
Immune Checkpoints in Recurrent Pregnancy Loss: New Insights into a Detrimental and Elusive Disorder
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
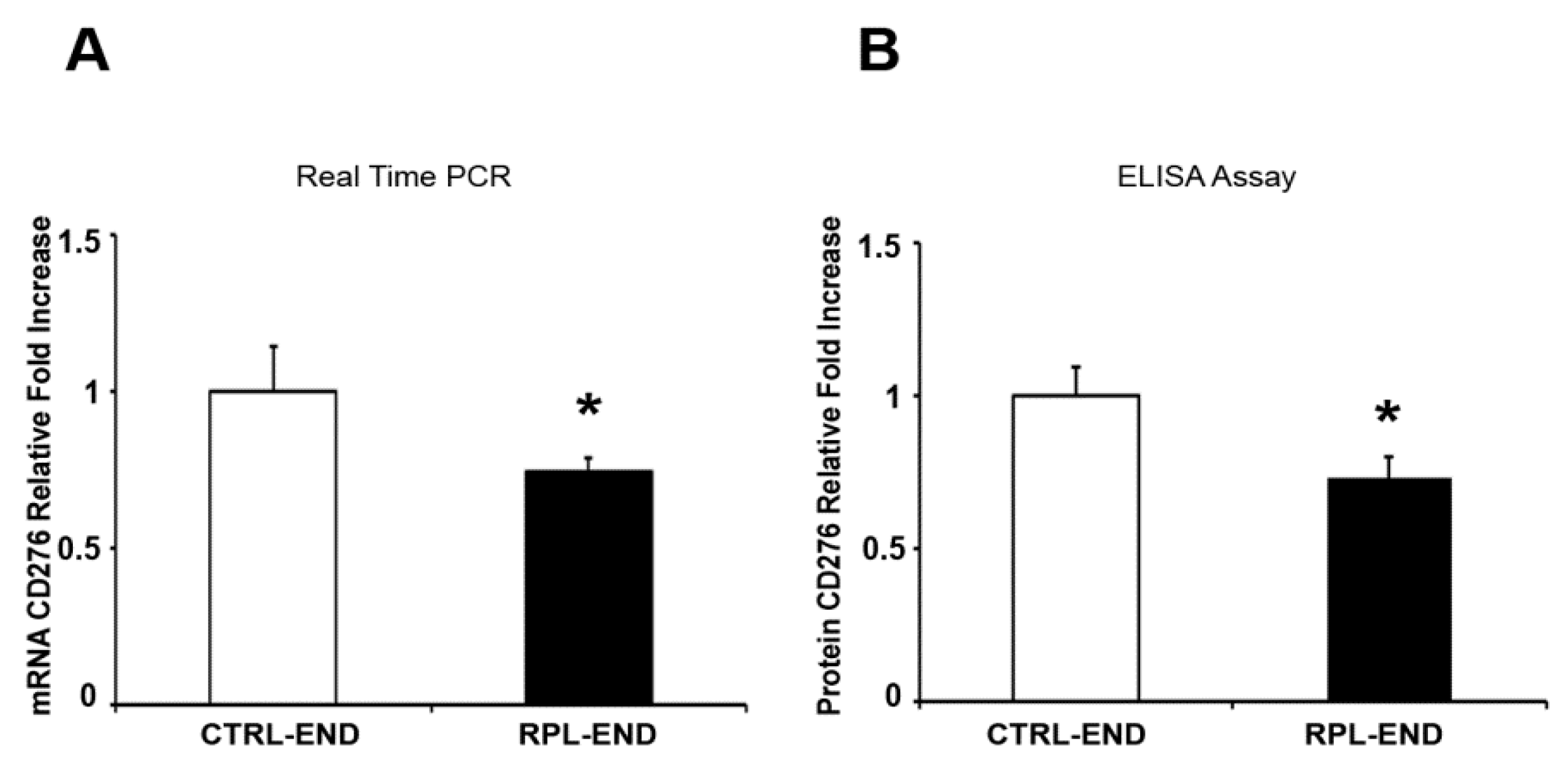
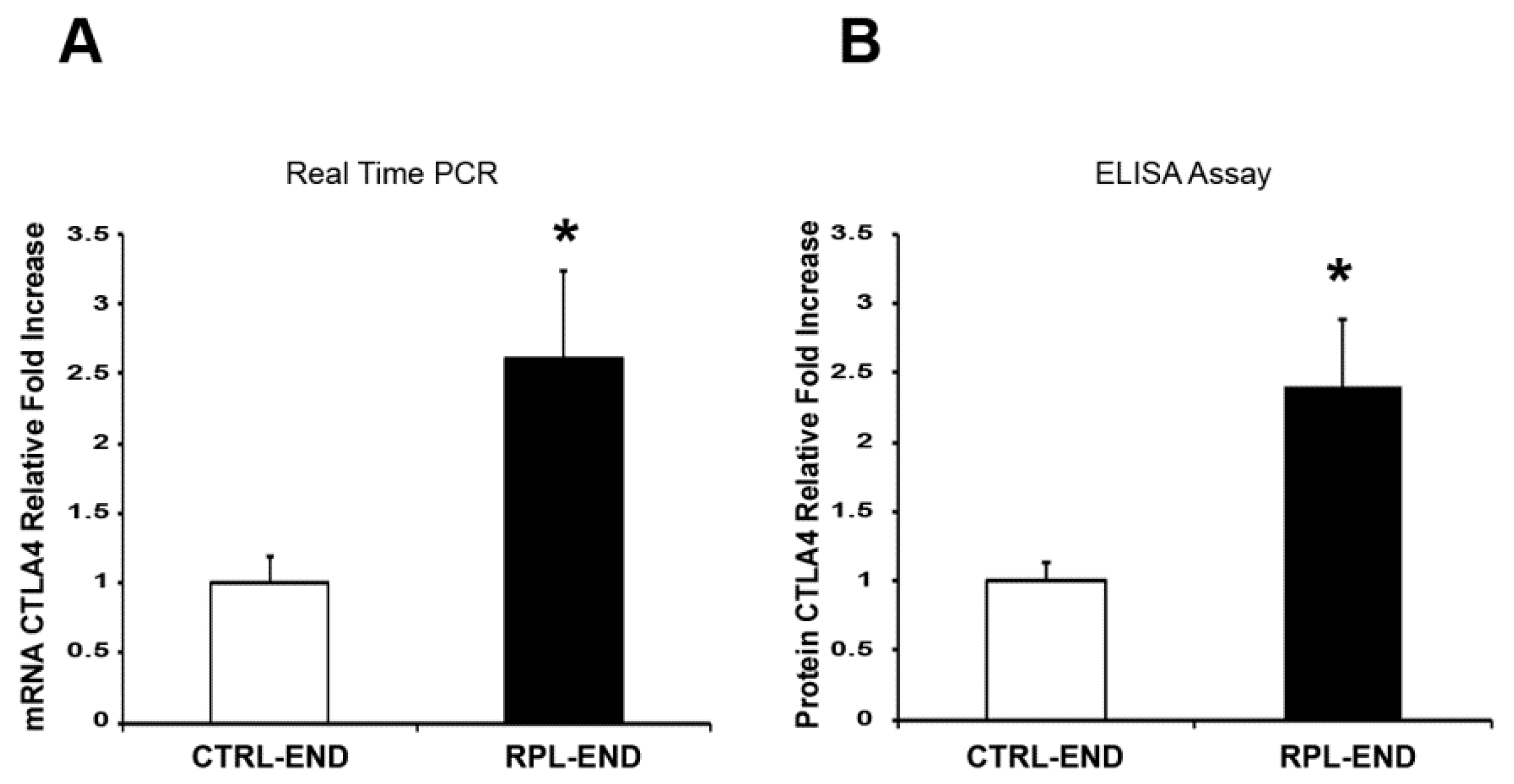
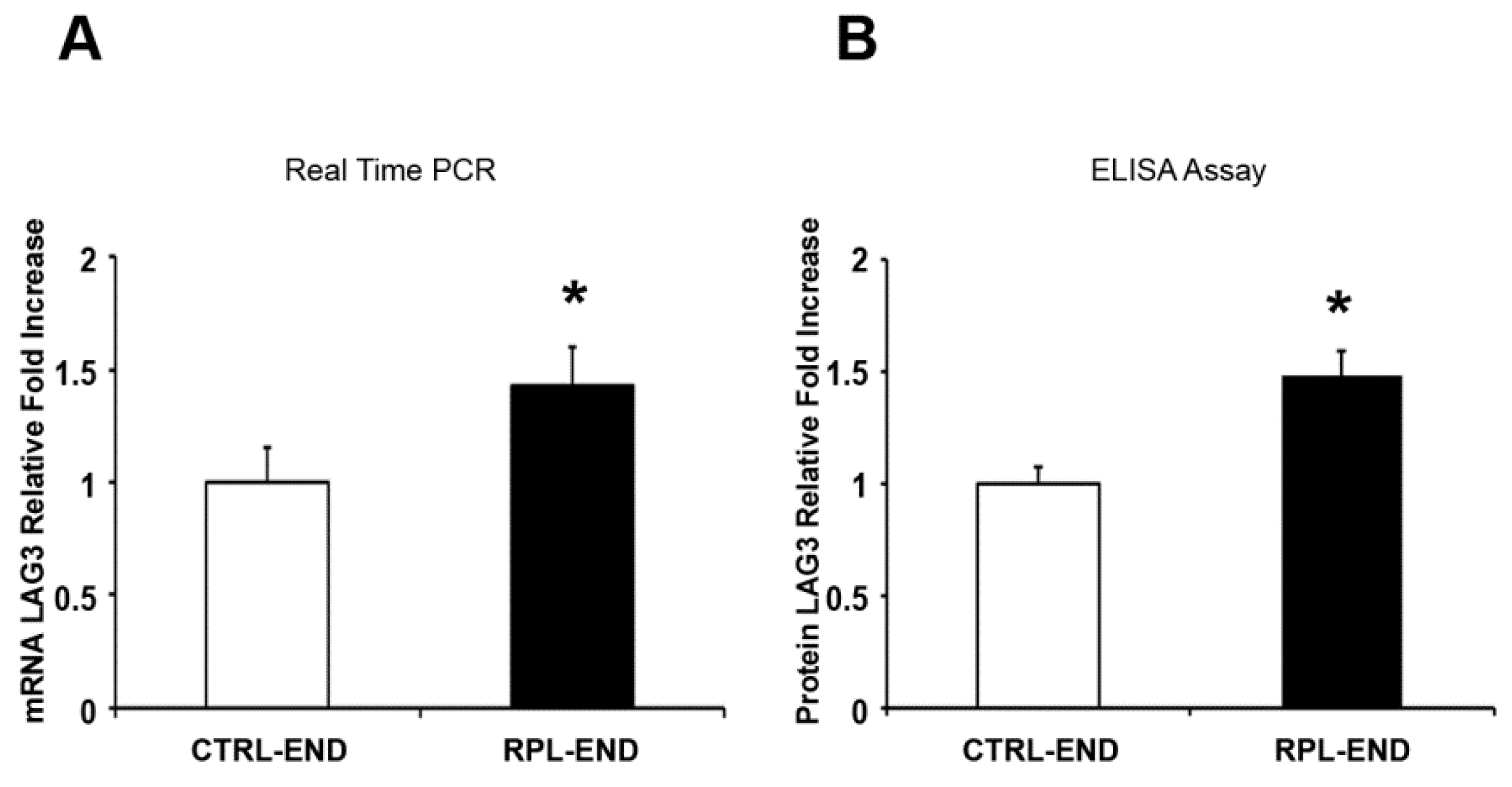
2. Results
3. Discussion
4. Materials and Methods
4.1. Immunohistochemistry (IHC)
4.2. RNA Isolation and Real-Time PCR
4.3. Protein Isolation and Enzyme-Linked ImmunoSorbent Assay (ELISA)
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ticconi, C.; Pietropolli, A.; Di Simone, N.; Piccione, E.; Fazleabas, A. Endometrial Immune Dysfunction in Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2019, 20, 5332. [Google Scholar] [CrossRef] [PubMed]
- Jaslow, C.R.; Carney, J.L.; Kutteh, W.H. Diagnostic Factors Identified in 1020 Women with Two versus Three or More Recurrent Pregnancy Losses. Fertil. Steril. 2010, 93, 1234–1243. [Google Scholar] [CrossRef] [PubMed]
- Chapoval, A.I.; Ni, J.; Lau, J.S.; Wilcox, R.A.; Flies, D.B.; Liu, D.; Dong, H.; Sica, G.L.; Zhu, G.; Tamada, K.; et al. B7-H3: A Costimulatory Molecule for T Cell Activation and IFN-Gamma Production. Nat. Immunol. 2001, 2, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Shen, Y.; Qu, Q.-X.; Chen, X.-Q.; Zhang, X.-G.; Huang, J.-A. Induced Expression of B7-H3 on the Lung Cancer Cells and Macrophages Suppresses T-Cell Mediating Anti-Tumor Immune Response. Exp. Cell Res. 2013, 319, 96–102. [Google Scholar] [CrossRef]
- Li, G.; Quan, Y.; Che, F.; Wang, L. B7-H3 in Tumors: Friend or Foe for Tumor Immunity? Cancer Chemother. Pharmacol. 2018, 81, 245–253. [Google Scholar] [CrossRef]
- Sasaki, Y.; Sakai, M.; Miyazaki, S.; Higuma, S.; Shiozaki, A.; Saito, S. Decidual and Peripheral Blood CD4+CD25+ Regulatory T Cells in Early Pregnancy Subjects and Spontaneous Abortion Cases. Mol. Hum. Reprod. 2004, 10, 347–353. [Google Scholar] [CrossRef]
