
Calcium Handling Remodeling Underlies Impaired Sympathetic Stress Response in Ventricular Myocardium from Cacna1c Haploinsufficient Rats
 ,
,  , , , , ,
, , , , ,  and
and 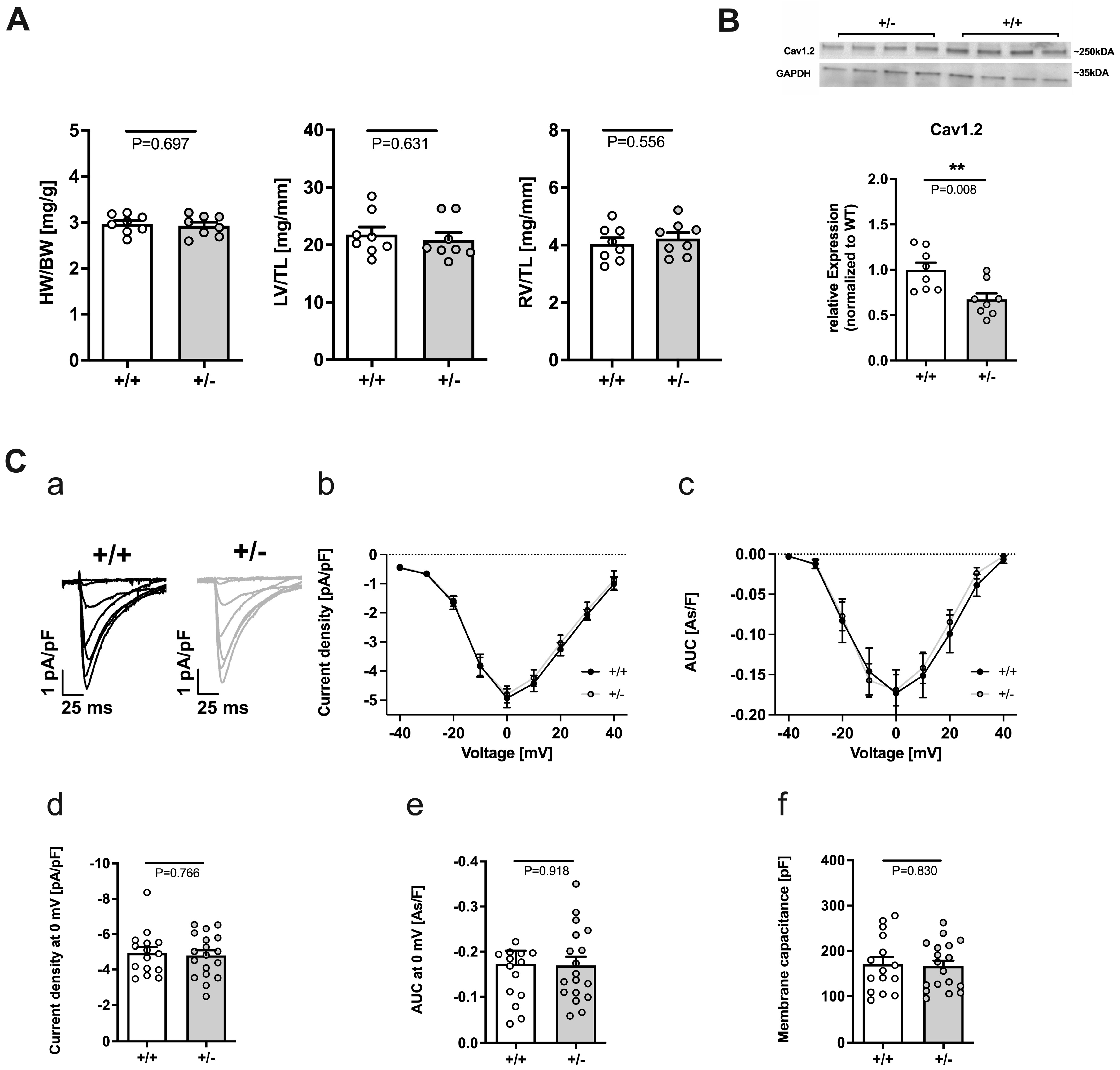