


Assembly of Synaptic Protein–DNA Complexes: Critical Role of Non-Specific Interactions

Abstract
:1. Introduction
2. Results
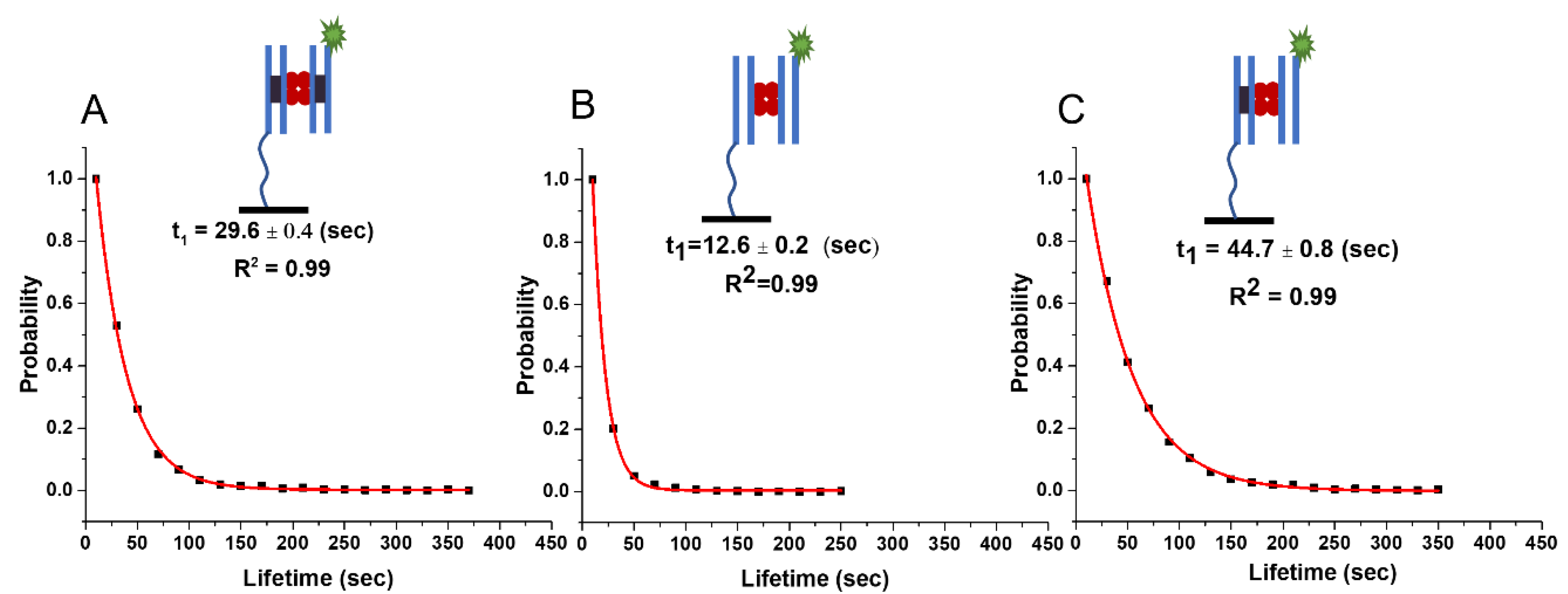
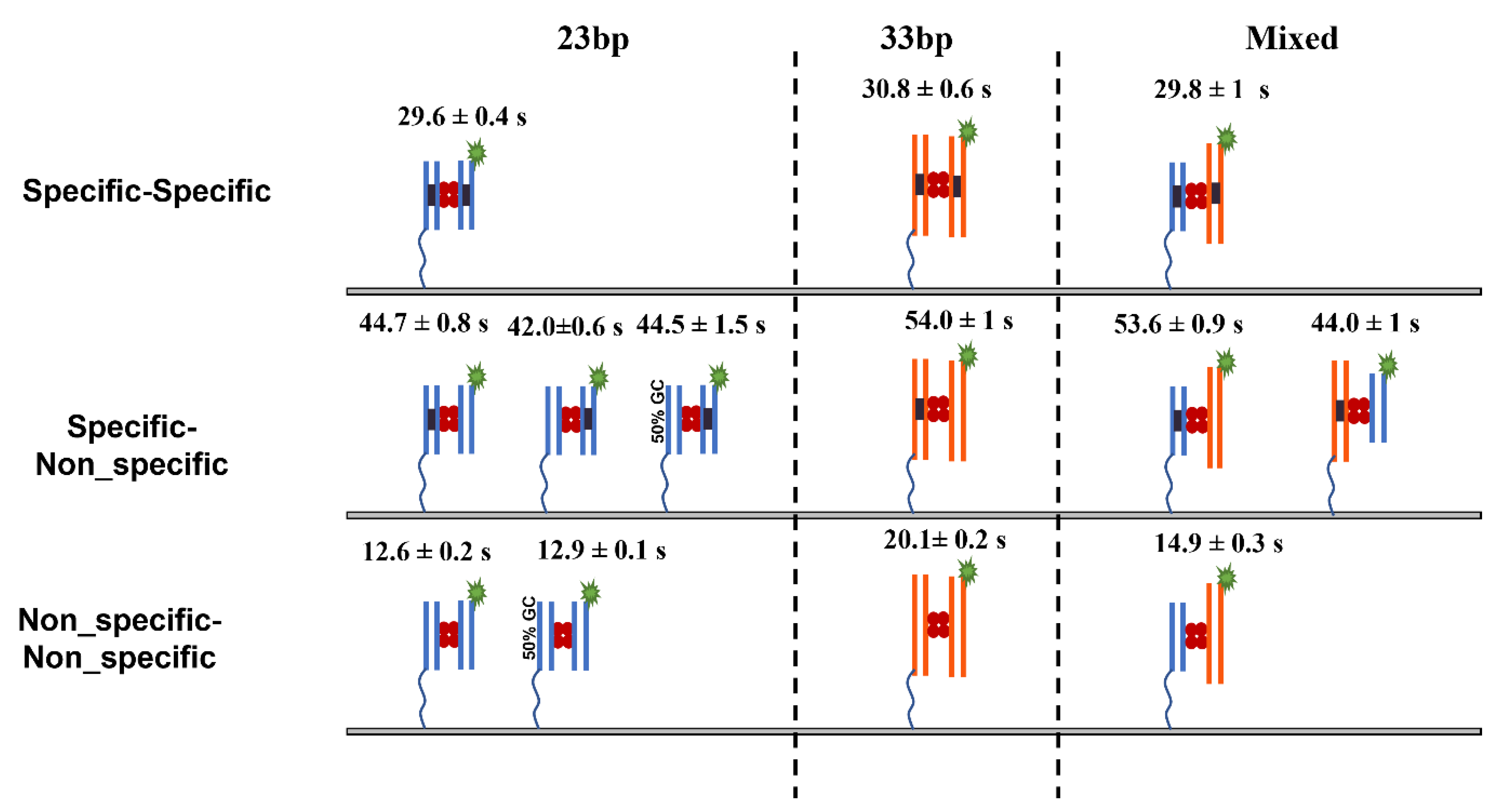
2.1. Lifetime of SfiI–DNA Complexes Using 23 bp Substrates
2.1.1. Lifetime of Synaptic Specific Complexes (ss)
2.1.2. Lifetime of Non-Specific Complexes (nn)
2.1.3. Lifetime of Pre-Synaptic Non-Specific–Specific Complexes (ns)
2.2. Theoretical Model for SfiI–DNA Complex Lifetimes
- (1)
- After the bond is broken with the first duplex, we assume that the specific/specific (ss) complex can only return to the ss complex, and no non-specific bonds between protein and DNA molecules can be formed.
- (2)
- The non-specific/specific (ns) pre-synaptic complex first breaks the bond with the non-specific duplex (the weakest one), and after that, it will have multiple possibilities to return to ns complex due to the creation of 23 − 13 + 1 = 11 possible bonds with the non-specific duplex. This also reflects the possibility of sliding in the ns conformation. We call this effect an effective entropic effect. Our hypothesis is that this is the primary source of the larger lifetimes for ns complexes compared to ss complexes. Although a single ns interaction is weaker than the ss interactions, the number of such interactions due to different locations are cumulative, making the overall ns interaction stronger.