- Walker, L.S.K. EFIS Lecture: Understanding the CTLA-4 Checkpoint in the Maintenance of Immune Homeostasis. Immunol. Lett. 2017, 184, 43–50. [Google Scholar] [CrossRef]
- Walker, L.S.K.; Sansom, D.M. Confusing Signals: Recent Progress in CTLA-4 Biology. Trends Immunol. 2015, 36, 63–70. [Google Scholar] [CrossRef]
- Chambers, C.A.; Kuhns, M.S.; Egen, J.G.; Allison, J.P. CTLA-4-Mediated Inhibition in Regulation of T Cell Responses: Mechanisms and Manipulation in Tumor Immunotherapy. Annu. Rev. Immunol. 2001, 19, 565–594. [Google Scholar] [CrossRef]
- Jin, L.-P.; Fan, D.-X.; Zhang, T.; Guo, P.-F.; Li, D.-J. The Costimulatory Signal Upregulation Is Associated with Th1 Bias at the Maternal-Fetal Interface in Human Miscarriage. Am. J. Reprod. Immunol. 2011, 66, 270–278. [Google Scholar] [CrossRef]
- Pardoll, D.M. The Blockade of Immune Checkpoints in Cancer Immunotherapy. Nat. Rev. Cancer 2012, 12, 252–264. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Rivard, C.J.; Rozeboom, L.; Yu, H.; Ellison, K.; Kowalewski, A.; Zhou, C.; Hirsch, F.R. Lymphocyte-Activation Gene-3, an Important Immune Checkpoint in Cancer. Cancer Sci. 2016, 107, 1193–1197. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-T.; Workman, C.J.; Flies, D.; Pan, X.; Marson, A.L.; Zhou, G.; Hipkiss, E.L.; Ravi, S.; Kowalski, J.; Levitsky, H.I.; et al. Role of LAG-3 in Regulatory T Cells. Immunity 2004, 21, 503–513. [Google Scholar] [CrossRef] [PubMed]
- Kisielow, M.; Kisielow, J.; Capoferri-Sollami, G.; Karjalainen, K. Expression of Lymphocyte Activation Gene 3 (LAG-3) on B Cells Is Induced by T Cells. Eur. J. Immunol. 2005, 35, 2081–2088. [Google Scholar] [CrossRef] [PubMed]
- Grosso, J.F.; Kelleher, C.C.; Harris, T.J.; Maris, C.H.; Hipkiss, E.L.; De Marzo, A.; Anders, R.; Netto, G.; Getnet, D.; Bruno, T.C.; et al. LAG-3 Regulates CD8+ T Cell Accumulation and Effector Function in Murine Self- and Tumor-Tolerance Systems. J. Clin. Investig. 2007, 117, 3383–3392. [Google Scholar] [CrossRef]
