
GADD45A and GADD45B as Novel Biomarkers Associated with Chromatin Regulators in Renal Ischemia-Reperfusion Injury

Abstract
:1. Introduction
2. Results
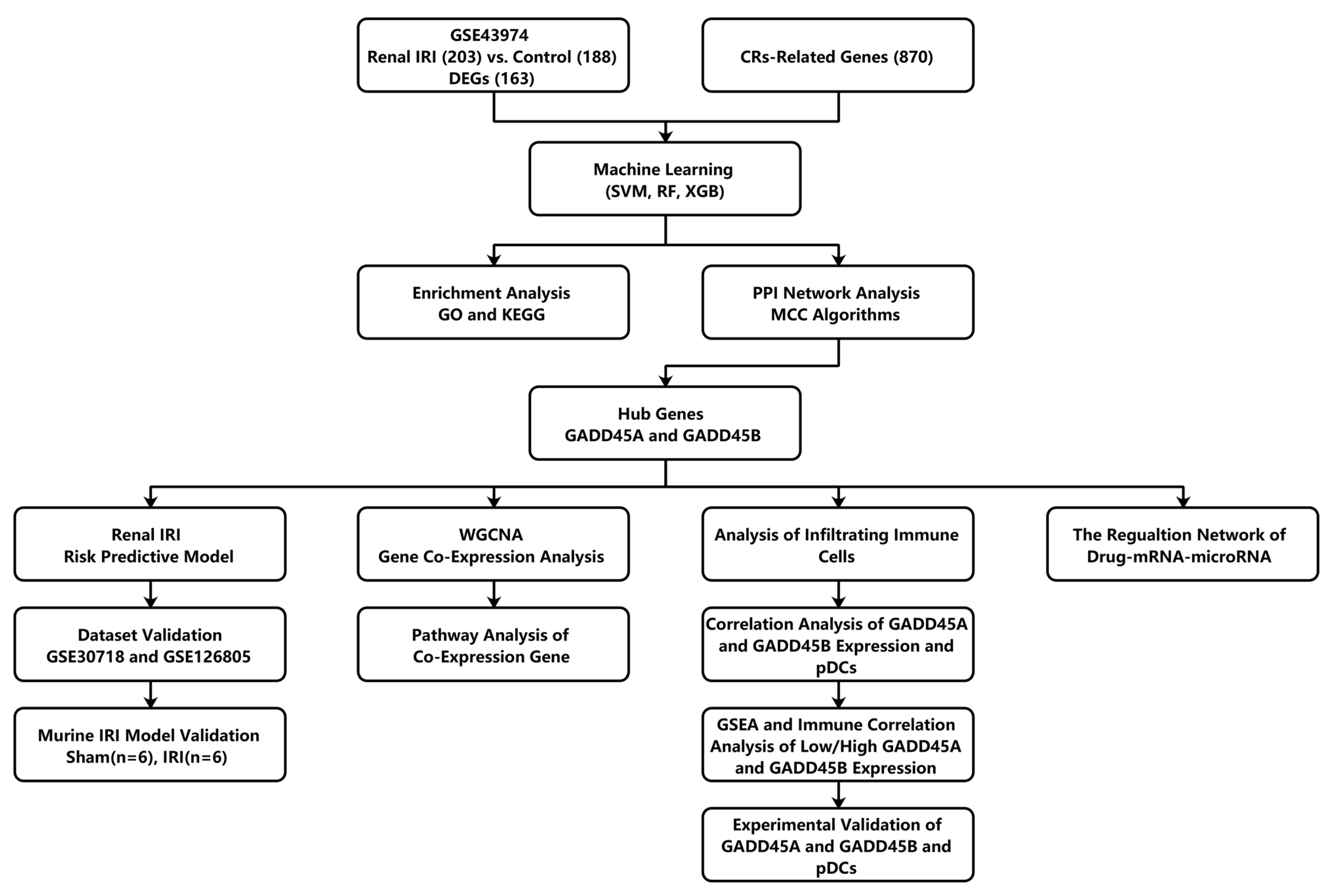
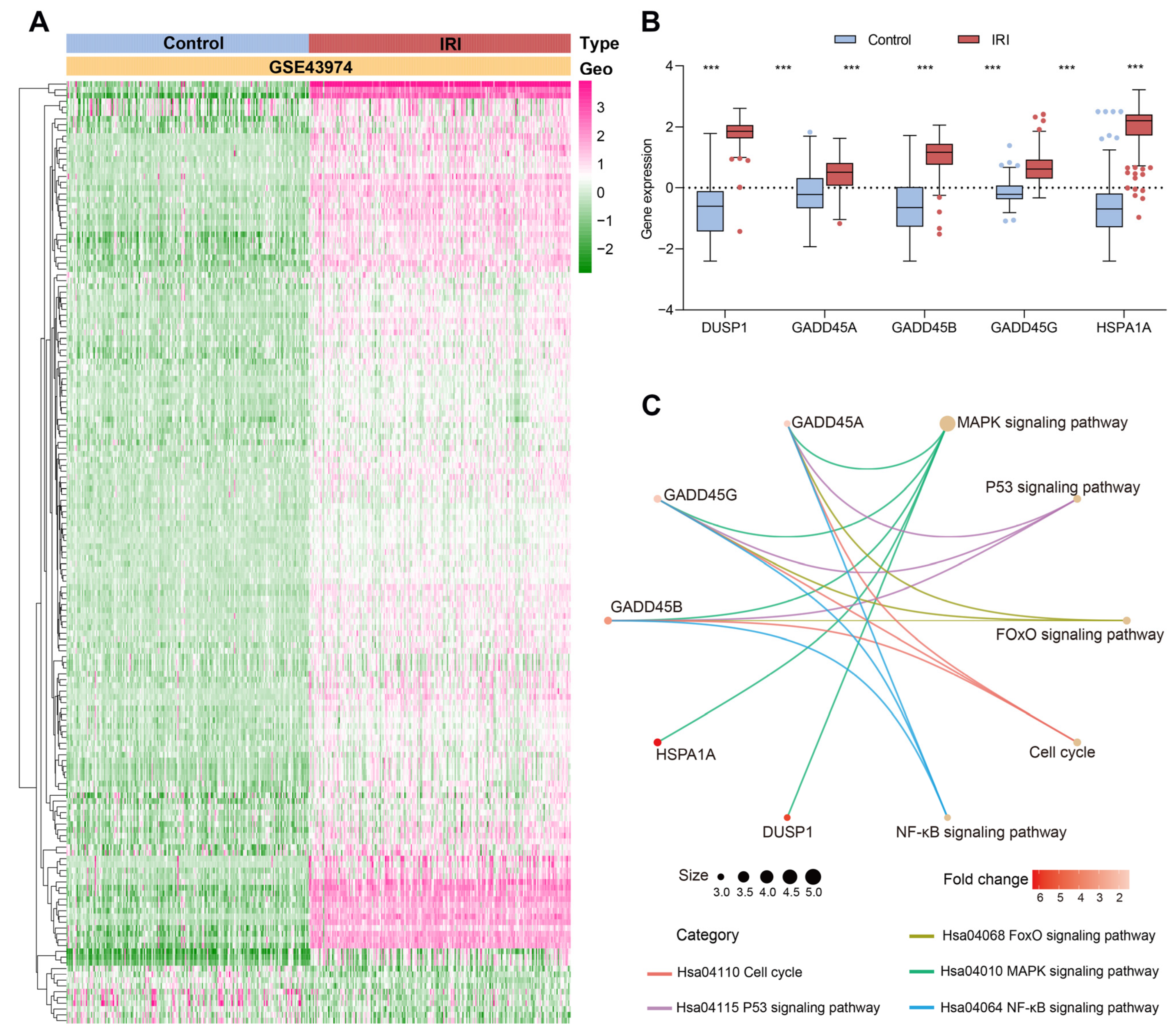
2.1. Identification and Enrichment Analysis of CRs-Associated DEGs in Renal Ischemia-Reperfusion Injury Using Three Machine Learning Models
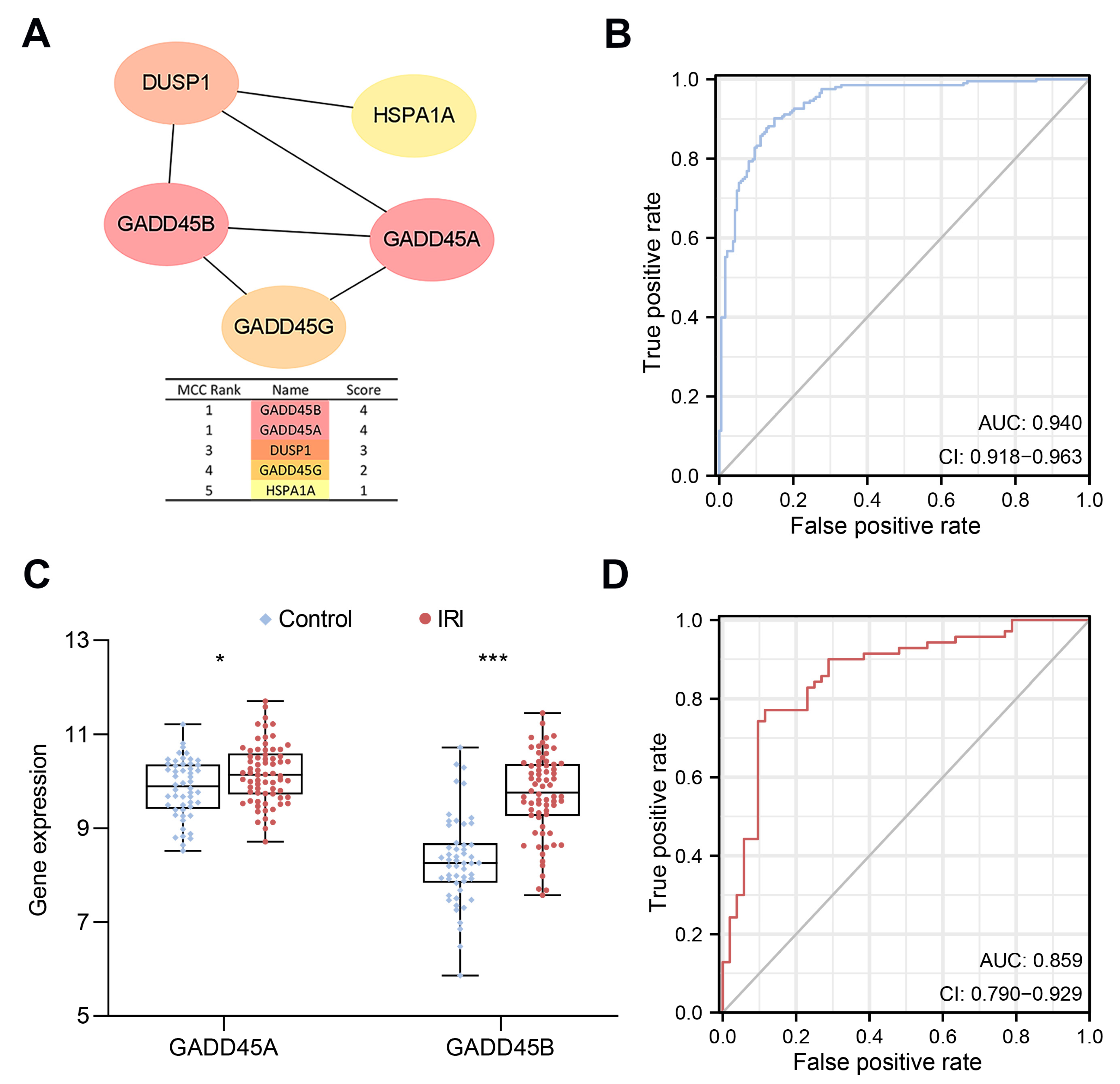
2.2. Construction of a CRs-Associated Predictive Nomogram and Validation
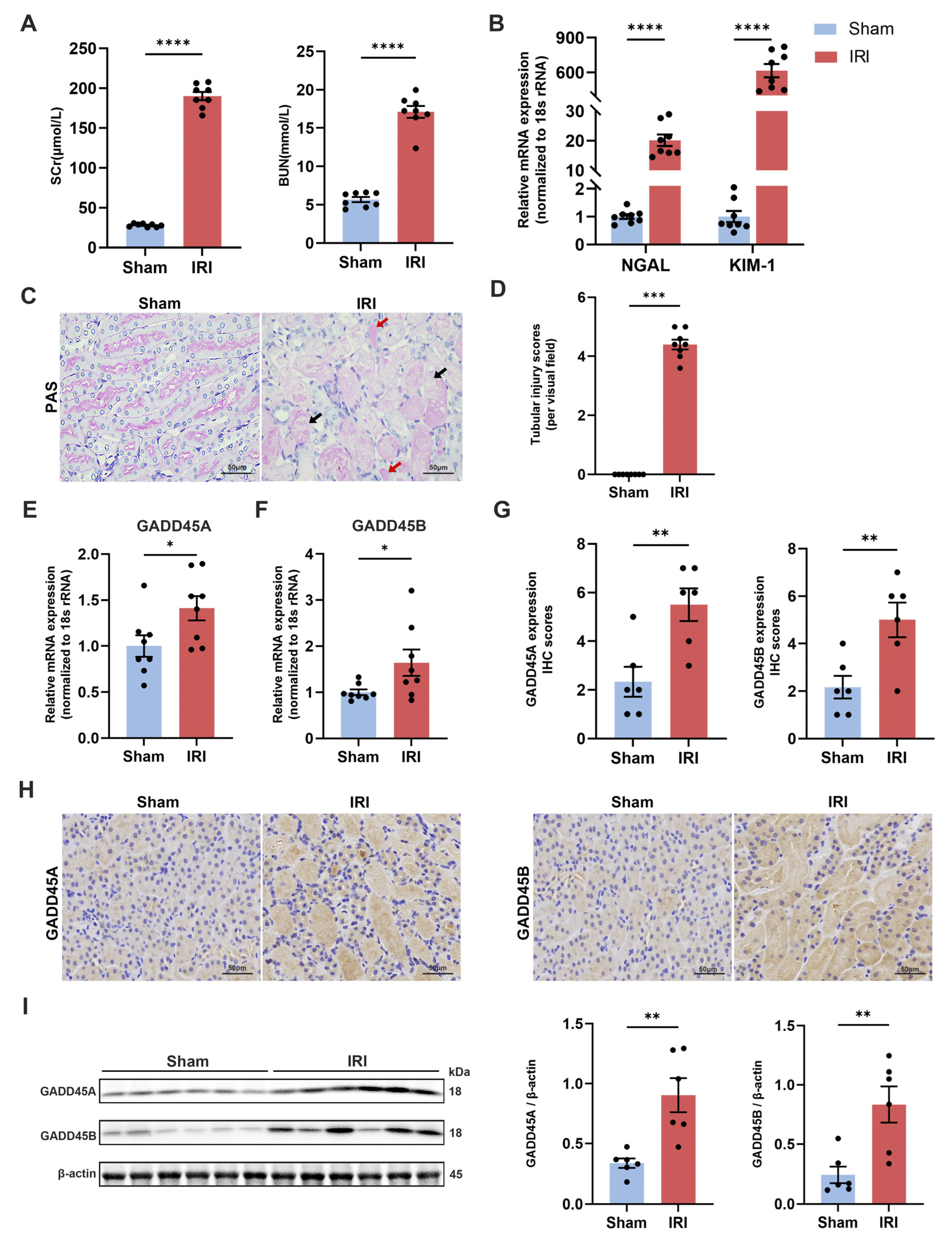
2.3. Validation of GADD45A and GADD45B Expression in Mice Renal IRI Model
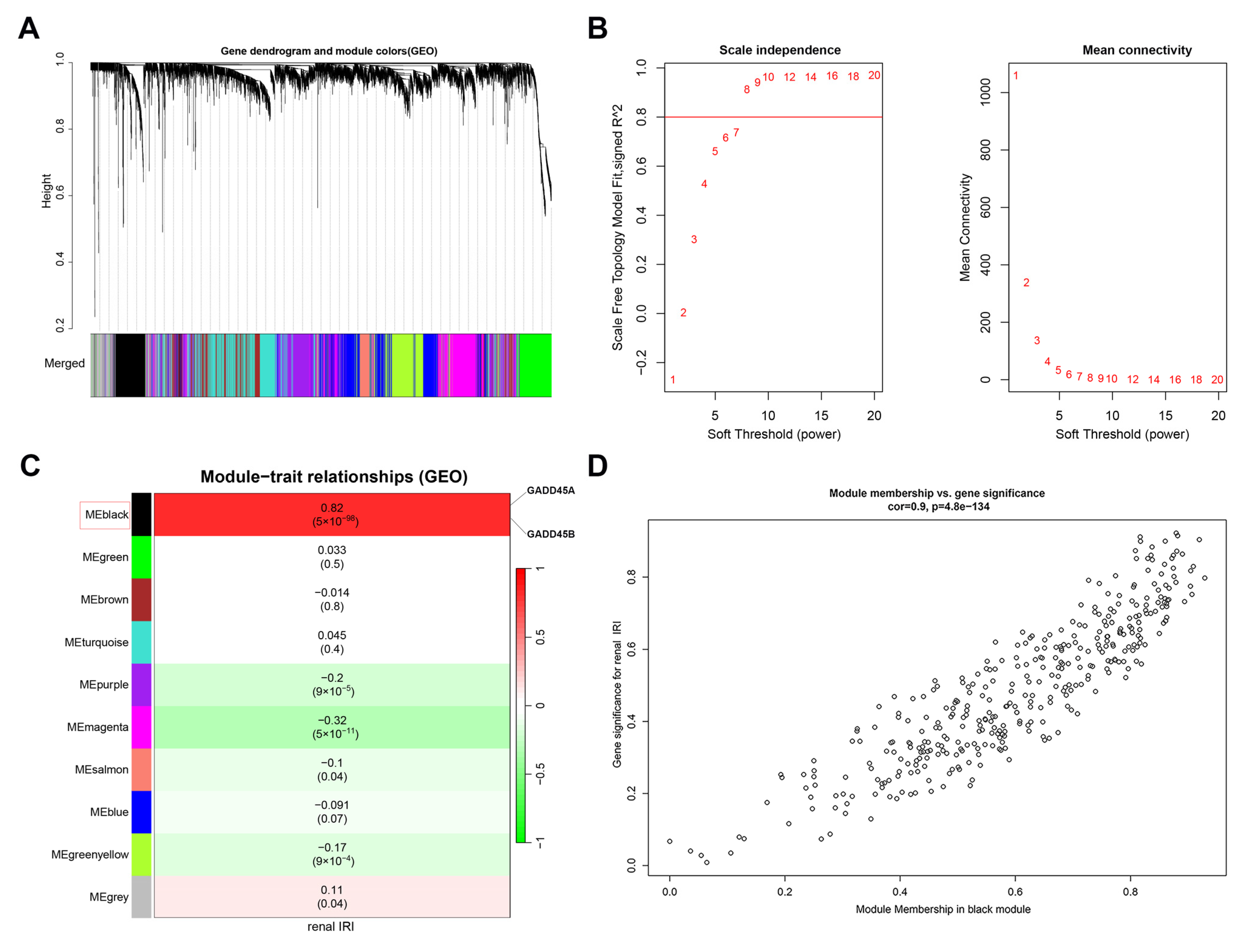
2.4. Identification of GADD45A and GADD45B Co-Expression Genes Network in Renal Ischemia-Reperfusion Injury Using WGCNA
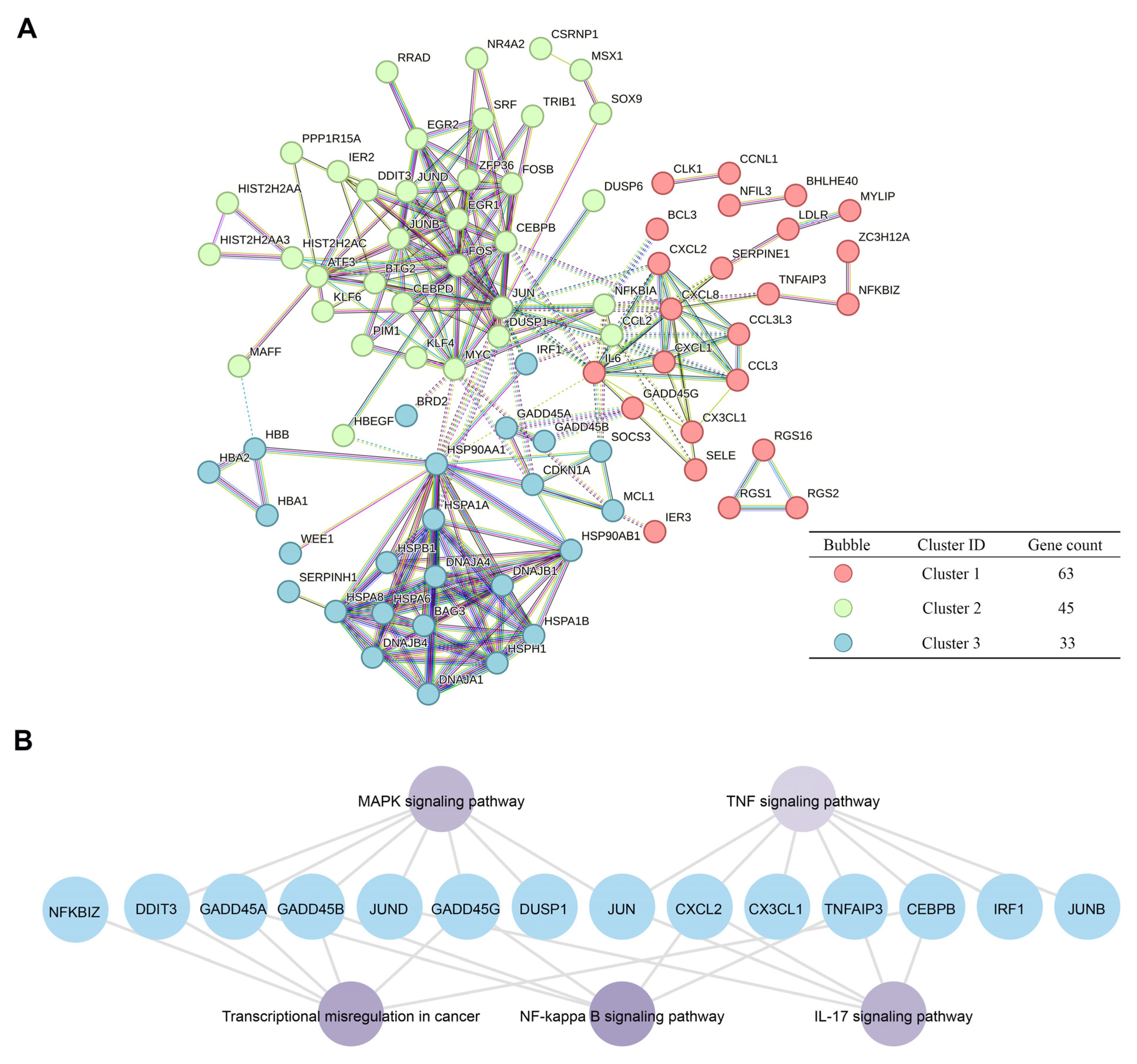
2.5. Pathway Analysis of GADD45A and GADD45B Co-Expression Genes Network in Renal Ischemia-Reperfusion Injury
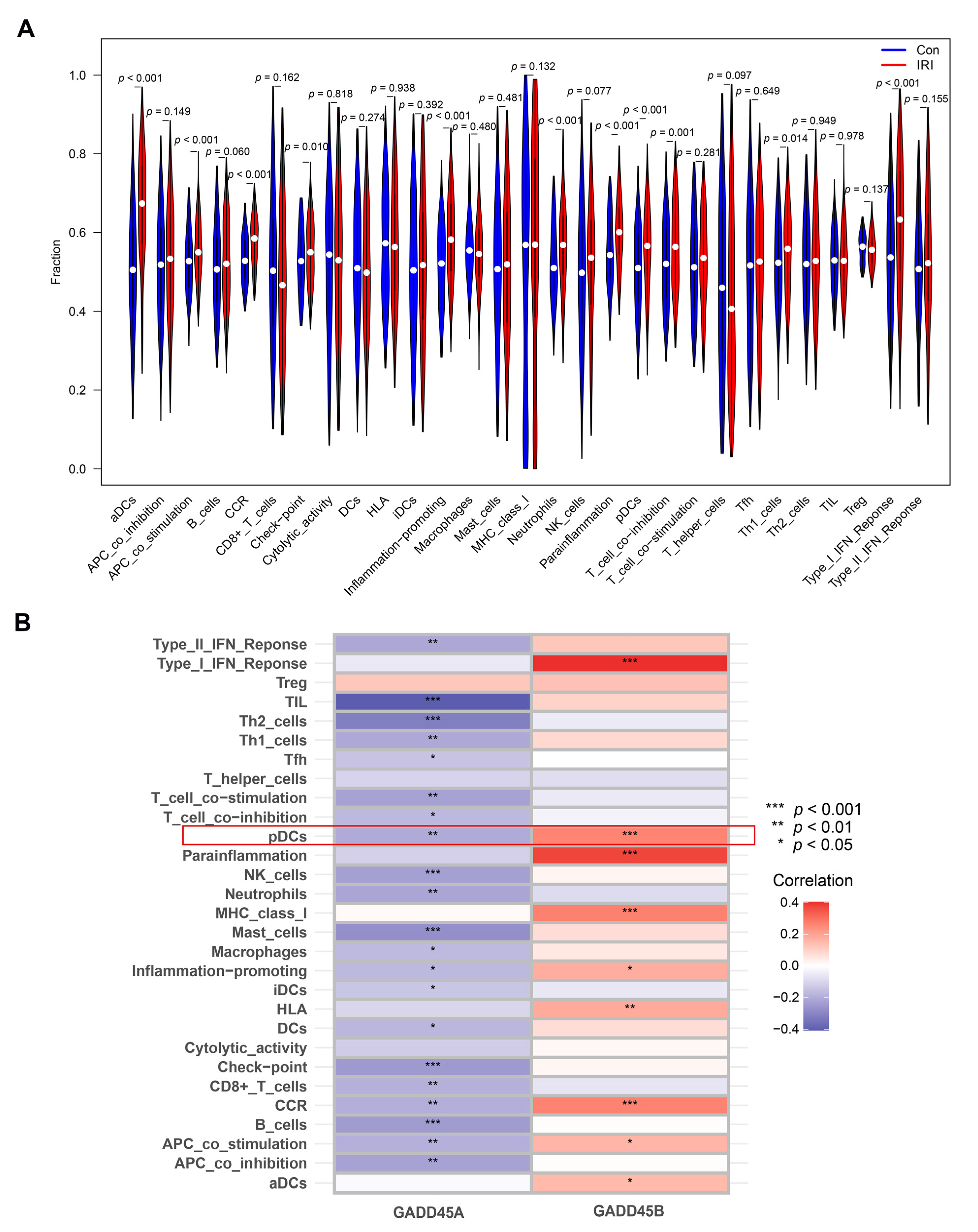
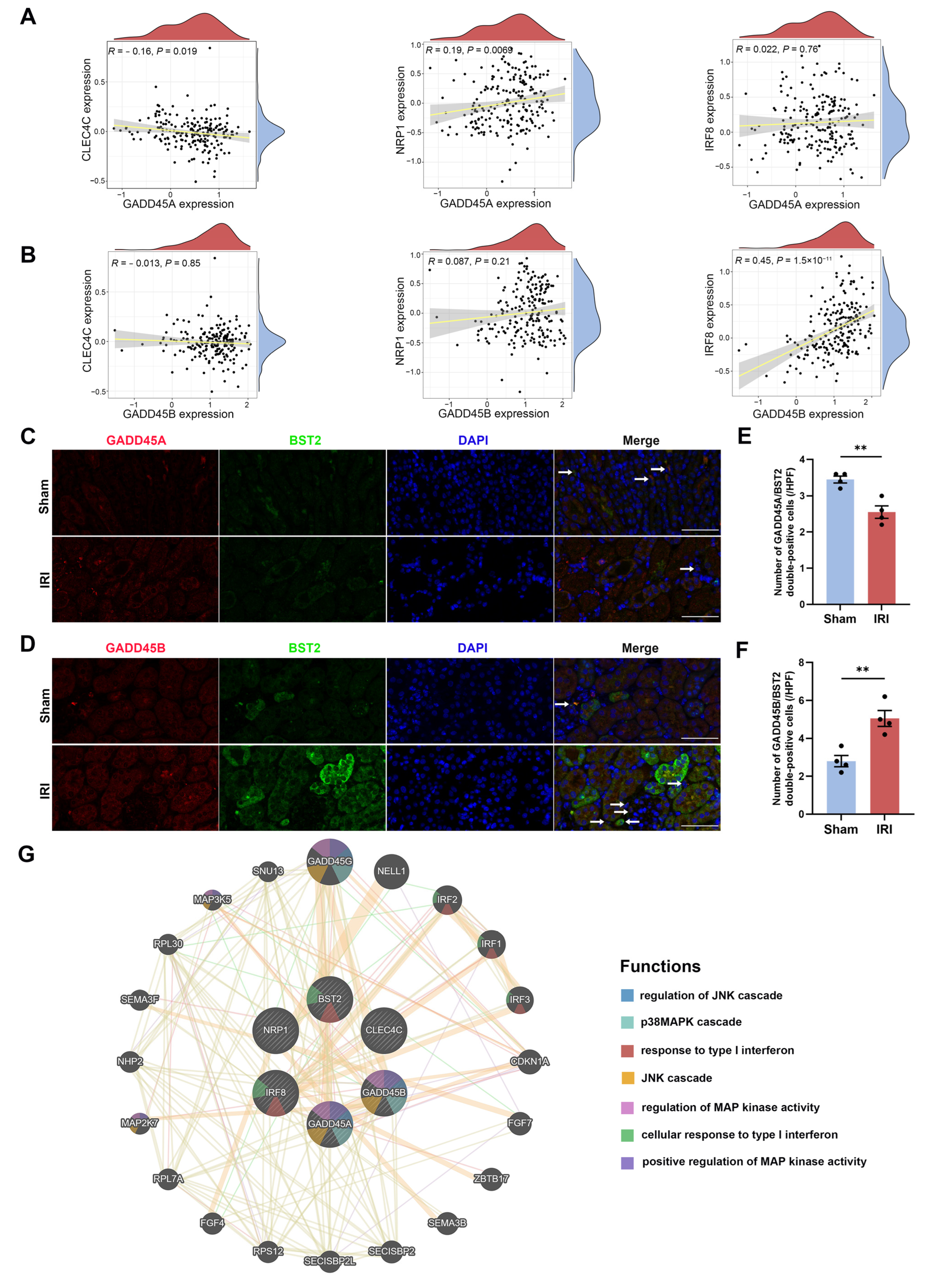
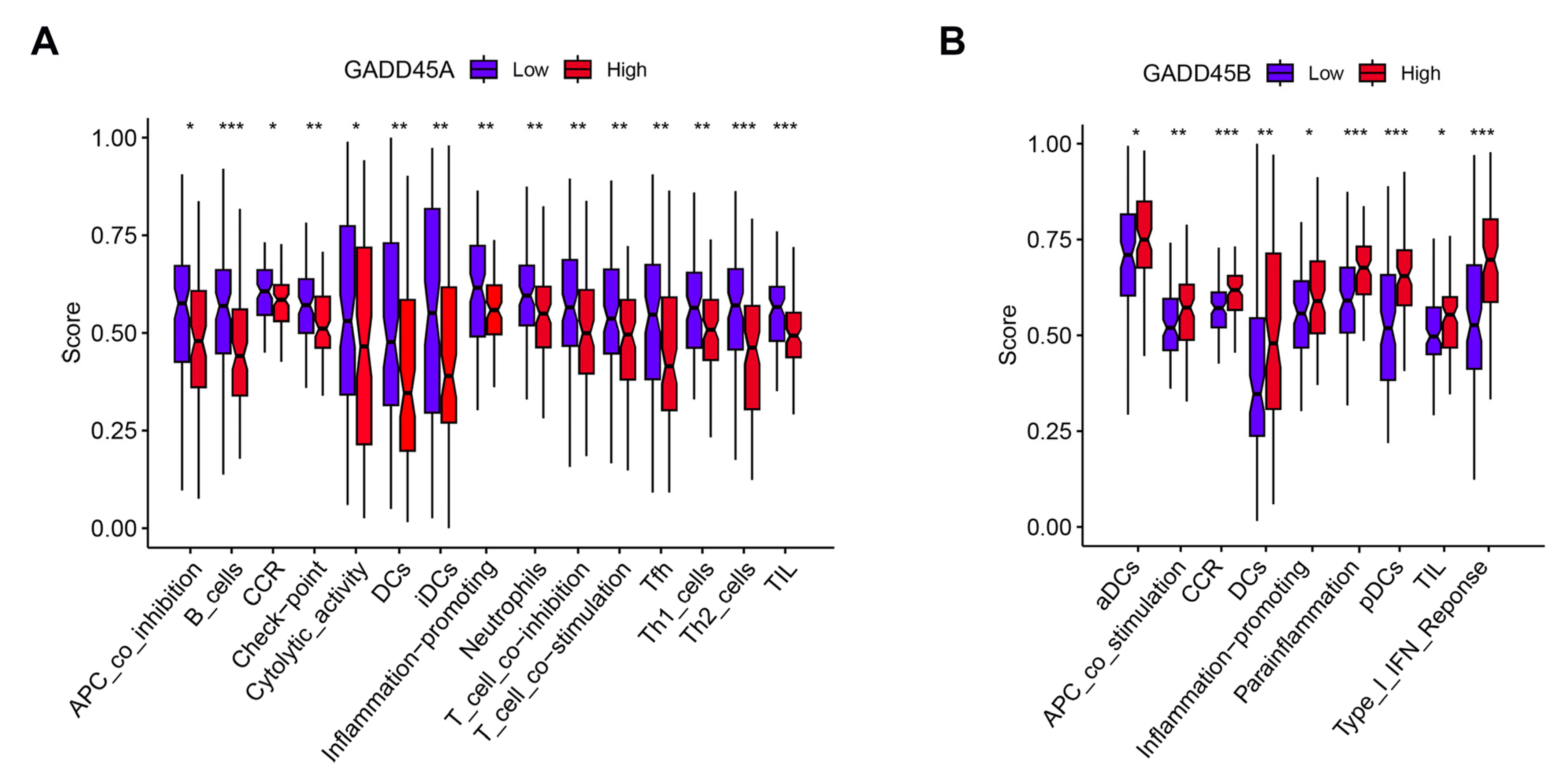
2.6. Association of GADD45A and GADD45B with Infiltrating Immune Cells and Functions in Renal IRI and Validation
2.7. GSEA and Immune Correlation Analysis of Low/High GADD45A and GADD45B Expression in Renal IRI
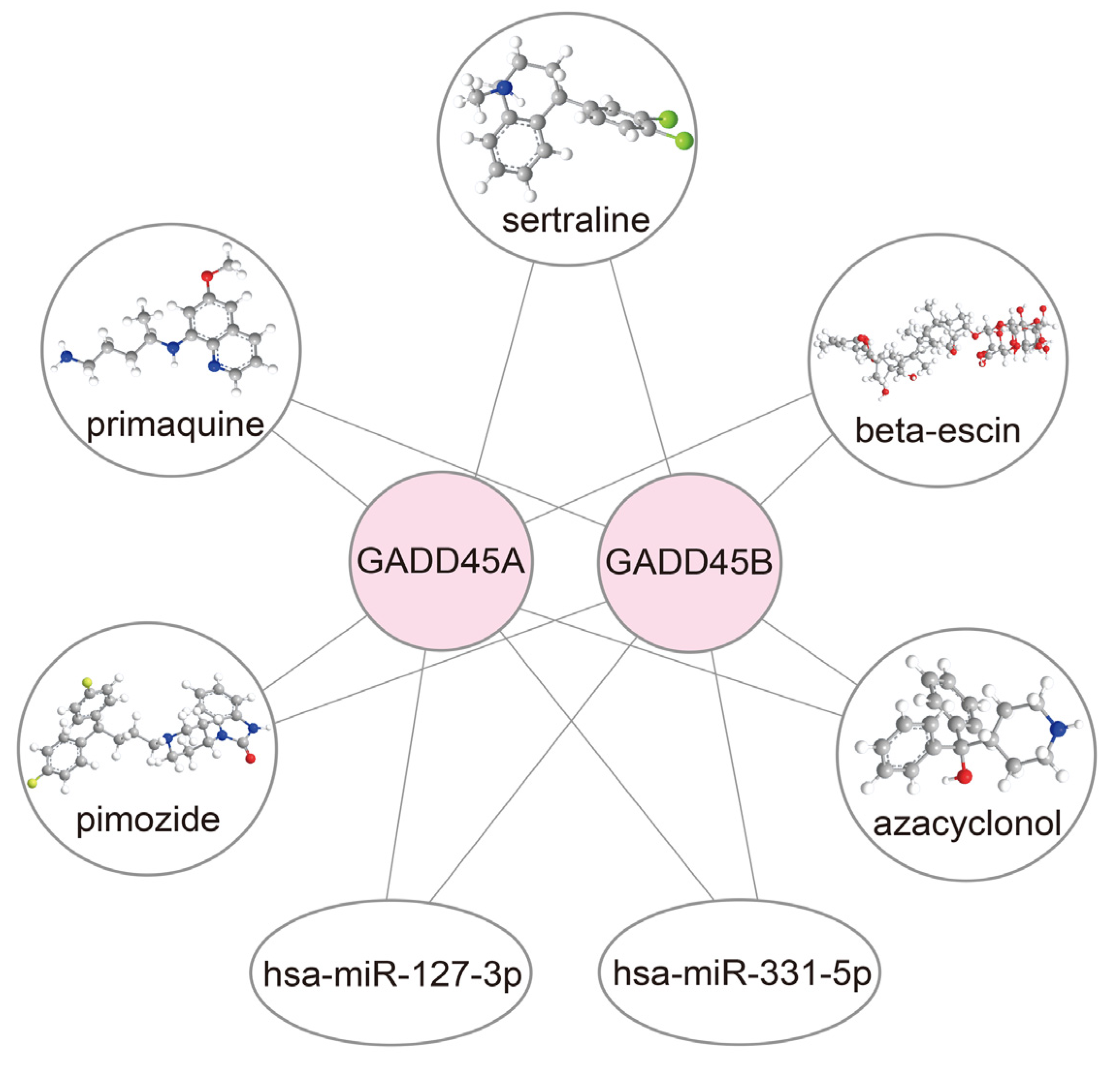
2.8. Construction of Drug-mRNA-miRNA Network
3. Discussion
4. Materials and Methods
4.1. Datasets Collection
4.2. Identification of Chromatin Regulators-Associated Differentially Expressed Genes
4.3. Identification CR-Related Characteristic Biomarkers Using Machine Learning Approach
4.4. Gene Ontology (GO), Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis and Gene Set Enrichment Analysis
4.5. Protein-Protein Interaction (PPI) Network Construction
4.6. Screening the Co-Expression Genes Using WGCNA Network Analysis
4.7. Analysis of Infiltrating Immune Cells
4.8. Evaluation of Relationship between Hub Genes and Cell Surface Markers of Plasmacytoid Dendritic Cells
4.9. Construction of a CRs-Associated Risk Predictive Model for Renal Ischemia-Reperfusion Injury
4.10. Validation of Bioinformatics Data by Murine Renal IRI Model
4.11. Validation of Hub Genes and Kidney Injury Biomarkers Expression Using Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.12. Renal Histopathology
4.13. Candidate Drugs and Microrna-Targeted Screening
4.14. Western Blot Analysis
4.15. Immunohistochemistry (IHC) and Immunofluorescence (IF) Staining
4.16. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eltzschig, H.K.; Eckle, T. Ischemia and reperfusion—From mechanism to translation. Nat. Med. 2011, 17, 1391–1401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellum, J.A.; Romagnani, P.; Ashuntantang, G.; Ronco, C.; Zarbock, A.; Anders, H.-J. Acute kidney injury. Nat. Rev. Dis. Prim. 2021, 7, 52. [Google Scholar] [CrossRef]
- Kellum, J.A.; Chawla, L.S.; Keener, C.; Singbartl, K.; Palevsky, P.M.; Pike, F.L.; Yealy, D.M.; Huang, D.T.; Angus, D.C.; ProCESS and ProGReSS-AKI Investigators. The Effects of Alternative Resuscitation Strategies on Acute Kidney Injury in Patients with Septic Shock. Am. J. Respir. Crit. Care Med. 2016, 193, 281–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loef, B.G.; Epema, A.H.; Smilde, T.D.; Henning, R.H.; Ebels, T.; Navis, G.; Stegeman, C.A. Immediate Postoperative Renal Function Deterioration in Cardiac Surgical Patients Predicts In-Hospital Mortality and Long-Term Survival. J. Am. Soc. Nephrol. 2005, 16, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Hobson, C.E.; Yavas, S.; Segal, M.S.; Schold, J.D.; Tribble, C.G.; Layon, A.J.; Bihorac, A. Acute Kidney Injury Is Associated with Increased Long-Term Mortality After Cardiothoracic Surgery. Circulation 2009, 119, 2444–2453. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Alam, A.; Soo, A.P.; George, A.J.T.; Ma, D. Ischemia-Reperfusion Injury Reduces Long Term Renal Graft Survival: Mechanism and Beyond. EBioMedicine 2018, 28, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pefanis, A.; Ierino, F.L.; Murphy, J.M.; Cowan, P.J. Regulated necrosis in kidney ischemia-reperfusion injury. Kidney Int. 2019, 96, 291–301. [Google Scholar] [CrossRef]