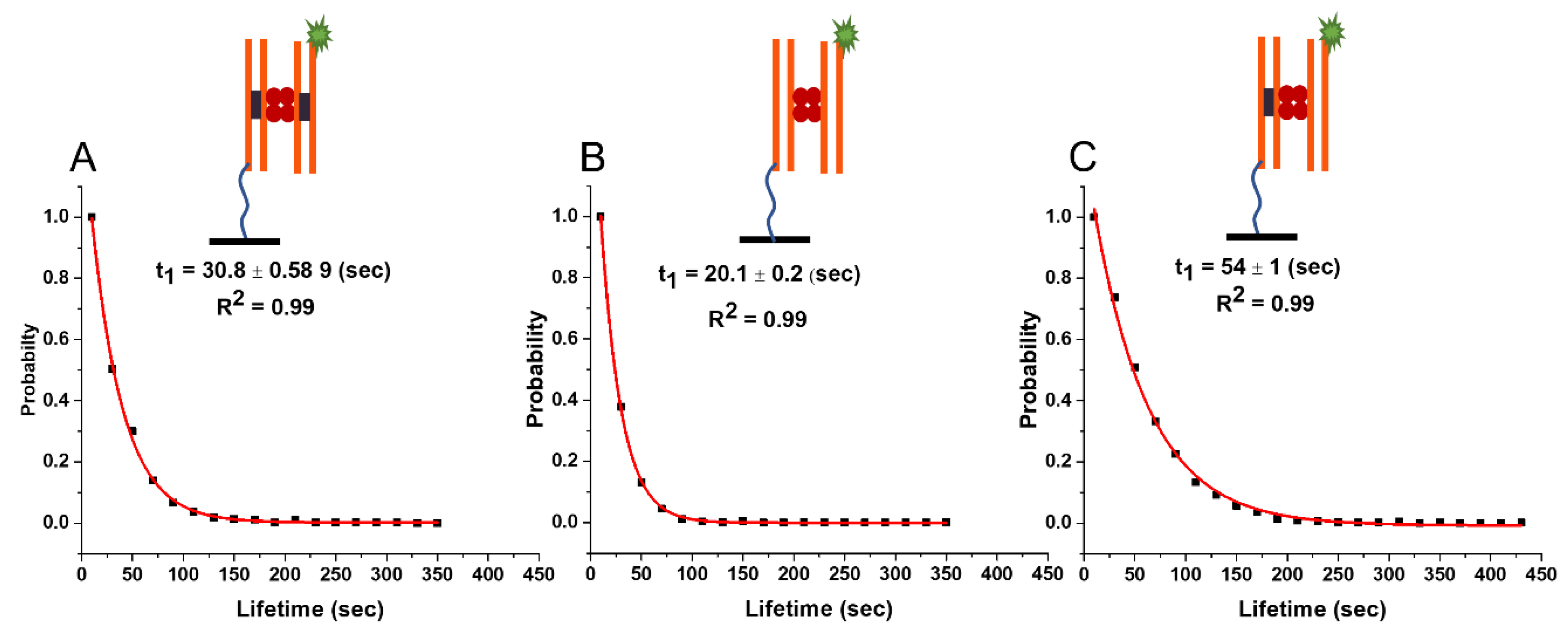
2.3. Experiment: Lifetime of SfiI–DNA Complexes with 33 bp DNA Substrates
2.3.1. SfiI–DNA Complexes with Symmetric Substrates
2.3.2. Lifetime of Synaptic Complexes (ss)
2.3.3. Lifetime of the Non-Specific Complexes (nn)
2.3.4. Lifetime of the Pre-Synaptic Complexes (ns)
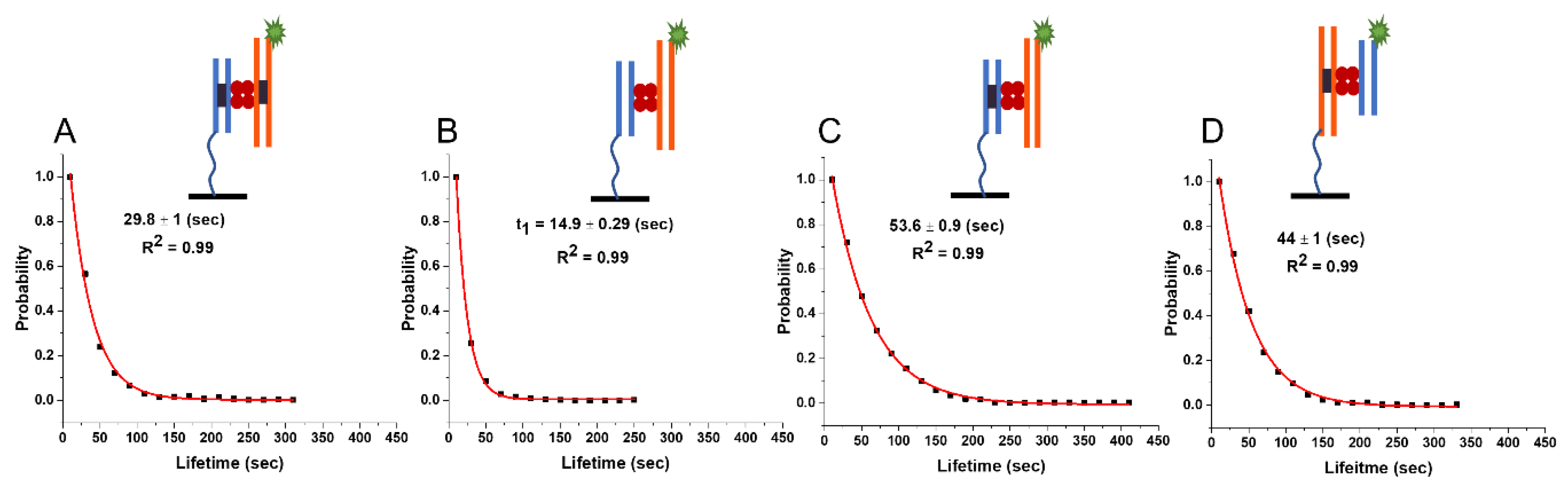
2.4. SfiI–DNA Complexes with Asymmetric Substrates
2.4.1. Lifetime of Synaptic Complexes (ss)
2.4.2. Lifetime of the Non-Specific Complexes (nn)
2.4.3. Lifetime of the Pre-Synaptic Complexes (ns)
3. Discussion
4. Materials and Methods
4.1. Preparation of DNA Duplexes
4.2. Annealing and Melting of DNA Duplexes
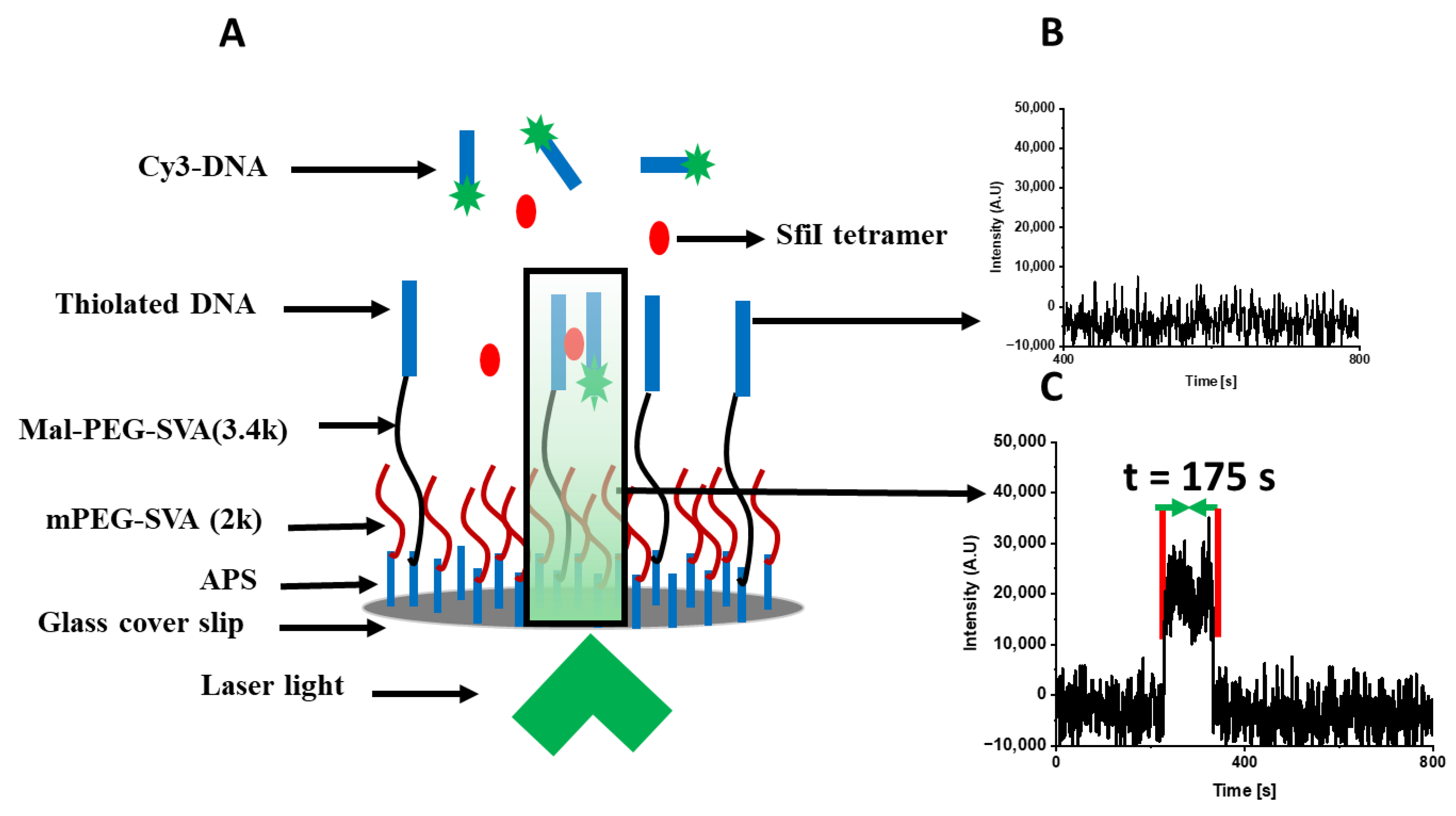
4.3. Single-Molecule Fluorescence with TAPIN Approach
4.3.1. Surface Modification
4.3.2. TAPIN Data Acquisition of SfiI–DNA Complexes
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Grindley, N.D.; Whiteson, K.L.; Rice, P.A. Mechanisms of site-specific recombination. Annu. Rev. Biochem. 2006, 75, 567–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Virnik, K.; Lyubchenko, Y.L.; Karymov, M.A.; Dahlgren, P.; Tolstorukov, M.Y.; Semsey, S.; Zhurkin, V.B.; Adhya, S. “Antiparallel” DNA loop in gal repressosome visualized by atomic force microscopy. J. Mol. Biol. 2003, 334, 53–63. [Google Scholar] [CrossRef]
- Lyubchenko, Y.L.; Shlyakhtenko, L.S.; Aki, T.; Adhya, S. Atomic force microscopic demonstration of DNA looping by GalR and HU. Nucleic Acids Res. 1997, 25, 873–876. [Google Scholar] [CrossRef] [PubMed]
- Vetcher, A.A.; Lushnikov, A.Y.; Navarra-Madsen, J.; Scharein, R.G.; Lyubchenko, Y.L.; Darcy, I.K.; Levene, S.D. DNA topology and geometry in Flp and Cre recombination. J. Mol. Biol. 2006, 357, 1089–1104. [Google Scholar] [CrossRef] [PubMed]
- Hickman, A.B.; Dyda, F. Mechanisms of DNA Transposition. Microbiol. Spectr. 2015, 3, Mdna3-0034-2014. [Google Scholar] [CrossRef] [Green Version]
