
Nicotianamine: A Key Player in Metal Homeostasis and Hyperaccumulation in Plants

Abstract
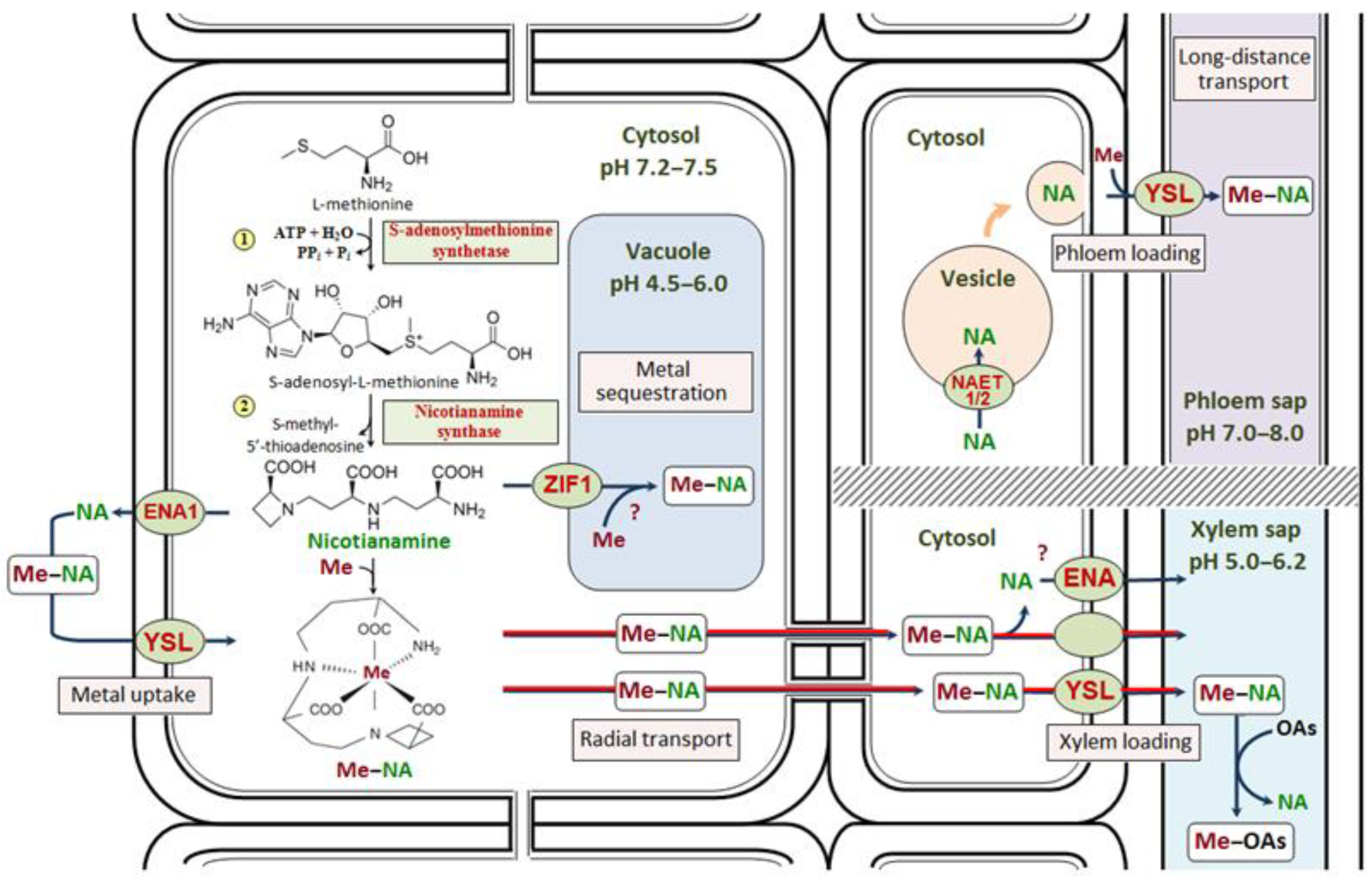
:1. Introduction
2. Formation of Metal–Nicotianamine Complexes
3. Biosynthesis and Accumulation of Nicotianamine in Plants
3.1. Biosynthesis of Nicotianamine
3.2. The Expression of NAS Genes in Plant Organs
3.3. The Expression of NAS Genes in Plant Tissues
3.4. Regulation of NA Biosynthesis
3.5. Nicotianamine Accumulation in Metal Hyperaccumulators and Excluders
3.6. Localization of NAS Proteins in Metal Hyperaccumulators and Excluders
3.7. Nicotianamine as a Precursor of Phytosiderophores in Cereals
4. Transport and Physiological Role of Nicotianamine in Plants
4.1. The Role of Yellow Stripe-like Transporters in Metal Transport in Plants
4.2. Secretion of Nicotianamine
4.3. Transport of Nicotianamine into the Vacuole
4.4. Participation of Nicotianamine in Metal Radial Transport in Roots
4.5. Nicotianamine-Dependent Long-Distance Transport of Metals
4.6. Nicotianamine-Dependent Metal Transport in Shoots
4.7. The Role of Nicotianamine in Nitrogen Fixation in Legumes
4.8. Nicotianamine and Metal Tolerance
5. Conclusions, Perspectives and Outlook
5.1. Theoretical Importance
5.2. Practical Importance
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brooks, R.R.; Lee, J.; Reeves, R.D.; Jaffré, T. Detection of nickeliferous rocks by analysis of herbarium specimens of indicator plants. J. Geochem. Explor. 1977, 7, 49–57. [Google Scholar] [CrossRef]
- Jaffré, T.; Pillon, Y.; Thomine, S.; Merlot, S. The metal hyperaccumulators from New Caledonia can broaden our understanding of nickel accumulation in plants. Front. Plant Sci. 2013, 4, 279. [Google Scholar] [CrossRef] [Green Version]
- van der Ent, A.; Vinya, R.; Erskine, P.D.; Malaisse, F.; Przybyłowicz, W.J.; Barnabas, A.D.; Harris, H.H.; Mesjasz-Przybyłowicz, J. Elemental distribution and chemical speciation of copper and cobalt in three metallophytes from the copper-cobalt belt in Northern Zambia. Metallomics 2020, 12, 682–701. [Google Scholar] [CrossRef]
- Verbruggen, N.; Hermans, C.; Schat, H. Molecular mechanisms of metal hyperaccumulation in plants. New Phytol. 2009, 181, 759–776. [Google Scholar] [CrossRef]
- Krämer, U. Metal hyperaccumulation in plants. Annu. Rev. Plant Biol. 2010, 61, 517–534. [Google Scholar] [CrossRef]
- Reeves, R.D.; Baker, A.J.; Jaffré, T.; Erskine, P.D.; Echevarria, G.; van der Ent, A. A global database for plants that hyperaccumulate metal and metalloid trace elements. New Phytol. 2018, 218, 407–411. [Google Scholar] [CrossRef] [Green Version]
- Manara, A.; Fasani, E.; Furini, A.; DalCorso, G. Evolution of the metal hyperaccumulation and hypertolerance traits. Plant Cell Environ. 2020, 43, 2969–2986. [Google Scholar] [CrossRef]
- Padmavathiamma, P.K.; Li, L.Y. Phytoremediation technology: Hyper-accumulation metals in plants. Water Air Soil Pollut. 2007, 184, 105–126. [Google Scholar] [CrossRef]
- van der Ent, A.; Baker, A.J.M.; Reeves, R.D.; Chaney, R.L.; Anderson, C.W.N.; Meech, J.A.; Erskine, P.D.; Simonnot, M.O.; Vaughan, J.; Morel, J.L.; et al. Agromining: Farming for metals in the future? Environ. Sci. Technol. 2015, 49, 4773–4780. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Physiological role of nickel and its toxic effects on higher plants. Russ. J. Plant Physiol. 2006, 53, 257–277. [Google Scholar] [CrossRef]
- Jeong, J.; Guerinot, M.L. Homing in on iron homeostasis in plants. Trends Plant Sci. 2009, 14, 280–285. [Google Scholar] [CrossRef]
- Shahzad, B.; Tanveer, M.; Rehman, A.; Cheema, S.A.; Fahad, S.; Rehman, S.; Sharma, A. Nickel; whether toxic or essential for plants and environment—A review. Plant Physiol. Biochem. 2018, 132, 641–651. [Google Scholar] [CrossRef]
- Ghori, N.-H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy metal stress and responses in plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- Angulo-Bejarano, P.I.; Puente-Rivera, J.; Cruz-Ortega, R. Metal and metalloid toxicity in plants: An overview on molecular aspects. Plants 2021, 10, 635. [Google Scholar] [CrossRef]
- Kaur, H.; Garg, N. Zinc toxicity in plants: A review. Planta 2021, 253, 129. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, V.B.; Zhukovskaya, N.V. Effect of heavy metals on root growth and the use of roots as test objects. Russ. J. Plant Physiol. 2021, 68, S1–S25. [Google Scholar] [CrossRef]
- Pourret, O.; Hursthouse, A. It’s time to replace the term “heavy metals” with “potentially toxic elements” when reporting environmental research. Int. J. Environ. Res. Public Health 2019, 16, 4446. [Google Scholar] [CrossRef] [Green Version]
- Gonneau, C.; Genevois, N.; Frérot, H.; Sirguey, C.; Sterckeman, T. Variation of trace metal accumulation, major nutrient uptake and growth parameters and their correlations in 22 populations of Noccaea caerulescens. Plant Soil 2014, 384, 271–287. [Google Scholar] [CrossRef]
- Stein, R.J.; Höreth, S.; de Melo, J.R.F.; Syllwasschy, L.; Lee, G.; Garbin, M.L.; Clemens, S.; Krämer, U. Relationships between soil and leaf mineral composition are element-specific, environment-dependent and geographically structured in the emerging model Arabidopsis halleri. New Phytol. 2017, 213, 1274–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sterckeman, T.; Cazes, Y.; Gonneau, C.; Sirguey, C. Phenotyping 60 populations of Noccaea caerulescens provides a broader knowledge of variation in traits of interest for phytoextraction. Plant Soil 2017, 418, 523–540. [Google Scholar] [CrossRef]
- Corso, M.; Schvartzman, M.S.; Guzzo, F.; Souard, F.; Malkowski, E.; Hanikenne, M.; Verbruggen, N. Contrasting cadmium resistance strategies in two metallicolous populations of Arabidopsis halleri. New Phytol. 2018, 218, 283–297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozhevnikova, A.D.; Seregin, I.V.; Aarts, M.G.M.; Schat, H. Intra-specific variation in zinc, cadmium and nickel hypertolerance and hyperaccumulation capacities in Noccaea caerulescens. Plant Soil 2020, 452, 479–498. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D.; Schat, H. Correlated variation of the Zn accumulation and tolerance capacities among populations and ecotypes of the Zn hyperaccumulator, Noccaea caerulescens. Russ. J. Plant Physiol. 2021, 68, S26–S36. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D.; Schat, H. Nickel tolerance and accumulation capacities in different populations of the hyperaccumulator Noccaea caerulescens. Russ. J. Plant Physiol. 2022, 69, 70. [Google Scholar] [CrossRef]
- Salt, D.E.; Baxter, I.; Lahner, B. Ionomics and the study of the plant ionome. Annu. Rev. Plant Biol. 2008, 59, 709–733. [Google Scholar] [CrossRef] [Green Version]
- Baxter, I.; Dilkes, B.P. Elemental profiles reflect plant adaptations to the environment. Science 2012, 336, 1661–1663. [Google Scholar] [CrossRef] [Green Version]
- Hanikenne, M.; Nouet, C. Metal hyperaccumulation and hypertolerance: A model for plant evolutionary genomics. Curr. Opin. Plant Biol. 2011, 14, 252–259. [Google Scholar] [CrossRef]
- Corso, M.; García de la Torre, V.S. Biomolecular approaches to understanding metal tolerance and hyperaccumulation in plants. Metallomics 2020, 12, 840–859. [Google Scholar] [CrossRef]
- Merlot, S.; de la Torre, V.S.; Hanikenne, M. Physiology and molecular biology of trace element hyperaccumulation. In Agromining: Farming for Metals. Mineral Resource Reviews; van der Ent, A., Baker, A.J., Echevarria, G., Simonnot, M.O., Morel, J.L., Eds.; Springer: Cham, Switzerland, 2021; pp. 155–181. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Low-molecular-weight ligands in plants: Role in metal homeostasis and hyperaccumulation. Photosynth. Res. 2021, 150, 51–96. [Google Scholar] [CrossRef]
- Clemens, S. Metal ligands in micronutrient acquisition and homeostasis. Plant Cell Environ. 2019, 42, 2902–2912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zlobin, I.E.; Kartashov, A.V.; Nosov, A.V.; Fomenkov, A.A.; Kuznetsov, V.V. The labile zinc pool in plant cells. Funct. Plant Biol. 2019, 46, 796–805. [Google Scholar] [CrossRef]
- Callahan, D.L.; Baker, A.J.M.; Kolev, S.D.; Wedd, A.G. Metal ion ligands in hyperaccumulating plants. J. Biol. Inorg. Chem. 2006, 11, 2–12. [Google Scholar] [CrossRef]
- Haydon, M.J.; Cobbett, C.S. Transporters of ligands for essential metal ions in plants. New Phytol. 2007, 174, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Curie, C.; Cassin, G.; Couch, D.; Divol, F.; Higuchi, K.; Le Jean, M.; Misson, J.; Schikora, A.; Czernic, P.; Mari, S. Metal movement within the plant: Contribution of nicotianamine and yellow stripe 1-like transporters. Ann. Bot. 2009, 103, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Clemens, S.; Deinlein, U.; Ahmadi, H.; Höreth, S.; Uraguchi, S. Nicotianamine is a major player in plant Zn homeostasis. BioMetals 2013, 26, 623–632. [Google Scholar] [CrossRef]
- Kobayashi, T.; Nozoye, T.; Nishizawa, N.K. Iron transport and its regulation in plants. Free Radic. Biol. Med. 2019, 133, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Noma, M.; Noguchi, M.; Tamaki, E. A new amino acid, nicotianamine, from tobacco leaves. Tetrahedron Lett. 1971, 12, 2017–2020. [Google Scholar] [CrossRef]
- Kristensen, I.B.; Larsen, P.O. Azetidine-2-carboxylic acid derivatives from seeds of Fagus silvatica L. and a revised structure for nicotianamine. Phytochemistry 1974, 13, 2791–2798. [Google Scholar] [CrossRef]
- Laffont, C.; Arnoux, P. The ancient roots of nicotianamine: Diversity, role, regulation and evolution of nicotianamine-like metallophores. Metallomics 2020, 12, 1480–1493. [Google Scholar] [CrossRef]