- Smith, S.F.; Hosgood, S.A.; Nicholson, M.L. Ischemia-reperfusion injury in renal transplantation: 3 key signaling pathways in tubular epithelial cells. Kidney Int. 2019, 95, 50–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, M.; Tang, C.; Ma, Z.; Huang, S.; Dong, Z. DNA damage response in nephrotoxic and ischemic kidney injury. Toxicol. Appl. Pharmacol. 2016, 313, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Livingston, M.J.; Wang, J.; Zhou, J.; Wu, G.; Ganley, I.G.; Hill, J.A.; Yin, X.-M.; Dong, Z. Clearance of damaged mitochondria via mitophagy is important to the protective effect of ischemic preconditioning in kidneys. Autophagy 2019, 15, 2142–2162. [Google Scholar] [CrossRef] [Green Version]
- Havasi, A.; Borkan, S.C. Apoptosis and acute kidney injury. Kidney Int. 2011, 80, 29–40. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Dong, G.; Liang, X.; Dong, Z. Epigenetic regulation in AKI and kidney repair: Mechanisms and therapeutic implications. Nat. Rev. Nephrol. 2019, 15, 220–239. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Xu, J.; Li, J.; Pan, T.; Bai, J.; Wang, L.; Jin, X.; Lin, X.; Zhang, Y.; Li, Y.; et al. FACER: Comprehensive molecular and functional characterization of epigenetic chromatin regulators. Nucleic Acids Res. 2018, 46, 10019–10033. [Google Scholar] [CrossRef]
- Wang, Y.; Wysocka, J.; Sayegh, J.; Lee, Y.-H.; Perlin, J.R.; Leonelli, L.; Sonbuchner, L.S.; McDonald, C.H.; Cook, R.G.; Dou, Y.; et al. Human PAD4 Regulates Histone Arginine Methylation Levels via Demethylimination. Science 2004, 306, 279–283. [Google Scholar] [CrossRef]
- Rabadi, M.; Kim, M.; D’Agati, V.; Lee, H.T. Peptidyl arginine deiminase-4-deficient mice are protected against kidney and liver injury after renal ischemia and reperfusion. Am. J. Physiol. Physiol. 2016, 311, F437–F449. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chen, Z.; Weng, X.; Chen, H.; Du, Y.; Diao, C.; Liu, X.; Wang, L. Enhancer of zeste homolog 2 modulates oxidative stress-mediated pyroptosis in vitro and in a mouse kidney ischemia-reperfusion injury model. FASEB J. 2019, 34, 835–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Wang, W.; Weng, X.; Chen, H.; Chen, Z.; Du, Y.; Liu, X.; Wang, L. The H3K9 histone methyltransferase G9a modulates renal ischemia reperfusion injury by targeting Sirt1. Free Radic. Biol. Med. 2021, 172, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Feng, W.; Su, X.; Luo, D.; Li, Z.; Zhou, Y.; Zhu, Y.; Zhang, M.; Chen, J.; Liu, B.; et al. SIRT6 protects vascular smooth muscle cells from osteogenic transdifferentiation via Runx2 in chronic kidney disease. J. Clin. Investig. 2022, 132, e150051. [Google Scholar] [CrossRef] [PubMed]
- Li, L.X.; Fan, L.X.; Zhou, J.X.; Grantham, J.J.; Calvet, J.P.; Sage, J.; Li, X. Lysine methyltransferase SMYD2 promotes cyst growth in autosomal dominant polycystic kidney disease. J. Clin. Investig. 2017, 127, 2751–2764. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Xu, C.; Wang, Y.; Meng, C.; Liu, W.; Zhao, Y.; Huang, X.-R.; You, W.; Feng, B.; Zheng, Z.-H.; et al. Lethal (3) malignant brain tumor-like 2 (L3MBTL2) protein protects against kidney injury by inhibiting the DNA damage–p53–apoptosis pathway in renal tubular cells. Kidney Int. 2018, 93, 855–870. [Google Scholar] [CrossRef]
- Jang, H.R.; Rabb, H. Immune cells in experimental acute kidney injury. Nat. Rev. Nephrol. 2014, 11, 88–101. [Google Scholar] [CrossRef]
- Rice, S.B.; Nenadic, G.; Stapley, B.J. Mining protein function from text using term-based support vector machines. BMC Bioinform. 2005, 6 (Suppl. S1), S22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liaw, A.; Wiener, M.C. Classification and Regression by RandomForest. R News 2002, 2, 18–22. [Google Scholar]
