
Regulatory Mechanisms Underlying Arsenic Uptake, Transport, and Detoxification in Rice

Abstract
:1. Introduction
2. The Toxic Effects of As
3. The Molecular Mechanisms of As Uptake, Transport, and Redistribution in Rice
3.1. As Uptake
3.1.1. Uptake of Arsenite in Rice
3.1.2. Uptake of Arsenate in Rice
3.1.3. Uptake of Other As Species in Rice
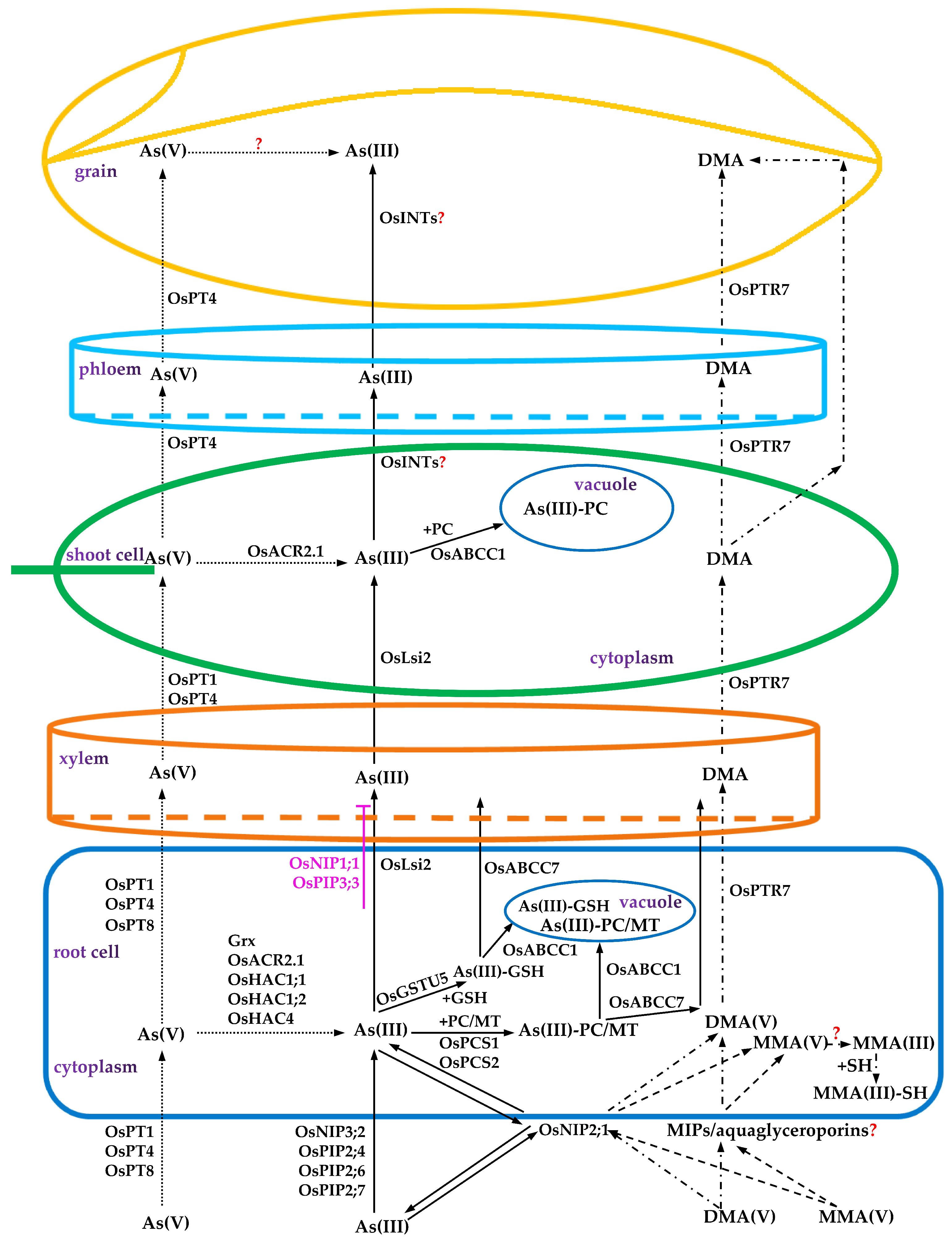
3.2. Arsenic Transport
3.3. Arsenic Redistribution
4. Regulatory Mechanisms
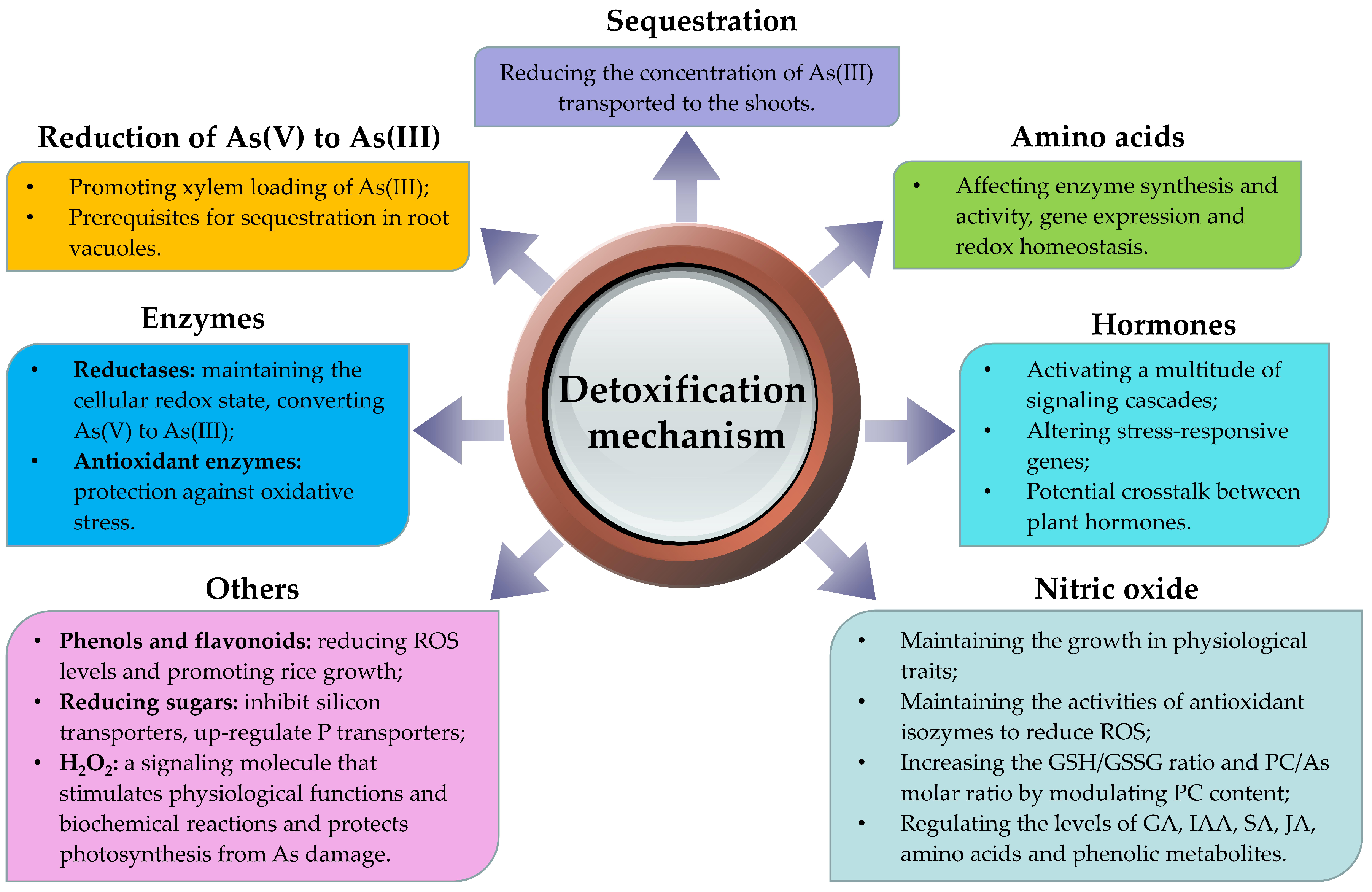
5. Detoxification Mechanisms
5.1. As Speciation Conversion
5.2. Chelation and Vacuolar Compartmentalization
5.3. Enzymes
5.4. Amino Acids
5.5. Nitric Oxide (NO)
5.6. Hormones
5.7. Other Small Molecules
6. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| Acr3p | arsenical resistance3 |
| APX | ascorbate peroxidase |
| As | arsenic |
| As(III) | arsenite |
| As(III)-GSH | As(III) glutathione conjugate |
| As(III)-PC/MT | As(III)-phytochelatin/metallothionein conjugates |
| As(V) | arsenate |
| AsGlu3 | tris-As-glutathione |
| ATP | adenosine triphosphate |
| CAD8 | cadherin-8 |
| CAT | catalase |
| DAR | dehydroascorbate reductase |
| DMA | dimethyl arsenic acid |
| DNA | deoxyribonucleic acid |
| FeRB | Fe(III)-reducing bacteria |
| GA | gibberellin |
| GR | glutathione reductase |
| Grx | glutaredoxin |
| GSH | glutathione |
| H2O2 | hydrogen peroxide |
| IAA | auxin |
| INTs | inositol transporters |
| JA | jasmonate |
| MATE | multidrug and toxic compound extrusion |
| Me-JA | methyl jasmonate |
| MIPs | membrane intrinsic proteins |
| miRNAs | micro ribonucleic acids |
| MMA(III)-SH | monomethyl arsenic acid sulfhydryl conjugate |
| MMA | monomethyl arsenic acid |
| MT | metallothionein |
| NIPs | nodulin 26-like intrinsic proteins |
| NO | nitric oxide |
| NRAMP | natural resistance-associated macrophage protein |
| OPR | oxophytodienoate reductase |
| OsABCC | C-TYPE ABC subfamily transporter |
| OsACR2.1 | arsenate reductase 2.1 |
| OsADH2 | alcohol dehydrogenase 2 |
| OsARM1 | arsenite-responsive MYB1 |
| OsCLT1 | CRT-like transporter 1 |
| OsDHFR | dihydrofolate reductase |
| OsGA2ox4 | GA 2-oxidases 4 |
| OsGSTU5 | glutathione-S-transferase 5 |
| OsHAC | high arsenic content |
| OsHIR1 | heavy metal-induced ring E3 ligase 1 |
| OsIPT3 | adenylate isopentenyl transferase 3 |
| OsLAC3 | laccase gene 3 |
| OsLsi | low silicon |
| OsNAS3 | nicotianamine synthase gene 3 |
| OsNCED2 | 9-cis-epoxycarotenoid dioxygenase 2 |
| OsNLA1 | nitrogen limitation adaptation 1 |
| OsOASTL-A1 | O-acetyl serine(thiol) lyase 1 |
| OsPCS | phytochelatin synthase |
| OsPHF1 | phosphate transporter traffic facilitator 1 |
| OsPHR2 | phosphate starvation response 2 |
| OsPIN9 | probable auxin efflux carrier component 9 |
| OsPRX38 | peroxidase 38 |
| OsPT(OsPht) | phosphate transporter |
| OsPTR7 | putative peptide transporter 7 |
| OsWNK9 | with no lysine 9 |
| OsVIT1 | vacuolar membrane transporter 1 |
| OsZIP7 | ZRT and IRT-like protein 7 |
| PCs | phytochelatins |
| PIPs | plasma membrane intrinsic proteins |
| PTMs | post-translational modifications |
| psb | photosystem subunit |
| RING | really interesting new gene |
| RNS | reactive nitrogen species |
| ROS | reactive oxygen species |
| SA | salicylic acid |
| SH | sulfhydryl |
| Si | silicon |
| SIPs | small basic intrinsic proteins |
| SL | strigolactone |
| SOD | superoxide dismutase |
| SRB | sulfate-reducing bacteria |
| TMA(V) | trimethylarsine |
| TIPs | tonoplast intrinsic proteins |
| μ-XRF | synchrotron micro-X-ray fluorescence |
| WT | wild type |
References
- IARC Working Group on the Evaluation of Carcinogenic Risks to Humans. Some drinking-water disinfectants and contaminants, including arsenic. IARC Monogr. Eval. Carcinog. Risks Hum. 2004, 84, 1–477. [Google Scholar]
- Biswas, P.; Hossain, M.; Patra, P.K. Arsenic hydrogeochemistry, quality assessment, and associated health risks of groundwater through the novel water pollution index (WPI) and GIS approach. Groundw. Sustain. Dev. 2023, 21, 100944. [Google Scholar] [CrossRef]
- Hassan, A. Inorganic-based pesticides: A review article. Egypt. Sci. J. Pestic. 2019, 5, 39–52. [Google Scholar]
- Palmer, M.J.; Jamieson, H.E.; Borčinová Radková, A.; Maitland, K.; Oliver, J.; Falck, H.; Richardson, M. Mineralogical, geospatial, and statistical methods combined to estimate geochemical background of arsenic in soils for an area impacted by legacy mining pollution. Sci. Total Environ. 2021, 776, 145926. [Google Scholar] [CrossRef] [PubMed]
- de Mello, J.W.V.; Talbott, J.L.; Scott, J.; Roy, W.R.; Stucki, J.W. Arsenic speciation in arsenic-rich Brazilian soils from gold mining sites under anaerobic incubation. Environ. Sci. Pollut. Res. Int. 2007, 14, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Buccino, M.; Daliri, M.; Buttarazzi, M.N.; Del Giudice, G.; Calabrese, M.; Somma, R. Arsenic contamination at the Bagnoli Bay seabed (South Italy) via particle tracking numerical modeling: Pollution patterns from stationary climatic forcings. Chemosphere 2022, 303, 134955. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Naghibi, S.A.; Motevalli, A.; Hashemi, H. Human-induced arsenic pollution modeling in surface waters—An integrated approach using machine learning algorithms and environmental factors. J. Environ. Manag. 2022, 305, 114347. [Google Scholar] [CrossRef]
- Cüce, H.; Kalıpcı, E.; Ustaoğlu, F.; Kaynar, İ.; Baser, V.; Türkmen, M. Multivariate statistical methods and GIS based evaluation of the health risk potential and water quality due to arsenic pollution in the Kızılırmak river. Int. J. Sediment. Res. 2022, 37, 754–765. [Google Scholar] [CrossRef]
- Brammer, H.; Ravenscroft, P. Arsenic in groundwater: A threat to sustainable agriculture in south and south-east Asia. Environ. Int. 2009, 35, 647–654. [Google Scholar] [CrossRef]
- Bundschuh, J.; Niazi, N.K.; Alam, M.A.; Berg, M.; Herath, I.; Tomaszewska, B.; Maity, J.P.; Ok, Y.S. Global arsenic dilemma and sustainability. J. Hazard. Mater. 2022, 436, 129197. [Google Scholar] [CrossRef]
- Shaji, E.; Santosh, M.; Sarath, K.V.; Prakash, P.; Deepchand, V.; Divya, B.V. Arsenic contamination of groundwater: A global synopsis with focus on the Indian Peninsula. Geosci. Front. 2021, 12, 101079. [Google Scholar] [CrossRef]