- Buděšínský, M.; Budzikiewicz, H.; Procházka, Ž.; Ripperger, H.; Römer, A.; Scholz, G.; Schreiber, K. Nicotianamine, a possible phytosiderophore of general occurrence. Phytochemistry 1980, 19, 2295–2297. [Google Scholar] [CrossRef]
- Procházka, Ž.; Scholz, G. Nicotianamine, the ‘normalizing factor’ for the auxotroph tomato mutant Chloronerva; a representative of a new class of plant effectors. Experientia 1984, 40, 794–801. [Google Scholar] [CrossRef]
- Scholz, G.; Schlesier, G.; Seifert, K. Effect of nicotianamine on iron uptake by the tomato mutant ‘chloronerva’. Physiol. Plant. 1985, 63, 99–104. [Google Scholar] [CrossRef]
- Rudolph, A.; Becker, R.; Scholz, G.; Procházka, Z.; Toman, J.; Macek, T.; Herout, V. The occurrence of the amino acid nicotianamine in plants and microorganisms. A reinvestigation. Biochem. Physiol. Pflanzen 1985, 180, 557–563. [Google Scholar] [CrossRef]
- Stephan, U.W.; Scholz, G. Nicotianamine: Mediator of transport of iron and heavy metals in the phloem? Physiol. Plant. 1993, 88, 522–529. [Google Scholar] [CrossRef]
- Stephan, U.W.; Schmidke, I.; Stephan, V.W.; Scholz, G. The nicotianamine molecule is made-to-measure for complexation of metal micronutrients in plants. Biometals 1996, 9, 84–90. [Google Scholar] [CrossRef]
- Curie, C.; Briat, J.-F. Iron transport and signaling in plants. Annu. Rev. Plant Biol. 2003, 54, 183–206. [Google Scholar] [CrossRef]
- Vacchina, V.; Mari, S.; Czernic, P.; Marquès, L.; Pianelli, K.; Schaumlöffel, D.; Lebrun, M.; Łobiński, R. Speciation of nickel in a hyperaccumulating plant by high-performance liquid chromatography-inductively coupled plasma mass spectrometry and electrospray MS/MS assisted by cloning using yeast complementation. Anal. Chem. 2003, 75, 2740–2745. [Google Scholar] [CrossRef]
- Mari, S.; Gendre, D.; Pianelli, K.; Ouerdane, L.; Lobinski, R.; Briat, J.-F.; Lebrun, M.; Czernic, P. Root-to-shoot long-distance circulation of nicotianamine and nicotianamine-nickel chelates in the metal hyperaccumulator Thlaspi caerulescens. J. Exp. Bot. 2006, 57, 4111–4122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banakar, R.; Alvarez Fernandez, A.; Zhu, C.; Abadia, J.; Capell, T.; Christou, P. The ratio of phytosiderophores nicotianamine to deoxymugenic acid controls metal homeostasis in rice. Planta 2019, 250, 1339–1354. [Google Scholar] [CrossRef]
- Kińska, K.; Cruzado-Tafur, E.; Parailloux, M.; Torró, L.; Lobinski, R.; Szpunar, J. Speciation of metals in indigenous plants growing in post-mining areas: Dihydroxynicotianamine identified as the most abundant Cu and Zn ligand in Hypericum laricifolium. Sci. Total Environ. 2022, 809, 151090. [Google Scholar] [CrossRef] [PubMed]
- Chao, Z.-F.; Chao, D.-Y. Similarities and differences in iron homeostasis strategies between graminaceous and nongraminaceous plants. New Phytol. 2022, 236, 1655–1660. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Murata, J.; Namba, K. Unraveling the new biological roles and possible applications of phytosiderophores in plants and mammals. Metallomics Res. 2022, 2, rev-1. [Google Scholar]
- Andresen, E.; Peiter, E.; Küpper, H. Trace metal metabolism in plants. J. Exp. Bot. 2018, 69, 909–954. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, J.T.; Borthakur, D. Methods for metal chelation in plant homeostasis: Review. Plant Physiol. Biochem. 2021, 163, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Nozoye, T.; Nagasaka, S.; Kobayashi, T.; Takahashi, M.; Sato, Y.; Sato, Y.; Uozumi, N.; Nakanishi, H.; Nishizawa, N.K. Phytosiderophore efflux transporters are crucial for iron acquisition in graminaceous plants. J. Biol. Chem. 2011, 286, 5446–5454. [Google Scholar] [CrossRef] [Green Version]
- Nozoye, T.; Nakanishi, H.; Nishizawa, N.K. Characterizing the crucial components of iron homeostasis in the maize mutants ys1 and ys3. PLoS ONE 2013, 8, e62567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Xiao, K.; Li, S.; Li, J.; Huang, J.; Chen, R.; Pang, S.; Zhou, X. Genome-wide analysis of the NAAT, DMAS, TOM, and ENA gene families in maize suggests their roles in mediating iron homeostasis. BMC Plant Biol. 2022, 22, 37. [Google Scholar] [CrossRef]
- Suzuki, M.; Nozoye, T.; Nagasaka, S.; Nakanishi, H.; Nishizawa, N.K.; Mori, S. The detection of endogenous 2′-deoxymugineic acid in olives (Olea europaea L.) indicates the biosynthesis of mugineic acid family phytosiderophores in non-graminaceous plants. Soil Sci. Plant Nutr. 2016, 62, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Astolfi, S.; Pii, Y.; Mimmo, T.; Lucini, L.; Miras-Moreno, M.B.; Coppa, E.; Violino, S.; Celletti, S.; Cesco, S. Single and combined Fe and S deficiency differentially modulate root exudate composition in tomato: A double strategy for Fe acquisition? Int. J. Mol. Sci. 2020, 21, 4038. [Google Scholar] [CrossRef]
- Marastoni, L.; Lucini, L.; Miras-Moreno, B.; Trevisan, M.; Sega, D.; Zamboni, A.; Varanini, Z. Changes in physiological activities and root exudation profile of two grapevine rootstocks reveal common and specific strategies for Fe acquisition. Sci. Rep. 2020, 10, 18839. [Google Scholar] [CrossRef]
- Tsednee, M.; Yang, S.-C.; Lee, D.-C.; Yeh, K.-C. Root-secreted nicotianamine from Arabidopsis halleri facilitates zinc hypertolerance by regulating zinc bioavailability. Plant Physiol. 2014, 166, 839–852. [Google Scholar] [CrossRef] [Green Version]
- Beneš, I.; Schreiber, K.; Ripperger, H.; Kircheiss, A. Metal complex formation by nicotianamine, a possible phytosiderophore. Experientia 1983, 39, 261–262. [Google Scholar] [CrossRef]
- Blindauer, C.A.; Schmid, R. Cytosolic metal handling in plants: Determinants for zinc specificity in metal transporters and metallothioneins. Metallomics 2010, 2, 510–529. [Google Scholar] [CrossRef]
- von Wirén, N.; Klair, S.; Bansal, S.; Briat, J.-F.; Khodr, H.; Shioiri, T.; Leigh, R.A.; Hider, R.C. Nicotianamine chelates both FeIII and FeII. Implications for metal transport in plants. Plant Physiol. 1999, 119, 1107–1114. [Google Scholar] [CrossRef] [Green Version]
- Müller, B.; Kovács, K.; Pham, H.-D.; Kavak, Y.; Pechoušek, J.; Machala, L.; Zbořil, R.; Szenthe, K.; Abadía, J.; Fodor, F.; et al. Chloroplasts preferentially take up ferric–citrate over iron–nicotianamine complexes in Brassica napus. Planta 2019, 249, 751–763. [Google Scholar] [CrossRef]
- Gopika, S.; Augustine, C. Theoretical studies on the coordination chemistry of phytosiderophores with special reference to Fe-nicotianamine complexes in graminaceous plants. J. Mol. Model. 2022, 28, 71. [Google Scholar] [CrossRef] [PubMed]
- Reichman, S.M.; Parker, D.R. Revisiting the metal-binding chemistry of nicotianamine and 2′-deoxymugineic acid. Implications for iron nutrition in strategy II Plants. Plant Physiol. 2002, 129, 1435–1438. [Google Scholar] [CrossRef] [Green Version]
- Nishiyama, R.; Kato, M.; Nagata, S.; Yanagisawa, S.; Yoneyama, T. Identification of Zn-nicotianamine and Fe-2′-deoxymugineic acid in the phloem sap from rice plants (Oryza sativa L.). Plant Cell Physiol. 2012, 53, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Hazama, K.; Nagata, S.; Fujimori, T.; Yanagisawa, S.; Yoneyama, T. Concentrations of metals and potential metal-binding compounds and speciation of Cd, Zn and Cu in phloem and xylem saps from castor bean plants (Ricinus communis) treated with four levels of cadmium. Physiol. Plant. 2015, 154, 243–255. [Google Scholar] [CrossRef]
- Flis, P.; Ouerdane, L.; Grillet, L.; Curie, C.; Mari, S.; Lobinski, R. Inventory of metal complexes circulating in plant fluids: A reliable method based on HPLC coupled with dual elemental and high-resolution molecular mass spectrometric detection. New Phytol. 2016, 211, 1129–1141. [Google Scholar] [CrossRef] [Green Version]
- Wojcieszek, J.; Jiménez-Lamana, J.; Bierla, K.; Asztemborska, M.; Ruzik, L.; Jarosz, M.; Szpunar, J. Elucidation of the fate of zinc in model plants using single particle ICP-MS and ESI tandem MS. J. Anal. At. Spectrom. 2019, 34, 683–693. [Google Scholar] [CrossRef]
- Alchoubassi, G.; Kińska, K.; Bierla, K.; Lobinski, R.; Szpunar, J. Speciation of essential nutrient trace elements in coconut water. Food Chem. 2021, 339, 127680. [Google Scholar] [CrossRef]
- Mijovilovich, A.; Leitenmaier, B.; Meyer-Klaucke, W.; Kroneck, P.M.H.; Götz, B.; Küpper, H. Complexation and toxicity of copper in higher plants. II. Different mechanisms for copper versus cadmium detoxification in the copper-sensitive cadmium/zinc hyperaccumulator Thlaspi caerulescens (Ganges Ecotype). Plant Physiol. 2009, 151, 715–731. [Google Scholar] [CrossRef] [Green Version]
- Kutrowska, A.; Szelag, M. Low-molecular weight organic acids and peptides involved in the long-distance transport of trace metals. Acta Physiol. Plant. 2014, 36, 1957–1968. [Google Scholar] [CrossRef] [Green Version]
- Rellán-Álvarez, R.; Abadía, J.; Álvarez-Fernández, A. Formation of metal-nicotianamine complexes as affected by pH, ligand exchange with citrate and metal exchange. A study by electrospray ionization time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2008, 22, 1553–1562. [Google Scholar] [CrossRef]
- Shojima, S.; Nishizawa, N.-K.; Fushiya, S.; Nozoe, S.; Irifune, T.; Mori, S. Biosynthesis of phytosiderophores. In vitro biosynthesis of 2ʹ-deoxymugineic acid from L-methionine and nicotianamine. Plant Physiol. 1990, 93, 1497–1503. [Google Scholar] [CrossRef] [Green Version]
