
Taxifolin Reduces Blood Pressure via Improvement of Vascular Function and Mitigating the Vascular Inflammatory Response in Spontaneously Hypertensive Rats
 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Basic Characteristics of Experimental Animals
2.2. Vascular Responses of Isolated Femoral Arteries
2.2.1. NA-Induced Contraction of Isolated Femoral Arteries
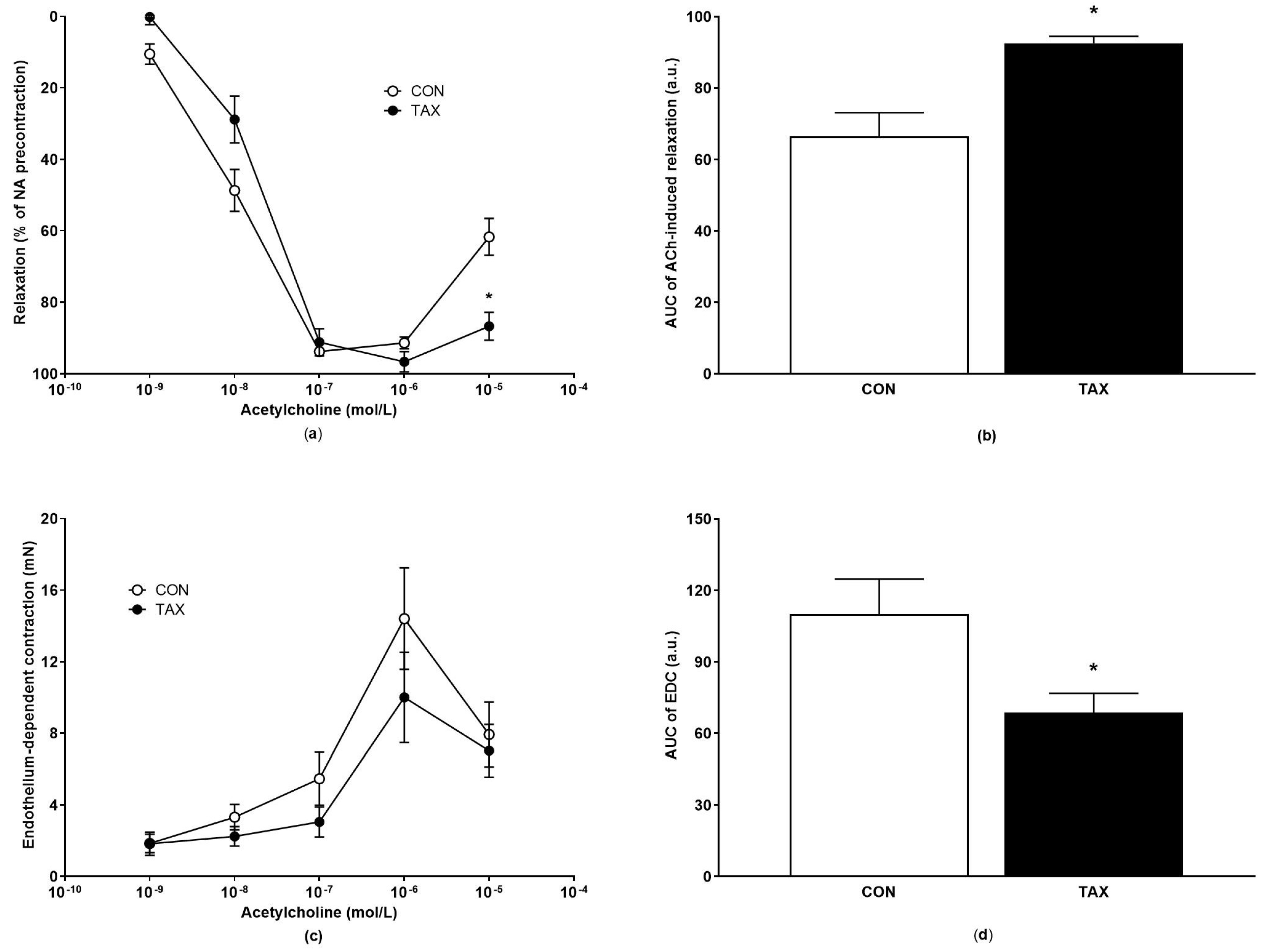
2.2.2. ACh-Induced Endothelium-Dependent Relaxation and Endothelium-Dependent Contraction of Isolated Femoral Arteries
2.3. Vascular Responses of Isolated Thoracic Aortas
2.4. Total NOS Activity and Superoxide Production in Tissue of Abdominal Aorta
2.5. Protein Expressions of eNOS, iNOS, and COX2 in Tissue and Gene Expression of Corresponding Genes in Abdominal Aorta
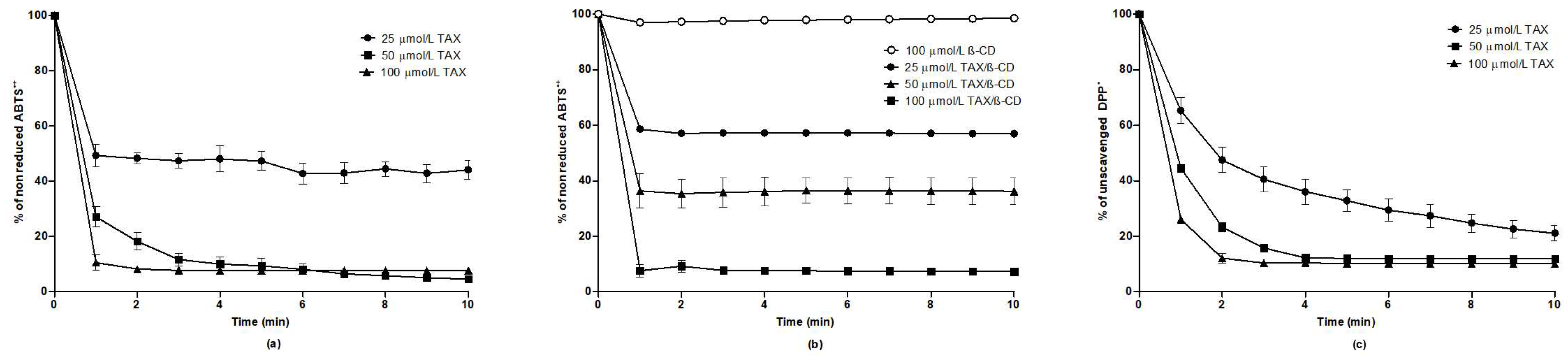
2.6. Evaluation of Antioxidant Effects of TAX In Vitro
3. Discussion
4. Materials and Methods
4.1. Description of Experimental Model and Blood Pressure Measurement
4.2. Assessment of Vascular Responses
4.3. Protein Expression
4.4. Gene Expressions
4.5. Superoxide Production Measurement
4.6. Total NOS Activity Measurement
4.7. Measurements of Antioxidant and Free Radical Scavenging Properties of TAX
4.8. Chemicals
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uchmanowicz, B.; Chudiak, A.; Mazur, G. The influence of quality of life on the level of adherence to therapeutic recommendations among elderly hypertensive patients. Patient Prefer. Adherence 2018, 12, 2593–2603. [Google Scholar] [CrossRef] [Green Version]
- Shubina, V.S.; Shatalin, Y.V. Antioxidant and iron-chelating properties of taxifolin and its condensation product with glyoxylic acid. J. Food Sci. Technol. 2017, 54, 1467–1475. [Google Scholar] [CrossRef] [Green Version]
- Bernatova, I.; Liskova, S. Mechanisms modified by (−)-epicatechin and taxifolin relevant for the treatment of hypertension and viral infection: Knowledge from preclinical studies. Antioxidants 2021, 10, 467. [Google Scholar] [CrossRef]
- Hämäläinen, M.; Nieminen, R.; Asmawi, M.Z.; Vuorela, P.; Vapaatalo, H.; Moilanen, E. Effects of flavonoids on prostaglandin E2 production and on COX-2 and MPGES-1 expressions in activated macrophages. Planta Med. 2011, 77, 1504–1511. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Sellner, M.; Neranjan, S.; Smieško, M.; Lill, M.A. Potential inhibitors for novel coronavirus protease identified by virtual screening of 606 million compounds. Int. J. Mol. Sci. 2020, 21, 3626. [Google Scholar] [CrossRef] [PubMed]