- Shlyakhtenko, L.S.; Gilmore, J.; Kriatchko, A.N.; Kumar, S.; Swanson, P.C.; Lyubchenko, Y.L. Molecular mechanism underlying RAG1/RAG2 synaptic complex formation. J. Biol. Chem. 2009, 284, 20956–20965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craigie, R. The molecular biology of HIV integrase. Future Virol. 2012, 7, 679–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghavendra, N.K.; Engelman, A. LEDGF/p75 interferes with the formation of synaptic nucleoprotein complexes that catalyze full-site HIV-1 DNA integration in vitro: Implications for the mechanism of viral cDNA integration. Virology 2007, 360, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Halford, S.E.; Marko, J.F. How do site-specific DNA-binding proteins find their targets? Nucleic Acids Res. 2004, 32, 3040–3052. [Google Scholar] [CrossRef] [Green Version]
- Halford, S.E. Hopping, Jumping and Looping by Restriction Enzymes; Portland Press Ltd.: London, UK, 2001. [Google Scholar]
- Berg, O.G.; Blomberg, C. Association kinetics with coupled diffusional flows: Special application to the lac repressor-operator system. Biophys. Chem. 1976, 4, 367–381. [Google Scholar] [CrossRef]
- Berg, O.G.; Blomberg, C. Association kinetics with coupled diffusion. An extension to coiled-chain macromolecules applied to the lac repressor-operator system. Biophys. Chem. 1977, 7, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Berg, O.G.; Blomberg, C. Association kinetics with coupled diffusion: III. Ionic-strength dependence of the lac repressor-operator association. Biophys. Chem. 1978, 8, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Berg, O.G.; Winter, R.B.; Von Hippel, P.H. Diffusion-driven mechanisms of protein translocation on nucleic acids. 1. Models and theory. Biochemistry 1981, 20, 6929–6948. [Google Scholar] [CrossRef] [PubMed]