- Heidari, P.; Mazloomi, F.; Nussbaumer, T.; Barcaccia, G. Insights into the SAM synthetase gene family and its roles in tomato seedlings under abiotic stresses and hormone treatments. Plants 2020, 9, 586. [Google Scholar] [CrossRef]
- Shojima, S.; Nishizawa, N.K.; Fushiya, S.; Nozoe, S.; Kumashiro, T.; Nagata, T.; Ohata, T.; Mori, S. Biosynthesis of nicotianamine in the suspension-cultured cells of tobacco (Nicotiana megalosiphon). Biol. Met. 1989, 2, 142–145. [Google Scholar] [CrossRef]
- Higuchi, K.; Kanazawa, K.; Nishizawa, N.-K.; Chino, M.; Mori, S. Purification and characterization of nicotianamine synthase from Fe-deficient barley roots. Plant Soil 1994, 165, 173–179. [Google Scholar] [CrossRef]
- Higuchi, K.; Suzuki, K.; Nakanishi, H.; Yamaguchi, H.; Nishizawa, N.-K.; Mori, S. Cloning of nicotianamine synthase genes, novel genes involved in the biosynthesis of phytosiderophores. Plant Physiol. 1999, 119, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.S.; Dietz, K.-J. The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef] [Green Version]
- Herbik, A.; Koch, G.; Mock, H.-P.; Dushkov, D.; Czihal, A.; Thielmann, J.; Stephan, U.W.; Bäumlein, H. Isolation, characterization and cDNA cloning of nicotianamine synthase from barley. A key enzyme for iron homeostasis in plants. Eur. J. Biochem. 1999, 265, 231–239. [Google Scholar] [CrossRef] [Green Version]
- Douchkov, D.; Herbik, A.; Koch, G.; Mock, H.-P.; Melzer, M.; Stephan, U.W.; Bäumlein, H. Nicotianamine synthase: Gene isolation, gene transfer and application for the manipulation of plant iron assimilation. Plant Soil 2002, 241, 115–119. [Google Scholar] [CrossRef]
- Bonneau, J.; Baumann, U.; Beasley, J.; Li, Y.; Johnson, A.A.T. Identification and molecular characterization of the nicotianamine synthase gene family in bread wheat. Plant Biotechnol. J. 2016, 14, 2228–2239. [Google Scholar] [CrossRef] [Green Version]
- Kumar, N.; Tiwari, A.; Pal, N.; Kumar, S. Genome-wide identification, characterization and relative expression analysis of putative iron homeostasis genes: NAS, NAAT, and DMAS in hexaploid wheat and its progenitors. J. Cereal Sci. 2022, 105, 103466. [Google Scholar] [CrossRef]
- van de Mortel, J.E.; Villanueva, L.A.; Schat, H.; Kwekkeboom, J.; Coughlan, S.; Moerland, P.D.; van Themaat, E.V.L.; Koornneef, M.; Aarts, M.G.M. Large expression differences in genes for iron and zinc homeostasis, stress response, and lignin biosynthesis distinguish roots of Arabidopsis thaliana and the related metal hyperaccumulator Thlaspi caerulescens. Plant Physiol. 2006, 142, 1127–1147. [Google Scholar] [CrossRef] [Green Version]
- van de Mortel, J.E.; Schat, H.; Moerland, P.D.; van Themaat, E.V.L.; van der Ent, S.; Blankestijn, H.; Ghandilyan, A.; Tsiatsiani, S.; Aarts, M.G.M. Expression differences for genes involved in lignin, glutathione and sulphate metabolism in response to cadmium in Arabidopsis thaliana and the related Zn/Cd-hyperaccumulator Thlaspi caerulescens. Plant Cell Environ. 2008, 31, 301–324. [Google Scholar] [CrossRef]
- Klatte, M.; Schuler, M.; Wirtz, M.; Fink-Straube, C.; Hell, R.; Bauer, P. The analysis of Arabidopsis nicotianamine synthase mutants reveals functions for nicotianamine in seed iron loading and iron deficiency responses. Plant Physiol. 2009, 150, 257–271. [Google Scholar] [CrossRef] [Green Version]
- Becher, M.; Talke, I.N.; Krall, L.; Krämer, U. Cross-species microarray transcript profiling reveals high constitutive expression of metal homeostasis genes in shoots of the zinc hyperaccumulator Arabidopsis halleri. Plant J. 2004, 37, 251–268. [Google Scholar] [CrossRef]
- Weber, M.; Harada, E.; Vess, C.; Roepenack-Lahaye, E.V.; Clemens, S. Comparative microarray analysis of Arabidopsis thaliana and Arabidopsis halleri roots identifies nicotianamine synthase, a ZIP transporter and other genes as potential metal hyperaccumulation factors. Plant J. 2004, 37, 269–281. [Google Scholar] [CrossRef]
- Chen, S.; Sahito, Z.A.; Zhang, M.; Feng, Y.; Yang, Q.; Yang, X. Identification and characterization of four nicotianamine synthase genes in Sedum alfredii Hance. J. Biobased Mater. Bio. 2018, 12, 551–559. [Google Scholar] [CrossRef]
- Chen, S.; Zhang, M.; Feng, Y.; Sahito, Z.A.; Tian, S.; Yang, X. Nicotianamine synthase gene 1 from the hyperaccumulator Sedum alfredii Hance is associated with Cd/Zn tolerance and accumulation in plants. Plant Soil 2019, 443, 413–427. [Google Scholar] [CrossRef]
- Singh, R.; Misra, A.N.; Sharma, P. Effect of arsenate toxicity on antioxidant enzymes and expression of nicotianamine synthase in contrasting genotypes of bioenergy crop Ricinus communis. Environ. Sci. Pollut. Res. 2021, 28, 31421–31430. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, K.; Watanabe, S.; Takahashi, M.; Kawasaki, S.; Nakanishi, H.; Nishizawa, N.K.; Mori, S. Nicotianamine synthase gene expression differs in barley and rice under Fe-deficient conditions. Plant J. 2001, 25, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Perovic, D.; Tiffin, P.; Douchkov, D.; Bäumlein, H.; Graner, A. An integrated approach for the comparative analysis of a multigene family: The nicotianamine synthase genes of barley. Funct. Integr. Genom. 2007, 7, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Higuchi, K.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Three rice nicotianamine synthase genes, OsNAS1, OsNAS2, and OsNAS3 are expressed in cells involved in long-distance transport of iron and differentially regulated by iron. Plant J. 2003, 36, 366–381. [Google Scholar] [CrossRef]
- Aung, M.S.; Masuda, H.; Kobayashi, T.; Nishizawa, N.K. Physiological and transcriptomic analysis of responses to different levels of iron excess stress in various rice tissues. Soil Sci. Plant Nutr. 2018, 64, 370–385. [Google Scholar] [CrossRef]
- Aung, M.S.; Masuda, H.; Nozoye, T.; Kobayashi, T.; Jeon, J.-S.; An, G.; Nishizawa, N.K. Nicotianamine synthesis by OsNAS3 is important for mitigating iron excess stress in rice. Front. Plant. Sci. 2019, 10, 660. [Google Scholar] [CrossRef] [Green Version]
- Du, X.; Wang, H.; He, J.; Zhu, B.; Guo, J.; Hou, W.; Weng, Q.; Zhang, X. Identification of nicotianamine synthase genes in Triticum monococcum and their expression under different Fe and Zn concentrations. Gene 2018, 672, 1–7. [Google Scholar] [CrossRef]
- Mizuno, D.; Higuchi, K.; Sakamoto, T.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Three nicotianamine synthase genes isolated from maize are differentially regulated by iron nutritional status. Plant Physiol. 2003, 132, 1989–1997. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.L.; Qi, L.P.; Pang, J.F.; Zhang, Q.; Lei, Z.; Tang, Y.X.; Zhu, X.M.; Shao, J.R.; Wu, Y.M. Nicotianamine synthase gene family as central components in heavy metal and phytohormone response in maize. Funct. Integr. Genom. 2013, 13, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Li, S.; Zhao, Q.; Liu, X.; Zhang, S.; Sun, C.; Fan, Y.; Zhang, C.; Chen, R. Genome-wide identification, classification and expression profiling of nicotianamine synthase (NAS) gene family in maize. BMC Genom. 2013, 14, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, D.-G.; Yang, G.-H.; Xu, K.-D.; Shao, Q.; Yu, Z.-Y.; Wang, B.; Ge, Q.-L.; Yu, Y. Overexpression of a Malus xiaojinensis Nas1 gene influences flower development and tolerance to iron stress in transgenic tobacco. Plant Mol. Biol. Rep. 2013, 31, 802–809. [Google Scholar] [CrossRef]
- Han, D.; Zhang, Z.; Ni, B.; Ding, H.; Liu, W.; Li, W.; Chai, L.; Yang, G. Isolation and functional analysis of MxNAS3 involved in enhanced iron stress tolerance and abnormal flower in transgenic Arabidopsis. J. Plant Interact. 2018, 13, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Li, J.; Liu, W.; Yu, Z.; Shi, Y.; Lv, B.; Wang, B.; Han, D. Molecular cloning and characterization of MxNAS2, a gene encoding nicotianamine synthase in Malus xiaojinensis, with functions in tolerance to iron stress and misshapen flower in transgenic tobacco. Sci. Hortic. 2015, 183, 77–86. [Google Scholar] [CrossRef]
- Ling, H.-Q.; Koch, G.; Bäumlein, H.; Ganal, M.W. Map-based cloning of chloronerva, a gene involved in iron uptake of higher plants encoding nicotianamine synthase. Proc. Natl. Acad. Sci. USA 1999, 96, 7098–7103. [Google Scholar] [CrossRef] [Green Version]
- Aprile, A.; Sabella, E.; Francia, E.; Milc, J.; Ronga, D.; Pecchioni, N.; Ferrari, E.; Luvisi, A.; Vergine, M.; De Bellis, L. Combined effect of cadmium and lead on durum wheat. Int. J. Mol. Sci. 2019, 20, 5891. [Google Scholar] [CrossRef] [Green Version]
- Halimaa, P.; Blande, D.; Baltzi, E.; Aarts, M.G.M.; Granlund, L.; Keinänen, M.; Kärenlampi, S.O.; Kozhevnikova, A.D.; Peräniemi, S.; Schat, H.; et al. Transcriptional effects of cadmium on iron homeostasis differ in calamine accessions of Noccaea caerulescens. Plant J. 2019, 97, 306–320. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Zhao, M.; Chen, J.; Yang, S.; Chen, J.; Xue, Y. Comparative transcriptome analysis reveals complex physiological response and gene regulation in peanut roots and leaves under manganese toxicity stress. Int. J. Mol. Sci. 2023, 24, 1161. [Google Scholar] [CrossRef]
- Wintz, H.; Fox, T.; Wu, Y.-Y.; Feng, V.; Chen, W.; Chang, H.-S.; Zhu, T.; Vulpe, C. Expression profiles of Arabidopsis thaliana in mineral deficiencies reveal novel transporters involved in metal homeostasis. J. Biol. Chem. 2003, 278, 47644–47653. [Google Scholar] [CrossRef] [Green Version]