- Zicha, J.; Kunes, J. Ontogenetic aspects of hypertension development: Analysis in the rat. Physiol. Rev. 1999, 79, 1227–1282. [Google Scholar] [CrossRef] [PubMed]
- Arribas, S.M.; Hinek, A.; González, M.C. Elastic fibres and vascular structure in hypertension. Pharmacol. Ther. 2006, 111, 771–791. [Google Scholar] [CrossRef] [PubMed]
- Berenyiova, A.; Drobna, M.; Cebova, M.; Kristek, F.; Cacanyiova, S. Changes in the vasoactive effects of nitric oxide, hydrogen sulfide and the structure of the rat thoracic aorta: The role of age and essential hypertension. J. Physiol. Pharmacol. 2018, 69, 4. [Google Scholar] [CrossRef]
- Kluknavsky, M.; Balis, P.; Puzserova, A.; Radosinska, J.; Berenyiova, A.; Drobna, M.; Lukac, S.; Muchova, J.; Bernatova, I. (−)-epicatechin prevents blood pressure increase and reduces locomotor hyperactivity in young spontaneously hypertensive rats. Oxid. Med. Cell. Longev. 2016, 2016, 6949020. [Google Scholar] [CrossRef] [Green Version]
- Líšková, S.; Petrová, M.; Karen, P.; Kuneš, J.; Zicha, J. Effects of aging and hypertension on the participation of endothelium-derived constricting factor (EDCF) in norepinephrine-induced contraction of rat femoral artery. Eur. J. Pharmacol. 2011, 667, 265–270. [Google Scholar] [CrossRef]
- Bernatova, I. Endothelial dysfunction in experimental models of arterial hypertension: Cause or consequence? BioMed Res. Int. 2014, 2014, 598271. [Google Scholar] [CrossRef] [Green Version]
- Koga, T.; Takata, Y.; Kobayashi, K.; Takishita, S.; Yamashita, Y.; Fujishima, M. Age and hypertension promote endothelium-dependent contractions to acetylcholine in the aorta of the rat. Hypertension 1989, 14, 542–548. [Google Scholar] [CrossRef] [Green Version]
- Heijnen, B.F.J.; Van Essen, H.; Schalkwijk, C.G.; Janssen, B.J.A.; Struijker-Boudier, H.A.J. Renal inflammatory markers during the onset of hypertension in spontaneously hypertensive rats. Hypertens. Res. 2014, 37, 100–109. [Google Scholar] [CrossRef]
- Dinh, Q.N.; Drummond, G.R.; Sobey, C.G.; Chrissobolis, S. Roles of inflammation, oxidative stress, and vascular dysfunction in hypertension. BioMed Res. Int. 2014, 2014, 406960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solak, Y.; Afsar, B.; Vaziri, N.D.; Aslan, G.; Yalcin, C.E.; Covic, A.; Kanbay, M. Hypertension as an autoimmune and inflammatory disease. Hypertens. Res. 2016, 39, 567–573. [Google Scholar] [CrossRef]
- Murray, E.C.; Nosalski, R.; MacRitchie, N.; Tomaszewski, M.; Maffia, P.; Harrison, D.G.; Guzik, T.J. Therapeutic targeting of inflammation in hypertension: From novel mechanisms to translational perspective. Cardiovasc. Res. 2021, 117, 2589–2609. [Google Scholar] [CrossRef]