- Winter, R.B.; Berg, O.G.; Von Hippel, P.H. Diffusion-driven mechanisms of protein translocation on nucleic acids. 3. The Escherichia coli lac repressor-operator interaction: Kinetic measurements and conclusions. Biochemistry 1981, 20, 6961–6977. [Google Scholar] [CrossRef] [PubMed]
- Karymov, M.A.; Krasnoslobodtsev, A.V.; Lyubchenko, Y.L. Dynamics of synaptic SfiI-DNA complex: Single-molecule fluorescence analysis. Biophys. J. 2007, 92, 3241–3250. [Google Scholar] [CrossRef] [Green Version]
- Krasnoslobodtsev, A.V.; Shlyakhtenko, L.S.; Lyubchenko, Y.L. Probing interactions within the synaptic DNA-SfiI complex by AFM force spectroscopy. J. Mol. Biol. 2007, 365, 1407–1416. [Google Scholar] [CrossRef] [Green Version]
- Lushnikov, A.Y.; Potaman, V.N.; Oussatcheva, E.A.; Sinden, R.R.; Lyubchenko, Y.L. DNA Strand Arrangement within the SfiI− DNA Complex: Atomic Force Microscopy Analysis. Biochemistry 2006, 45, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Nobbs, T.J.; Szczelkun, M.D.; Wentzell, L.M.; Halford, S.E. DNA excision by the SfiI restriction endonuclease. J. Mol. Biol. 1998, 281, 419–432. [Google Scholar] [CrossRef]
- Suzuki, Y.; Gilmore, J.L.; Yoshimura, S.H.; Henderson, R.M.; Lyubchenko, Y.L.; Takeyasu, K. Visual analysis of concerted cleavage by type IIF restriction enzyme SfiI in subsecond time region. Biophys. J. 2011, 101, 2992–2998. [Google Scholar] [CrossRef] [Green Version]
- Vanamee, E.S.; Viadiu, H.; Kucera, R.; Dorner, L.; Picone, S.; Schildkraut, I.; Aggarwal, A.K. A view of consecutive binding events from structures of tetrameric endonuclease SfiI bound to DNA. EMBO J. 2005, 24, 4198–4208. [Google Scholar] [CrossRef] [Green Version]
- Vemulapalli, S.; Hashemi, M.; Kolomeisky, A.B.; Lyubchenko, Y.L. DNA Looping Mediated by Site-Specific SfiI–DNA Interactions. J. Phys. Chem. B 2021, 125, 4645–4653. [Google Scholar] [CrossRef] [PubMed]
