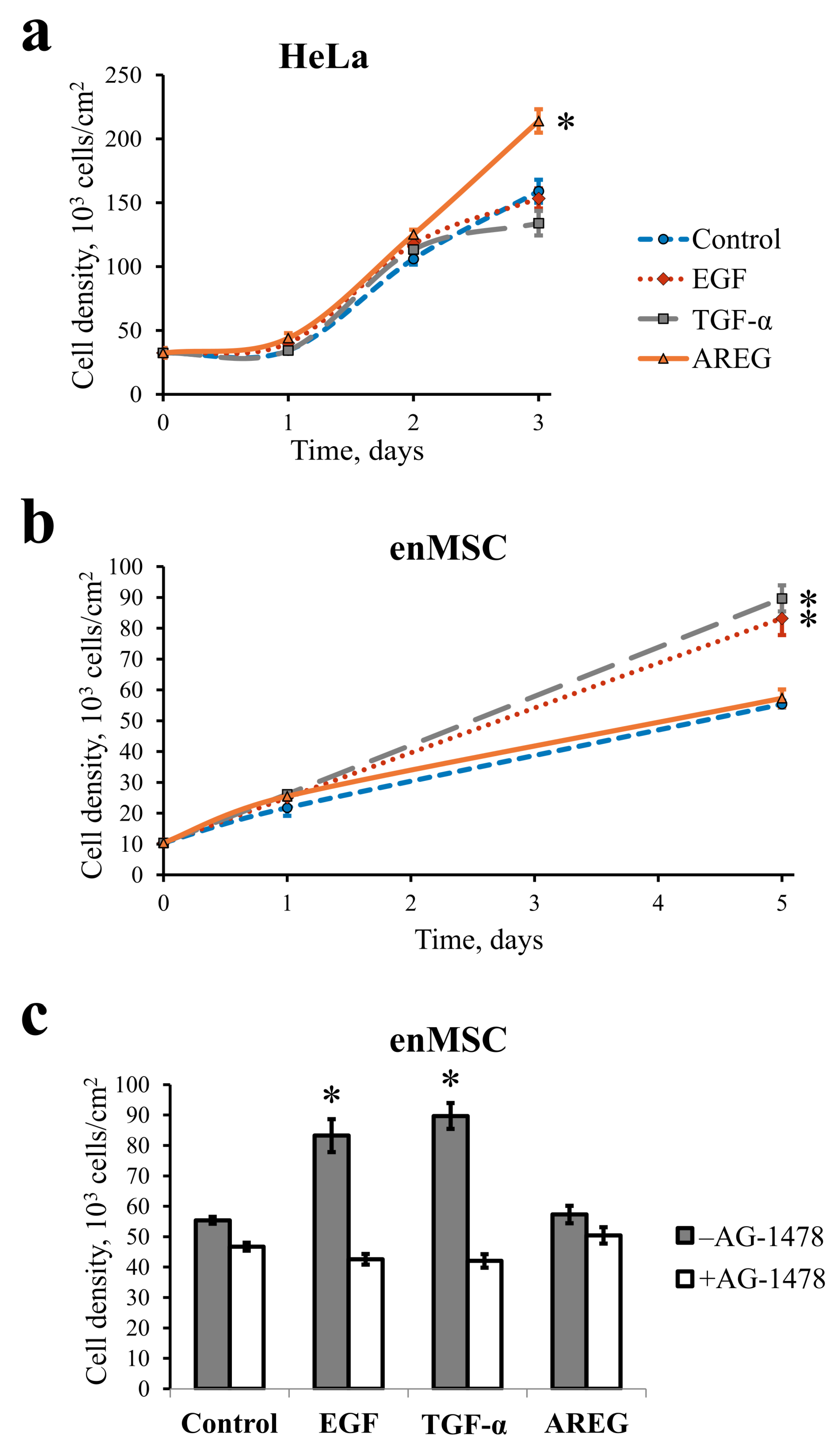
2.2. EGF, TGF- and AREG Differentially Affect HeLa and enMSC Cell Proliferation
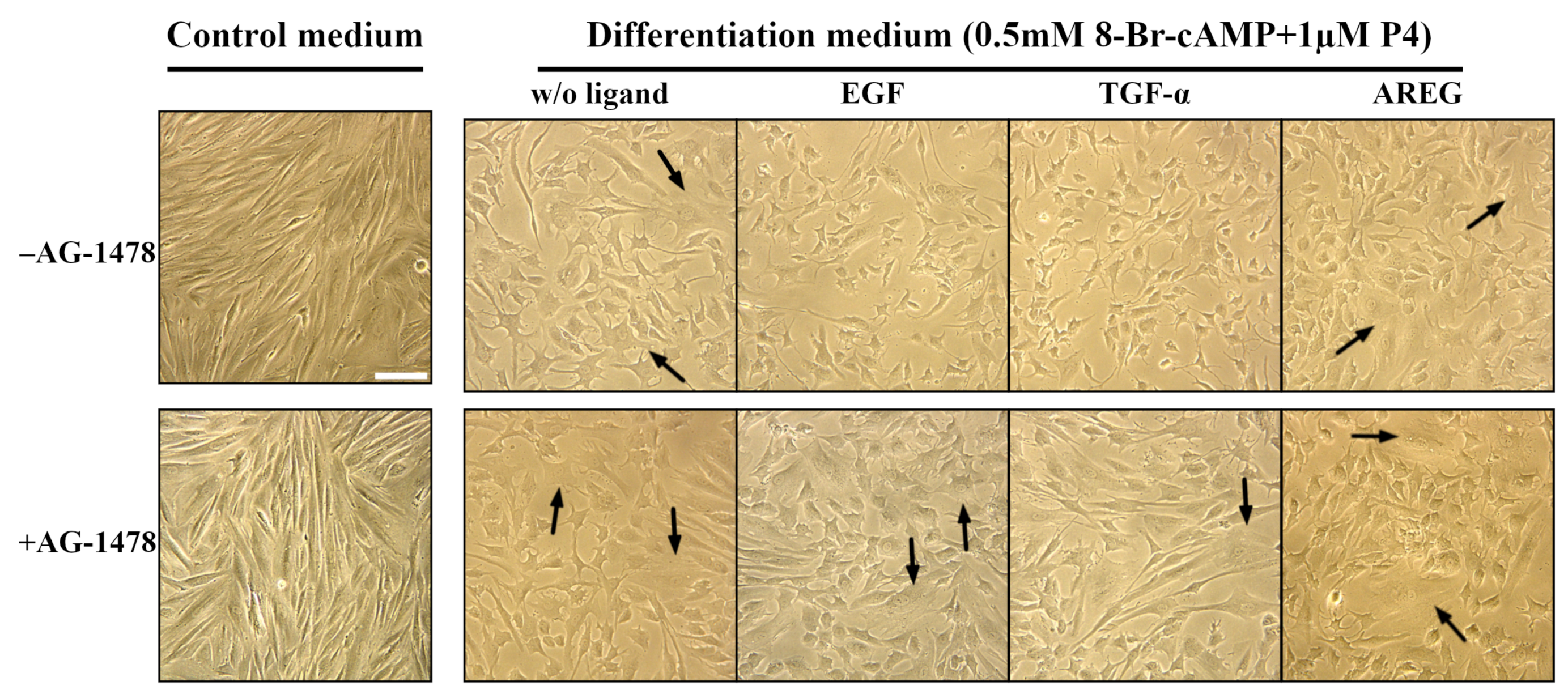
2.3. EGF and TGF- Suppress Decidualization of Human enMSCs
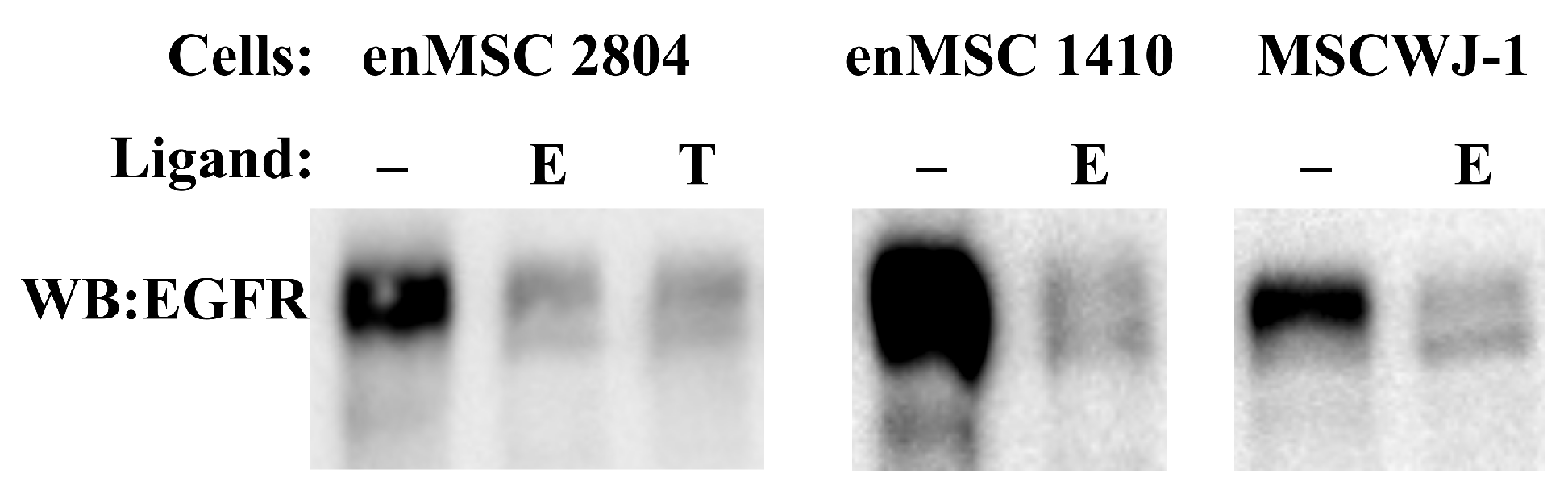
2.4. AREG Is Secreted by Human enMSCs
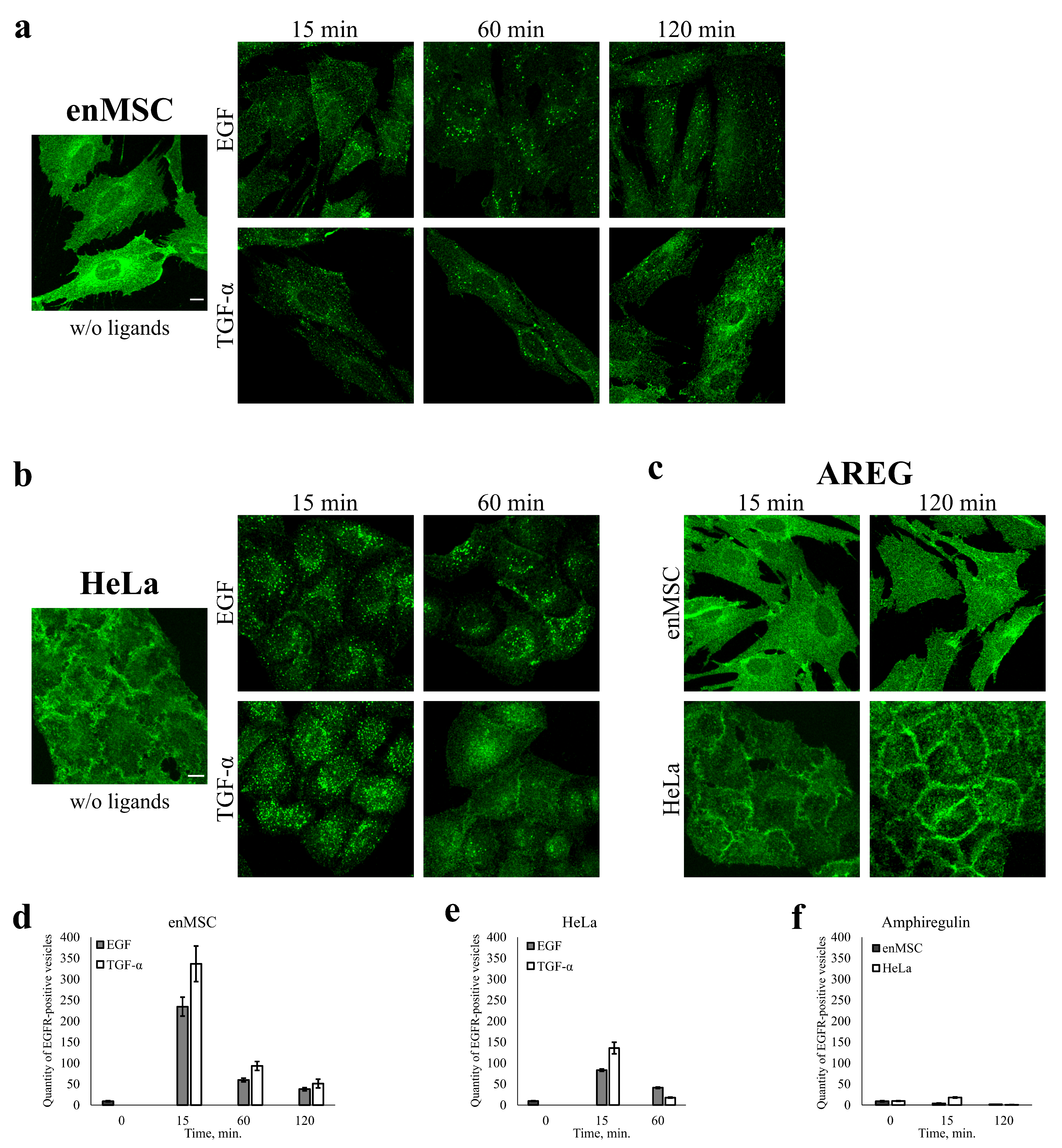
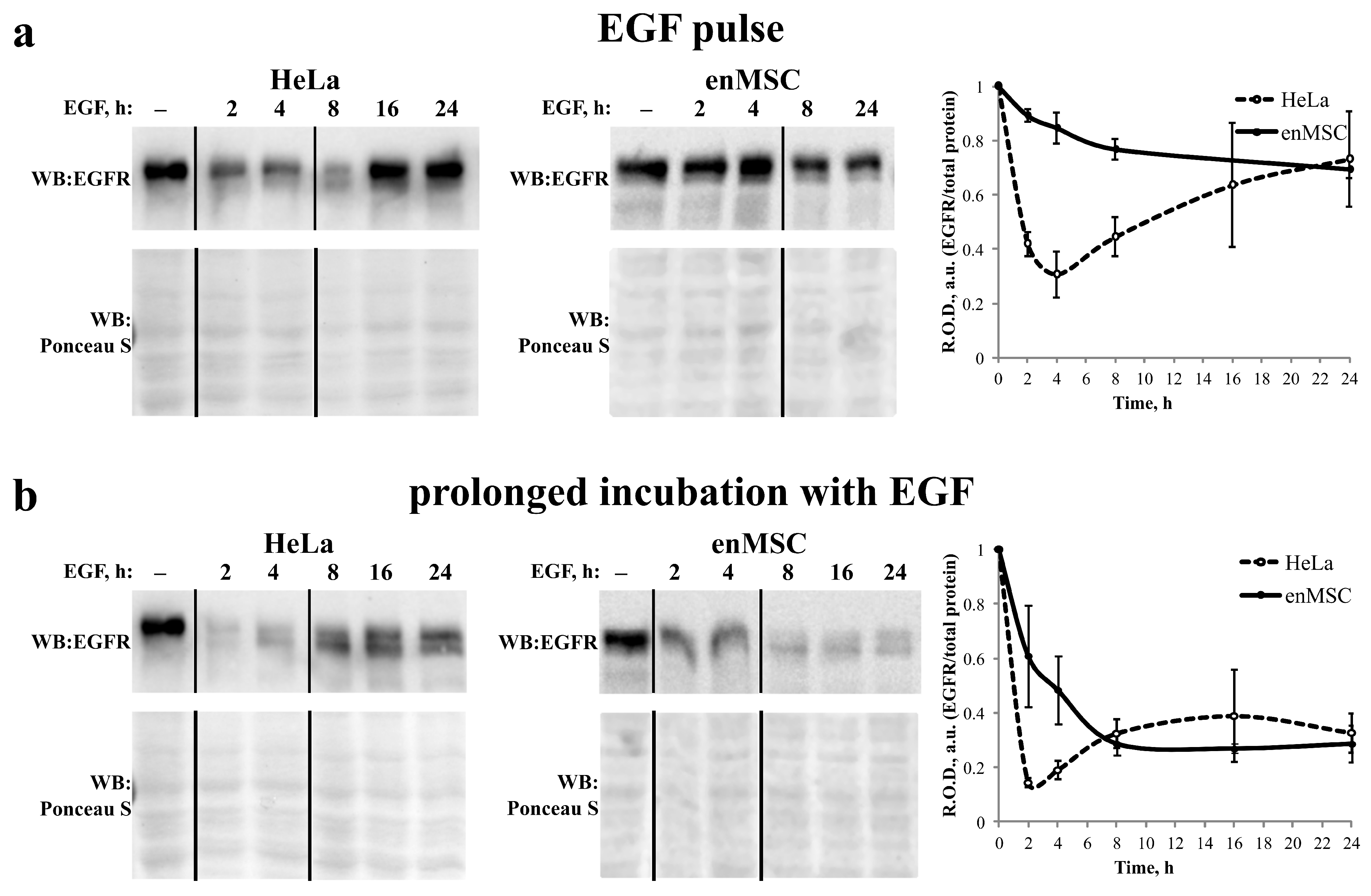
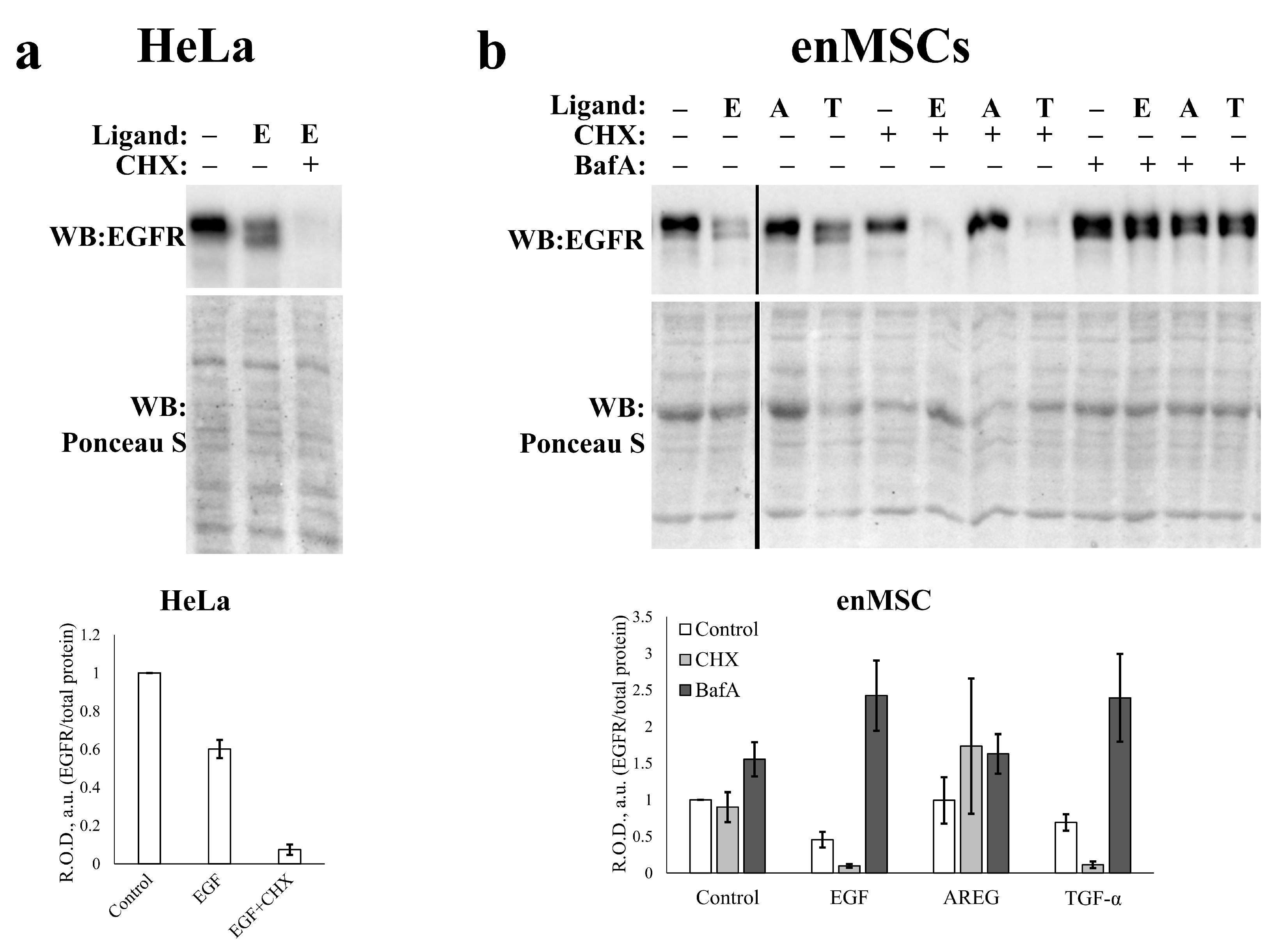
2.5. EGFR Endocytosis Is Stimulated by EGF and TGF- but Not by AREG in Both enMSCs and HeLa Cells
2.6. EnMSC Incubation with EGF Leads to a Long-Term Decrease in the Level of EGFR
3. Discussion
4. Materials and Methods
4.1. Cell Cultures
4.2. Reagents
4.3. Proliferation Assay
4.4. Decidualization Induction
4.5. Western Blotting
4.6. Immunofluorescence and Laser Scanning Confocal Microscopy
4.7. ELISA
4.8. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AREG | amphiregulin |
| BafA | bafilomycin A |
| CHX | cycloheximide |
| EGF | epidermal growth factor |
| EGFR | EGF receptor |
| enMSC | endometrium-derived MSC |
| MSC | mesenchymal stromal/stem cells |
| TGF- | transforming growth factor- |
References
- Ling, L.; Nurcombe, V.; Cool, S.M. Wnt signaling controls the fate of mesenchymal stem cells. Gene 2009, 433, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kurpinski, K.; Lam, H.; Chu, J.; Wang, A.; Kim, A.; Tsay, E.; Agrawal, S.; Schaffer, D.V.; Li, S. Transforming Growth Factor-β and Notch Signaling Mediate Stem Cell Differentiation into Smooth Muscle Cells. Stem Cells 2010, 28, 734–742. [Google Scholar] [CrossRef]
- Takam Kamga, P.; Bazzoni, R.; Dal Collo, G.; Cassaro, A.; Tanasi, I.; Russignan, A.; Tecchio, C.; Krampera, M. The Role of Notch and Wnt Signaling in MSC Communication in Normal and Leukemic Bone Marrow Niche. Front. Cell Dev. Biol. 2021, 8, 599276. [Google Scholar] [CrossRef]
- Schuldiner, M.; Yanuka, O.; Itskovitz-Eldor, J.; Melton, D.A.; Benvenisty, N. Effects of eight growth factors on the differentiation of cells derived from human embryonic stem cells. Proc. Natl. Acad. Sci. USA 2000, 97, 11307–11312. [Google Scholar] [CrossRef]
- Scalabrino, G. Epidermal Growth Factor in the CNS: A Beguiling Journey from Integrated Cell Biology to Multiple Sclerosis. An Extensive Translational Overview. Cell. Mol. Neurobiol. 2022, 42, 891–916. [Google Scholar] [CrossRef] [PubMed]