- Chen, T.; Guestrin, C. XGBoost: A Scalable Tree Boosting System. In Proceedings of the 22nd ACM SIGKDD International Conference on Knowledge Discovery and Data Mining, San Francisco, CA, USA, 13–17 August 2016; pp. 785–794. [Google Scholar]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. CytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene Set Knowledge Discovery with Enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minaga, K.; Watanabe, T.; Arai, Y.; Shiokawa, M.; Hara, A.; Yoshikawa, T.; Kamata, K.; Yamashita, K.; Kudo, M. Activation of interferon regulatory factor 7 in plasmacytoid dendritic cells promotes experimental autoimmune pancreatitis. J. Gastroenterol. 2020, 55, 565–576. [Google Scholar] [CrossRef]
- Kitamura, H.; Tanigawa, T.; Kuzumoto, T.; Nadatani, Y.; Otani, K.; Fukunaga, S.; Hosomi, S.; Tanaka, F.; Kamata, N.; Nagami, Y.; et al. Interferon-α exerts proinflammatory properties in experimental radiation-induced esophagitis: Possible involvement of plasmacytoid dendritic cells. Life Sci. 2022, 289, 120215. [Google Scholar] [CrossRef]
- Watanabe, T.; Yamashita, K.; Arai, Y.; Minaga, K.; Kamata, K.; Nagai, T.; Komeda, Y.; Takenaka, M.; Hagiwara, S.; Ida, H.; et al. Chronic Fibro-Inflammatory Responses in Autoimmune Pancreatitis Depend on IFN-α and IL-33 Produced by Plasmacytoid Dendritic Cells. J. Immunol. 2017, 198, 3886–3896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warde-Farley, D.; Donaldson, S.L.; Comes, O.; Zuberi, K.; Badrawi, R.; Chao, P.; Franz, M.; Grouios, C.; Kazi, F.; Lopes, C.T.; et al. The GeneMANIA prediction server: Biological network integration for gene prioritization and predicting gene function. Nucleic Acids Res. 2010, 38, W214–W220. [Google Scholar] [CrossRef] [Green Version]
- Yoo, M.; Shin, J.; Kim, J.; Ryall, K.A.; Lee, K.; Lee, S.; Jeon, M.; Kang, J.; Tan, A.C. DSigDB: Drug signatures database for gene set analysis. Bioinformatics 2015, 31, 3069–3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agarwal, V.; Bell, G.W.; Nam, J.-W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Wu, Q.; Tian, R.; Liu, J.; Ou, C.; Li, Y.; Fu, X. Deciphering comprehensive features of tumor microenvironment controlled by chromatin regulators to predict prognosis and guide therapies in uterine corpus endometrial carcinoma. Front. Immunol. 2023, 14, 1139126. [Google Scholar] [CrossRef]
- Niu, X.; Cheuk, Y.C.; Li, X.; Rong, R.; Xu, X.; Xu, C.; Luo, Y.; Zhang, P.; Guo, J. Identification of the Subtypes of Renal Ischemia-Reperfusion Injury Based on Pyroptosis-Related Genes. Biomolecules 2023, 13, 275. [Google Scholar] [CrossRef]
- Wang, Z.; He, Z.; Xuan, Q.; Zhang, Y.; Xu, J.; Lin, J.; Li, H.; Chen, W.; Jiang, T. Analysis of the potential ferroptosis mechanism and multitemporal expression change of central ferroptosis-related genes in cardiac ischemia–reperfusion injury. Front. Physiol. 2022, 13, 934901. [Google Scholar] [CrossRef]
- Sun, S.; Xue, J.; Guo, Y.; Li, J. Bioinformatics analysis of genes related to ferroptosis in hepatic ischemia-reperfusion injury. Front. Genet. 2022, 13, 1072544. [Google Scholar] [CrossRef]
- Zhu, L.; Lian, W.; Yao, Z.; Yang, X.; Wang, Z.; Lai, Y.; Xu, S.; Zhao, B.; Liu, K. Integrated Analysis of Ferroptosis and Immunity-Related Genes Associated with Intestinal Ischemia/Reperfusion Injury. J. Inflamm. Res. 2022, 15, 2397–2411. [Google Scholar] [CrossRef]
- He, S.; He, L.; Yan, F.; Li, J.; Liao, X.; Ling, M.; Jing, R.; Pan, L. Identification of hub genes associated with acute kidney injury induced by renal ischemia–reperfusion injury in mice. Front. Physiol. 2022, 13, 951855. [Google Scholar] [CrossRef] [PubMed]