- Wentzell, L.M.; Nobbs, T.J.; Halford, S.E. TheSfiI Restriction Endonuclease Makes a Four-strand DNA Break at Two Copies of its Recognition Sequence. J. Mol. Biol. 1995, 248, 581–595. [Google Scholar] [CrossRef] [PubMed]
- Vemulapalli, S.; Hashemi, M.; Lyubchenko, Y.L. Site-Search Process for Synaptic Protein-DNA Complexes. Int. J. Mol. Sci. 2022, 23, 212. [Google Scholar] [CrossRef] [PubMed]
- Shlyakhtenko, L.S.; Gilmore, J.; Portillo, A.; Tamulaitis, G.; Siksnys, V.; Lyubchenko, Y.L. Direct visualization of the EcoRII−DNA triple synaptic complex by atomic force microscopy. Biochemistry 2007, 46, 11128–11136. [Google Scholar] [CrossRef]
- Gilmore, J.L.; Suzuki, Y.; Tamulaitis, G.; Siksnys, V.; Takeyasu, K.; Lyubchenko, Y.L. Single-molecule dynamics of the DNA− Eco RII protein complexes revealed with high-speed atomic force microscopy. Biochemistry 2009, 48, 10492–10498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Z.; Krasnoslobodtsev, A.V.; Zhang, Y.; Ysselstein, D.; Rochet, J.C.; Blanchard, S.C.; Lyubchenko, Y.L. Direct Detection of α-Synuclein Dimerization Dynamics: Single-Molecule Fluorescence Analysis. Biophys. J. 2015, 108, 2038–2047. [Google Scholar] [CrossRef] [Green Version]
- Maity, S.; Hashemi, M.; Lyubchenko, Y.L. Nano-assembly of amyloid β peptide: Role of the hairpin fold. Sci. Rep. 2017, 7, 2344. [Google Scholar] [CrossRef]
- Maity, S.; Pramanik, A.; Lyubchenko, Y.L. Probing Intermolecular Interactions within the Amyloid β Trimer Using a Tethered Polymer Nanoarray. Bioconjug. Chem. 2018, 29, 2755–2762. [Google Scholar] [CrossRef]
- Maity, S.; Viazovkina, E.; Gall, A.; Lyubchenko, Y.L. Single-molecule probing of amyloid nano-ensembles using the polymer nanoarray approach. Phys. Chem. Chem. Phys. 2017, 19, 16387–16394. [Google Scholar] [CrossRef]