- Large, M.J.; Wetendorf, M.; Lanz, R.B.; Hartig, S.M.; Creighton, C.J.; Mancini, M.A.; Kovanci, E.; Lee, K.F.; Threadgill, D.W.; Lydon, J.P.; et al. The Epidermal Growth Factor Receptor Critically Regulates Endometrial Function during Early Pregnancy. PLoS Genet. 2014, 10, e1004451. [Google Scholar] [CrossRef]
- Liu, L.P.; Gong, Y.B. LncRNA-TCL6 promotes early abortion and inhibits placenta implantation via the EGFR pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 7105–7112. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, P.J.; Berger, J.E.; Meneses, J.; Phung, Y.; Pedersen, R.A.; Werb, Z.; Derynck, R. Epithelial immaturity and multiorgan failure in mice lacking epidermal growth factor receptor. Nature 1995, 376, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Ejskjær, K.; Sørensen, B.; Poulsen, S.; Mogensen, O.; Forman, A.; Nexø, E. Expression of the epidermal growth factor system in human endometrium during the menstrual cycle. MHR Basic Sci. Reprod. Med. 2005, 11, 543–551. [Google Scholar] [CrossRef]
- Boutin, A.; Marcus-Samuels, B.; Eliseeva, E.; Neumann, S.; Gershengorn, M.C. Opposing Effects of EGF Receptor Signaling on Proliferation and Differentiation Initiated by EGF or TSH/EGF Receptor Transactivation. Endocrinology 2022, 163, bqac136. [Google Scholar] [CrossRef]
- Bozorgmehr, M.; Gurung, S.; Darzi, S.; Nikoo, S.; Kazemnejad, S.; Zarnani, A.H.; Gargett, C.E. Endometrial and Menstrual Blood Mesenchymal Stem/Stromal Cells: Biological Properties and Clinical Application. Front. Cell Dev. Biol. 2020, 8, 497. [Google Scholar] [CrossRef]
- Saraiva, A.; Payan-Carreira, R.; Gärtner, F.; Faria, F.; Lourenço, L.; Pires, M. Changes in c-erbB-2 Immunoexpression in Feline Endometrial Adenocarcinomas. Reprod. Domest. Anim. 2016, 51, 33–39. [Google Scholar] [CrossRef]
- Irwin, J.C.; Utian, W.H.; Eckert, R.L. Sex Steroids and Growth Factors Differentially Regulate the Growth and Differentiation of Cultured Human Endometrial Stromal Cells. Endocrinology 1991, 129, 2385–2392. [Google Scholar] [CrossRef] [PubMed]
- Imai, T.; Kurachi, H.; Adachi, K.; Adachi, H.; Yoshimoto, Y.; Homma, H.; Tadokoro, C.; Takeda, S.; Yamaguchi, M.; Sakata, M.; et al. Changes in Epidermal Growth Factor Receptor and the Levels of its Ligands during Menstrual Cycle in Human Endometrium. Biol. Reprod. 1995, 52, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Aghajanova, L.; Bjuresten, K.; Altmäe, S.; Landgren, B.M.; Stavreus-Evers, A. HB-EGF but Not Amphiregulin or Their Receptors HER1 and HER4 Is Altered in Endometrium of Women with Unexplained Infertility. Reprod. Sci. 2008, 15, 484–492. [Google Scholar] [CrossRef]