- Huard, B.; Prigent, P.; Pagès, F.; Bruniquel, D.; Triebel, F. T Cell Major Histocompatibility Complex Class II Molecules Down-Regulate CD4+ T Cell Clone Responses Following LAG-3 Binding. Eur. J. Immunol. 1996, 26, 1180–1186. [Google Scholar] [CrossRef]
- Hannier, S.; Triebel, F. The MHC Class II Ligand Lymphocyte Activation Gene-3 Is Co-Distributed with CD8 and CD3-TCR Molecules after Their Engagement by MAb or Peptide-MHC Class I Complexes. Int. Immunol. 1999, 11, 1745–1752. [Google Scholar] [CrossRef]
- Gleicher, N.; Kushnir, V.A.; Barad, D.H. Redirecting Reproductive Immunology Research toward Pregnancy as a Period of Temporary Immune Tolerance. J. Assist. Reprod. Genet. 2017, 34, 425–430. [Google Scholar] [CrossRef]
- Rimmer, M.P.; Fishwick, K.; Henderson, I.; Chinn, D.; Al Wattar, B.H.; Quenby, S. Quantifying CD138+ Cells in the Endometrium to Assess Chronic Endometritis in Women at Risk of Recurrent Pregnancy Loss: A Prospective Cohort Study and Rapid Review. J. Obstet. Gynaecol. Res. 2021, 47, 689–697. [Google Scholar] [CrossRef]
- Yahata, T.; Kurabayashi, T.; Honda, A.; Takakuwa, K.; Tanaka, K.; Abo, T. Decrease in the Proportion of Granulated CD56+ T-Cells in Patients with a History of Recurrent Abortion. J. Reprod. Immunol. 1998, 38, 63–73. [Google Scholar] [CrossRef]
- Lachapelle, M.H.; Hemmings, R.; Roy, D.C.; Falcone, T.; Miron, P. Flow Cytometric Evaluation of Leukocyte Subpopulations in the Follicular Fluids of Infertile Patients. Fertil. Steril. 1996, 65, 1135–1140. [Google Scholar] [CrossRef] [PubMed]
- Galgani, M.; Insabato, L.; Calì, G.; Della Gatta, A.N.; Mirra, P.; Papaccio, F.; Santopaolo, M.; Alviggi, C.; Mollo, A.; Strina, I.; et al. Regulatory T Cells, Inflammation, and Endoplasmic Reticulum Stress in Women with Defective Endometrial Receptivity. Fertil. Steril. 2015, 103, 1579–1586.e1. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Zhu, G.; Xu, H.; Yao, S.; Zhou, G.; Zhu, Y.; Tamada, K.; Huang, L.; Flies, A.D.; Broadwater, M.; et al. B7-H3 Promotes Pathogenesis of Autoimmune Disease and Inflammation by Regulating the Activity of Different T Cell Subsets. PLoS ONE 2015, 10, e0130126. [Google Scholar] [CrossRef]
- Prasad, D.V.R.; Nguyen, T.; Li, Z.; Yang, Y.; Duong, J.; Wang, Y.; Dong, C. Murine B7-H3 Is a Negative Regulator of T Cells. J. Immunol. 2004, 173, 2500–2506. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Cao, N.-N.; Wang, S.; Man, H.-W.; Li, P.-F.; Shan, B.-E. Roles of Coinhibitory Molecules B7-H3 and B7-H4 in Esophageal Squamous Cell Carcinoma. Tumour Biol. 2016, 37, 2961–2971. [Google Scholar] [CrossRef]
- Zhao, Y.; Zheng, Q.; Jin, L. The Role of B7 Family Molecules in Maternal-Fetal Immunity. Front. Immunol. 2020, 11, 458. [Google Scholar] [CrossRef] [PubMed]
- Gardner, D.; Jeffery, L.E.; Sansom, D.M. Understanding the CD28/CTLA-4 (CD152) Pathway and Its Implications for Costimulatory Blockade. Am. J. Transplant. 2014, 14, 1985–1991. [Google Scholar] [CrossRef]
- Ding, J.-L.; Diao, L.-H.; Yin, T.-L.; Huang, C.-Y.; Yin, B.; Chen, C.; Zhang, Y.; Li, J.; Cheng, Y.-X.; Zeng, Y.; et al. Aberrant Expressions of Endometrial Id3 and CTLA-4 Are Associated with Unexplained Repeated Implantation Failure and Recurrent Miscarriage. Am. J. Reprod. Immunol. 2017, 78, e12632. [Google Scholar] [CrossRef]
- Tafuri, A.; Alferink, J.; Möller, P.; Hämmerling, G.J.; Arnold, B. T Cell Awareness of Paternal Alloantigens during Pregnancy. Science 1995, 270, 630–633. [Google Scholar] [CrossRef]
- Mellor, A.L.; Munn, D.H. Immunology at the Maternal-Fetal Interface: Lessons for T Cell Tolerance and Suppression. Annu. Rev. Immunol. 2000, 18, 367–391. [Google Scholar] [CrossRef]
- Moffett, A.; Loke, Y.W. The Immunological Paradox of Pregnancy: A Reappraisal. Placenta 2004, 25, 1–8. [Google Scholar] [CrossRef]