- Styblo, M.; Del Razo, L.M.; Vega, L.; Germolec, D.; Lecluyse, E.; Hamilton, G.; Reed, W.; Wang, C.; Cullen, W.; Thomas, D. Comparative toxicity of trivalent and pentavalent inorganic and methylated arsenicals in rat and human cells. Arch. Toxicol. 2000, 74, 289–299. [Google Scholar] [CrossRef]
- Emily, S. Contamination: The toxic side of rice. Nature 2014, 514, S62–S63. [Google Scholar]
- Su, Y.H.; McGrath, S.P.; Zhao, F.J. Rice is more efficient in arsenite uptake and translocation than wheat and barley. Plant Soil 2010, 328, 27–34. [Google Scholar] [CrossRef]
- Piacentini, D.; Della Rovere, F.; Lanni, F.; Cittadin i, M.; Palombi, M.; Fattorini, L.; Cecchetti, V.; Altamura, M.M.; Falasca, G. Brassinosteroids interact with nitric oxide in the response of rice root systems to arsenic stress. Environ. Exp. Bot. 2023, 209, 105287. [Google Scholar] [CrossRef]
- Li, Y.; Chu, Y.T.; Sun, H.Y.; Bao, Q.L.; Huang, Y.Z. Melatonin alleviates arsenite toxicity by decreasing the arsenic accumulation in cell protoplasts and increasing the antioxidant capacity in rice. Chemosphere 2023, 312 Pt 1, 137292. [Google Scholar] [CrossRef] [PubMed]
- Guillod-Magnin, R.; Brüschweiler, B.J.; Aubert, R.; Haldimann, M. Arsenic species in rice and rice-based products consumed by toddlers in Switzerland. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2018, 35, 1164–1178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindsay, E.R.; Maathuis, F.J.M. New molecular mechanisms to reduce arsenic in crops. Trends Plant Sci. 2017, 22, 1016–1026. [Google Scholar] [CrossRef]
- Das, S.; Biswas, A.K. Comparative study of silicon and selenium to modulate chloroplast pigments levels, hill activity, photosynthetic parameters and carbohydrate metabolism under arsenic stress in rice seedlings. Environ. Sci. Pollut. Res. Int. 2022, 29, 19508–19529. [Google Scholar] [CrossRef]
- Faizan, M.; Sehar, S.; Rajput, V.D.; Faraz, A.; Afzal, S.; Minkina, T.; Sushkova, S.; Adil, M.F.; Yu, F.; Alatar, A.A.; et al. Modulation of cellular redox status and antioxidant defense system after synergistic application of zinc oxide nanoparticles and salicylic acid in rice (Oryza sativa) plant under arsenic stress. Plants 2021, 10, 2254. [Google Scholar] [CrossRef]
- Bidi, H.; Fallah, H.; Niknejad, Y.; Barari Tari, D. Iron oxide nanoparticles alleviate arsenic phytotoxicity in rice by improving iron uptake, oxidative stress tolerance and diminishing arsenic accumulation. Plant Physiol. Biochem. 2021, 163, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Bhadwal, S.; Sharma, S. Selenium alleviates carbohydrate metabolism and nutrient composition in arsenic stressed rice plants. Rice Sci. 2022, 29, 385–396. [Google Scholar] [CrossRef]
- Begum, M.C.; Islam, M.S.; Islam, M.; Amin, R.; Parvez, M.S.; Kabir, A.H. Biochemical and molecular responses underlying differential arsenic tolerance in rice (Oryza sativa L.). Plant Physiology and Biochemistry 2016, 104, 266–277. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Siddika, A.; Rahman, K.; Nahar, K. Supplementation with Ascophyllum nodosum extracts mitigates arsenic toxicity by modulating reactive oxygen species metabolism and reducing oxidative stress in rice. Ecotoxicol. Environ. Saf. 2023, 255, 114819. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.X.; Li, J.; Lou, B.; Wu, R.R.; Wang, G.; Lu, C.W.; Wang, H.H.; Pi, J.B.; Xu, Y.Y. The role of reactive oxygen species in arsenic toxicity. Biomolecules 2020, 10, 240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvarenga, I.F.S.; Dos Santos, F.E.; Silveira, G.L.; Andrade-Vieira, L.F.; Martins, G.C.; Guilherme, L.R.G. Investigating arsenic toxicity in tropical soils: A cell cycle and DNA fragmentation approach. Sci. Total Environ. 2020, 698, 134272. [Google Scholar] [CrossRef]
- Shukla, J.; Narayan, S.; Shirke, P.A.; Mishra, A.; Kumar, M. Reduction of arsenic accumulation in rice grain by endophytic fungus Serendipita indica. Rhizosphere 2023, 26, 100680. [Google Scholar] [CrossRef]
- Niazi, N.K.; Hussain, M.M.; Bibi, I.; Natasha, S.M.; Ali, F.; Iqbal, J.; Shaheen, S.M.; Abdelrahman, H.; Akhtar, W.; Wang, H.; et al. significance of eighteen rice genotypes on arsenic accumulation, physiological response and potential health risk. Sci. Total Environ. 2022, 832, 155004. [Google Scholar] [CrossRef]
- Majumder, B.; Das, S.; Pal, B.; Biswas, A.K. Evaluation of arsenic induced toxicity based on arsenic accumulation, translocation and its implications on physio-chemical changes and genomic instability in indica rice (Oryza sativa L.) cultivars. Ecotoxicology 2020, 29, 13–34. [Google Scholar] [CrossRef]
- Kumar, A.; Basu, S.; Kumar, G. Evaluating the effect of seed-priming for improving arsenic tolerance in rice. J. Plant Biochem. Biotechnol. 2022, 31, 197–201. [Google Scholar] [CrossRef]
- Kumar, A.; Basu, S.; Kumari, S.; Shekhar, S.; Kumar, G. Effective antioxidant defense prevents nitro-oxidative stress under arsenic toxicity: A study in rice genotypes of eastern indo-gangetic plains. Environ. Exp. Bot. 2022, 204, 105084. [Google Scholar] [CrossRef]
- Ghorbani, A.; Tafteh, M.; Roudbari, N.; Pishkar, L.; Zhang, W.Y.; Wu, C. Piriformospora indica augments arsenic tolerance in rice (Oryza sativa) by immobilizing arsenic in roots and improving iron translocation to shoots. Ecotoxicol. Environ. Saf. 2021, 209, 111793. [Google Scholar] [CrossRef] [PubMed]
- Thongnok, S.; Siripornadulsil, W.; Siripornadulsil, S. Responses to arsenic stress of rice varieties coinoculated with the heavy metal-resistant and rice growth-promoting bacteria Pseudomonas stutzeri and Cupriavidus taiwanensis. Plant Physiol. Biochem. 2022, 191, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, L.M.V.P.; Martins, S.C.V.; Detmann, K.C.; Silva, P.E.M.; Lavinsky, A.O.; Silva, M.M.; Detmann, E.; Araújo, W.L.; DaMatta, F.M. Silicon nutrition alleviates the negative impacts of arsenic on the photosynthetic apparatus of rice leaves: An analysis of the key limitations of photosynthesis. Physiol. Plant. 2014, 152, 355–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murugaiyan, V.; Ali, J.; Mahender, A.; Aslam, U.M.; Jewel, Z.A.; Pang, Y.L.; Marfori-Nazarea, C.M.; Wu, L.B.; Frei, M.; Li, Z.K. Mapping of genomic regions associated with arsenic toxicity stress in a backcross breeding populations of rice (Oryza sativa L.). Rice 2019, 12, 61. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Dubey, R.S.; Tripathi, R.D.; Chakrabarty, D.; Trivedi, P.K. Omics and biotechnology of arsenic stress and detoxification in plants: Current updates and prospective. Environ. Int. 2015, 74, 221–230. [Google Scholar] [CrossRef]