- Reis, F.M.; Lhullier, C.; Edelweiss, M.I.; Spritzer, P.M. In Vivo Assessment of the Regulation of Transforming Growth Factor Alpha, Epidermal Growth Factor (EGF), and EGF Receptor in the Human Endometrium by Medroxyprogesterone Acetate. J. Assist. Reprod. Genet. 2005, 22, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Lockwood, C.J.; Krikun, G.; Schatz, F. Decidual Cell-Expressed Tissue Factor Maintains Hemostasis in Human Endometrium. Ann. N. Y. Acad. Sci. 2001, 943, 77–88. [Google Scholar] [CrossRef]
- Morishige, K.; Kurachi, H.; Amemiya, K.; Adachi, H.; Adachi, K.; Sakoyama, Y.; Miyake, A.; Tanizawa, O. Menstrual stage-specific expression of epidermal growth factor and transforming growth factor-alpha in human oviduct epithelium and their role in early embryogenesis. Endocrinology 1993, 133, 199–207. [Google Scholar] [CrossRef]
- Bulletti, C.; Jasonni, V.M.; Polli, V.; Cappuccini, F.; Galassi, A.; Flamigni, C. Basement Membrane in Human Endometrium: Possible Role of Proteolytic Enzymes in Developing Hyperplasia and Carcinoma. Ann. N. Y. Acad. Sci. 1991, 622, 376–382. [Google Scholar] [CrossRef]
- Yu, M.; Li, X.; Wu, X.; Wang, W.; Li, Y.; Zhang, Y.; Zhang, S.; Wang, Y. Continuous Vaginal Bleeding Induced By EGFR-TKI in Premenopausal Female Patients with EGFR Mutant NSCLC. Front. Oncol. 2022, 12, 805538. [Google Scholar] [CrossRef]
- Capuani, F.; Conte, A.; Argenzio, E.; Marchetti, L.; Priami, C.; Polo, S.; Di Fiore, P.P.; Sigismund, S.; Ciliberto, A. Quantitative analysis reveals how EGFR activation and downregulation are coupled in normal but not in cancer cells. Nat. Commun. 2015, 6, 7999. [Google Scholar] [CrossRef] [PubMed]
- Fabricant, R.N.; De Larco, J.E.; Todaro, G.J. Nerve growth factor receptors on human melanoma cells in culture. Proc. Natl. Acad. Sci. USA 1977, 74, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Radinsky, R.; Risin, S.; Fan, D.; Dong, Z.; Bielenberg, D.; Bucana, C.D.; Fidler, I.J. Level and function of epidermal growth factor receptor predict the metastatic potential of human colon carcinoma cells. Clin. Cancer Res. 1995, 1, 19–31. [Google Scholar]
- Sigismund, S.; Avanzato, D.; Lanzetti, L. Emerging functions of the EGFR in cancer. Mol. Oncol. 2018, 12, 3–20. [Google Scholar] [CrossRef]