- Cardini, A.; Pellegrino, E.; White, P.J.; Mazzolai, B.; Mascherpa, M.C.; Ercoli, L. Transcriptional regulation of genes involved in zinc uptake, sequestration and redistribution following foliar zinc application to Medicago sativa. Plants 2021, 10, 476. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.-R.; Qu, B.-Y.; Tong, Y.-P.; Zou, C.-Q. Iron and zinc accumulation in winter wheat regulated by NICOTIANAMINE SYNTHASE responded to increasing nitrogen levels. J. Plant Nutr. 2019, 42, 1624–1636. [Google Scholar] [CrossRef]
- Ishimaru, Y.; Suzuki, M.; Ogo, Y.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Synthesis of nicotianamine and deoxymugineic acid is regulated by OsIRO2 in Zn excess rice plants. Soil Sci. Plant Nutr. 2008, 54, 417–423. [Google Scholar] [CrossRef]
- Mallikarjuna, M.G.; Thirunavukkarasu, N.; Sharma, R.; Shiriga, K.; Hossain, F.; Bhat, J.S.; Mithra, A.C.R.; Marla, S.S.; Manjaiah, K.M.; Rao, A.R.; et al. Comparative transcriptome analysis of iron and zinc deficiency in maize (Zea mays L.). Plants 2020, 9, 1812. [Google Scholar] [CrossRef] [PubMed]
- Nozoye, T.; Inoue, H.; Takahashi, M.; Ishimaru, Y.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. The expression of iron homeostasis-related genes during rice germination. Plant Mol. Biol. 2007, 64, 35–47. [Google Scholar] [CrossRef]
- Escudero, V.; Abreu, I.; del Sastre, E.; Tejada-Jiménez, M.; Larue, C.; Novoa-Aponte, L.; Castillo-González, J.; Wen, J.; Mysore, K.S.; Abadía, J.; et al. Nicotianamine synthase 2 is required for symbiotic nitrogen fixation in Medicago truncatula nodules. Front. Plant Sci. 2020, 10, 1780. [Google Scholar] [CrossRef]
- Palmer, C.M.; Hindt, M.N.; Schmidt, H.; Clemens, S.; Guerinot, M.L. MYB10 and MYB72 are required for growth under iron-limiting conditions. PLoS Genet. 2013, 9, e1003953. [Google Scholar] [CrossRef] [Green Version]
- Long, T.A.; Tsukagoshi, H.; Busch, W.; Lahner, B.; Salt, D.E.; Benfey, P.N. The bHLH transcription factor POPEYE regulates response to iron deficiency in Arabidopsis roots. Plant Cell 2010, 22, 2219–2236. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Ye, J.; Ma, Y.; Wang, T.; Shou, H.; Zheng, L. OsIRO3 plays an essential role in iron deficiency responses and regulates iron homeostasis in rice. Plants 2020, 9, 1095. [Google Scholar] [CrossRef]
- Wang, H.; Liao, S.; Li, M.; Wei, J.; Zhu, B.; Gu, L.; Li, L.; Du, X. TmNAS3 from Triticum monococcum directly regulated by TmbHLH47 increases Fe content of wheat grain. Gene 2022, 811, 146096. [Google Scholar] [CrossRef]
- Ivanov, R.; Brumbarova, T.; Bauer, P. Fitting into the harsh reality: Regulation of iron-deficiency responses in dicotyledonous plants. Mol. Plant 2012, 5, 27–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lilay, G.H.; Persson, D.P.; Castro, P.H.; Liao, F.; Alexander, R.D.; Aarts, M.G.M.; Assunção, A.G.L. Arabidopsis bZIP19 and bZIP23 act as zinc sensors to control plant zinc status. Nat. Plants 2021, 7, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Assunção, A.G.L. The F-bZIP-regulated Zn deficiency response in land plants. Planta 2022, 256, 108. [Google Scholar] [CrossRef] [PubMed]
- Liao, F.; Lilay, G.H.; Castro, P.H.; Azevedo, H.; Assunção, A.G.L. Regulation of the zinc deficiency response in the legume model Medicago truncatula. Front. Plant Sci. 2022, 13, 916168. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-K.; Chung, P.J.; Jeong, J.S.; Jang, G.; Bang, S.W.; Jung, H.; Kim, Y.S.; Ha, S.-H.; Choi, Y.D.; Kim, J.-K. The rice OsNAC6 transcription factor orchestrates multiple molecular mechanisms involving root structural adaptions and nicotianamine biosynthesis for drought tolerance. Plant Biotechnol. J. 2017, 15, 754–764. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Weber, M.; Clemens, S. Elevated root nicotianamine concentrations are critical for Zn hyperaccumulation across diverse edaphic environments. Plant Cell Environ. 2019, 42, 2003–2014. [Google Scholar] [CrossRef]
- Wu, J.; Wang, C.; Zheng, L.; Wang, L.; Chen, Y.; Whelan, J.; Shou, H. Ethylene is involved in the regulation of iron homeostasis by regulating the expression of iron-acquisition-related genes in Oryza sativa. J. Exp. Bot. 2011, 62, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Jeon, U.S.; Lee, S.J.; Kim, Y.-K.; Persson, D.P.; Husted, S.; Schjørring, J.K.; Kakei, Y.; Masuda, H.; Nishizawa, N.K.; et al. Iron fortification of rice seeds through activation of the nicotianamine synthase gene. Proc. Natl. Acad. Sci. USA 2009, 106, 22014–22019. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Persson, D.P.; Hansen, T.H.; Husted, S.; Schjoerring, J.K.; Kim, Y.-S.; Jeon, U.S.; Kim, Y.-K.; Kakei, Y.; Masuda, H.; et al. Bio-available zinc in rice seeds is increased by activation tagging of nicotianamine synthase. Plant Biotechnol. J. 2011, 9, 865–873. [Google Scholar] [CrossRef]
- Díaz-Benito, P.; Banakar, R.; Rodríguez-Menéndez, S.; Capell, T.; Pereiro, R.; Christou, P.; Abadía, J.; Fernández, B.; Álvarez-Fernández, A. Iron and zinc in the embryo and endosperm of rice (Oryza sativa L.) seeds in contrasting 2′-deoxymugineic acid/nicotianamine scenarios. Front. Plant Sci. 2018, 9, 1190. [Google Scholar] [CrossRef] [Green Version]
- Beasley, J.T.; Bonneau, J.P.; Sánchez-Palacios, J.T.; Moreno-Moyano, L.T.; Callahan, D.L.; Tako, E.; Glahn, R.P.; Lombi, E.; Johnson, A.A.T. Metabolic engineering of bread wheat improves grain iron concentration and bioavailability. Plant Biotechnol. J. 2019, 17, 1514–1526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, C.; Yuan, M.; Zhai, L.; Li, D.; Zhang, X.; Wu, T.; Xu, X.; Wang, Y.; Han, Z. Iron deficiency stress can induce MxNAS1 protein expression to facilitate iron redistribution in Malus xiaojinensis. Plant Biol. 2018, 20, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Deinlein, U.; Weber, M.; Schmidt, H.; Rensch, S.; Trampczynska, A.; Hansen, T.H.; Husted, S.; Schjoerring, J.K.; Talke, I.N.; Krämer, U.; et al. Elevated nicotianamine levels in Arabidopsis halleri roots play a key role in zinc hyperaccumulation. Plant Cell 2012, 24, 708–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montacié, C.; Riondet, C.; Wei, L.; Darriere, T.; Weiss, A.; Pontvianne, F.; Escande, M.L.; de Bures, A.; Jobet, E.; Barbarossa, A.; et al. nicotianamine synthase activity affects nucleolar iron accumulation and impacts rDNA silencing and RNA methylation in Arabidopsis. J. Exp. Bot. 2023, erad180. [Google Scholar] [CrossRef] [PubMed]
- Talke, I.N.; Hanikenne, M.; Krämer, U. Zinc-dependent global transcriptional control, transcriptional deregulation, and higher gene copy number for genes in metal homeostasis of the hyperaccumulator Arabidopsis halleri. Plant Physiol. 2006, 142, 148–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schvartzman, M.S.; Corso, M.; Fataftah, N.; Scheepers, M.; Nouet, C.; Bosman, B.; Carnol, M.; Motte, P.; Verbruggen, N.; Hanikenne, M. Adaptation to high zinc depends on distinct mechanisms in metallicolous populations of Arabidopsis halleri. New Phytol. 2018, 218, 269–282. [Google Scholar] [CrossRef] [Green Version]
- Callahan, D.L.; Kolev, S.D.; O’Hair, R.A.J.; Salt, D.E.; Baker, A.J.M. Relationships of nicotianamine and other amino acids with nickel, zinc and iron in Thlaspi hyperaccumulators. New Phytol. 2007, 176, 836–848. [Google Scholar] [CrossRef]
- Foroughi, S.; Baker, A.J.M.; Roessner, U.; Johnson, A.A.T.; Bacic, A.; Callahan, D.L. Hyperaccumulation of zinc by Noccaea caerulescens results in a cascade of stress responses and changes in the elemental profile. Metallomics 2014, 6, 1671–1682. [Google Scholar] [CrossRef]
- Halimaa, P.; Lin, Y.-F.; Ahonen, V.H.; Blande, D.; Clemens, S.; Gyenesei, A.; Häikiö, E.; Kärenlampi, S.O.; Laiho, A.; Aarts, M.G.M.; et al. Gene expression differences between Noccaea caerulescens ecotypes help to identify candidate genes for metal phytoremediation. Environ. Sci. Technol. 2014, 48, 3344–3353. [Google Scholar] [CrossRef]
- Visioli, G.; Gullì, M.; Marmiroli, N. Noccaea caerulescens populations adapted to grow in metalliferous and non-metalliferous soils: Ni tolerance, accumulation and expression analysis of genes involved in metal homeostasis. Environ. Exp. Bot. 2014, 105, 10–17. [Google Scholar] [CrossRef]
- Seregin, I.V.; Kozhevnikova, A.D. Roles of root and shoot tissues in transport and accumulation of cadmium, lead, nickel, and strontium. Russ. J. Plant Physiol. 2008, 55, 1–22. [Google Scholar] [CrossRef]
- Schneider, T.; Persson, D.P.; Husted, S.; Schellenberg, M.; Gehrig, P.; Lee, Y.; Martinoia, E.; Schjoerring, J.K.; Meyer, S. A proteomics approach to investigate the process of Zn hyperaccumulation in Noccaea caerulescens (J & C. Presl) F.K. Meyer. Plant J. 2013, 73, 131–142. [Google Scholar] [CrossRef] [PubMed]
- Nozoye, T.; Nagasaka, S.; Bashir, K.; Takahashi, M.; Kobayashi, T.; Nakanishi, H.; Nishizawa, N.K. Nicotianamine synthase 2 localizes to the vesicles of iron-deficient rice roots, and its mutation in the YXXφ or LL motif causes the disruption of vesicle formation or movement in rice. Plant J. 2014, 77, 246–260. [Google Scholar] [CrossRef] [PubMed]
- Bashir, K.; Nozoye, T.; Nagasaka, S.; Rasheed, S.; Miyauchi, N.; Seki, M.; Nakanishi, H.; Nishizawa, N.K. Paralogs and mutants show that one DMA synthase functions in iron homeostasis in rice. J. Exp. Bot. 2017, 68, 1785–1795. [Google Scholar] [CrossRef] [Green Version]
- Gautam, T.; Jan, I.; Batra, R.; Singh, K.; Pandey, R.; Sharma, P.K.; Balyan, H.S.; Gupta, P.K. Further studies on nicotianamine aminotransferase (NAAT) genes involved in biofortification in bread wheat (Triticum aestivum L.). Plant Gene 2023, 33, 100389. [Google Scholar] [CrossRef]