- You, R.; Heyang, Z.; Ma, Y.; Xia, P.; Zheng, H.; Lin, J.; Ji, P.; Chen, L. Identification of biomarkers, immune infiltration landscape, and treatment targets of ischemia–reperfusion acute kidney injury at an early stage by bioinformatics methods. Hereditas 2022, 159, 24. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.-T.; Lv, L.-L.; Pan, M.-M.; Wen, Y.; Wang, B.; Li, Z.-L.; Wu, M.; Wang, F.-M.; Crowley, S.D.; Liu, B.-C. Hydroxychloroquine attenuates renal ischemia/reperfusion injury by inhibiting cathepsin mediated NLRP3 inflammasome activation. Cell Death Dis. 2018, 9, 351. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Gusella, G.L.; He, J.C. Epithelial proliferation and cell cycle dysregulation in kidney injury and disease. Kidney Int. 2021, 100, 67–78. [Google Scholar] [CrossRef]
- Tang, C.; Ma, Z.; Zhu, J.; Liu, Z.; Liu, Y.; Liu, Y.; Cai, J.; Dong, Z. P53 in kidney injury and repair: Mechanism and therapeutic potentials. Pharmacol. Ther. 2019, 195, 5–12. [Google Scholar] [CrossRef]
- Peng, J.; Li, X.; Zhang, D.; Chen, J.-K.; Su, Y.; Smith, S.B.; Dong, Z. Hyperglycemia, p53, and mitochondrial pathway of apoptosis are involved in the susceptibility of diabetic models to ischemic acute kidney injury. Kidney Int. 2015, 87, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Humayun, A.; Fornace, A.J. GADD45 in Stress Signaling, Cell Cycle Control, and Apoptosis. In Gadd45 Stress Sensor Genes; Springer International Publishing: Cham, Switzerland, 2022; pp. 1–22. [Google Scholar] [CrossRef]
- Zhang, W.; Bae, I.; Krishnaraju, K.; Azam, N.; Fan, W.; Smith, K.; Hoffman, B.; A Liebermann, D. CR6: A third member in the MyD118 and Gadd45 gene family which functions in negative growth control. Oncogene 1999, 18, 4899–4907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, K.; Murray, M.G.; Whelan, K.A. Roles for GADD45 in Development and Cancer. In Gadd45 Stress Sensor Genes; Springer International Publishing: Cham, Switzerland, 2022; pp. 23–39. [Google Scholar] [CrossRef]
- Zaidi, M.R.; Liebermann, D.A. Gadd45 in Senescence. 2022, 1360, 109–116. Adv. Exp. Med. Biol. 2022, 1360, 109–116. [Google Scholar] [CrossRef]
- Aslan, G.; Atessahin, A.; Sahna, E. The inhibition of apoptosis through myocardial postconditioning by affecting Fas/FasIg signaling through miR139-3p and miR181a-1. J. Card. Surg. 2019, 35, 564–570. [Google Scholar] [CrossRef]
- Gu, Q.-L.; Jiang, P.; Ruan, H.-F.; Tang, H.; Liang, Y.-B.; Ma, Z.-F.; Zhan, H. The expression of oxidative stress genes related to myocardial ischemia reperfusion injury in patients with ST-elevation myocardial infarction. World J. Emerg. Med. 2022, 13, 106. [Google Scholar] [CrossRef]
- Liu, C.; Liu, H.; Sun, Q.; Zhang, P. MicroRNA 1283 alleviates cardiomyocyte damage caused by hypoxia /reoxygenation via targeting GADD45A and inactivating the JNK and p38 MAPK signaling pathways. Kardiol. Pol. 2021, 79, 147–155. [Google Scholar] [CrossRef]
- Wang, L.; Zhou, C.; Wang, Z.; Liu, J.; Jing, Z.; Zhang, Z.; Wang, Y. Dynamic variation of genes profiles and pathways in the hippocampus of ischemic mice: A genomic study. Brain Res. 2011, 1372, 13–21. [Google Scholar] [CrossRef]
- He, G.-Q.; Xu, W.-M.; Liao, H.-J.; Jiang, C.; Li, C.-Q.; Zhang, W. Silencing Huwe1 Reduces Apoptosis of Cortical Neurons Exposed to Oxygen-Glucose Deprivation and Reperfusion. Neural Regen. Res. 2019, 14, 1977. [Google Scholar] [CrossRef] [PubMed]
- Fallsehr, C.; Zapletal, C.; Kremer, M.; Demir, R.; Doeberitz, M.V.K.; Klar, E. Identification of differentially expressed genes after partial rat liver ischemia/reperfusion by suppression subtractive hybridization. World J. Gastroenterol. 2005, 11, 1303–1316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Ming, Y.; Cheng, K.; Niu, Y.; Ye, Q. Gene Expression Profiling in Ischemic Postconditioning to Alleviate Mouse Liver Ischemia/Reperfusion Injury. Int. J. Med. Sci. 2019, 16, 343–354. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhu, N.; Jia, J.; Gu, L.; Du, Y.; Tang, G.; Wang, X.; Yang, M.; Yuan, W. Trimethylamine N-oxide mediated Y-box binding protein-1 nuclear translocation promotes cell cycle progression by directly downregulating Gadd45a expression in a cellular model of chronic kidney disease. Life Sci. 2021, 271, 119173. [Google Scholar] [CrossRef]