{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Cacna1c+/− Rats Exhibit No Signs of Hypo- or Hypertrophy
2.2. Calcium Handling in Cacna1c+/− Ventricular Myocardium under Baseline Conditions
2.2.1. Reduced Expression of Cav1.2 but Unaltered L-Type Ca2+ Current in Cacna1c+/−
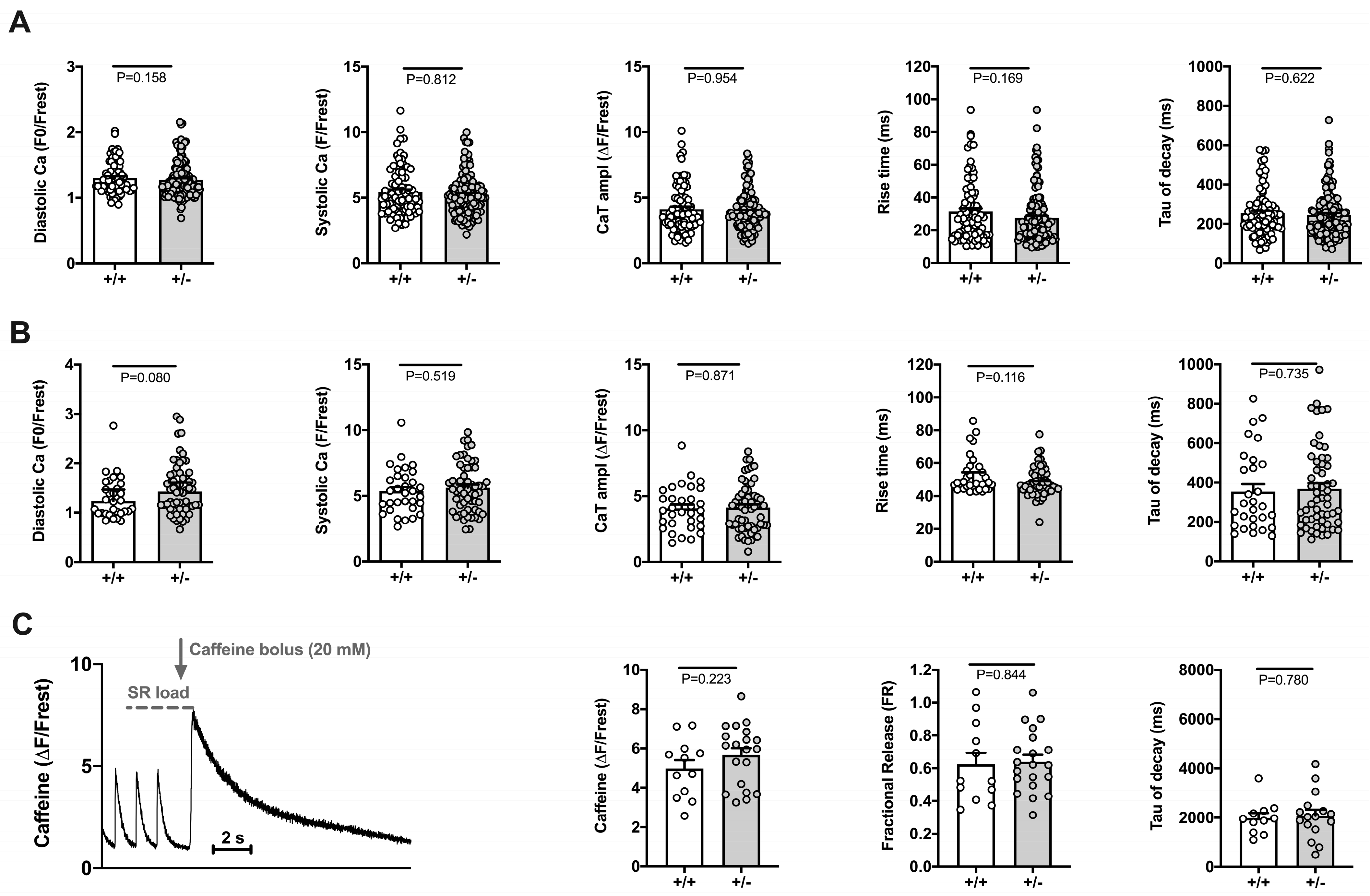
2.2.2. Electrically Stimulated Ca2+ Transients and Sarcomere Shortenings Are