- Baixeras, E.; Huard, B.; Miossec, C.; Jitsukawa, S.; Martin, M.; Hercend, T.; Auffray, C.; Triebel, F.; Piatier-Tonneau, D. Characterization of the Lymphocyte Activation Gene 3-Encoded Protein. A New Ligand for Human Leukocyte Antigen Class II Antigens. J. Exp. Med. 1992, 176, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Annunziato, F.; Manetti, R.; Tomasévic, I.; Guidizi, M.G.; Biagiotti, R.; Giannò, V.; Germano, P.; Mavilia, C.; Maggi, E.; Romagnani, S. Expression and Release of LAG-3-Encoded Protein by Human CD4+ T Cells Are Associated with IFN-Gamma Production. FASEB J. 1996, 10, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Avice, M.N.; Sarfati, M.; Triebel, F.; Delespesse, G.; Demeure, C.E. Lymphocyte Activation Gene-3, a MHC Class II Ligand Expressed on Activated T Cells, Stimulates TNF-Alpha and IL-12 Production by Monocytes and Dendritic Cells. J. Immunol. 1999, 162, 2748–2753. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K. Impaired Endometrial Function and Unexplained Recurrent Pregnancy Loss. Hypertens. Res. Pregnancy 2019, 7, 16–21. [Google Scholar] [CrossRef]
- Haudebourg, T.; Dugast, A.-S.; Coulon, F.; Usal, C.; Triebel, F.; Vanhove, B. Depletion of LAG-3 Positive Cells in Cardiac Allograft Reveals Their Role in Rejection and Tolerance. Transplantation 2007, 84, 1500–1506. [Google Scholar] [CrossRef]
- Poirier, N.; Haudebourg, T.; Brignone, C.; Dilek, N.; Hervouet, J.; Minault, D.; Coulon, F.; de Silly, R.V.; Triebel, F.; Blancho, G.; et al. Antibody-Mediated Depletion of Lymphocyte-Activation Gene-3 (LAG-3(+) )-Activated T Lymphocytes Prevents Delayed-Type Hypersensitivity in Non-Human Primates. Clin. Exp. Immunol. 2011, 164, 265–274. [Google Scholar] [CrossRef]
- Ellis, J.; Marks, D.; Barrett, C.; Hopkins, T.; Richards, A.; Fuhr, R.; Albayaty, M.; Coenen, M.; Liefaard, L.; Leavens, K.; et al. P449 Selective Depletion of LAG3+ Cells in T-Cell-Driven Inflammation: A Randomised, Double-Blind, Placebo-Controlled, FTIH Phase I/Ib Clinical Trial. J. Crohn’s Colitis 2019, 13, S336. [Google Scholar] [CrossRef]