- Rodriguez, S.M.B.; Kamel, A.; Ciubotaru, G.V.; Onose, G.; Sevastre, A.S.; Sfredel, V.; Danoiu, S.; Dricu, A.; Tataranu, L.G. An Overview of EGFR Mechanisms and Their Implications in Targeted Therapies for Glioblastoma. Int. J. Mol. Sci. 2023, 24, 11110. [Google Scholar] [CrossRef] [PubMed]
- Goustin, A.S.; Leof, E.B.; Shipley, G.D.; Moses, H.L. Growth factors and cancer. Cancer Res. 1986, 46, 1015–1029. [Google Scholar]
- Lindsey, S.; Langhans, S.A. Epidermal Growth Factor Signaling in Transformed Cells. In International Review of Cell and Molecular Biology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 314, pp. 1–41. [Google Scholar] [CrossRef]
- Landry, J.J.M.; Pyl, P.T.; Rausch, T.; Zichner, T.; Tekkedil, M.M.; Stütz, A.M.; Jauch, A.; Aiyar, R.S.; Pau, G.; Delhomme, N.; et al. The genomic and transcriptomic landscape of a HeLa cell line. G3 2013, 3, 1213–1224. [Google Scholar] [CrossRef]
- Chung, I.; Akita, R.; Vandlen, R.; Toomre, D.; Schlessinger, J.; Mellman, I. Spatial control of EGF receptor activation by reversible dimerization on living cells. Nature 2010, 464, 783–787. [Google Scholar] [CrossRef]
- Needham, S.R.; Roberts, S.K.; Arkhipov, A.; Mysore, V.P.; Tynan, C.J.; Zanetti-Domingues, L.C.; Kim, E.T.; Losasso, V.; Korovesis, D.; Hirsch, M.; et al. EGFR oligomerization organizes kinase-active dimers into competent signalling platforms. Nat. Commun. 2016, 7, 13307. [Google Scholar] [CrossRef]
- Sawano, A.; Takayama, S.; Matsuda, M.; Miyawaki, A. Lateral Propagation of EGF Signaling after Local Stimulation Is Dependent on Receptor Density. Dev. Cell 2002, 3, 245–257. [Google Scholar] [CrossRef]
- Wilson, K.J.; Gilmore, J.L.; Foley, J.; Lemmon, M.A.; Riese, D.J. Functional selectivity of EGF family peptide growth factors: Implications for cancer. Pharmacol. Ther. 2009, 122, 1–8. [Google Scholar] [CrossRef]
- Rusch, V.; Klimstra, D.; Venkatraman, E.; Pisters, P.W.; Langenfeld, J.; Dmitrovsky, E. Overexpression of the epidermal growth factor receptor and its ligand transforming growth factor alpha is frequent in resectable non-small cell lung cancer but does not predict tumor progression. Clin. Cancer Res. 1997, 3, 515–522. [Google Scholar]
- Yarden, Y.; Pines, G. The ERBB network: At last, cancer therapy meets systems biology. Nat. Rev. Cancer 2012, 12, 553–563. [Google Scholar] [CrossRef] [PubMed]
- Roskoski, R. The ErbB/HER family of protein-tyrosine kinases and cancer. Pharmacol. Res. 2014, 79, 34–74. [Google Scholar] [CrossRef]