- Xu, X.Y.; McGrath, S.P.; Meharg, A.A.; Zhao, F.J. Growing rice aerobically markedly decreases arsenic accumulation. Environ. Sci. Technol. 2008, 42, 5574–5579. [Google Scholar] [CrossRef]
- de Oliveira, L.M.; Suchismita, D.; Da Silva, E.B.; Gao, P.; Vardanyan, L.; Liu, Y.G.; Ma, L.Q. Interactive effects of chromate and arsenate on their uptake and speciation in Pteris ensiformis. Plant Soil 2018, 422, 515–526. [Google Scholar] [CrossRef]
- Kamiya, T.; Islam, R.; Duan, G.; Uraguchi, S.; Fujiwara, T. Phosphate deficiency signaling pathway is a target of arsenate and phosphate transporter OsPT1 is involved in As accumulation in shoots of rice. Soil Sci. Plant Nutr. 2013, 59, 580–590. [Google Scholar] [CrossRef]
- Cao, Y.; Sun, D.; Ai, H.; Mei, H.Y.; Liu, X.; Sun, S.B.; Xu, G.H.; Liu, Y.G.; Chen, Y.S.; Ma, L.Q. Knocking out OsPT4 gene decreases arsenate uptake by rice plants and inorganic arsenic accumulation in rice grains. Environ. Sci. Technol. 2017, 51, 12131–12138. [Google Scholar] [CrossRef]
- Wu, Z.C.; Ren, H.Y.; McGrath, S.P.; Wu, P.; Zhao, F.J. Investigating the contribution of the phosphate transport pathway to arsenic accumulation in rice. Plant Physiol. 2011, 157, 498–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.T.; Zhang, W.T.; Mao, C.Z.; Xu, G.H.; Zhao, F.J. The role of OsPT8 in arsenate uptake and varietal difference in arsenate tolerance in rice. J. Exp. Bot. 2016, 67, 6051–6059. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.Y.; Tian, Z.H.; Yang, X.L.; Liu, B.H.; Yang, J.; Lin, H.H. The role of OsNLA1 in regulating arsenate uptake and tolerance in rice. J. Plant Physiol. 2019, 236, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.K.; Chen, Y.; Che, J.; Konishi, N.; Tang, Z.; Miller, A.J.; Ma, J.F.; Zhao, F.J. Decreasing arsenic accumulation in rice by overexpressing OsNIP1;1 and OsNIP3;3 through disrupting arsenite radial transport in roots. New Phytol. 2018, 219, 641–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Xu, X.Y.; Su, Y.H.; McGrath, S.P.; Zhao, F.J. Transporters of arsenite in rice and their role in arsenic accumulation in rice grain. Proc. Natl. Acad. Sci. USA 2008, 105, 9931–9935. [Google Scholar] [CrossRef] [PubMed]
- Kumarathilaka, P.; Seneweera, S.; Meharg, A.; Bundschuh, J. Arsenic accumulation in rice (Oryza sativa L.) is influenced by environment and genetic factors. Sci. Total Environ. 2018, 642, 485–496. [Google Scholar] [CrossRef] [Green Version]
- Li, R.Y.; Ago, Y.; Liu, W.J.; Mitani, N.; Feldmann, J.; McGrath, S.P.; Ma, J.F.; Zhao, F.J. The rice aquaporin Lsi1 mediates uptake of methylated arsenic species. Plant Physiol. 2009, 150, 2071–2080. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Sun, S.K.; Tang, Z.; Liu, G.; Moore, K.L.; Maathuis, F.J.M.; Miller, A.J.; McGrath, S.P.; Zhao, F.J. The Nodulin 26-like intrinsic membrane protein OsNIP3;2 is involved in arsenite uptake by lateral roots in rice. J. Exp. Bot. 2017, 68, 3007–3016. [Google Scholar] [CrossRef] [Green Version]
- Mosa, K.A.; Kumar, K.; Chhikara, S.; Mcdermott, J.; Liu, Z.; Musante, C.; White, J.C.; Dhankher, O.P. Members of rice plasma membrane intrinsic proteins subfamily are involved in arsenite permeability and tolerance in plants. Transgenic Res. 2012, 21, 1265–1277. [Google Scholar] [CrossRef]
- Verma, P.K.; Verma, S.; Pande, V.; Mallick, S.; Deo Tripathi, R.; Dhankher, O.P.; Chakrabarty, D. Overexpression of rice glutaredoxin OsGrx_C7 and OsGrx_C2.1 reduces intracellular arsenic accumulation and increases tolerance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 740. [Google Scholar]
- Verma, P.K.; Verma, S.; Tripathi, R.D.; Chakrabarty, D. A rice glutaredoxin regulate the expression of aquaporin genes and modulate root responses to provide arsenic tolerance. Ecotoxicol. Environ. Saf. 2020, 195, 110471. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Yamaki, T.; Yamaji, N.; Ko, D.; Jung, K.; Fujii-Kashino, M.; An, G.; Martinoia, E.; Lee, Y.; Ma, J.F. A rice ABC transporter, OsABCC1, reduces arsenic accumulation in the grain. Proc. Natl. Acad. Sci. USA 2014, 111, 15699–15704. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Chen, Y.; Miller, A.J.; Zhao, F.J. The C-type ATP-binding cassette transporter OsABCC7 is involved in the root-to-shoot translocation of arsenic in rice. Plant Cell Physiol. 2019, 60, 1525–1535. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, M.; Sharma, D.; Dwivedi, S.; Singh, M.; Tripathi, R.D.; Trivedi, P.K. Expression in Arabidopsis and cellular localization reveal involvement of rice NRAMP, OsNRAMP1, in arsenic transport and tolerance. Plant Cell Environ. 2014, 37, 140–152. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Chen, Y.; Chen, F.; Ji, Y.C.; Zhao, F.J. OsPTR7 (OsNPF8.1), a putative peptide transporter in rice, is involved in dimethylarsenate accumulation in rice grain. Plant Cell Physiol. 2017, 58, 904–913. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.T.; Xu, X.; Tang, Z.; Zhang, W.W.; Huang, X.Y.; Zhao, F.J. OsWRKY28 regulates phosphate and arsenate accumulation, root system architecture and fertility in rice. Front. Plant Sci. 2018, 9, 1330. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.; Kuramata, M.; Abe, T.; Yamaguchi, N.; Takagi, H.; Tanikawa, H.; Iino, M.; Sugimoto, K.; Ishikawa, S. Deficiency in alcohol dehydrogenase 2 reduces arsenic in rice grains by suppressing silicate transporters. Plant Physiol. 2021, 186, 611–623. [Google Scholar] [CrossRef]
- Das, N.; Bhattacharya, S.; Bhattacharyya, S.; Maiti, M.K. Expression of rice MATE family transporter OsMATE2 modulates arsenic accumulation in tobacco and rice. Plant Mol. Biol. 2018, 98, 101–120. [Google Scholar] [CrossRef]