- Bouet, P.-E.; El Hachem, H.; Monceau, E.; Gariépy, G.; Kadoch, I.-J.; Sylvestre, C. Chronic Endometritis in Women with Recurrent Pregnancy Loss and Recurrent Implantation Failure: Prevalence and Role of Office Hysteroscopy and Immunohistochemistry in Diagnosis. Fertil. Steril. 2016, 105, 106–110. [Google Scholar] [CrossRef]
- McQueen, D.B.; Perfetto, C.O.; Hazard, F.K.; Lathi, R.B. Pregnancy Outcomes in Women with Chronic Endometritis and Recurrent Pregnancy Loss. Fertil. Steril. 2015, 104, 927–931. [Google Scholar] [CrossRef]
- Kitaya, K. Prevalence of Chronic Endometritis in Recurrent Miscarriages. Fertil. Steril. 2011, 95, 1156–1158. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Kimura, F.; Zheng, L.; Ishida, M.; Niwa, Y.; Hirata, K.; Takebayashi, A.; Takashima, A.; Takahashi, K.; Kushima, R.; et al. Chronic Endometritis Modifies Decidualization in Human Endometrial Stromal Cells. Reprod. Biol. Endocrinol. 2017, 15, 16. [Google Scholar] [CrossRef] [PubMed]
- Cicinelli, E.; Matteo, M.; Tinelli, R.; Pinto, V.; Marinaccio, M.; Indraccolo, U.; De Ziegler, D.; Resta, L. Chronic Endometritis Due to Common Bacteria Is Prevalent in Women with Recurrent Miscarriage as Confirmed by Improved Pregnancy Outcome after Antibiotic Treatment. Reprod. Sci. 2014, 21, 640–647. [Google Scholar] [CrossRef] [PubMed]
- Confavreux, C.; Hutchinson, M.; Hours, M.M.; Cortinovis-Tourniaire, P.; Moreau, T. Rate of Pregnancy-Related Relapse in Multiple Sclerosis. Pregnancy in Multiple Sclerosis Group. N. Engl. J. Med. 1998, 339, 285–291. [Google Scholar] [CrossRef]
- Buchel, E.; Van Steenbergen, W.; Nevens, F.; Fevery, J. Improvement of Autoimmune Hepatitis during Pregnancy Followed by Flare-up after Delivery. Am. J. Gastroenterol. 2002, 97, 3160–3165. [Google Scholar] [CrossRef]