- Althunibat, O.Y.; Abukhalil, M.H.; Jghef, M.M.; Alfwuaires, M.A.; Algefare, A.I.; Alsuwayt, B.; Alazragi, R.; Abourehab, M.A.S.; Almuqati, A.F.; Karimulla, S.; et al. Hepatoprotective effect of taxifolin on cyclophosphamide-induced oxidative stress, inflammation, and apoptosis in mice: Involvement of NRF2/HO-1 signaling. Biomol. Biomed. 2023, 23, 649–660. [Google Scholar] [CrossRef] [PubMed]
- Arutyunyan, T.V.; Korystova, A.F.; Kublik, L.N.; Levitman, M.; Shaposhnikova, V.V.; Korystov, Y.N. Effects of taxifolin on the activity of angiotensin-converting enzyme and reactive oxygen and nitrogen species in the aorta of aging rats and rats treated with the nitric oxide synthase inhibitor and dexamethasone. Age 2013, 35, 2089–2097. [Google Scholar] [CrossRef] [Green Version]
- Plotnikov, M.B.; Aliev, O.I.; Sidekhmenova, A.V.; Shamanaev, A.Y.; Anishchenko, A.M.; Nosarev, A.V.; Pushkina, E.A. Modes of hypotensive action of dihydroquercetin in arterial hypertension. Bull. Exp. Biol. Med. 2017, 162, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Slashcheva, G.A.; Rykov, V.A.; Lobanov, A.V.; Murashev, A.N.; Kim, Y.A.; Arutyunyan, T.V.; Korystova, A.F.; Kublik, L.N.; Levitman, M.K.; Shaposhnikona, V.V.; et al. Dihydroquercetin does not affect age-dependent increase in blood pressure and angiotensin-converting enzyme activity in the aorta of hypertensive rats. Bull. Exp. Biol. Med. 2016, 161, 670–673. [Google Scholar] [CrossRef] [PubMed]
- Berenyiova, A.; Dovinova, I.; Kvandova, M.; Kristek, F.; Jansen, E.; Majzunova, M.; Cacanyiova, S. The effect of chronic no synthase inhibition on the vasoactive and structural properties of thoracic aorta, no synthase activity, and oxidative stress biomarkers in young SHR. Oxid. Med. Cell. Longev. 2018, 2018, 2502843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernátová, I.; Kopincová, J.; Púzserová, A.; Janega, P.; Babál, P. Chronic low-dose L-NAME treatment increases nitric oxide production and vasorelaxation in normotensive rats. Physiol. Res. 2007, 56 (Suppl. S2), S17–S24. [Google Scholar] [CrossRef]
- Leloup, A.J.; Van Hove, C.E.; Heykers, A.; Schrijvers, D.M.; De Meyer, G.R.; Fransen, P. Elastic and muscular arteries differ in structure, basal no production and voltage-gated Ca2+-channels. Front. Physiol. 2015, 6, 375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, I.S.; Sidekhmenova, A.V.; Nosarev, A.V.; Tyukavkina, N.A.; Plotnikov, M.B. Effect of dihydroquercetin on the tone of isolated rat veins. Bull. Exp. Biol. Med. 2013, 155, 65–66. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Fusi, F.; Valoti, M. Perivascular adipose tissue modulates the effects of flavonoids on rat aorta rings: Role of superoxide anion and β(3) receptors. Pharmacol. Res. 2022, 180, 106231. [Google Scholar] [CrossRef]