- Wang, F.Z.; Chen, M.X.; Yu, L.J.; Xie, L.J.; Yuan, L.B.; Qi, H.; Xiao, M.; Guo, W.X.; Chen, Z.; Yi, K.; et al. OsARM1, an R2R3 MYB transcription factor, is involved in regulation of the response to arsenic stress in rice. Front. Plant Sci. 2017, 8, 1868. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.J.; Ma, J.F.; Meharg, A.A.; McGrath, S.P. Arsenic uptake and metabolism in plants. New Phytol. 2009, 181, 777–794. [Google Scholar] [CrossRef]
- Xu, W.Z.; Dai, W.T.; Yan, H.L.; Li, S.; Shen, H.L.; Chen, Y.S.; Xu, H.; Sun, Y.Y.; He, Z.Y.; Ma, M. Arabidopsis NIP3,1 plays an important role in arsenic uptake and root-to-shoot translocation under arsenite stress conditions. Mol. Plant 2015, 8, 722–733. [Google Scholar] [CrossRef] [Green Version]
- Katsuhara, M.; Sasano, S.; Horie, T.; Matsumoto, T.; Rhee, J.; Shibasaka, M. Functional and molecular characteristics of rice and barley NIP aquaporins transporting water, hydrogen peroxide and arsenite. Plant Biotechnol. 2014, 31, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef]
- Chen, Y.; Moore, K.L.; Miller, A.J.; McGrath, S.P.; Ma, J.F.; Zhao, F.J. The role of nodes in arsenic storage and distribution in rice. J. Exp. Bot. 2015, 66, 3717–3724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef] [PubMed]
- Mitani, N.; Chiba, Y.; Yamaji, N.; Ma, J.F. Identification and characterization of maize and barley Lsi2-like silicon efflux transporters reveals a distinct silicon uptake system from that in rice. Plant Cell 2009, 21, 2133–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maurel, C.; Boursiac, Y.; Luu, D.; Santoni, V.; Shahzad, Z.; Verdoucq, L. Aquaporins in plants. Physiol. Rev. 2015, 95, 1321–1358. [Google Scholar] [CrossRef]
- Takahashi, Y.; Minamikawa, R.; Hattori, K.H.; Kurishima, K.; Kihou, N.; Yuita, K. Arsenic behavior in paddy fields during the cycle of flooded and non-flooded periods. Environ. Sci. Technol. 2004, 38, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Paszkowski, U.; Kroken, S.; Roux, C.; Briggs, S.P. Rice phosphate transporters include an evolutionarily divergent gene specifically activated in arbuscular mycorrhizal symbiosis. Proc. Natl. Acad. Sci. USA 2002, 99, 13324–13329. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Li, P.; Xu, T.Q.; Zeng, L.T.; Cheng, D.; Yang, M.; Luo, J.; Lian, X.M. OsPT4 contributes to arsenate uptake and transport in rice. Front. Plant Sci. 2017, 8, 2197. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.H.; Zhang, R.; Lilley, R.M. Methylation of arsenic in vitro by cell extracts from bentgrass (Agrostis tenuis): Effect of acute exposure of plants to arsenate. Funct. Plant Biol. 2002, 29, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Huang, H.; Sun, G.X.; Zhao, F.J.; Zhu, Y.G. Pathways and relative contributions to arsenic volatilization from rice plants and paddy soil. Environ. Sci. Technol. 2012, 46, 8090–8096. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tang, Z.; Chen, X.P.; Wang, X.; Zhou, W.X.; Tang, Z.; Zhang, J.; Zhao, F.J. Water management impacts the soil microbial communities and total arsenic and methylated arsenicals in rice grains. Environ. Pollut. 2019, 247, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Rensing, C.; Rosen, B.P.; Zhu, Y.G. Arsenic biomethylation by photosynthetic organisms. Trends Plant Sci. 2012, 17, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Kadohashi, K.; Maki, T.; Hasegawa, H. Transport of DMAA and MMAA into rice (Oryza sativa L.) roots. Environ. Exp. Bot. 2011, 72, 41–46. [Google Scholar] [CrossRef] [Green Version]
- Chandrakar, V.; Naithani, S.C.; Keshavkant, S. Arsenic-induced metabolic disturbances and their mitigation mechanisms in crop plants: A review. Biologia 2016, 71, 367–377. [Google Scholar] [CrossRef]
- Mitra, A.; Chatterjee, S.; Gupta, D. Uptake, Transport, and remediation of arsenic by Algae and higher plants. In Arsenic Contamination in the Environment: The Issues and Solutions; Springer: Cham, Switzerland, 2017; pp. 145–169. [Google Scholar]
- Zhang, X.; Wu, S.L.; Ren, B.H.; Chen, B.D. Water management, rice varieties and mycorrhizal inoculation influence arsenic concentration and speciation in rice grains. Mycorrhiza 2016, 26, 299–309. [Google Scholar] [CrossRef]
- Zhao, F.J.; Zhu, Y.G.; Meharg, A.A. Methylated arsenic species in rice: Geographical variation, origin, and uptake mechanisms. Environ. Sci. Technol. 2013, 47, 3957–3966. [Google Scholar] [CrossRef]
- Zhao, F.J.; Stroud, J.L.; Khan, M.A.; McGrath, S.P. Arsenic translocation in rice investigated using radioactive 73As tracer. Plant Soil. 2012, 350, 413–420. [Google Scholar] [CrossRef]
- Seyfferth, A.L.; Webb, S.M.; Andrews, J.C.; Fendorf, S. Defining the distribution of arsenic species and plant nutrients in rice (Oryza sativa L.) from the root to the grain. Geochim. Cosmochim. Acta 2011, 75, 6655–6671. [Google Scholar] [CrossRef]
- Punshon, T.; Jackson, B.P.; Meharg, A.A.; Warczack, T.; Scheckel, K.; Guerinot, M.L. Understanding arsenic dynamics in agronomic systems to predict and prevent uptake by crop plants. Sci. Total Environ. 2017, 581–582, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Suriyagoda, L.D.B.; Dittert, K.; Lambers, H. Mechanism of arsenic uptake, translocation and plant resistance to accumulate arsenic in rice grains. Agric. Ecosyst. Environ. 2018, 253, 23–37. [Google Scholar] [CrossRef]
- Lindsay, E.R.; Maathuis, F.J.M. Arabidopsis thaliana NIP7;1 is involved in tissue arsenic distribution and tolerance in response to arsenate. FEBS Lett. 2016, 590, 779–786. [Google Scholar] [CrossRef] [Green Version]
- Duan, G.L.; Hu, Y.; Schneider, S.; McDermott, J.; Chen, J.; Sauer, N.; Rosen, B.P.; Daus, B.; Liu, Z.; Zhu, Y.G. Inositol transporters AtINT2 and AtINT4 regulate arsenic accumulation in Arabidopsis seeds. Nat. Plants 2016, 2, 15202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, P.G.; Koch, I.; Reimer, K.J. An investigation of arsenic compounds in fur and feathers using X-ray absorption spectroscopy speciation and imaging. Sci. Total Environ. 2008, 390, 198–204. [Google Scholar] [CrossRef]