- Singh, A.B.; Harris, R.C. Autocrine, paracrine and juxtacrine signaling by EGFR ligands. Cell. Signal. 2005, 17, 1183–1193. [Google Scholar] [CrossRef] [PubMed]
- Isokane, M.; Hieda, M.; Hirakawa, S.; Shudou, M.; Nakashiro, K.; Hashimoto, K.; Hamakawa, H.; Higashiyama, S. Plasma-membrane-anchored growth factor pro-amphiregulin binds A-type lamin and regulates global transcription. J. Cell Sci. 2008, 121, 3608–3618. [Google Scholar] [CrossRef]
- Sanders, J.M.; Wampole, M.E.; Thakur, M.L.; Wickstrom, E. Molecular Determinants of Epidermal Growth Factor Binding: A Molecular Dynamics Study. PLoS ONE 2013, 8, e54136. [Google Scholar] [CrossRef]
- Kong, C.; Su, X.; Chen, P.I.; Stahl, P.D. Rin1 Interacts with Signal-transducing Adaptor Molecule (STAM) and Mediates Epidermal Growth Factor Receptor Trafficking and Degradation. J. Biol. Chem. 2007, 282, 15294–15301. [Google Scholar] [CrossRef]
- De Melker, A.A.; Van Der Horst, G.; Calafat, J.; Jansen, H.; Borst, J. c-Cbl ubiquitinates the EGF receptor at the plasma membrane and remains receptor associated throughout the endocytic route. J. Cell Sci. 2001, 114, 2167–2178. [Google Scholar] [CrossRef] [PubMed]
- Ebner, R.; Derynck, R. Epidermal growth factor and transforming growth factor-α: Differential intracellular routing and processing of ligand-receptor complexes. Cell Regul. 1991, 2, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Roepstorff, K.; Grandal, M.V.; Henriksen, L.; Knudsen, S.L.J.; Lerdrup, M.; Grøvdal, L.; Willumsen, B.M.; van Deurs, B. Differential Effects of EGFR Ligands on Endocytic Sorting of the Receptor. Traffic 2009, 10, 1115–1127. [Google Scholar] [CrossRef]
- Zhang, F.; Wang, S.; Yin, L.; Yang, Y.; Guan, Y.; Wang, W.; Xu, H.; Tao, N. Quantification of Epidermal Growth Factor Receptor Expression Level and Binding Kinetics on Cell Surfaces by Surface Plasmon Resonance Imaging. Anal. Chem. 2015, 87, 9960–9965. [Google Scholar] [CrossRef]
- Chen, T.R. SK-UT-1B, a human tumorigenic diploid cell line. Cancer Genet. Cytogenet. 1988, 33, 77–81. [Google Scholar] [CrossRef]
- Favaro, R.; Abrahamsohn, P.A.; Zorn, M.T. Decidualization and Endometrial Extracellular Matrix Remodeling. In The Guide to Investigation of Mouse Pregnancy; Elsevier: Amsterdam, The Netherlands, 2014; pp. 125–142. [Google Scholar] [CrossRef]