- Venancio, V.P.; Almeida, M.R.; Antunes, L.M.G. Cocoplum (Chrysobalanus icaco L.) decreases doxorubicin-induced DNA damage and downregulates Gadd45a, Il-1β and Tnf-α in vivo. Food Res. Int. 2017, 105, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Wang, Z.; Xu, Q.; Zhang, Y.; Zhai, Y.; Bai, J.; Liu, M.; Hui, Z.; Xu, N. Inhibition of STAT1 sensitizes renal cell carcinoma cells to radiotherapy and chemotherapy. Cancer Biol. Ther. 2012, 13, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, T.A.; Biazi, B.I.; Coatti, G.C.; Baranoski, A.; Marques, L.A.; Corveloni, A.C.; Mantovani, M.S. Dimethoxycurcumin reduces proliferation and induces apoptosis in renal tumor cells more efficiently than demethoxycurcumin and curcumin. Chem. -Biol. Interact. 2021, 338, 109410. [Google Scholar] [CrossRef]
- Liu, B.; Zhang, Y.-H.; Jiang, Y.; Li, L.-L.; Chen, Q.; He, G.-Q.; Tan, X.-D.; Li, C.-Q. Gadd45b is a Novel Mediator of Neuronal Apoptosis in Ischemic Stroke. Int. J. Biol. Sci. 2015, 11, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Li, L.-L.; Tan, X.-D.; Zhang, Y.-H.; Jiang, Y.; He, G.-Q.; Chen, Q.; Li, C.-Q. Gadd45b Mediates Axonal Plasticity and Subsequent Functional Recovery After Experimental Stroke in Rats. Mol. Neurobiol. 2014, 52, 1245–1256. [Google Scholar] [CrossRef]
- Cho, C.H.; Byun, H.-R.; Jover-Mengual, T.; Pontarelli, F.; DeJesus, C.; Cho, A.-R.; Zukin, R.S.; Hwang, J.-Y. Gadd45b Acts as Neuroprotective Effector in Global Ischemia-Induced Neuronal Death. Int. Neurourol. J. 2019, 23, S11–S21. [Google Scholar] [CrossRef] [Green Version]
- Yin, Q.; Du, T.; Yang, C.; Li, X.; Zhao, Z.; Liu, R.; Yang, B.; Liu, B. Gadd45b is a novel mediator of depression-like behaviors and neuroinflammation after cerebral ischemia. Biochem. Biophys. Res. Commun. 2021, 554, 107–113. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, Q.; Deng, J.; Li, J.; Li, J.; Wen, L.; Ma, J.; Li, C. ALK5 signaling pathway mediates neurogenesis and functional recovery after cerebral ischemia/reperfusion in rats via Gadd45b. Cell Death Dis. 2019, 10, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, G.; Xu, W.; Tong, L.; Li, S.; Su, S.; Tan, X.; Li, C. Gadd45b prevents autophagy and apoptosis against rat cerebral neuron oxygen-glucose deprivation/reperfusion injury. Apoptosis 2016, 21, 390–403. [Google Scholar] [CrossRef]
- Sheng, M.; Huang, Z.; Pan, L.; Yu, M.; Yi, C.; Teng, L.; He, L.; Gu, C.; Xu, C.; Li, J. SOCS2 exacerbates myocardial injury induced by ischemia/reperfusion in diabetic mice and H9c2 cells through inhibiting the JAK-STAT-IGF-1 pathway. Life Sci. 2017, 188, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Tang, R.; Meng, T.; Lin, W.; Shen, C.; Ooi, J.D.; Eggenhuizen, P.J.; Jin, P.; Ding, X.; Chen, J.; Tang, Y.; et al. A Partial Picture of the Single-Cell Transcriptomics of Human IgA Nephropathy. Front. Immunol. 2021, 12, 645988. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Sun, H.; Xu, R.; Wang, Y.; Guo, J.; Li, X.; Cheng, Y.; Xu, C.; Tang, C.; Sun, B.; et al. GADD45B Promotes Glucose-Induced Renal Tubular Epithelial-Mesenchymal Transition and Apoptosis via the p38 MAPK and JNK Signaling Pathways. Front. Physiol. 2020, 11, 1074. [Google Scholar] [CrossRef]
- Scott-Browne, J.; Shih, H. Stability and change in epigenetic regulation of immune cells. Immunol. Rev. 2022, 305, 5–8. [Google Scholar] [CrossRef]
- Swiecki, M.; Colonna, M. The multifaceted biology of plasmacytoid dendritic cells. Nat. Rev. Immunol. 2015, 15, 471–485. [Google Scholar] [CrossRef]