- Seong, E.H.; Gong, D.S.; Shiwakoti, S.; Adhikari, D.; Kim, H.J.; Oak, M.H. Taxifolin as a major bioactive compound in the vasorelaxant effect of different pigmented rice bran extracts. Front. Pharmacol. 2022, 13, 799064. [Google Scholar] [CrossRef]
- Choi, S.E.; Park, K.H.; Han, B.H.; Jeong, M.S.; Seo, S.J.; Lee, D.I.; Joo, S.S.; Lee, M.W. Inhibition of inducible nitric oxide synthase and cyclooxygenase-2 expression by phenolic compounds from roots of rhododendron mucronulatum. Phytother. Res. 2011, 25, 1301–1305. [Google Scholar] [CrossRef]
- Zhang, X.; Lian, X.; Li, H.; Zhao, W.; Li, X.; Zhou, F.; Zhou, Y.; Cui, T.; Wang, Y.; Liu, C. Taxifolin attenuates inflammation via suppressing MAPK signal pathway in vitro and in silico analysis. Chin. Herb. Med. 2022, 14, 554–562. [Google Scholar] [CrossRef]
- Hou, J.; Hu, M.; Zhang, L.; Gao, Y.; Ma, L.; Xu, Q. Dietary taxifolin protects against dextran sulfate sodium-induced colitis via nf-κb signaling, enhancing intestinal barrier and modulating gut microbiota. Front. Immunol. 2020, 11, 631809. [Google Scholar] [CrossRef]
- Okkay, U.; Ferah Okkay, I.; Cicek, B.; Aydin, I.C.; Ozkaraca, M. Hepatoprotective and neuroprotective effect of taxifolin on hepatic encephalopathy in rats. Metab. Brain Dis. 2022, 37, 1541–1556. [Google Scholar] [CrossRef]
- Tinsley, J.H.; South, S.; Chiasson, V.L.; Mitchell, B.M. Interleukin-10 reduces inflammation, endothelial dysfunction, and blood pressure in hypertensive pregnant rats. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2010, 298, R713–R719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berenyiova, A.; Bernatova, I.; Zemancikova, A.; Drobna, M.; Cebova, M.; Golas, S.; Balis, P.; Liskova, S.; Valaskova, Z.; Krskova, K.; et al. Vascular effects of low-dose ACE2 inhibitor MLN-4760 benefit or detriment in essential hypertension? Biomedicines 2022, 10, 38. [Google Scholar] [CrossRef] [PubMed]
- Liskova, S.; Petrova, M.; Karen, P.; Behuliak, M.; Zicha, J. Contribution of Ca²⁺-dependent Cl⁻ channels to norepinephrine-induced contraction of femoral artery is replaced by increasing EDCF contribution during ageing. BioMed Res. Int. 2014, 2014, 289361. [Google Scholar] [CrossRef] [PubMed]