- Wu, C.; Ye, Z.; Shu, W.; Zhu, Y.; Wong, M. Arsenic accumulation and speciation in rice are affected by root aeration and variation of genotypes. J. Exp. Bot. 2011, 62, 2889–2898. [Google Scholar] [CrossRef] [Green Version]
- Talukder, A.S.M.H.; Meisner, C.A.; Sarkar, M.A.R.; Islam, M.S.; Sayre, K.D. Effects of water management, arsenic and phosphorus levels on rice yield in high-arsenic soil-water system. Rice Sci. 2014, 21, 99–107. [Google Scholar] [CrossRef]
- Sarwar, T.; Khan, S.; Muhammad, S.; Amin, S. Arsenic speciation, mechanisms, and factors affecting rice uptake and potential human health risk: A systematic review. Environ. Technol. Innov. 2021, 22, 101392. [Google Scholar] [CrossRef]
- Raab, A.; Williams, P.N.; Meharg, A.; Feldmann, J. Uptake and translocation of inorganic and methylated arsenic species by plants. Environ. Chem. 2007, 4, 197. [Google Scholar] [CrossRef] [Green Version]
- Lomax, C.; Liu, W.J.; Wu, L.; Xue, K.; Xiong, J.; Zhou, J.; McGrath, S.P.; Meharg, A.A.; Miller, A.J.; Zhao, F.J. Methylated arsenic species in plants originate from soil microorganisms. New Phytol. 2012, 193, 665–672. [Google Scholar] [CrossRef]
- Carey, A.; Scheckel, K.G.; Lombi, E.; Newville, M.; Choi, Y.; Norton, G.J.; Charnock, J.M.; Feldmann, J.; Price, A.H.; Meharg, A.A. Grain unloading of arsenic species in rice. Plant Physiol. 2010, 152, 309–319. [Google Scholar] [CrossRef] [Green Version]
- Carey, A.; Norton, G.J.; Deacon, C.; Scheckel, K.G.; Lombi, E.; Punshon, T.; Guerinot, M.L.; Lanzirotti, A.; Newville, M.; Choi, Y.; et al. Phloem transport of arsenic species from flag leaf to grain during grain filling. New Phytol. 2011, 192, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Ye, W.L.; Zhang, J.J.; Fan, T.; Lu, H.J.; Chen, H.Y.; Li, X.D.; Hua, R.M. Arsenic speciation in the phloem exudates of rice and its role in arsenic accumulation in rice grains. Ecotoxicol. Environ. Saf. 2017, 143, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Zheng, M.Z.; Li, G.; Sun, G.X.; Shim, H.; Cai, C. Differential toxicity and accumulation of inorganic and methylated arsenic in rice. Plant Soil 2013, 365, 227–238. [Google Scholar] [CrossRef]
- Jia, Y.; Huang, H.; Zhong, M.; Wang, F.H.; Zhang, L.M.; Zhu, Y.G. Microbial arsenic methylation in soil and rice rhizosphere. Environ. Sci. Technol. 2013, 47, 3141–3148. [Google Scholar] [CrossRef]
- Zhang, J.; Wysocki, R.; Li, F.B.; Yu, M.; Martinoia, E.; Song, W.Y. Role of ubiquitination in arsenic tolerance in plants. Trends Plant Sci. 2023. [Google Scholar] [CrossRef]
- Hwang, S.; Chapagain, S.; Lee, J.W.; Han, A.R.; Jang, C.S. Genome-wide transcriptome profiling of genes associated with arsenate toxicity in an arsenic-tolerant rice mutant. Plant Physiol. Biochem. 2017, 120, 40–51. [Google Scholar] [CrossRef]
- Yu, L.J.; Luo, Y.F.; Liao, B.; Xie, L.J.; Chen, L.; Xiao, S.; Li, J.T.; Hu, S.N.; Shu, W.S. Comparative transcriptome analysis of transporters, phytohormone and lipid metabolism pathways in response to arsenic stress in rice (Oryza sativa). New Phytol. 2012, 195, 97–112. [Google Scholar] [CrossRef]
- Sharma, D.; Tiwari, M.; Lakhwani, D.; Tripathi, R.D.; Trivedi, P.K. Differential expression of microRNAs by arsenate and arsenite stress in natural accessions of rice. Metallomics 2015, 7, 174–187. [Google Scholar] [CrossRef]
- Liu, Q.P.; Hu, H.C.; Zhu, L.Y.; Li, R.C.; Feng, Y.; Zhang, L.Q.; Yang, Y.Y.; Liu, X.Q.; Zhang, H.M. Involvement of miR528 in the regulation of arsenite tolerance in rice (Oryza sativa L.). J. Agric. Food Chem. 2015, 63, 8849–8861. [Google Scholar] [CrossRef] [PubMed]
- Dametto, A.; Buffon, G.; Dos Reis Blasi, É.A.; Sperotto, R.A. Ubiquitination pathway as a target to develop abiotic stress tolerance in rice. Plant Signal. Behav. 2015, 10, e1057369. [Google Scholar] [CrossRef] [Green Version]
- Jochem, M.; Ende, L.; Isasa, M.; Ang, J.; Schnell, H.; Guerra-Moreno, A.; Micoogullari, Y.; Bhanu, M.; Gygi, S.P.; Hanna, J. Targeted degradation of glucose transporters protects against arsenic toxicity. Mol. Cell. Biol. 2019, 39, e00559-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, R.T.; Menezes, R.A.; Rodrigues-Pousada, C. E4-Ubiquitin ligase Ufd2 stabilizes Yap8 and modulates arsenic stress responses independent of the U-box motif. Biol. Open 2015, 4, 1122–1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.; Park, H.M.; Han, A.R.; Jang, C.S. Molecular characterization of Oryza sativa arsenic-induced RING E3 ligase 1 (OsAIR1): Uxpression patterns, localization, functional interaction, and heterogeneous overexpression. J. Plant Physiol. 2016, 191, 140–148. [Google Scholar] [CrossRef]
- Lim, S.D.; Hwang, J.G.; Han, A.R.; Park, Y.C.; Lee, C.; Ok, Y.S.; Jang, C.S. Positive regulation of rice RING E3 ligase OsHIR1 in arsenic and cadmium uptakes. Plant Mol. Biol. 2014, 85, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Ying, Y.; Wang, C.; Zhao, Y.; Dong, C.; Whelan, J.; Shou, H. OsNLA1, a RING-type ubiquitin ligase, maintains phosphate homeostasis in Oryza sativa via degradation of phosphate transporters. Plant J. Cell Mol. Biol. 2017, 90, 1040–1051. [Google Scholar] [CrossRef] [Green Version]
- Batista, B.L.; Nigar, M.; Mestrot, A.; Rocha, B.A.; Barbosa Júnior, F.; Price, A.H.; Raab, A.; Feldmann, J. Identification and quantification of phytochelatins in roots of rice to long-term exposure: Evidence of individual role on arsenic accumulation and translocation. J. Exp. Bot. 2014, 65, 1467–1479. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, S.; Kuramata, M.; Abe, T.; Takagi, H.; Ozawa, K.; Ishikawa, S. Phytochelatin synthase OsPCS1 plays a crucial role in reducing arsenic levels in rice grains. Plant J. Cell Mol. Biol. 2017, 91, 840–848. [Google Scholar] [CrossRef] [Green Version]
- Dave, R.; Tripathi, R.D.; Dwivedi, S.; Tripathi, P.; Dixit, G.; Sharma, Y.K.; Trivedi, P.K.; Corpas, F.J.; Barroso, J.B.; Chakrabarty, D. Arsenate and arsenite exposure modulate antioxidants and amino acids in contrasting arsenic accumulating rice (Oryza sativa L.) genotypes. J. Hazard. Mater. 2013, 262, 1123–1131. [Google Scholar] [CrossRef]