- Deng, B.; Lin, Y.; Chen, Y.; Ma, S.; Cai, Q.; Wang, W.; Li, B.; Liu, T.; Zhou, P.; He, R.; et al. Plasmacytoid dendritic cells promote acute kidney injury by producing interferon-α. Cell. Mol. Immunol. 2020, 18, 219–229. [Google Scholar] [CrossRef]
- Castellaneta, A.; Yoshida, O.; Kimura, S.; Yokota, S.; Geller, D.A.; Murase, N.; Thomson, A.W. Plasmacytoid dendritic cell-derived IFN-α promotes murine liver ischemia/reperfusion injury by induction of hepatocyte IRF-1. Hepatology 2014, 60, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Lai, L.; Zhang, A.; Yang, B.; Charles, E.J.; Kron, I.L.; Yang, Z. Plasmacytoid Dendritic Cells Mediate Myocardial Ischemia/Reperfusion Injury by Secreting Type I Interferons. J. Am. Heart Assoc. 2021, 10, e020754. [Google Scholar] [CrossRef]
- Salerno, D.M.; Tront, J.S.; Hoffman, B.; Liebermann, D.A. Gadd45a and Gadd45b modulate innate immune functions of granulocytes and macrophages by differential regulation of p38 and JNK signaling. J. Cell. Physiol. 2012, 227, 3613–3620. [Google Scholar] [CrossRef] [PubMed]
- Zazzeroni, F.; Papa, S.; Algeciras-Schimnich, A.; Alvarez, K.; Melis, T.; Bubici, C.; Majewski, N.; Hay, N.; De Smaele, E.; Peter, M.E.; et al. Gadd45β mediates the protective effects of CD40 costimulation against Fas-induced apoptosis. Blood 2003, 102, 3270–3279. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Wang, L.; Zhang, Y.; Jiang, W.; Sheng, H.; Cao, Q.; Wu, J.; Shen, B.; Shen, T.; Zhang, J.Z.; et al. Role of GADD45β in the regulation of synovial fluid T cell apoptosis in rheumatoid arthritis. Clin. Immunol. 2008, 128, 238–247. [Google Scholar] [CrossRef]
- Osawa, Y.; Iho, S.; Takauji, R.; Takatsuka, H.; Yamamoto, S.; Takahashi, T.; Horiguchi, S.; Urasaki, Y.; Matsuki, T.; Fujieda, S. Collaborative Action of NF-κB and p38 MAPK Is Involved in CpG DNA-Induced IFN-α and Chemokine Production in Human Plasmacytoid Dendritic Cells. J. Immunol. 2006, 177, 4841–4852. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Ferrandino, A.F.; A Flavell, R. Gadd45β is important for perpetuating cognate and inflammatory signals in T cells. Nat. Immunol. 2003, 5, 38–44. [Google Scholar] [CrossRef]
- Salvador, J.M.; Mittelstadt, P.R.; Belova, G.I.; Fornace, A.J.; Ashwell, J.D. The autoimmune suppressor Gadd45α inhibits the T cell alternative p38 activation pathway. Nat. Immunol. 2005, 6, 396–402. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, G.P.; Janakiraman, U.; Essa, M.M.; Thenmozhi, A.J.; Manivasagam, T. Escin attenuates behavioral impairments, oxidative stress and inflammation in a chronic MPTP/probenecid mouse model of Parkinson’s disease. Brain Res. 2014, 1585, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Mariani, N.; Everson, J.; Pariante, C.M.; Borsini, A. Modulation of microglial activation by antidepressants. J. Psychopharmacol. 2022, 36, 131–150. [Google Scholar] [CrossRef]
- Zanucoli, F.; Thomé, R.; Bonfanti, A.P.; de Carvalho, A.C.; Issayama, L.K.; da Costa, T.A.; Di Gangi, R.; Ferreira, I.T.; Bombeiro, A.L.; de Oliveira, A.L.R.; et al. Primaquine Treatment Suppresses Experimental Autoimmune Encephalomyelitis Severity. CNS Neurosci. Ther. 2014, 20, 1061–1064. [Google Scholar] [CrossRef]
- Mothi, M.; Sampson, S. Pimozide for schizophrenia or related psychoses. Cochrane Database Syst. Rev. 2013. [Google Scholar] [CrossRef]
- Hu, B.; Li, Y.; Gao, L.; Guo, Y.; Zhang, Y.; Chai, X.; Xu, M.; Yan, J.; Lu, P.; Ren, S.; et al. Hepatic Induction of Fatty Acid Binding Protein 4 Plays a Pathogenic Role in Sepsis in Mice. Am. J. Pathol. 2017, 187, 1059–1067. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemere, F. Failure of Azacyclonol (Frenquel) to Relieve Non-Experimental Mental Confusion and Hallucinations. Am. J. Psychiatry 1957, 113, 840. [Google Scholar] [CrossRef]