- Gellersen, B.; Brosens, J.J. Cyclic Decidualization of the Human Endometrium in Reproductive Health and Failure. Endocr. Rev. 2014, 35, 851–905. [Google Scholar] [CrossRef] [PubMed]
- Gorsek Sparovec, T.; Markert, U.R.; Reif, P.; Schoell, W.; Moser, G.; Feichtinger, J.; Mihalic, Z.N.; Kargl, J.; Gargett, C.E.; Gold, D. The fate of human SUSD2+ endometrial mesenchymal stem cells during decidualization. Stem Cell Res. 2022, 60, 102671. [Google Scholar] [CrossRef] [PubMed]
- Shoyab, M.; McDonald, V.L.; Bradley, J.G.; Todaro, G.J. Amphiregulin: A bifunctional growth-modulating glycoprotein produced by the phorbol 12-myristate 13-acetate-treated human breast adenocarcinoma cell line MCF-7. Proc. Natl. Acad. Sci. USA 1988, 85, 6528–6532. [Google Scholar] [CrossRef] [PubMed]
- Kamentseva, R.; Kosheverova, V.; Kharchenko, M.; Zlobina, M.; Salova, A.; Belyaeva, T.; Nikolsky, N.; Kornilova, E. Functional cycle of EEA1-positive early endosome: Direct evidence for pre-existing compartment of degradative pathway. PLoS ONE 2020, 15, e0232532. [Google Scholar] [CrossRef] [PubMed]
- Yoshimori, T.; Yamamoto, A.; Moriyama, Y.; Futai, M.; Tashiro, Y. Bafilomycin A1, a specific inhibitor of vacuolar-type H+-ATPase, inhibits acidification and protein degradation in lysosomes of cultured cells. J. Biol. Chem. 1991, 266, 17707–17712. [Google Scholar] [CrossRef]
- Tamama, K.; Kawasaki, H.; Wells, A. Epidermal Growth Factor (EGF) Treatment on Multipotential Stromal Cells (MSCs). Possible Enhancement of Therapeutic Potential of MSC. J. Biomed. Biotechnol. 2010, 2010, 1–10. [Google Scholar] [CrossRef]
- Erikson, D.W.; Chen, J.C.; Piltonen, T.T.; Conti, M.; Irwin, J.C.; Giudice, L.C. Inhibition of epidermal growth factor receptor restores decidualization markers in stromal fibroblasts from women with endometriosis. J. Endometr. Pelvic Pain Disord. 2014, 6, 196–211. [Google Scholar] [CrossRef]
- Niikura, H.; Sasano, H.; Kaga, K.; Sato, S.; Yajima, A. Expression of epidermal growth factor family proteins and epidermal growth factor receptor in human endometrium. Hum. Pathol. 1996, 27, 282–289. [Google Scholar] [CrossRef] [PubMed]
- Gill, G.N. Regulation of EGF receptor expression and function. Mol. Reprod. Dev. 1990, 27, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Macdonald-Obermann, J.L.; Pike, L.J. Different Epidermal Growth Factor (EGF) Receptor Ligands Show Distinct Kinetics and Biased or Partial Agonism for Homodimer and Heterodimer Formation. J. Biol. Chem. 2014, 289, 26178–26188. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, S.M.; Auger, R.; Dreux, C.; Mauduit, P. Regulated Cell Surface Pro-EGF Ectodomain Shedding Is a Zinc Metalloprotease-dependent Process. J. Biol. Chem. 2003, 278, 45255–45268. [Google Scholar] [CrossRef]