- Dubey, A.K.; Kumar, N.; Sahu, N.; Verma, P.K.; Chakrabarty, D.; Behera, S.K.; Mallick, S. Response of two rice cultivars differing in their sensitivity towards arsenic, differs in their expression of glutaredoxin and glutathione S transferase genes and antioxidant usage. Ecotoxicol. Environ. Saf. 2016, 124, 393–405. [Google Scholar] [CrossRef]
- Mishra, S.; Dubey, R.S. Inhibition of ribonuclease and protease activities in arsenic exposed rice seedlings: Role of proline as enzyme protectant. J. Plant Physiol. 2006, 163, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.L.; Wang, T.; Chen, Z.R.; Tang, Z.; Wu, Z.C.; Salt, D.E.; Chao, D.Y.; Zhao, F.J. OsHAC1;1 and OsHAC1;2 function as arsenate reductases and regulate arsenic accumulation. Plant Physiol. 2016, 172, 1708–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, D.Y.; Chen, Y.; Chen, J.G.; Shi, S.L.; Chen, R.Z.; Wang, C.C.; Danku, J.M.; Zhao, F.J.; Salt, D.E. Genome-wide association mapping identifies a new arsenate reductase enzyme critical for limiting arsenic accumulation in plants. PLoS Biol. 2014, 12, e1002009. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.M.; Shi, S.L.; Wang, L.; Tang, Z.; Lv, T.T.; Zhu, X.L.; Ding, X.M.; Wang, Y.F.; Zhao, F.J.; Wu, Z.C. OsHAC4 is critical for arsenate tolerance and regulates arsenic accumulation in rice. New Phytol. 2017, 215, 1090–1101. [Google Scholar] [CrossRef] [Green Version]
- Duan, G.L.; Zhou, Y.; Tong, Y.P.; Mukhopadhyay, R.; Rosen, B.P.; Zhu, Y.G. A CDC25 homologue from rice functions as an arsenate reductase. New Phytol. 2007, 174, 311–321. [Google Scholar] [CrossRef]
- Yamazaki, S.; Ueda, Y.; Mukai, A.; Ochiai, K.; Matoh, T. Rice phytochelatin synthases OsPCS1 and OsPCS2 make different contributions to cadmium and arsenic tolerance. Plant Direct 2018, 2, e00034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uraguchi, S.; Tanaka, N.; Hofmann, C.; Abiko, K.; Ohkama-Ohtsu, N.; Weber, M.; Kamiya, T.; Sone, Y.; Nakamura, R.; Takanezawa, Y.; et al. Phytochelatin synthase has contrasting effects on cadmium and arsenic accumulation in rice grains. Plant Cell Physiol. 2017, 58, 1730–1742. [Google Scholar] [CrossRef] [Green Version]
- Das, N.; Bhattacharya, S.; Bhattacharyya, S.; Maiti, M.K. Identification of alternatively spliced transcripts of rice phytochelatin synthase 2 gene OsPCS2 involved in mitigation of cadmium and arsenic stresses. Plant Mol. Biol. 2017, 94, 167–183. [Google Scholar] [CrossRef]
- Yang, J.; Gao, M.X.; Hu, H.; Ding, X.M.; Lin, H.W.; Wang, L.; Xu, J.M.; Mao, C.Z.; Zhao, F.J.; Wu, Z.C. OsCLT1, a CRT-like transporter 1, is required for glutathione homeostasis and arsenic tolerance in rice. New Phytol. 2016, 211, 658–670. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, M.; Kidwai, M.; Dutta, P.; Narayan, S.; Gautam, N.; Chawda, K.; Shirke, P.A.; Mishra, A.K.; Chakrabarty, D. A tau class glutathione-S-transferase (OsGSTU5) confers tolerance against arsenic toxicity in rice by accumulating more arsenic in root. J. Hazard. Mater. 2022, 426, 128100. [Google Scholar] [CrossRef]
- Kumar, S.; Khare, R.; Trivedi, P.K. Arsenic-responsive high-affinity rice sulphate transporter, OsSultr1;1, provides abiotic stress tolerance under limiting sulphur condition. J. Hazard. Mater. 2019, 373, 753–762. [Google Scholar] [CrossRef]
- Manuka, R.; Saddhe, A.A.; Srivastava, A.K.; Kumar, K.; Penna, S. Overexpression of rice OsWNK9 promotes arsenite tolerance in transgenic Arabidopsis plants. J. Biotechnol. 2021, 332, 114–125. [Google Scholar] [CrossRef]
- Kidwai, M.; Dhar, Y.V.; Gautam, N.; Tiwari, M.; Ahmad, I.Z.; Asif, M.H.; Chakrabarty, D. Oryza sativa class III peroxidase (OsPRX38) overexpression in Arabidopsis thaliana reduces arsenic accumulation due to apoplastic lignification. J. Hazard. Mater. 2019, 362, 383–393. [Google Scholar] [CrossRef]
- Wang, C.C.; Zheng, L.H.; Tang, Z.; Sun, S.K.; Ma, J.F.; Huang, X.Y.; Zhao, F.J. OASTL-A1 functions as a cytosolic cysteine synthase and affects arsenic tolerance in rice. J. Exp. Bot. 2020, 71, 3678–3689. [Google Scholar] [CrossRef] [PubMed]
- Shri, M.; Dave, R.; Diwedi, S.; Shukla, D.; Kesari, R.; Tripathi, R.D.; Trivedi, P.K.; Chakrabarty, D. Heterologous expression of Ceratophyllum demersum phytochelatin synthase, CdPCS1, in rice leads to lower arsenic accumulation in grain. Sci. Rep. 2014, 4, 5784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, M.K.; Majumdar, A.; Srivastava, A.K.; Bose, S.; Suprasanna, P.; Srivastava, S. Antioxidant enzymes and transporter genes mediate arsenic stress reduction in rice (Oryza sativa L.) upon thiourea supplementation. Chemosphere 2022, 292, 133482. [Google Scholar] [CrossRef] [PubMed]
- Mridha, D.; Paul, I.; De, A.; Ray, I.; Das, A.; Joardar, M.; Chowdhury, N.R.; Bhadoria, P.B.S.; Roychowdhury, T. Rice seed (IR64) priming with potassium humate for improvement of seed germination, seedling growth and antioxidant defense system under arsenic stress. Ecotoxicol. Environ. Saf. 2021, 219, 112313. [Google Scholar] [CrossRef]
- Yadav, P.; Srivastava, S. Effect of thiourea application on root, old leaf and young leaf of two contrasting rice varieties (Oryza sativa L.) grown in arsenic contaminated soil. Environ. Technol. Innov. 2021, 21, 101368. [Google Scholar] [CrossRef]
- Chakrabarty, D.; Trivedi, P.K.; Misra, P.; Tiwari, M.; Shri, M.; Shukla, D.; Kumar, S.; Rai, A.; Pandey, A.; Nigam, D.; et al. Comparative transcriptome analysis of arsenate and arsenite stresses in rice seedlings. Chemosphere 2009, 74, 688–702. [Google Scholar] [CrossRef]
- Shri, M.; Singh, P.K.; Kidwai, M.; Gautam, N.; Dubey, S.; Verma, G.; Chakrabarty, D. Recent advances in arsenic metabolism in plants: Current status, challenges and highlighted biotechnological intervention to reduce grain arsenic in rice. Metallomics 2019, 11, 519–532. [Google Scholar] [CrossRef]
- Jung, H.I.; Kong, M.S.; Lee, B.R.; Kim, T.H.; Chae, M.J.; Lee, E.J.; Jung, G.B.; Lee, C.H.; Sung, J.K.; Kim, Y.H. Exogenous glutathione increases arsenic translocation into shoots and alleviates arsenic-induced oxidative stress by sustaining ascorbate-glutathione homeostasis in rice seedlings. Front. Plant Sci. 2019, 10, 1089. [Google Scholar] [CrossRef]