- Conde, E.; Giménez-Moyano, S.; Martín-Gómez, L.; Rodríguez, M.; Ramos, M.E.; Aguado-Fraile, E.; Blanco-Sanchez, I.; Saiz, A.; García-Bermejo, M.L. HIF-1α induction during reperfusion avoids maladaptive repair after renal ischemia/reperfusion involving miR127-3p. Sci. Rep. 2017, 7, 41099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguado-Fraile, E.; Ramos, E.; Conde-Moreno, E.; Rodríguez, M.; Martín-Gómez, L.; Lietor, A.; Candela, Á.; Ponte, B.; Liaño, F.; García-Bermejo, M.L. A Pilot Study Identifying a Set of microRNAs As Precise Diagnostic Biomarkers of Acute Kidney Injury. PLoS ONE 2015, 10, e0127175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.-S.; Liu, M.-F.; Ji, X.-Y.; Jiang, C.-R.; Li, Z.-L.; OuYang, B. Gastrodin combined with rhynchophylline inhibits cerebral ischaemia-induced inflammasome activation via upregulating miR-21–5p and miR-331–5p. Life Sci. 2019, 239, 116935. [Google Scholar] [CrossRef]
- Winkler, I.; Heisinger, T.; Hammerl, M.; Huber, E.; Urbanek, M.; Kiechl-Kohlendorfer, U.; Griesmaier, E.; Posod, A. MicroRNA Expression Profiles as Diagnostic and Prognostic Biomarkers of Perinatal Asphyxia and Hypoxic-Ischaemic Encephalopathy. Neonatology 2022, 119, 204–213. [Google Scholar] [CrossRef]
- Chen, B.; Wang, M.; Huang, R.; Liao, K.; Wang, T.; Yang, R.; Zhang, W.; Shi, Z.; Ren, L.; Lv, Q.; et al. Circular RNA circLGMN facilitates glioblastoma progression by targeting miR-127-3p/LGMN axis. Cancer Lett. 2021, 522, 225–237. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Wang, D.; Yu, Y.; Zhao, T.; Min, N.; Wu, Y.; Kang, L.; Zhao, Y.; Du, L.; Zhang, M.; et al. Legumain promotes tubular ferroptosis by facilitating chaperone-mediated autophagy of GPX4 in AKI. Cell Death Dis. 2021, 12, 65. [Google Scholar] [CrossRef]
- Hu, X.; Ding, C.; Ding, X.; Fan, P.; Zheng, J.; Xiang, H.; Li, X.; Qiao, Y.; Xue, W.; Li, Y. Inhibition of myeloid differentiation protein 2 attenuates renal ischemia/reperfusion-induced oxidative stress and inflammation via suppressing TLR4/TRAF6/NF-kB pathway. Life Sci. 2020, 256, 117864. [Google Scholar] [CrossRef]
- Damman, J.; Bloks, V.W.; Daha, M.R.; Van Der Most, P.J.; Sanjabi, B.; Van Der Vlies, P.; Snieder, H.; Ploeg, R.J.; Krikke, C.; Leuvenink, H.G.D.; et al. Hypoxia and Complement-and-Coagulation Pathways in the Deceased Organ Donor as the Major Target for Intervention to Improve Renal Allograft Outcome. Transplantation 2015, 99, 1293–1300. [Google Scholar] [CrossRef]
- Famulski, K.S.; de Freitas, D.G.; Kreepala, C.; Chang, J.; Sellares, J.; Sis, B.; Einecke, G.; Mengel, M.; Reeve, J.; Halloran, P.F. Molecular Phenotypes of Acute Kidney Injury in Kidney Transplants. J. Am. Soc. Nephrol. 2012, 23, 948–958. [Google Scholar] [CrossRef] [Green Version]
- Cippà, P.E.; Sun, B.; Liu, J.; Chen, L.; Naesens, M.; McMahon, A.P. Transcriptional trajectories of human kidney injury progression. J. Clin. Investig. 2018, 3, e123151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, A.; Su, J.; Liu, R.; Zhao, S.; Li, W.; Xu, X.; Li, D.; Shi, J.; Bin Gu, B.; Zhang, J.; et al. Sexual dimorphism in glucose metabolism is shaped by androgen-driven gut microbiome. Nat. Commun. 2021, 12, 7080. [Google Scholar] [CrossRef]
- Robin, X.; Turck, N.; Hainard, A.; Tiberti, N.; Lisacek, F.; Sanchez, J.-C.; Müller, M. pROC: An open-source package for R and S+ to analyze and compare ROC curves. BMC Bioinform. 2011, 12, 77. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R Package for Comparing Biological Themes Among Gene Clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Kirsch, R.; Koutrouli, M.; Nastou, K.; Mehryary, F.; Hachilif, R.; Gable, A.L.; Fang, T.; Doncheva, N.T.; Pyysalo, S.; et al. The STRING database in 2023: Protein–protein association networks and functional enrichment analyses for any sequenced genome of interest. Nucleic Acids Res. 2022, 51, D638–D646. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. Fast R Functions for Robust Correlations and Hierarchical Clustering. J. Stat. Softw. 2012, 46, i11. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Zhang, H.; Liu, Y.; Liu, Y.-X.; Huang, L. EVenn: Easy to create repeatable and editable Venn diagrams and Venn networks online. J. Genet. Genom. 2021, 48, 863–866. [Google Scholar] [CrossRef]