- Yen, M.R.; Tseng, Y.H.; Saier, M.H., Jr. Maize Yellow Stripe1, an iron-phytosiderophore uptake transporter, is a member of the oligopeptide transporter (OPT) family. Microbiology 2001, 147, 2881–2883. [Google Scholar] [CrossRef] [Green Version]
- Kurt, F.; Filiz, E. Functional divergence in oligopeptide transporters in plants. In Cation Transporters in Plants; Kumar, S., Ed.; Academic Press: Cambridge, MA, USA, 2022; pp. 167–183. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, C.; Guo, H.; Hu, Y.; He, Y.; Jiang, D. Overexpression of a Miscanthus sacchariflorus Yellow Stripe-Like transporter MsYSL1 enhances resistance of Arabidopsis to cadmium by mediating metal ion reallocation. Plant Growth Regul. 2018, 85, 101–111. [Google Scholar] [CrossRef]
- Koike, S.; Inoue, H.; Mizuno, D.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. OsYSL2 is a rice metal-nicotianamine transporter that is regulated by iron and expressed in the phloem. Plant J. 2004, 39, 415–424. [Google Scholar] [CrossRef]
- Inoue, H.; Kobayashi, T.; Nozoye, T.; Takahashi, M.; Kakei, Y.; Suzuki, K.; Nakazono, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Rice OsYSL15 is an iron-regulated iron (III)-deoxymugineic acid transporter expressed in the roots and is essential for iron uptake in early growth of the seedlings. J. Biol. Chem. 2009, 284, 3470–3479. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, T.; Kobayashi, T.; Takahashi, M.; Nagasaka, S.; Usuda, K.; Kakei, Y.; Ishimaru, Y.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. OsYSL18 is a rice iron(III)-deoxymugineic acid transporter specifically expressed in reproductive organs and phloem of lamina joints. Plant Mol. Biol. 2009, 70, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Yamaji, N.; Xia, J.; Ma, J.F. OsYSL6 is involved in the detoxification of excess manganese in rice. Plant Physiol. 2011, 157, 1832–1840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, R.; Murata, J.; Murata, Y. A novel barley yellow stripe 1-like transporter (HvYSL2) localized to the root endodermis transports metal-phytosiderophore complexes. Plant Cell Physiol. 2011, 52, 1931–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.; Wang, N.; Xiong, H.; Qiu, W.; Nakanishi, H.; Kobayashi, T.; Nishizawa, N.K.; Zuo, Y. The Yellow Stripe-Like (YSL) gene functions in internal copper transport in peanut. Genes 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Shinwari, K.I.; Luo, L.; Zheng, L. OsYSL13 is involved in iron distribution in rice. Int. J. Mol. Sci. 2018, 19, 3537. [Google Scholar] [CrossRef] [Green Version]
- Foroozani, M. The role of the Yellow Stripe-Like transporter BdYSL3 in copper homeostasis in Brachypodium. Plant Physiol. 2021, 186, 204–205. [Google Scholar] [CrossRef]
- Chowdhury, R.; Nallusamy, S.; Shanmugam, V.; Loganathan, A.; Muthurajan, R.; Sivathapandian, S.K.; Paramasivam, J.; Duraialagaraja, S. Genome-wide understanding of evolutionary and functional relationships of rice Yellow Stripe-Like (YSL) transporter family in comparison with other plant species. Biologia 2022, 77, 39–53. [Google Scholar] [CrossRef]
- Sheng, H.; Jiang, Y.; Rahmati, M.; Chia, J.-C.; Dokuchayeva, T.; Kavulych, Y.; Zavodna, T.-O.; Mendoza, P.N.; Huang, R.; Smieshka, L.M.; et al. YSL3-mediated copper distribution is required for fertility, seed size and protein accumulation in Brachypodium. Plant Physiol. 2021, 186, 655–676. [Google Scholar] [CrossRef]
- Murata, Y.; Ma, J.F.; Yamaji, N.; Ueno, D.; Nomoto, K.; Iwashita, T. A specific transporter for iron(III)-phytosiderophore in barley roots. Plant J. 2006, 46, 563–572. [Google Scholar] [CrossRef]
- Harada, E.; Sugase, K.; Namba, K.; Iwashita, T.; Murata, Y. Structural element responsible for the Fe(III)-phytosiderophore specific transport by HvYS1 transporter in barley. FEBS Lett. 2007, 581, 4298–4302. [Google Scholar] [CrossRef] [Green Version]
- Banakar, R.; Alvarez Fernández, Á.; Abadía, J.; Capell, T.; Christou, P. The expression of heterologous Fe (III) phytosiderophore transporter HvYS1 in rice increases Fe uptake, translocation and seed loading and excludes heavy metals by selective Fe transport. Plant Biotechnol. J. 2017, 15, 423–432. [Google Scholar] [CrossRef] [Green Version]
- Ishimaru, Y.; Masuda, H.; Bashir, K.; Inoue, H.; Tsukamoto, T.; Takahashi, M.; Nakanishi, H.; Aoki, N.; Hirose, T.; Ohsugi, R.; et al. Rice metal-nicotianamine transporter, OsYSL2, is required for the long-distance transport of iron and manganese. Plant J. 2010, 62, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Senoura, T.; Sakashita, E.; Kobayashi, T.; Takahashi, M.; Aung, M.S.; Masuda, H.; Nakanishi, H.; Nishizawa, N.K. The iron-chelate transporter OsYSL9 plays a role in iron distribution in developing rice grains. Plant Mol. Biol. 2017, 95, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Chiecko, J.C.; Kim, S.A.; Walker, E.L.; Lee, Y.; Guerinot, M.L.; An, G. Disruption of OsYSL15 leads to iron inefficiency in rice plants. Plant Physiol. 2009, 150, 786–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakei, Y.; Ishimaru, Y.; Kobayashi, T.; Yamakawa, T.; Nakanishi, H.; Nishizawa, N.K. OsYSL16 plays a role in the allocation of iron. Plant Mol. Biol. 2012, 79, 583–594. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Yamaji, N.; Yokosho, K.; Ma, J.F. YSL16 is a phloem-localized transporter of the copper-nicotianamine complex that is responsible for copper distribution in rice. Plant Cell 2012, 24, 3767–3782. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Lu, W.; Yang, Y.; Shen, Z.; Ma, J.F.; Zheng, L. OsYSL16 is required for preferential Cu distribution to floral organs in rice. Plant Cell Physiol. 2018, 59, 2039–2051. [Google Scholar] [CrossRef]
- Curie, C.; Panaviene, Z.; Loulergue, C.; Dellaporta, S.L.; Briat, J.-F.; Walker, E.L. Maize yellow stripe1 encodes a membrane protein directly involved in Fe(III) uptake. Nature 2001, 409, 346–349. [Google Scholar] [CrossRef]
- Roberts, L.A.; Pierson, A.J.; Panaviene, Z.; Walker, E.L. Yellow Stripe1. Expanded roles for the maize iron-phytosiderophore transporter. Plant Physiol. 2004, 135, 112–120. [Google Scholar] [CrossRef] [Green Version]
- Schaaf, G.; Ludewig, U.; Erenoglu, B.E.; Mori, S.; Kitahara, T.; von Wirén, N. ZmYS1 Functions as a proton-coupled symporter for phytosiderophore- and nicotianamine-chelated metals. J. Biol. Chem. 2004, 279, 9091–9096. [Google Scholar] [CrossRef] [Green Version]
- Zang, J.; Huo, Y.; Liu, J.; Zhang, H.; Liu, J.; Chen, H. Maize YSL2 is required for iron distribution and development in kernels. J. Exp. Bot. 2020, 71, 5896–5910. [Google Scholar] [CrossRef]
- He, Y.; Yang, Q.; Yang, J.; Wang, Y.-F.; Sun, X.; Wang, S.; Qi, W.; Ma, Z.; Song, R. shrunken4 is a mutant allele of ZmYSL2 that affects aleurone development and starch synthesis in maize. Genetics 2021, 218, iyab070. [Google Scholar] [CrossRef] [PubMed]
- Chao, Z.-F.; Chen, Y.-Y.; Ji, C.; Wang, Y.-L.; Huang, X.; Zhang, C.-Y.; Yang, J.; Song, T.; Wu, J.-C.; Guo, L.-X.; et al. A genome-wide association study identifies a transporter for zinc uploading to maize kernels. EMBO Rep. 2023, 24, e55542. [Google Scholar] [CrossRef] [PubMed]
- Le Jean, M.; Schikora, A.; Mari, S.; Briat, J.-F.; Curie, C. A Loss-of-function mutation in AtYSL1 reveals its role in iron and nicotianamine seed loading. Plant J. 2005, 44, 769–782. [Google Scholar] [CrossRef] [PubMed]
- Waters, B.M.; Chu, H.-H.; DiDonato, R.J.; Roberts, L.A.; Eisley, R.B.; Lahner, B.; Salt, D.E.; Walker, E.L. Mutations in Arabidopsis Yellow Stripe-Like1 and Yellow Stripe-Like3 reveal their roles in metal ion homeostasis and loading of metal ions in seeds. Plant Physiol. 2006, 141, 1446–1458. [Google Scholar] [CrossRef] [Green Version]
- Chu, H.-H.; Chiecko, J.; Punshon, T.; Lanzirotti, A.; Lahner, B.; Salt, D.E.; Walker, E.L. Successful reproduction requires the function of Arabidopsis YELLOW STRIPE-LIKE1 and YELLOW STRIPE-LIKE3 metal-nicotianamine transporters in both vegetative and reproductive structures. Plant Physiol. 2010, 154, 197–210. [Google Scholar] [CrossRef] [Green Version]
- DiDonato, R.J.; Roberts, L.A.; Sanderson, T.; Eisley, R.B.; Walker, E.L. Arabidopsis Yellow Stripe-Like2 (YSL2): A metal-regulated gene encoding a plasma membrane transporter of nicotianamine-metal complexes. Plant J. 2004, 39, 403–414. [Google Scholar] [CrossRef]
- Schaaf, G.; Schikora, A.; Häberle, J.; Vert, G.; Ludewig, U.; Briat, J.-F.; Curie, C.; von Wirén, N. A putative function for the Arabidopsis Fe-phytosiderophore transporter homolog AtYSL2 in Fe and Zn homeostasis. Plant Cell Physiol. 2005, 46, 762–774. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.K.; Chu, H.-H.; Abundis, C.; Vasques, K.; Rodriguez, D.C.; Chia, J.-C.; Huang, R.; Vatamaniuk, O.K.; Walker, E.L. Iron-nicotianamine transporters are required for proper long distance iron signaling. Plant Physiol. 2017, 175, 1254–1268. [Google Scholar] [CrossRef] [Green Version]
- Gendre, D.; Czernic, P.; Conéjéro, G.; Pianelli, K.; Briat, J.-F.; Lebrun, M.; Mari, S. TcYSL3, a member of the YSL gene family from the hyperaccumulator Thlaspi caerulescens, encodes a nicotianamine-Ni/Fe transporter. Plant J. 2006, 49, 1–15. [Google Scholar] [CrossRef]
- Xiong, H.; Kakei, Y.; Kobayashi, T.; Guo, X.; Nakazono, M.; Takahashi, H.; Nakanishi, H.; Shen, H.; Zhang, F.; Nishizawa, N.K.; et al. Molecular evidence for phytosiderophore-induced improvement of iron nutrition of peanut intercropped with maize in calcareous soil. Plant Cell Environ. 2013, 36, 1888–1902. [Google Scholar] [CrossRef]