- Dodge, Y. The Concise Encyclopedia of Statistics; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Blainey, P.C.; Luo, G.; Kou, S.; Mangel, W.F.; Verdine, G.L.; Bagchi, B.; Xie, X.S. Nonspecifically bound proteins spin while diffusing along DNA. Nat. Struct. Mol. Biol. 2009, 16, 1224–1229. [Google Scholar] [CrossRef] [Green Version]
- Davidson, I.F.; Barth, R.; Zaczek, M.; van der Torre, J.; Tang, W.; Nagasaka, K.; Janissen, R.; Kerssemakers, J.; Wutz, G.; Dekker, C.; et al. CTCF is a DNA-tension-dependent barrier to cohesin-mediated loop extrusion. Nature 2023, 616, 822–827. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Complex Type | Experiment 23 bp/23 bp | Experiment 33 bp/33 bp | Experiment 33 bp/23 bp (Fluo) | Theory 33 bp | Theory 23 bp/33 bp |
|---|---|---|---|---|---|
| Specific (ss) | 29.6 ± 0.4 s | 30.8 ± 0.6 s | 29.8 ± 1 s | 30.1 s | 30 s |
| Presynaptic (ns) | 44.7 ± 0.8 s | 54.1 ± 0.1 s | 44 ± 1.2 s | 58 s | 58 s/36 s |
| Non-specific (nn) | 12.6 ± 0.2 s | 20.1 ± 0.2 s | 14.9 ± 0.3 s | 20 s | 16 s |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vemulapalli, S.; Hashemi, M.; Kolomeisky, A.B.; Lyubchenko, Y.L. Assembly of Synaptic Protein–DNA Complexes: Critical Role of Non-Specific Interactions. Int. J. Mol. Sci. 2023, 24, 9800. https://doi.org/10.3390/ijms24129800
Vemulapalli S, Hashemi M, Kolomeisky AB, Lyubchenko YL. Assembly of Synaptic Protein–DNA Complexes: Critical Role of Non-Specific Interactions. International Journal of Molecular Sciences. 2023; 24(12):9800. https://doi.org/10.3390/ijms24129800
Chicago/Turabian StyleVemulapalli, Sridhar, Mohtadin Hashemi, Anatoly B. Kolomeisky, and Yuri L. Lyubchenko. 2023. "Assembly of Synaptic Protein–DNA Complexes: Critical Role of Non-Specific Interactions" International Journal of Molecular Sciences 24, no. 12: 9800. https://doi.org/10.3390/ijms24129800