- Choudhury, B.; Chowdhury, S.; Biswas, A. Regulation of growth and metabolism in rice (Oryza sativa L.) by arsenic and its possible reversal by phosphate. J. Plant Interact. 2011, 6, 15–24. [Google Scholar] [CrossRef]
- Chandrakar, V.; Yadu, B.; Meena, R.K.; Dubey, A.; Keshavkant, S. Arsenic-induced genotoxic responses and their amelioration by diphenylene iodonium, 24-epibrassinolide and proline in Glycine max L. Plant Physiol. Biochem. 2017, 112, 74–86. [Google Scholar] [CrossRef]
- Asgher, M.; Sehar, Z.; Rehaman, A.; Rashid, S.; Ahmed, S.; Per, T.S.; Alyemeni, M.N.; Khan, N.A. Exogenously-applied L-glutamic acid protects photosynthetic functions and enhances arsenic tolerance through increased nitrogen assimilation and antioxidant capacity in rice (Oryza sativa L.). Environ. Pollut. 2022, 301, 119008. [Google Scholar] [CrossRef]
- Huang, S.H.; Yang, X.L.; Chen, G.; Wang, X. Application of glutamic acid improved As tolerance in aromatic rice at early growth stage. Chemosphere 2023, 322, 138173. [Google Scholar] [CrossRef]
- Begara Morales, J.; Sánchez-Calvo, B.; Chaki, M.; Valderrama, R.; Mata-Pérez, C.; Padilla, M.; Corpas, F.; Barroso, J. Antioxidant systems are regulated by nitric oxide-mediated post-translational modifications (NO-PTMs). Front. Plant Sci. 2016, 7, 152. [Google Scholar] [CrossRef] [Green Version]
- Asgher, M.; Per, T.S.; Masood, A.; Fatma, M.; Freschi, L.; Corpas, F.J.; Khan, N.A. Nitric oxide signaling and its crosstalk with other plant growth regulators in plant responses to abiotic stress. Environ. Sci. Pollut. Res. Int. 2017, 24, 2273–2285. [Google Scholar] [CrossRef]
- Sami, F.; Faizan, M.; Faraz, A.; Siddiqui, H.; Yusuf, M.; Hayat, S. Nitric oxide-mediated integrative alterations in plant metabolism to confer abiotic stress tolerance, NO crosstalk with phytohormones and NO-mediated post translational modifications in modulating diverse plant stress. Nitric Oxide 2018, 73, 22–38. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.J.; Zhang, M.L.; Wei, S.H.; Zhang, J.; Wang, C.L.; Liao, W.B. Roles of nitric oxide in heavy metal stress in plants: Cross-talk with phytohormones and protein S-nitrosylation. Environ. Pollut. 2020, 259, 113943. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.K.; Indoliya, Y.; Chauhan, A.S.; Singh, S.P.; Singh, A.P.; Dwivedi, S.; Tripathi, R.D.; Chakrabarty, D. Nitric oxide mediated transcriptional modulation enhances plant adaptive responses to arsenic stress. Sci. Rep. 2017, 7, 3592. [Google Scholar] [CrossRef] [PubMed]
- Praveen, A.; Gupta, M. Nitric oxide confronts arsenic stimulated oxidative stress and root architecture through distinct gene expression of auxin transporters, nutrient related genes and modulates biochemical responses in Oryza sativa L. Environ. Pollut. 2018, 240, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Mostofa, M.; Alam, M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Calcium mitigates arsenic toxicity in rice seedlings by reducing arsenic uptake and modulating the antioxidant defense and glyoxalase systems and stress markers. BioMed Res. Int. 2015, 2015, 340812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, G.; Srivastava, D.; Narayan, S.; Shirke, P.A.; Chakrabarty, D. Exogenous application of methyl jasmonate alleviates arsenic toxicity by modulating its uptake and translocation in rice (Oryza sativa L.). Ecotoxicol. Environ. Saf. 2020, 201, 110735. [Google Scholar] [CrossRef]
- Rivas-San, V.M.; Plasencia, J. Salicylic acid beyond defence: Its role in plant growth and development. J. Exp. Bot. 2011, 62, 3321–3338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.P.; Dixit, G.; Kumar, A.; Mishra, S.; Kumar, N.; Dixit, S.; Singh, P.K.; Dwivedi, S.; Trivedi, P.K.; Pandey, V.; et al. A protective role for nitric oxide and salicylic acid for arsenite phytotoxicity in rice (Oryza sativa L.). Plant Physiol. Biochem. 2017, 115, 163–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.P.; Dixit, G.; Mishra, S.; Dwivedi, S.; Tiwari, M.; Mallick, S.; Pandey, V.; Trivedi, P.K.; Chakrabarty, D.; Tripathi, R.D. Salicylic acid modulates arsenic toxicity by reducing its root to shoot translocation in rice (Oryza sativa L.). Front. Plant Sci. 2015, 6, 340. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.I.R.; Jahan, B.; AlAjmi, M.F.; Rehman, M.T.; Iqbal, N.; Irfan, M.; Sehar, Z.; Khan, N.A. Crosstalk of plant growth regulators protects photosynthetic performance from arsenic damage by modulating defense systems in rice. Ecotoxicol. Environ. Saf. 2021, 222, 112535. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Rahman, M.M.; Nguyen, K.H.; Li, W.; Watanabe, Y.; Tran, C.D.; Zhang, M.; Itouga, M.; Fujita, M.; Tran, L.P. Strigolactones regulate arsenate uptake, vacuolar-sequestration and antioxidant defense responses to resist arsenic toxicity in rice roots. J. Hazard. Mater. 2021, 415, 125589. [Google Scholar] [CrossRef]
- Asgher, M.; Ahmed, S.; Sehar, Z.; Gautam, H.; Gandhi, S.G.; Khan, N.A. Hydrogen peroxide modulates activity and expression of antioxidant enzymes and protects photosynthetic activity from arsenic damage in rice (Oryza sativa L.). J. Hazard. Mater. 2021, 401, 123365. [Google Scholar] [CrossRef]
- Choudhury, B.; Mitra, S.; Biswas, A.K. Regulation of sugar metabolism in rice (Oryza sativa L.) seedlings under arsenate toxicity and its improvement by phosphate. Physiol. Mol. Biol. Plants 2010, 16, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, P.; Samal, A.C.; Majumdar, J.; Banerjee, S.; Santra, S.C. In vitro assessment on the impact of soil arsenic in the eight rice varieties of west Bengal, India. J. Hazard. Mater. 2013, 262, 1091–1097. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene Name | Gene ID | Main Expression Tissues or Organs | Subcellular Localization | Function | Ref. |
|---|---|---|---|---|---|
| OsPht1;1 (OsPT1) | LOC_Os03g05620 | Root, shoot | Plasma membrane | Involved in As(V) uptake from the soil or apoplast and involved in root-to-shoot As(V) transport | [39] |
| OsPht1;4 (OsPT4) | LOC_Os09g37200 | Root, shoot | Plasma membrane | Takes up and transports As(V) from the root to the shoot | [40] |
| OsPht1;8 (OsPT8) | LOC_Os10g30790 | Root, shoot | Plasma membrane | Transports As(V) into the root or xylem | [41,42] |