- Patas, K.; Engler, J.B.; Friese, M.A.; Gold, S.M. Pregnancy and Multiple Sclerosis: Feto-Maternal Immune Cross Talk and Its Implications for Disease Activity. J. Reprod. Immunol. 2013, 97, 140–146. [Google Scholar] [CrossRef]
- Coyle, P.K. Pregnancy and Multiple Sclerosis. Neurol. Clin. 2012, 30, 877–888. [Google Scholar] [CrossRef]
- Mor, G.; Aldo, P.; Alvero, A.B. The Unique Immunological and Microbial Aspects of Pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Characteristic | RPL Group (N = 27) | Control Group (N = 29) | p-Value |
|---|---|---|---|
| Age at hysteroscopy—years, mean ± SE | 36.4 ± 0.78 | 38.1 ± 0.74 | NS |
| Body-mass index (kg/m2)—mean ± SE | 23.4 ± 0.24 | 24.2 ± 0.27 | NS |
| Tobacco use—no. (%) | 2 (7.4) | 1 (3.7) | NS |
| Caucasian ethnicity—no. (%) | 23 (85.2) | 24 (82.7) | NS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marozio, L.; Nuzzo, A.M.; Gullo, E.; Moretti, L.; Canuto, E.M.; Tancredi, A.; Goia, M.; Cosma, S.; Revelli, A.; Rolfo, A.; et al. Immune Checkpoints in Recurrent Pregnancy Loss: New Insights into a Detrimental and Elusive Disorder. Int. J. Mol. Sci. 2023, 24, 13071. https://doi.org/10.3390/ijms241713071
Marozio L, Nuzzo AM, Gullo E, Moretti L, Canuto EM, Tancredi A, Goia M, Cosma S, Revelli A, Rolfo A, et al. Immune Checkpoints in Recurrent Pregnancy Loss: New Insights into a Detrimental and Elusive Disorder. International Journal of Molecular Sciences. 2023; 24(17):13071. https://doi.org/10.3390/ijms241713071
Chicago/Turabian StyleMarozio, Luca, Anna Maria Nuzzo, Eugenio Gullo, Laura Moretti, Emilie M. Canuto, Annalisa Tancredi, Margherita Goia, Stefano Cosma, Alberto Revelli, Alessandro Rolfo, and et al. 2023. "Immune Checkpoints in Recurrent Pregnancy Loss: New Insights into a Detrimental and Elusive Disorder" International Journal of Molecular Sciences 24, no. 17: 13071. https://doi.org/10.3390/ijms241713071