- Feng, S.; Tan, J.; Zhang, Y.; Liang, S.; Xiang, S.; Wang, H.; Chai, T. Isolation and characterization of a novel cadmium-regulated Yellow Stripe-Like transporter (SnYSL3) in Solanum nigrum. Plant Cell Rep. 2017, 36, 281–296. [Google Scholar] [CrossRef] [PubMed]
- von Wirén, N.; Marschner, H.; Römheld, V. Roots of iron-efficient maize also absorb phytosiderophore-chelated zinc. Plant Physiol. 1996, 111, 1119–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yordem, B.K.; Conte, S.S.; Ma, J.F.; Yokosho, K.; Vasques, K.A.; Gopalsamy, S.N.; Walker, E.L. Brachypodium distachyon as a new model system for understanding iron homeostasis in grasses: Phylogenetic and expression analysis of Yellow Stripe-Like (YSL) transporters. Ann. Bot. 2011, 108, 821–833. [Google Scholar] [CrossRef] [PubMed]
- Sinclair, S.A.; Krämer, U. The zinc homeostasis network of land plants. Biochim. Biophys. Acta Mol. Cell Res. 2012, 1823, 1553–1567. [Google Scholar] [CrossRef]
- Grotz, N.; Guerinot, M.L. Molecular aspects of Cu, Fe and Zn homeostasis in plants. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 595–608. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Wang, T.; Song, H.; Jia, Y.; Ma, Q.; Tao, M.; Zhu, X.; Cao, S. The transcription factor WRKY12 negatively regulates iron entry into seeds in Arabidopsis. J. Exp. Bot. 2023, 74, 415–426. [Google Scholar] [CrossRef]
- Yang, J.; Chen, J.; Chen, X.; Ma, G.; Wang, P.; Fabrice, M.R.; Zhang, S.; Wu, J. Phylogenetic and expression analysis of pear Yellow Stripe-Like transporters and functional verification of PbrYSL4 in pear pollen. Plant Mol. Biol. Report. 2016, 34, 737–747. [Google Scholar] [CrossRef]
- Das, S.; Sen, M.; Saha, C.; Chakraborty, D.; Das, A.; Banerjee, M.; Seal, A. Isolation and expression analysis of partial sequences of heavy metal transporters from Brassica juncea by coupling high throughput cloning with a molecular fingerprinting technique. Planta 2011, 234, 139–156. [Google Scholar] [CrossRef]
- Divol, F.; Couch, D.; Conéjéro, G.; Roschzttardtz, H.; Mari, S.; Curie, C. The Arabidopsis Yellow Stripe Like4 and 6 transporters control iron release from the chloroplast. Plant Cell 2013, 25, 1040–1055. [Google Scholar] [CrossRef] [Green Version]
- Vigani, G.; Solti, Á.; Thomine, S.; Philippar, K. Essential and detrimental—An update on intracellular iron trafficking and homeostasis. Plant Cell Physiol. 2019, 60, 1420–1439. [Google Scholar] [CrossRef]
- Jaquinod, M.; Villiers, F.; Kieffer-Jaquinod, S.; Hugouvieux, V.; Bruley, C.; Garin, J.; Bourguignon, J. A proteomics dissection of Arabidopsis thaliana vacuoles isolated from cell culture. Mol. Cell. Proteom. 2007, 6, 394–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conte, S.S.; Chu, H.H.; Chan-Rodriguez, D.; Punshon, T.; Vasques, K.A.; Salt, D.E.; Walker, E.L. Arabidopsis thaliana Yellow Stripe1-Like4 and Yellow Stripe1-Like6 localize to internal cellular membranes and are involved in metal ion homeostasis. Front. Plant Sci. 2013, 4, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarts, M.G.M. Nicotianamine secretion for zinc excess tolerance. Plant Physiol. 2014, 166, 751–752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nozoye, T.; von Wirén, N.; Sato, Y.; Higashiyama, T.; Nakanishi, H.; Nishizawa, N.K. Characterization of the nicotianamine exporter ENA1 in rice. Front. Plant Sci. 2019, 10, 502. [Google Scholar] [CrossRef] [PubMed]
- Negishi, T.; Nakanishi, H.; Yazaki, J.; Kishimoto, N.; Fujii, F.; Shimbo, K.; Yamamoto, K.; Sakata, K.; Sasaki, T.; Kikuchi, S.; et al. cDNA microarray analysis of gene expression during Fe-deficiency stress in barley suggests that polar transport of vesicles is implicated in phytosiderophore secretion in Fe-deficient barley roots. Plant J. 2002, 30, 83–94. [Google Scholar] [CrossRef] [Green Version]
- Nozoye, T.; Itai, R.N.; Nagasaka, S.; Takahashi, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Diurnal changes in the expression of genes that participate in phytosiderophore synthesis in rice. Soil Sci. Plant Nutr. 2004, 50, 1125–1131. [Google Scholar] [CrossRef]
- Nozoye, T.; Tsunoda, K.; Nagasaka, S.; Bashir, K.; Takahashi, M.; Kobayashi, T.; Nakanishi, H.; Nishizawa, N.K. Rice nicotianamine synthase localizes to particular vesicles for proper function. Plant Signal. Behav. 2014, 9, e28660. [Google Scholar] [CrossRef] [Green Version]
- Takagi, S.-I.; Nomoto, K.; Takemoto, T. Physiological aspect of mugineic acid, a possible phytosiderophore of graminaceous plants. J. Plant Nutr. 1984, 7, 469–477. [Google Scholar] [CrossRef]
- Pich, A.; Hillmer, S.; Manteuffel, R.; Scholz, G. First immunohistochemical localization of the endogenous Fe2+-chelator nicotianamine. J. Exp. Bot. 1997, 48, 759–767. [Google Scholar] [CrossRef]
- Pich, A.; Manteuffel, R.; Hillmer, S.; Scholz, G.; Schmidt, W. Fe homeostasis in plant cells: Does nicotianamine play multiple roles in the regulation of cytoplasmic Fe concentration? Planta 2001, 213, 967–976. [Google Scholar] [CrossRef]
- Haydon, M.J.; Kawachi, M.; Wirtz, M.; Hillmer, S.; Hell, R.; Krämer, U. Vacuolar nicotianamine has critical and distinct roles under iron deficiency and for zinc sequestration in Arabidopsis. Plant Cell 2012, 24, 724–737. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.S.; Dietz, K.-J.; Mimura, T. Vacuolar compartmentalization as indispensable component of heavy metal detoxification in plants. Plant Cell Environ. 2016, 39, 1112–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, M.; Terada, Y.; Nakai, I.; Nakanishi, H.; Yoshimura, E.; Mori, S.; Nishizawa, N.K. Role of nicotianamine in the intracellular delivery of metals and plant reproductive development. Plant Cell 2003, 15, 1263–1280. [Google Scholar] [CrossRef] [PubMed]
- Trampczynska, A.; Küpper, H.; Meyer-Klaucke, W.; Schmidt, H.; Clemens, S. Nicotianamine forms complexes with Zn (II) in vivo. Metallomics. 2010, 2, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Richau, K.H.; Kozhevnikova, A.D.; Seregin, I.V.; Vooijs, R.; Koevoets, P.L.M.; Smith, J.A.C.; Ivanov, V.B.; Schat, H. Chelation by histidine inhibits the vacuolar sequestration of nickel in roots of the hyperaccumulator Thlaspi caerulescens. New Phytol. 2009, 183, 106–116. [Google Scholar] [CrossRef] [PubMed]
- Kozhevnikova, A.D.; Seregin, I.V.; Erlikh, N.T.; Shevyreva, T.A.; Andreev, I.M.; Verweij, R.; Schat, H. Histidine-mediated xylem loading of zinc is a species-wide character in Noccaea caerulescens. New Phytol. 2014, 203, 508–519. [Google Scholar] [CrossRef]
- Schuler, M.; Rellán-Álvarez, R.; Fink-Straube, C.; Abadía, J.; Bauer, P. Nicotianamine functions in the phloem-based transport of iron to sink organs, in pollen development and pollen tube growth in Arabidopsis. Plant Cell 2012, 24, 2380–2400. [Google Scholar] [CrossRef] [Green Version]
- Cornu, J.-Y.; Deinlein, U.; Höreth, S.; Braun, M.; Schmidt, H.; Weber, M.; Persson, D.P.; Husted, S.; Schjoerring, J.K.; Clemens, S. Contrasting effects of nicotianamine synthase knockdown on zinc and nickel tolerance and accumulation in the zinc/cadmium hyperaccumulator Arabidopsis halleri. New Phytol. 2015, 206, 738–750. [Google Scholar] [CrossRef]
- Johnson, A.A.T.; Kyriacou, B.; Callahan, D.L.; Carruthers, L.; Stangoulis, J.; Lombi, E.; Tester, M. Constitutive overexpression of the OsNAS gene family reveals single-gene strategies for effective iron- and zinc-biofortification of rice endosperm. PLoS ONE 2011, 6, e24476. [Google Scholar] [CrossRef] [Green Version]
- Persson, D.P.; Chen, A.; Aarts, M.G.M.; Salt, D.E.; Schjoerring, J.K.; Husted, S. Multi-element bioimaging of Arabidopsis thaliana roots. Plant Physiol. 2016, 172, 835–847. [Google Scholar] [CrossRef] [Green Version]
- Pich, A.; Scholz, G.; Stephan, U.W. Iron-dependent changes of heavy metals, nicotianamine, and citrate in different plant organs and in the xylem exudate of two tomato genotypes. Nicotianamine as possible copper translocator. Plant Soil 1994, 165, 189–196. [Google Scholar] [CrossRef]
- Stephan, U.W.; Schmidke, I.; Pich, A. Phloem translocation of Fe, Cu, Mn, and Zn in Ricinus seedlings in relation to the concentrations of nicotianamine, an endogenous chelator of divalent metal ions, in different seedling parts. Plant Soil 1994, 165, 181–188. [Google Scholar] [CrossRef]
- Pich, A.; Scholz, G. Translocation of copper and other micronutrients in tomato plants (Lycopersicon esculentum Mill.): Nicotianamine-stimulated copper transport in the xylem. J. Exp. Bot. 1996, 47, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Liao, M.T.; Hedley, M.J.; Woolley, D.J.; Brooks, R.R.; Nichols, M.A. Copper uptake and translocation in chicory (Cichorium intybus L. cv. Grasslands Puna) and tomato (Lycopersicon esculentum Mill. cv. Rondy) plants grown in NFT System. II. The role of nicotianamine and histidine in xylem sap copper transport. Plant Soil 2000, 223, 243–252. [Google Scholar] [CrossRef]
- Irtelli, B.; Petrucci, W.A.; Navari-Izzo, F. Nicotianamine and histidine/proline are, respectively, the most important copper chelators in xylem sap of Brassica carinata under conditions of copper deficiency and excess. J. Exp. Bot. 2009, 60, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakei, Y.; Yamaguchi, I.; Kobayashi, T.; Takahashi, M.; Nakanishi, H.; Yamakawa, T.; Nishizawa, N.K. A highly sensitive, quick and simple quantification method for nicotianamine and 2′-deoxymugineic acid from minimum samples using LC/ESI-TOF-MS achieves functional analysis of these components in plants. Plant Cell Physiol. 2009, 50, 1988–1993. [Google Scholar] [CrossRef] [Green Version]