Unaltered in Cacna1c+/− Ventricular Myocytes
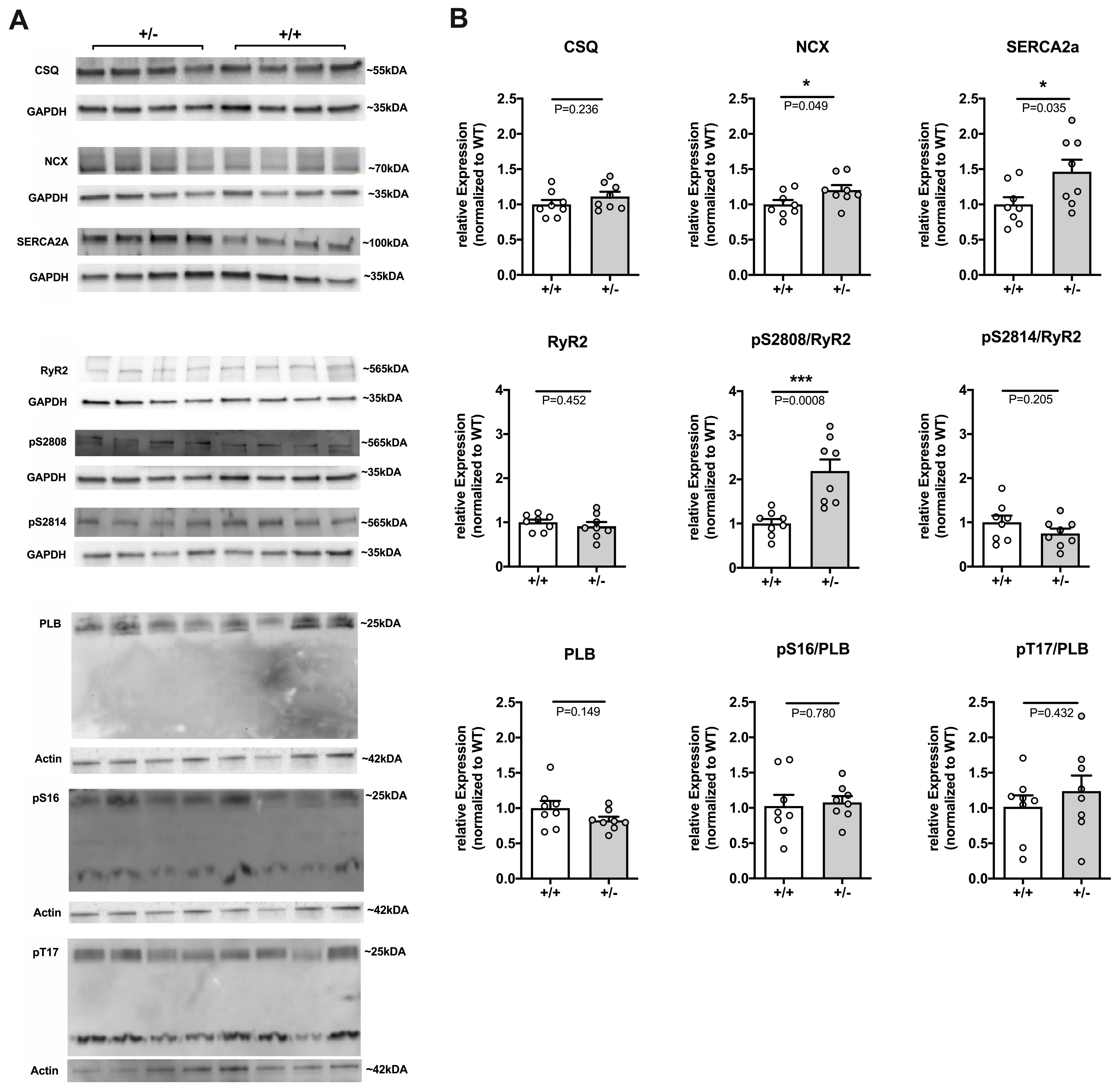
2.2.3. Remodeling of Major Ca2+ Handling Proteins in Cacna1c+/− LV Myocardium
2.3. Calcium Handling in Cacna1c+/− Ventricular