| OsPHF1 | LOC_Os07g09000 | Root | Plasma membrane | Regulates OsPT8 for the uptake and transport of As(V) | [41] |
| OsPHR2 | LOC_Os07g25710 | Root, shoot | Plasma membrane | Regulates phosphate transporters to affect As(V) transport to the root and xylem | [41] |
| OsNLA1 | LOC_Os07g47590 | Roots, shoot | Plasma membrane | Regulates As(V) uptake and tolerance mainly via regulating the amount of phosphate transporters | [43] |
| OsNIP1;1 | LOC_Os02g13870 | Root | Plasma membrane | Restricts As(III) loading into the xylem by providing a route for As exudation from the stele | [44] |
| OsNIP2;1 (OsLsi1) | LOC_Os02g51110 | Root, leaf, anther | Plasma membrane | Translocates organic species of As into the root, shoot, xylem/translocates As(III) into the root | [45,46,47] |
| OsLsi2 | LOC_Os03g01700 | Root | Plasma membrane | Involved in As(III) transport out of the root cells toward the stele | [45] |
| OsNIP2;2 (Os Lsi6) | LOC_Os06g12310 | Root, leaf, shoot | Plasma membrane | Weak capacity to absorb As(III) in rice | [45] |
| OsNIP3;1 | LOC_Os10g36924 | Root, leaf, shoot, anther | Membrane | Weak capacity to absorb As(III) in rice | [45] |
| OsNIP3;2 | LOC_Os08g05590 | Leaf, root, anther | Plasma membrane | Transports As(III) into the root | [48] |
| OsNIP3;3 | LOC_Os08g05600 | Root | Plasma membrane | Restricts As(III) loading into the xylem by providing a route for As exudation from the stele | [44] |
| OsPIP2;4 | LOC_Os07g26630 | Root | Plasma membrane | Plays a role in the permeability of As(III) in Arabidopsis | [49] |
| OsPIP2;6 | LOC_Os04g16450 | Root, leaf, shoot, anther | Plasma membrane | Transports As(III) into oocytes, and effluxes As(III) from the root of Arabidopsis | [49] |
| OsPIP2;7 | LOC_Os09g36930 | Root, leaf, anther | Plasma membrane | Plays a role in the permeability of As(III) in Arabidopsis | [49] |
| OsGrx_C7 | LOC_Os01g27140 | Root, shoot | / | Alters expression of As(III) transporters (aquaporins) in rice and Arabidopsis | [50,51] |
| OsABCC1 | LOC_Os04g52900 | Root, shoot, grain, leaf, nodes, peduncle, rachis | Tonoplast | Limits As transport to the grains by sequestering As in the vacuoles | [52] |
| OsABCC7 | LOC_Os04g49900 | Xylem parenchyma cells in the stele region of the primary and lateral roots | Plasma membrane | Involved in the translocation of As from the roots to the shoots in rice, likely by mediating the efflux of As(III)-thiol complexes for xylem loading | [53] |
| OsNRAMP1 | LOC_Os07g15460 | Root | Plasma membrane | Assists As(III) xylem loading for root-to-shoot mobilization in Arabidopsis | [54] |
| OsPTR7 | LOC_Os01g04950 | Root, shoot, leaf, nodes | Plasma membrane | Transports DMA to xylem, phloem, and grain | [55] |
| OsWRKY28 | LOC_Os06g44010 | Strong expression in the root tip, lateral root, and reproductive organs | Nuclei | Regulates As(V) accumulation in the shoot | [56] |
| OsADH2 | LOC_Os11g10510 | Root | / | Regulates silicate transporters to influence As(III) contents in aerial tissues of rice | [57] |
| OsMATE2 | LOC_Os05g48040 | Root, leaf, seed | / | Modulates As accumulation in rice grain | [58] |
| OsARM1 | LOC_Os05g37060 | Phloem of vascular bundles in basal and upper nodes | Nucleus | Regulates As(III) uptake and root-to-shoot translocation | [59] |
| Gene Name | Gene ID | Main Expression Tissues or Organs | Subcellular Localization | Function | Ref. |
|---|---|---|---|---|---|
| OsHAC1;1 | LOC_Os02g01220 | Root, shoot | Cytoplasm, nucleus | Reduces As(V) to As(III) | [113,114] |
| OsHAC1;2 | LOC_Os04g17660 | Root | Cytoplasm, nucleus | Reduces As(V) to As(III) | [113] |
| OsHAC4 | LOC_Os02g06290 | Root | Cytoplasm, nucleus | Reduces As(V) to As(III) | [115] |
| OsACR2.1 | LOC_Os10g39860 | Root, shoot | Cytoplasm, nucleus | Reduces As(V) to As(III) | [116] |
| OsPCS1 | LOC_Os05g34290 | Root, shoot, leaf, grain | Cytosol | Catalyzes the formation of phytochelatins that form As(III)-phytochelatin conjugates | [117,118] |
| OsPCS2 | LOC_Os06g01260 | Root, shoot, leaf, grain | Cytosol | Catalyzes the formation of phytochelatins that form As(III)-phytochelatin conjugates | [117,118,119] |
| OsGrx_C2.1 | LOC_Os02g40500 | Root, shoot | / | Involved in redox regulation and protection under oxidative stress/alters the transcripts of AtNIPs in Arabidopsis | [50] |
| OsCLT1 | LOC_Os01g72570 | Root | Envelope membrane of plastids | Maintains glutathione (GSH) homeostasis probably by mediating the export of γ-glutamylcysteine and GSH from the plastids to the cytoplasm, thereby affecting As detoxification in rice | [120] |
| OsGSTU5 | LOC_Os09g20220 | Root | Cytoplasm | Chelates As with GSH and sequesters it into the root cells vacuoles | [121] |
| OsSultr1;1 | LOC_Os03g09970 | / | / | Maintains ROS homeostasis, promotes the chelation of As with GSH and its sequestration into the root-cell vacuoles, and limits the acropetal translocation of As towards the shoot | [122] |
| OsWNK9 | LOC_Os12g06490 | Plant tissues | Nucleus | Involved in the regulation of arsenite-induced oxidative stress management by activating the antioxidant system and osmotic adjustment processes | [123] |
| OsPRX38 | LOC_Os03g13210 | Root | Apoplast | Enhances As(III) and As(V) tolerance by increasing ROS detoxification/reduces As accumulation due to high lignification | [124] |
| OsOASTL-A1 | LOC_Os03g53650 | Root | Cytosol | Plays an important role in non-protein thiol biosynthesis in roots for As detoxification | [125] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, A.; Lian, W.; Wang, X.; Chen, G. Regulatory Mechanisms Underlying Arsenic Uptake, Transport, and Detoxification in Rice. Int. J. Mol. Sci. 2023, 24, 11031. https://doi.org/10.3390/ijms241311031
Geng A, Lian W, Wang X, Chen G. Regulatory Mechanisms Underlying Arsenic Uptake, Transport, and Detoxification in Rice. International Journal of Molecular Sciences. 2023; 24(13):11031. https://doi.org/10.3390/ijms241311031
Chicago/Turabian StyleGeng, Anjing, Wenli Lian, Xu Wang, and Guang Chen. 2023. "Regulatory Mechanisms Underlying Arsenic Uptake, Transport, and Detoxification in Rice" International Journal of Molecular Sciences 24, no. 13: 11031. https://doi.org/10.3390/ijms241311031