- Líšková, S.; Bališ, P.; Mičurová, A.; Kluknavský, M.; Okuliarová, M.; Puzserová, A.; Škrátek, M.; Sekaj, I.; Maňka, J.; Valovič, P.; et al. Effect of iron oxide nanoparticles on vascular function and nitric oxide production in acute stress-exposed rats. Physiol. Res. 2020, 69, 1067–1083. [Google Scholar] [CrossRef] [PubMed]
- Cacanyiova, S.; Berenyiova, A. In vitro measurement of H2S-mediated vasoactive responses. Methods Mol. Biol. 2019, 2007, 89–107. [Google Scholar] [CrossRef]
- Lísková, S.; Petrová, M.; Karen, P.; Kunes, J.; Zicha, J. Influence of calcium-dependent potassium channel blockade and nitric oxide inhibition on norepinephrine-induced contractions in two forms of genetic hypertension. J. Am. Soc. Hypertens. 2010, 4, 128–134. [Google Scholar] [CrossRef]
- Kosutova, M.; Pechanova, O.; Barta, A.; Franova, S.; Cebova, M. Different adaptive NO-dependent mechanisms in normal and hypertensive conditions. Molecules 2019, 24, 1682. [Google Scholar] [CrossRef] [Green Version]
- Berenyiova, A.; Golas, S.; Drobna, M.; Cebova, M.; Cacanyiova, S. Fructose intake impairs the synergistic vasomotor manifestation of nitric oxide and hydrogen sulfide in rat aorta. Int. J. Mol. Sci. 2021, 22, 4749. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-delta delta c(t)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pechánová, O.; Bernátová, I.; Pelouch, V.; Simko, F. Protein remodelling of the heart in NO-deficient hypertension: The effect of captopril. J. Mol. Cell. Cardiol. 1997, 29, 3365–3374. [Google Scholar] [CrossRef]
- Valachová, K.; Mach, M.; Šoltés, L. Oxidative degradation of high-molar-mass hyaluronan: Effects of some indole derivatives to hyaluronan decay. Int. J. Mol. Sci. 2020, 21, 5609. [Google Scholar] [CrossRef] [PubMed]
- Heřmánková, E.; Zatloukalová, M.; Biler, M.; Sokolová, R.; Bancířová, M.; Tzakos, A.G.; Křen, V.; Kuzma, M.; Trouillas, P.; Vacek, J. Redox properties of individual quercetin moieties. Free Radic. Biol. Med. 2019, 143, 240–251. [Google Scholar] [CrossRef] [PubMed]
- Valachová, K.; Baňasová, M.; Topoľská, D.; Sasinková, V.; Juránek, I.; Collins, M.N.; Šoltés, L. Influence of tiopronin, captopril and levamisole therapeutics on the oxidative degradation of hyaluronan. Carbohydr. Polym. 2015, 134, 516–523. [Google Scholar] [CrossRef]
- Faul, F.; Erdfelder, E.; Lang, A.-G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Met. 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CON | TAX | |
|---|---|---|
| n = 8 | n = 8 | |
| Δ Body weight (g) | 17 ± 2 | 12 ± 3 |
| Δ BP (mmHg) | 20 ± 6 | −10 ± 4 * |
| Δ Heart rate (bpm) | −7 ± 27 | 16 ± 21 |
| CON | TAX | |
|---|---|---|