- Pan, T.; He, Y.; Chen, H.; Pei, J.; Li, Y.; Zeng, R.; Xia, J.; Zuo, Y.; Qin, L.; Chen, S.; et al. Identification and Validation of a Prognostic Gene Signature for Diffuse Large B-Cell Lymphoma Based on Tumor Microenvironment-Related Genes. Front. Oncol. 2021, 11, 614211. [Google Scholar] [CrossRef]
- Kim, N.; Han, I.W.; Ryu, Y.; Hwang, D.W.; Heo, J.S.; Choi, D.W.; Shin, S.H. Predictive Nomogram for Early Recurrence after Pancreatectomy in Resectable Pancreatic Cancer: Risk Classification Using Preoperative Clinicopathologic Factors. Cancers 2020, 12, 137. [Google Scholar] [CrossRef] [Green Version]
- Andrade-Oliveira, V.; Amano, M.T.; Correa-Costa, M.; Castoldi, A.; Felizardo, R.J.; de Almeida, D.C.; Bassi, E.J.; Moraes-Vieira, P.M.; Hiyane, M.I.; Rodas, A.C.; et al. Gut Bacteria Products Prevent AKI Induced by Ischemia-Reperfusion. J. Am. Soc. Nephrol. 2015, 26, 1877–1888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, R.-Y.; Fang, X.-L.; Zheng, X.-B.; Lv, W.-Z.; Li, Y.-J.; Rage, H.I.; He, Q.-L.; Zhu, W.-P.; Cui, T.-X. Salidroside and FG-4592 ameliorate high glucose-induced glomerular endothelial cells injury via HIF upregulation. Biomed. Pharmacother. 2019, 118, 109175. [Google Scholar] [CrossRef] [PubMed]
- Emal, D.; Rampanelli, E.; Stroo, I.; Butter, L.M.; Teske, G.J.; Claessen, N.; Stokman, G.; Florquin, S.; Leemans, J.C.; Dessing, M.C. Depletion of Gut Microbiota Protects against Renal Ischemia-Reperfusion Injury. J. Am. Soc. Nephrol. 2016, 28, 1450–1461. [Google Scholar] [CrossRef]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem 2023 update. Nucleic Acids Res. 2023, 51, D1373–D1380. [Google Scholar] [CrossRef] [PubMed]
- Kerwin, S.M. ChemBioOffice Ultra 2010 Suite. J. Am. Chem. Soc. 2010, 132, 2466–2467. [Google Scholar] [CrossRef]
- Xie, M.; Xie, R.; Xie, S.; Wu, Y.; Wang, W.; Li, X.; Xu, Y.; Liu, B.; Zhou, Y.; Wang, T.; et al. Thioredoxin interacting protein (TXNIP) acts as a tumor suppressor in human prostate cancer. Cell Biol. Int. 2020, 44, 2094–2106. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Description |
|---|---|
| GO:1900744 | regulation of p38 MAPK cascade |
| GO:0038066 | p38 MAPK cascade |
| GO:1900745 | positive regulation of p38 MAPK cascade |
| GO:0032872 | regulation of stress-activated MAPK cascade |
| GO:0070302 | regulation of stress-activated protein kinase signaling cascade |
| GO:0051403 | stress-activated MAPK cascade |
| GO:0016607 | nuclear speck |
| GO:0047485 | protein N-terminus binding |
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| 18S | CGATCCGAGGGCCTCACTA | AGTCCCTGCCCTTTGTACACA |
| NGAL | GCCTCAAGGACGACAACATC | CTGAACCATTGGGTCTCTGC |
| KIM-1 | TTGCCTTCCGTGTCTCTAAG | AGATGTTGTCTTCAGCTCGG |
| GADD45A | CCGAAAGGATGGACACGGTG | TTATCGGGGTCTACGTTGAGC |
| GADD45B | GCCAAACTGATGAATGTGGACC | GAACGACTGGATCAGGGTGA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, M.; Xie, R.; Huang, P.; Yap, D.Y.H.; Wu, P. GADD45A and GADD45B as Novel Biomarkers Associated with Chromatin Regulators in Renal Ischemia-Reperfusion Injury. Int. J. Mol. Sci. 2023, 24, 11304. https://doi.org/10.3390/ijms241411304
Xie M, Xie R, Huang P, Yap DYH, Wu P. GADD45A and GADD45B as Novel Biomarkers Associated with Chromatin Regulators in Renal Ischemia-Reperfusion Injury. International Journal of Molecular Sciences. 2023; 24(14):11304. https://doi.org/10.3390/ijms241411304
Chicago/Turabian StyleXie, Ming, Ruiyan Xie, Pengcheng Huang, Desmond Y. H. Yap, and Peng Wu. 2023. "GADD45A and GADD45B as Novel Biomarkers Associated with Chromatin Regulators in Renal Ischemia-Reperfusion Injury" International Journal of Molecular Sciences 24, no. 14: 11304. https://doi.org/10.3390/ijms241411304