- Centofanti, T.; Sayers, Z.; Cabello-Conejo, M.I.; Kidd, P.; Nishizawa, N.K.; Kakei, Y.; Davis, A.P.; Sicher, R.C.; Chaney, R.L. Xylem exudate composition and root-to-shoot nickel translocation in Alyssum species. Plant Soil 2013, 373, 59–75. [Google Scholar] [CrossRef]
- Schaumlöffel, D.; Ouerdane, L.; Bouyssiere, B.; Łobiński, R. Speciation analysis of nickel in the latex of a hyperaccumulating tree Sebertia acuminata by HPLC and CZE with ICP MS and electrospray MS-MS detection. J. Anal. At. Spectrom. 2003, 18, 120–127. [Google Scholar] [CrossRef]
- Ouerdane, L.; Mari, S.; Czernic, P.; Lebrun, M.; Łobiński, R. Speciation of non-covalent nickel species in plant tissue extracts by electrospray Q-TOFMS/MS after their isolation by 2D size exclusion-hydrophilic interaction LC (SEC-HILIC) monitored by ICP-MS. J. Anal. At. Spectrom. 2006, 21, 676–683. [Google Scholar] [CrossRef]
- Callahan, D.L.; Kolev, S.D.; O’Hair, R.A.J.; Baker, A.J.M. The detection of nicotianamine in nickel hyperaccumulating plants. In Proceedings of the 8th International Conference on the Biogeochemistry of Trace Elements (ICOBTE); Lombi, E., Ed.; CSIRO Land and Water Adelaide: Adelaide, Australia, 2005; pp. 822–823. [Google Scholar]
- Inoue, H.; Takahashi, M.; Kobayashi, T.; Suzuki, M.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Identification and localisation of the rice nicotianamine aminotransferase gene OsNAAT1 expression suggests the site of phytosiderophore synthesis in rice. Plant Mol. Biol. 2008, 66, 193–203. [Google Scholar] [CrossRef]
- Ando, Y.; Nagata, S.; Yanagisawa, S.; Yoneyama, T. Copper in xylem and phloem saps from rice (Oryza sativa): The effect of moderate copper concentrations in the growth medium on the accumulation of five essential metals and a speciation analysis of copper-containing compounds. Funct. Plant Biol. 2013, 40, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Chao, Z.-F.; Wang, Y.-L.; Chen, Y.-Y.; Zhang, C.-Y.; Wang, P.-Y.; Song, T.; Liu, C.-B.; Lv, Q.-Y.; Han, M.-L.; Wang, S.-S.; et al. NPF transporters in synaptic-like vesicles control delivery of iron and copper to seeds. Sci. Adv. 2021, 7, eabh2450. [Google Scholar] [CrossRef] [PubMed]
- Scholz, G.; Becker, R.; Pich, A.; Stephan, U.W. Nicotianamine—A common constituent of strategies I and II of iron acquisition by plants: A review. J. Plant Nutr. 1992, 15, 1647–1665. [Google Scholar] [CrossRef]
- Hakoyama, T.; Watanabe, H.; Tomita, J.; Yamamoto, A.; Sato, S.; Mori, Y.; Kouchi, H.; Suganuma, N. Nicotianamine synthase specifically expressed in root nodules of Lotus japonicus. Planta 2009, 230, 309–317. [Google Scholar] [CrossRef] [Green Version]
- Douchkov, D.; Gryczka, C.; Stephan, U.W.; Hell, R.; Bäumlein, H. Ectopic expression of nicotianamine synthase genes results in improved iron accumulation and increased nickel tolerance in transgenic tobacco. Plant Cell Environ. 2005, 28, 365–374. [Google Scholar] [CrossRef]
- Kim, S.; Takahashi, M.; Higuchi, K.; Tsunoda, K.; Nakanishi, H.; Yoshimura, E.; Mori, S.; Nishizawa, N.K. Increased nicotianamine biosynthesis confers enhanced tolerance of high levels of metals, in particular nickel, to plants. Plant Cell Physiol. 2005, 46, 1809–1818. [Google Scholar] [CrossRef] [Green Version]
- Pianelli, K.; Mari, S.; Marquès, L.; Lebrun, M.; Czernic, P. Nicotianamine over-accumulation confers resistance to nickel in Arabidopsis thaliana. Transgenic Res. 2005, 14, 739–748. [Google Scholar] [CrossRef]
- Glińska, S.; Gapińska, M.; Michlewska, S.; Skiba, E.; Kubicki, J. Analysis of Triticum aestivum seedling response to the excess of zinc. Protoplasma 2016, 253, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Feigl, G.; Molnár, Á.; Szőllősi, R.; Ördög, A.; Törőcsik, K.; Oláh, D.; Bodor, A.; Perei, K.; Kolbert, Z. Zinc-induced root architectural changes of rhizotron-grown B. napus correlate with a differential nitro-oxidative response. Nitric Oxide 2019, 90, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Hanikenne, M.; Esteves, S.M.; Fanara, S.; Rouached, H. Coordinated homeostasis of essential mineral nutrients: A focus on iron. J. Exp. Bot. 2021, 72, 2136–2153. [Google Scholar] [CrossRef]
- Ziller, A.; Fraissinet-Tachet, L. Metallothionein diversity and distribution in the tree of life: A multifunctional protein. Metallomics 2018, 10, 1549–1559. [Google Scholar] [CrossRef] [PubMed]
- Sterckeman, T.; Thomine, S. Mechanisms of cadmium accumulation in plants. Crit. Rev. Plant Sci. 2020, 39, 322–359. [Google Scholar] [CrossRef]
- Sytar, O.; Ghosh, S.; Malinska, H.; Zivcak, M.; Brestic, M. Physiological and molecular mechanisms of metal accumulation in hyperaccumulator plants. Physiol. Plant. 2021, 173, 148–166. [Google Scholar] [CrossRef] [PubMed]
- Seregin, I.V.; Kozhevnikova, A.D. Phytochelatins: Sulfur-containing metal(loid)-chelating ligands in plants. Int. J. Mol. Sci. 2023, 24, 2430. [Google Scholar] [CrossRef] [PubMed]
- van der Ent, A.; Purwadi, I.; Harris, H.H.; Kopittke, P.M.; Przybyłowicz, W.J.; Mesjasz-Przybyłowicz, J. Methods for visualizing elemental distribution in hyperaccumulator plants. In Agromining: Farming for Metals. Mineral Resource Reviews; van der Ent, A., Baker, A.J., Echevarria, G., Simonnot, M.O., Morel, J.L., Eds.; Springer: Cham, Switzerland, 2021; pp. 197–214. [Google Scholar] [CrossRef]
- Clemens, S. The cell biology of zinc. J. Exp. Bot. 2022, 73, 1688–1698. [Google Scholar] [CrossRef]
- Bashmakova, E.B.; Pashkovskiy, P.P.; Radyukina, N.L.; Kuznetsov, V.V. Possible mechanism of iron deficit development in Mimulus guttatus plants exposed to joint action of nickel and zinc salts. Russ. J. Plant Physiol. 2015, 62, 761–771. [Google Scholar] [CrossRef]
- Kozhevnikova, A.D.; Seregin, I.V.; Schat, H. Translocation of Ni and Zn in Odontarrhena corsica and Noccaea caerulescens: The effects of exogenous histidine and Ni/Zn interactions. Plant Soil 2021, 468, 295–318. [Google Scholar] [CrossRef]
- Negi, S.; Singh, D.; Pandey, H.; Singhal, R.K. Biofortification for high Fe and Zn in various Poaceae crops by using different molecular breeding and biotechnological approaches. Plant Physiol. Rep. 2021, 26, 636–646. [Google Scholar] [CrossRef]
- Harrington, S.A.; Connorton, J.M.; Nyangoma, N.I.M.; McNelly, R.; Morgan, Y.M.L.; Aslam, M.F.; Sharp, P.A.; Johnson, A.A.T.; Uauy, C.; Balk, J. A two-gene strategy increases iron and zinc concentrations in wheat flour, improving mineral bioaccessibility. Plant Physiol. 2023, 191, 528–541. [Google Scholar] [CrossRef]
- Clemens, S. Zn and Fe biofortification: The right chemical environment for human bioavailability. Plant Sci. 2014, 225, 52–57. [Google Scholar] [CrossRef]
- Wairich, A.; Ricachnevsky, F.K.; Lee, S. A tale of two metals: Biofortification of rice grains with iron and zinc. Front. Plant Sci. 2022, 13, 944624. [Google Scholar] [CrossRef] [PubMed]
- Connorton, J.M.; Balk, J. Iron biofortification of staple crops: Lessons and challenges in plant genetics. Plant Cell Physiol. 2019, 60, 1447–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beasley, J.T.; Johnson, A.A.T.; Kolba, N.; Bonneau, J.P.; Glahn, R.P.; Ozeri, L.; Koren, O.; Tako, E. Nicotianamine-chelated iron positively affects iron status, intestinal morphology and microbial populations in vivo (Gallus gallus). Sci. Rep. 2020, 10, 2297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beasley, J.T.; Bonneau, J.P.; Moreno-Moyano, L.T.; Callahan, D.L.; Howell, K.S.; Tako, E.; Taylor, J.; Glahn, R.P.; Appels, R.; Johnson, A.A. Multi-year field evaluation of nicotianamine biofortified bread wheat. Plant J. 2022, 109, 1168–1182. [Google Scholar] [CrossRef]
- Murata, Y.; Yoshida, M.; Sakamoto, N.; Morimoto, S.; Watanabe, T.; Namba, K. Iron uptake mediated by the plant-derived chelator nicotianamine in the small intestine. J. Biol. Chem. 2021, 296, 100195. [Google Scholar] [CrossRef]
- Takada, N.; Tanaka, M.; Ishidoya, A.; Hayakari, M. Nicotianamine-rich foods for improving the learning and memory functions and as an effective preventing agent of hypertension. Food Biosci. 2020, 37, 100729. [Google Scholar] [CrossRef]
- Singh, S.P.; Keller, B.; Gruissem, W.; Bhullar, N.K. Rice NICOTIANAMINE SYNTHASE 2 expression improves dietary iron and zinc levels in wheat. Theor. Appl. Genet. 2017, 130, 283–292. [Google Scholar] [CrossRef] [Green Version]
- Nozoye, T. The nicotianamine synthase gene is a useful candidate for improving the nutritional qualities and Fe-deficiency tolerance of various crops. Front. Plant Sci. 2018, 9, 340. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Rahman, M.M.; Nakanishi, H.; Nishizawa, N.K.; An, G.; Nam, H.G.; Jeon, J.-S. Concomitant activation of OsNAS2 and OsNAS3 contributes to the enhanced accumulation of iron and zinc in rice. Int. J. Mol. Sci. 2023, 24, 6568. [Google Scholar] [CrossRef]
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Chunduri, V.; Arora, P. Biofortified crops generated by breeding, agronomy, and transgenic approaches are improving lives of millions of people around the world. Front. Nutr. 2018, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; Usuda, K.; Kobayashi, T.; Ishimaru, Y.; Kakei, Y.; Takahashi, M.; Higuchi, K.; Nakanishi, H.; Mori, S.; Nishizawa, N.K. Overexpression of the barley nicotianamine synthase gene HvNAS1 increases iron and zinc concentrations in rice grains. Rice 2009, 2, 155–166. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; Cheng, Z.; Ai, C.; Jiang, X.; Bei, X.; Zheng, Y.; Glahn, R.P.; Welch, R.M.; Miller, D.D.; Lei, X.G.; et al. Nicotianamine, a novel enhancer of rice iron bioavailability to humans. PLoS ONE 2010, 5, e10190. [Google Scholar] [CrossRef] [PubMed]