- Möller, B. Expression of the angiogenic growth factors VEGF, FGF-2, EGF and their receptors in normal human endometrium during the menstrual cycle. Mol. Hum. Reprod. 2001, 7, 65–72. [Google Scholar] [CrossRef]
- Stern, K.; Place, T.; Lill, N. EGF and amphiregulin differentially regulate Cbl recruitment to endosomes and EGF receptor fate. Biochem. J. 2008, 410, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Deb, T.B.; Su, L.; Wong, L.; Bonvini, E.; Wells, A.; David, M.; Johnson, G.R. Epidermal Growth Factor (EGF) Receptor Kinase-independent Signaling by EGF. J. Biol. Chem. 2001, 276, 15554–15560. [Google Scholar] [CrossRef]
- Refaat, A.; Aminullah; Zhou, Y.; Kawanishi, M.; Tomaru, R.; Abdelhamed, S.; Shin, M.S.; Koizumi, K.; Yokoyama, S.; Saiki, I.; et al. Role of tyrosine kinase-independent phosphorylation of EGFR with activating mutation in cisplatin-treated lung cancer cells. Biochem. Biophys. Res. Commun. 2015, 458, 856–861. [Google Scholar] [CrossRef]
- Abrahamsen, H.; Stenmark, H.; Platta, H.W. Ubiquitination and phosphorylation of Beclin 1 and its binding partners: Tuning class III phosphatidylinositol 3-kinase activity and tumor suppression. FEBS Lett. 2012, 586, 1584–1591. [Google Scholar] [CrossRef]
- Pavlinov, I.; Salkovski, M.; Aldrich, L.N. Beclin 1–ATG14L Protein–Protein Interaction Inhibitor Selectively Inhibits Autophagy through Disruption of VPS34 Complex I. J. Am. Chem. Soc. 2020, 142, 8174–8182. [Google Scholar] [CrossRef]
- Birgisdottir, A.B.; Johansen, T. Autophagy and endocytosis – interconnections and interdependencies. J. Cell Sci. 2020, 133, jcs228114. [Google Scholar] [CrossRef]
- Alshammari, F.O.; Satari, A.O.; Aljabali, A.S.; Al-mahdy, Y.S.; Alabdallat, Y.J.; Al-sarayra, Y.M.; Alkhojah, M.A.; Alwardat, A.R.M.; Haddad, M.; Al-sarayreh, S.A.; et al. Glypican-3 Differentiates Intraductal Carcinoma and Paget’s Disease from Other Types of Breast Cancer. Medicina 2022, 59, 86. [Google Scholar] [CrossRef]