| n = 8 | n = 8 | |
| Length of the segments (mm) | 1.54 ± 0.03 | 1.56 ± 0.03 |
| EC50NA femoral a. (−log mol/L) | 5.12 ± 0.90 | 5.13 ± 0.11 |
| EmaxNA femoral a. (mN) | 14.90 ± 1.20 | 12.90 ± 1.20 |
| EC50ACh femoral a. (−log mol/L) | 8.22 ± 0.11 | 8.19 ± 0.09 |
| EmaxACh femoral a. (%) | 83.00 ± 2.00 | 85.00 ± 5.00 |
| EmaxKCl femoral a. (mN) | 24.00 ± 0.90 | 25.70 ± 1.00 |
| CON | TAX | |
|---|---|---|
| n = 8 | n = 8 | |
| EC50NA thoracic aorta (−log mol/L) | 7.71 ± 0.12 | 7.65 ± 0.15 |
| EmaxNA thoracic aorta (mN) | 0.46 ± 0.03 | 0.56 ± 0.05 |
| EC50ACh thoracic aorta (−log mol/L) | 7.66 ± 0.17 | 8.01 ± 0.06 |
| EmaxACh thoracic aorta (%) | 92.00 ± 7.00 | 75.00 ± 5.00 |
| Gene | Forward Primer | Reverse Primer | Tm | Amplicon Size |
|---|---|---|---|---|
| (°C) | (bp) | |||
| Nos2 (NM_012611.3) | AAA CGC TAC ACT TCC AAC GC | TGC TGA GAG CTT TGT TGA GGT C | 59 | 91 |
| Nos3 (NM_021838.2) | GAT CCC CCG GAG AAT GGA GA | TCG GAT TTT GTA ACT CTT GTG CT | 60 | 105 |
| Tnf (NM_012675.3) | CGT CAG CCG ATT TGC CAT TTC | TGG GCT CAT ACC AGG GCT T | 60 | 116 |
| Il10 (NM_012854.2) | GGC TCA GCA CTG CTA TGT TG | GAG CAT GTG GGT CTG GCT G | 59 | 117 |
| Ptgs2 (NM_017232.3) | CTA CCA TCT GGC TTC GGG AG | TGG AAC AGT CGC TCG TCA TC | 60 | 85 |
| Rpl10a (NM_031065.1) | TCC ACC TGG CTG TCA ACT TC | GGC AGC AAC GAG GTT TAT TGG | 60 | 134 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liskova, S.; Cacanyiova, S.; Cebova, M.; Berenyiova, A.; Kluknavsky, M.; Micurova, A.; Valachova, K.; Soltes, L.; Bernatova, I. Taxifolin Reduces Blood Pressure via Improvement of Vascular Function and Mitigating the Vascular Inflammatory Response in Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2023, 24, 12616. https://doi.org/10.3390/ijms241612616
Liskova S, Cacanyiova S, Cebova M, Berenyiova A, Kluknavsky M, Micurova A, Valachova K, Soltes L, Bernatova I. Taxifolin Reduces Blood Pressure via Improvement of Vascular Function and Mitigating the Vascular Inflammatory Response in Spontaneously Hypertensive Rats. International Journal of Molecular Sciences. 2023; 24(16):12616. https://doi.org/10.3390/ijms241612616
Chicago/Turabian StyleLiskova, Silvia, Sona Cacanyiova, Martina Cebova, Andrea Berenyiova, Michal Kluknavsky, Andrea Micurova, Katarina Valachova, Ladislav Soltes, and Iveta Bernatova. 2023. "Taxifolin Reduces Blood Pressure via Improvement of Vascular Function and Mitigating the Vascular Inflammatory Response in Spontaneously Hypertensive Rats" International Journal of Molecular Sciences 24, no. 16: 12616. https://doi.org/10.3390/ijms241612616