- Wirth, J.; Poletti, S.; Aeschlimann, B.; Yakandawala, N.; Drosse, B.; Osorio, S.; Tohge, T.; Fernie, A.R.; Günther, D.; Gruissem, W.; et al. Rice endosperm iron biofortification by targeted and synergistic action of nicotianamine synthase and ferritin. Plant Biotechnol. J. 2009, 7, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Trijatmiko, K.R.; Duenãs, C.; Tsakirpaloglou, N.; Torrizo, L.; Arines, F.M.; Adeva, C.; Balindong, J.; Oliva, N.; Sapasap, M.V.; Borrero, J.; et al. Biofortified indica rice attains iron and zinc nutrition dietary targets in the field. Sci. Rep. 2016, 6, 19792. [Google Scholar] [CrossRef] [Green Version]
- Tsakirpaloglou, N.; Bueno-Mota, G.M.; Soriano, J.C.; Arcillas, E.; Arines, F.M.; Yu, S.-M.; Stangoulis, J.; Trijatmiko, K.R.; Reinke, R.; Tohme, J.; et al. Proof of concept and early development stage of market-oriented high iron and zinc rice expressing dicot ferritin and rice nicotianamine synthase genes. Sci. Rep. 2023, 13, 676. [Google Scholar] [CrossRef] [PubMed]
- Banakar, R.; Fernandez, A.A.; Díaz-Benito, P.; Abadia, J.; Capell, T.; Christou, P. Phytosiderophores determine thresholds for iron and zinc accumulation in biofortified rice endosperm while inhibiting the accumulation of cadmium. J. Exp. Bot. 2017, 68, 4983–4995. [Google Scholar] [CrossRef] [Green Version]
- Krishna, T.P.A.; Maharajan, T.; Ceasar, S.A. The role of membrane transporters in the biofortification of zinc and iron in plants. Biol. Trace Elem. Res. 2023, 201, 464–478. [Google Scholar] [CrossRef]
- Zha, M.; Li, X.; Li, R.; Huang, J.; Fan, J.; Zhang, J.; Wang, Y.; Zhang, C. Overexpression of nicotianamine synthase (AtNAS1) increases iron accumulation in the tuber of potato. Plants 2022, 11, 2741. [Google Scholar] [CrossRef]
- Suman, J.; Uhlik, O.; Viktorova, J.; Macek, T. Phytoextraction of heavy metals: A promising tool for clean-up of polluted environment? Front. Plant Sci. 2018, 9, 1476. [Google Scholar] [CrossRef] [Green Version]
- Gunarathne, V.; Mayakaduwa, S.; Ashiq, A.; Weerakoon, S.R.; Biswas, J.K.; Vithanage, M. Transgenic Plants: Benefits, applications, and potential risks in phytoremediation. In Transgenic Plant Technology for Remediation of Toxic Metals and Metalloids; Elsevier: Amsterdam, the Netherlands, 2018; pp. 89–102. [Google Scholar]


{kind=link}
| Family | Species | Number of NAS Genes | References |
|---|---|---|---|
| Brassicaceae | Arabidopsis thaliana | 4 | [87,88,89] |
| Arabidopsis halleri | 4 | [90,91] | |
| Noccaea caerulescens | 4 | [84,87,88] | |
| Crassulaceae | Sedum alfredii | 5 | [92,93] |
| Euphorbiaceae | Ricinus communis | 3 | [94] |
| Poaceae | Aegilops speltoides | 5 | [86] |
| Aegilops tauschii | 5 | [86] | |
| Hordeum vulgare | 9 | [95,96] | |
| Oryza sativa | 3 | [95,97,98,99] | |
| Triticum aestivum (hexaploid varieties) | 20–21 | [85,86] | |
| Triticum monococcum | 8 | [100] | |
| Triticum urartu | 5 | [86] | |
| Zea mays | 9–10 | [101,102,103] | |
| Rosaceae | Malus xiaojinensis | 3 | [104,105,106] |
| Solanaceae | Solanum lycopersicum | 1 | [107] |
| Species | Transporters | Metal Transporting Capacity and Functional Role | Localization of the Transporter Gene Expression or the Transporter Itself | References |
|---|---|---|---|---|
| Poaceae | ||||
| Brachypodium distachyon | BdYSL3 | Cu in ionic forms; loading into the phloem and transport into generative organs | The expression was significantly upregulated by Cu starvation in roots, stems, mature leaves, flag leaves, and reproductive organs, mainly in the phloem of leaves and nodes. | [157,159] |
| Hordeum vulgare | HvYS1 | Fe(III) –DMA, Fe(III) –MA; Fe uptake | The expression was enhanced under Fe deficiency in root rhizodermal cells. | [160,161,162] |
| HvYSL2 | Fe(III)/Zn/Mn/Cu(II)/Co/Ni–DMA, Fe(II) –NA; metal transport | The expression was detected in root endodermis and shoots. The expression in roots was induced by Fe deficiency. | [154] | |
| Miscanthus sacchariflorus | MsYSL1 | Fe(II) –NA, Zn–NA | The expression was detected throughout the whole seedlings, with the highest level in the stem. The expression was enhanced in roots under Mn, Cd, and Pb excess as well as under Fe, Zn, and Cu deficiency. | [149] |
| Oryza sativa | OsYSL2 | Fe(II)–NA, Mn(II)–NA; Fe and Mn transport via the phloem, including translocation into the grains | The expression was induced by Fe deficiency in the phloem cells, especially in the companion cells in leaves, and in mature grains in the epithelium, the vascular bundle of the scutellum, leaf primordium, in the bran and coleoptile. | [116,150,163] |
| OsYSL6 | Mn(II)–NA; Mn transport | The expression was detected in all root and shoot tissues irrespective of metal deficiency or toxicity. | [153] | |
| OsYSL9 | Fe(III)–DMA, Fe(II)–NA; Fe transport from the endosperm into the embryo in developing kernels | The expression was induced in the vascular cylinder of roots but repressed in the non-juvenile leaves in response to Fe deficiency. At the grain filling stage, the expression was induced in the scutellum of the embryo and in the endosperm cells surrounding the embryo. | [164] | |
| OsYSL13 | Fe transport and redistribution, Fe translocation to the youngest leaves and seeds | The gene was highly expressed in leaves, especially in leaf blades, as well as in both Fe-sufficient and Fe-deficient root cortex. Gene expression was induced by Fe deficiency both in roots and shoots. | [151,156] | |
| OsYSL15 | Fe(III) –DMA, Fe(II) –NA; Fe uptake and transport via the phloem | Gene expression in roots was induced by Fe deficiency, was predominant in the rhizodermis / exodermis and phloem cells under Fe deficiency and was detected only in the phloem under Fe sufficiency. Gene expression was also shown in the flowers, developing seeds, and in the embryonic scutellar epithelial cells during seed germination. | [151,158,165] | |
| OsYSL16 | Fe(III)–DMA, Cu–NA; long-distance transport of metals and metal redistribution, including that of one to the floral organs | Gene expression was detected in the root rhizodermis, root and shoot conducting tissues, leaves, rachilla, palea, lemma, anther and ovary, and was induced by Fe deficiency. | [166,167,168] | |
| OsYSL18 | Fe(III)–DMA; Fe root-to-shoot transport, translocation into generative organs, metal redistribution | Gene expression was detected in generative organs including the pollen tube, and in vegetative organs in lamina joints, the inner cortex of crown roots, in phloem parenchyma and companion cells at the basal part of every leaf sheath. Gene expression in roots and discrimination center was induced by Fe excess. | [152] | |
| Zea mays | ZmYS1 | Fe(III)/Fe(II)/Cu(II)/ Mn(II)/Ni/Zn/Cd– DMA, Fe(II)/Fe(III)/Ni–NA; metal uptake | Gene expression was localized in root rhizodermal cells and shoots under Fe deficiency. | [35,169,170,171] |
| ZmYSL2 | Fe–NA, Zn–NA; redistribution of Fe and Zn in kernels | The transporter was localized in the maternal tissue of the basal endosperm transfer cell layer, in aleurone, sub-aleurone, and embryo cells. | [172,173,174] | |
| Brassicaceae | ||||
| Arabidopsis thaliana | AtYSL1 | Fe(II)–NA; Fe long-distance transport and translocation to the seeds | The expression was detected in roots, leaves, flowers, pollen, young siliques, and seeds. The expression in the xylem parenchyma of leaves was upregulated in response to Fe excess. The expression in shoots decreased under Fe deficiency. | [175,176,177] |
| AtYSL2 | Fe(II)–NA, Cu(II)–NA; Fe and Cu transport | Gene expression was found in the endodermal and pericycle cells opposite to the metaxylem vessels, in root and shoot conductive tissues, and particularly in xylem parenchyma. | [178,179] | |
| AtYSL3 | Fe(II)–NA; Fe long-distance transport | Gene expression was detected in the vasculature of roots and shoots as well as in flowers. The expression in shoots decreased under Fe deficiency. | [176,177,180] | |
| Noccaea caerulescens | NcYSL3 | Fe(II)–NA, Ni–NA; Fe and Ni transport | The gene was expressed constitutively in all organs, mainly in root central cylinder cells. | [181] |
| Fabaceae | ||||
| Arachis hypogaea | AhYSL1 | Fe(III)–DMA; absorption of Fe(III)–DMA (DMA is secreted by neighboring cereals) | The expression of this gene in the root rhizodermis was induced by Fe deficiency. | [182] |
| AhYSL3.1 | Fe(III)–DMA, Fe(II)–NA, Cu–NA; mainly Cu transport | The expression of this gene was detected in the main or lateral roots, but not the root tips, around the conducting tissues, and was upregulated by Cu deficiency. | [155] | |
| Solanaceae | ||||
| Solanum nigrum | SnYSL3 | Fe(II)/Cu/Zn/Cd–NA; metal transport | The expression was detected predominantly in the root rhizodermis and stem epidermis, as well as in conducting tissues of roots, stems and leaves, and was up-regulated by Cd and Fe excess as well as Cu deficiency. | [183] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seregin, I.V.; Kozhevnikova, A.D. Nicotianamine: A Key Player in Metal Homeostasis and Hyperaccumulation in Plants. Int. J. Mol. Sci. 2023, 24, 10822. https://doi.org/10.3390/ijms241310822
Seregin IV, Kozhevnikova AD. Nicotianamine: A Key Player in Metal Homeostasis and Hyperaccumulation in Plants. International Journal of Molecular Sciences. 2023; 24(13):10822. https://doi.org/10.3390/ijms241310822
Chicago/Turabian StyleSeregin, Ilya V., and Anna D. Kozhevnikova. 2023. "Nicotianamine: A Key Player in Metal Homeostasis and Hyperaccumulation in Plants" International Journal of Molecular Sciences 24, no. 13: 10822. https://doi.org/10.3390/ijms241310822