- Zemelko, V.I.; Grinchuk, T.M.; Domnina, A.P.; Artzibasheva, I.V.; Zenin, V.V.; Kirsanov, A.A.; Bichevaia, N.K.; Korsak, V.S.; Nikolsky, N.N. Multipotent mesenchymal stem cells of desquamated endometrium: Isolation, characterization, and application as a feeder layer for maintenance of human embryonic stem cells. Cell Tissue Biol. 2012, 6, 1–11. [Google Scholar] [CrossRef]
- Ochiai, A.; Kuroda, K.; Ozaki, R.; Ikemoto, Y.; Murakami, K.; Muter, J.; Matsumoto, A.; Itakura, A.; Brosens, J.J.; Takeda, S. Resveratrol inhibits decidualization by accelerating downregulation of the CRABP2-RAR pathway in differentiating human endometrial stromal cells. Cell Death Dis. 2019, 10, 276. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. Ilastik: Interactive Machine Learning for (Bio)Image Analysis. Nat. Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef] [PubMed]
- Bolte, S.; Cordelières, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2015. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Differentiation (0.5 M 8-Br-cAMP + 1 M P4) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| − AG-1478 | + AG-1478 | − AG-1478 | + AG-1478 | |||||||||
| w/o lig. | w/o lig. | w/o lig. | EGF | TGF- | AREG | w/o lig. | EGF | TGF- | AREG | |||
| IGFBP1 secretion | − | − | + | − | − | + | ++ | ++ | ++ | ++ | ||
| AREG Secretion, pg/mL | |||
|---|---|---|---|
| − (<15) | + (15–1000) | ++ (>1000) | |
| enMSC line 2804 | √ | ||
| enMSC line 2602 | √ | ||
| MSC-PL2 | √ | ||
| MSCWJ-1 | √ | ||
| MSC-DP | √ | ||
| HeLa | √ | ||
| MCF-7 | √ | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamentseva, R.S.; Kharchenko, M.V.; Gabdrahmanova, G.V.; Kotov, M.A.; Kosheverova, V.V.; Kornilova, E.S. EGF, TGF-α and Amphiregulin Differently Regulate Endometrium-Derived Mesenchymal Stromal/Stem Cells. Int. J. Mol. Sci. 2023, 24, 13408. https://doi.org/10.3390/ijms241713408
Kamentseva RS, Kharchenko MV, Gabdrahmanova GV, Kotov MA, Kosheverova VV, Kornilova ES. EGF, TGF-α and Amphiregulin Differently Regulate Endometrium-Derived Mesenchymal Stromal/Stem Cells. International Journal of Molecular Sciences. 2023; 24(17):13408. https://doi.org/10.3390/ijms241713408
Chicago/Turabian StyleKamentseva, Rimma Sergeevna, Marianna Viktorovna Kharchenko, Gulnara Vladikovna Gabdrahmanova, Michael Alexandrovich Kotov, Vera Vladislavovna Kosheverova, and Elena Sergeevna Kornilova. 2023. "EGF, TGF-α and Amphiregulin Differently Regulate Endometrium-Derived Mesenchymal Stromal/Stem Cells" International Journal of Molecular Sciences 24, no. 17: 13408. https://doi.org/10.3390/ijms241713408