Myocardium during Sympathetic Stress
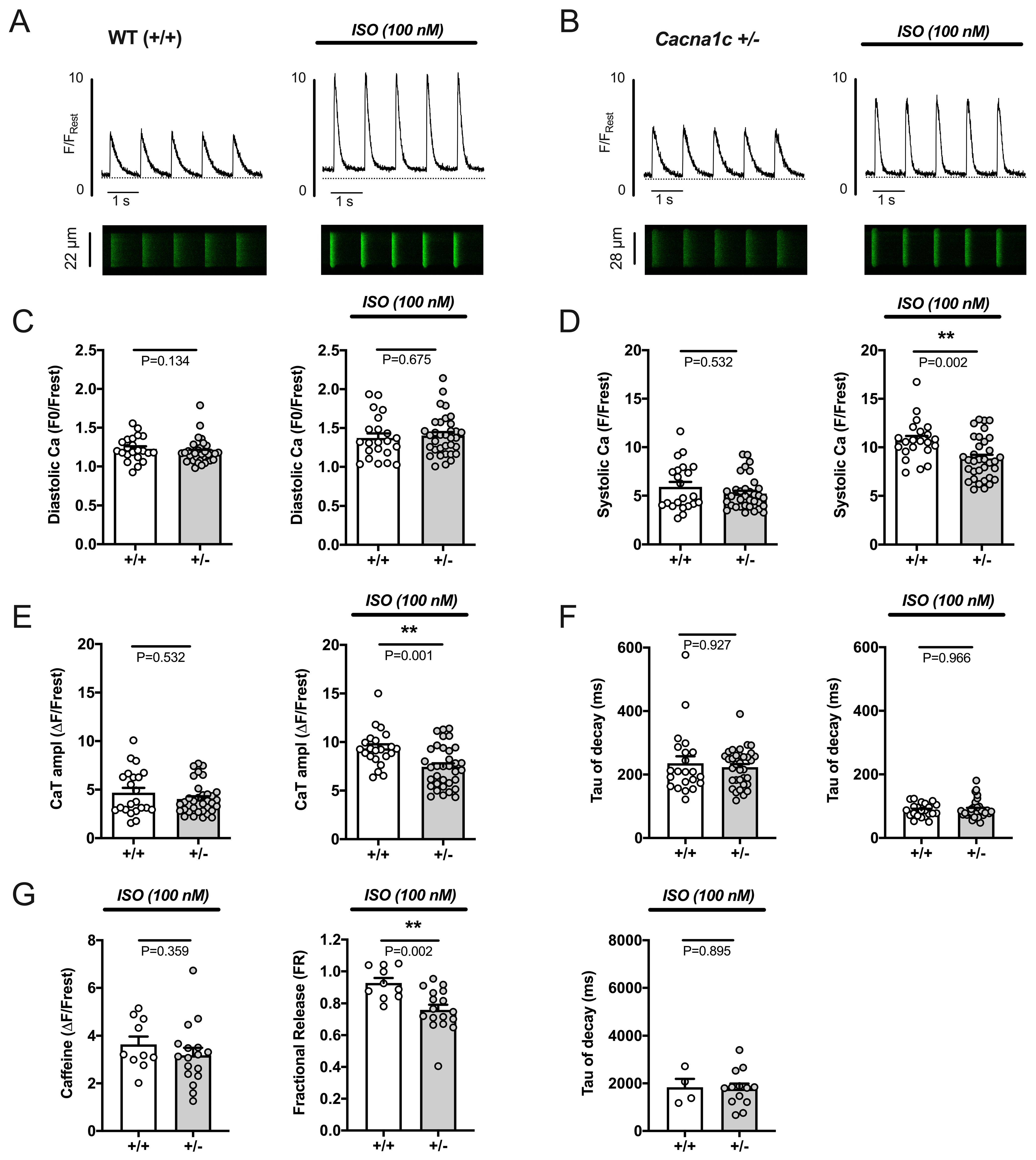
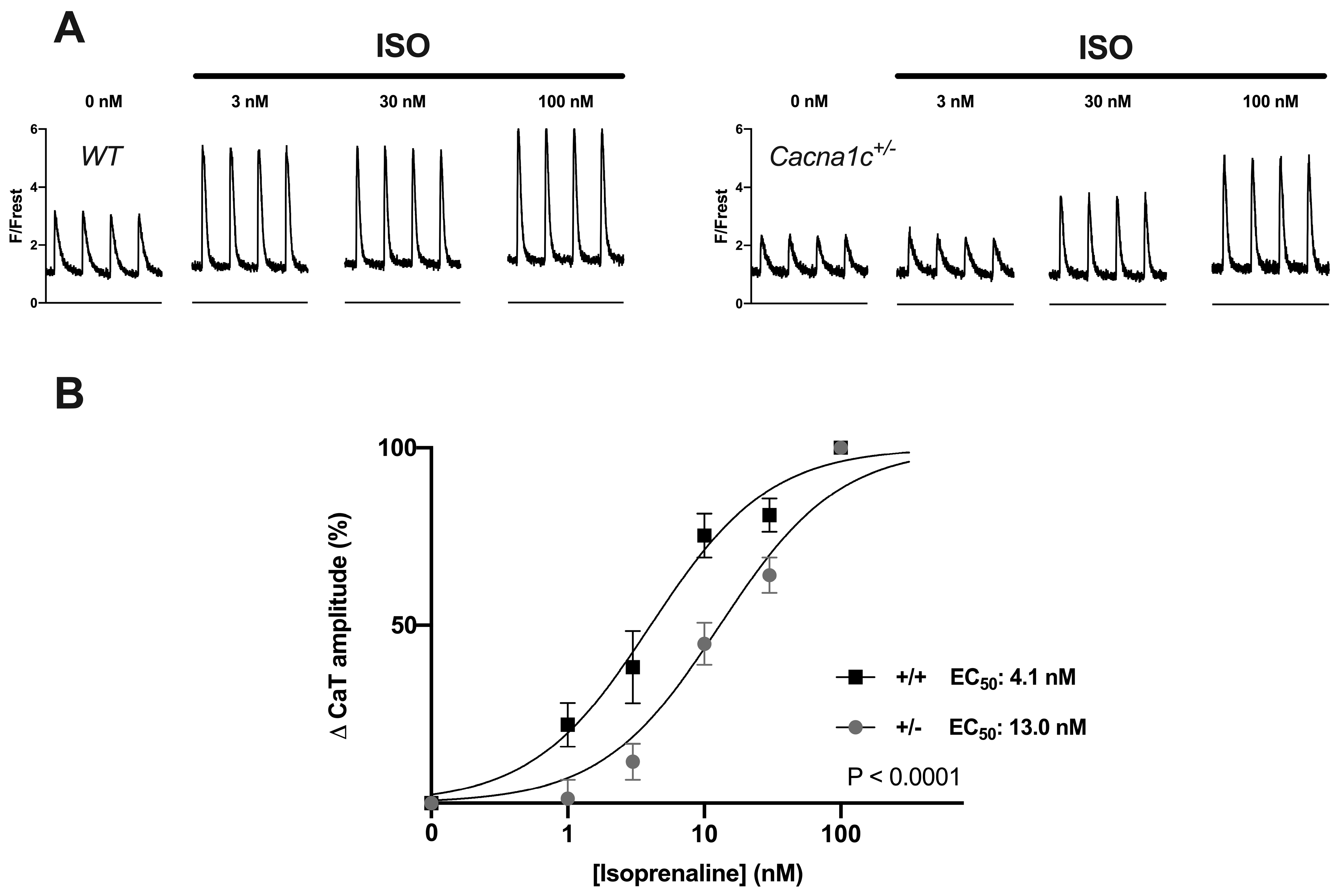
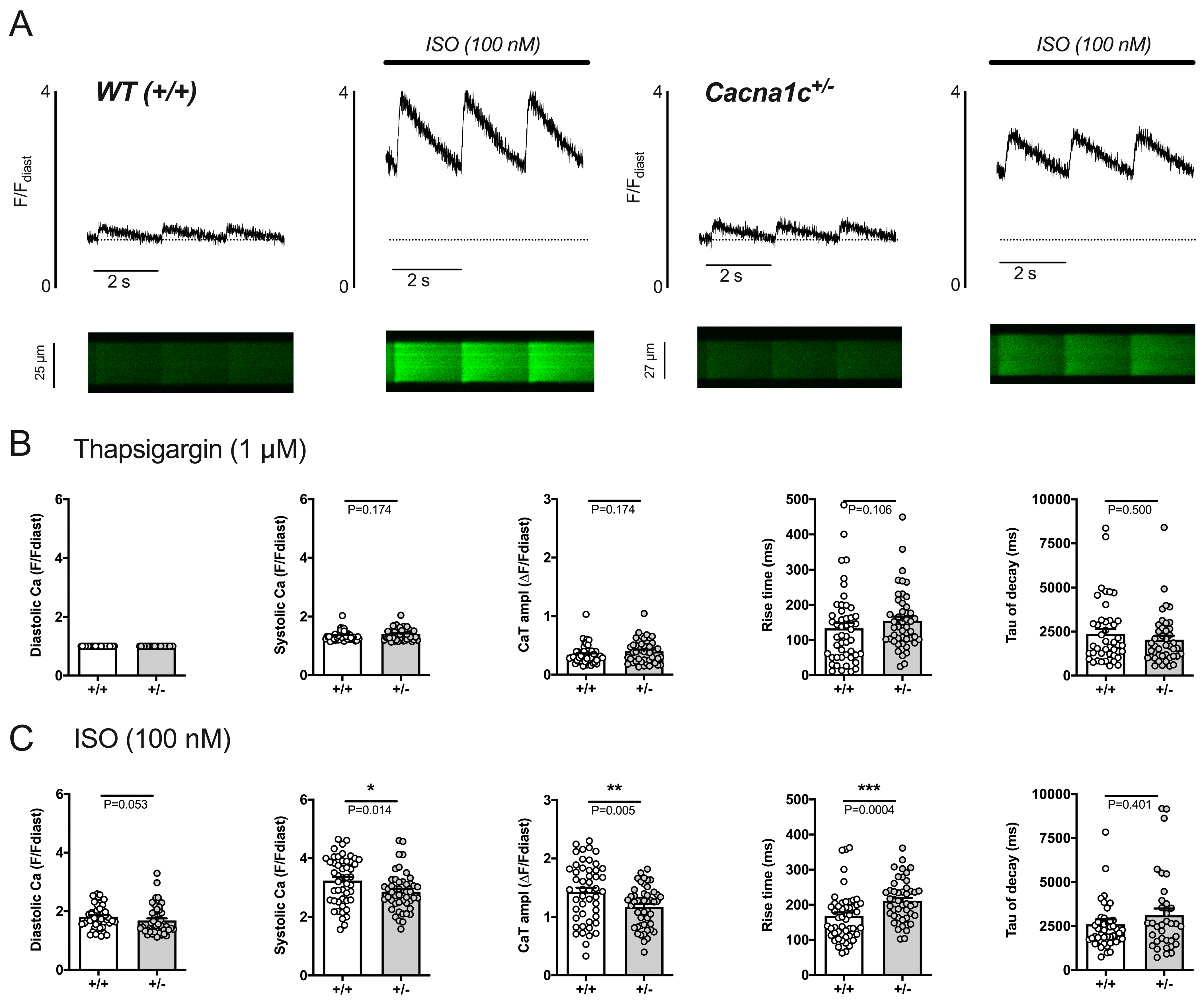
2.3.1. Attenuated Response of Ca2+ Transients and Sarcomere Shortenings to Stimulation by ISO in Cacna1c+/− Ventricular Myocytes
2.3.2. Reduced Potency of ISO to Stimulate Ca2+ Transients and Sarcomere Shortenings in Cacna1c+/− Ventricular Myocytes
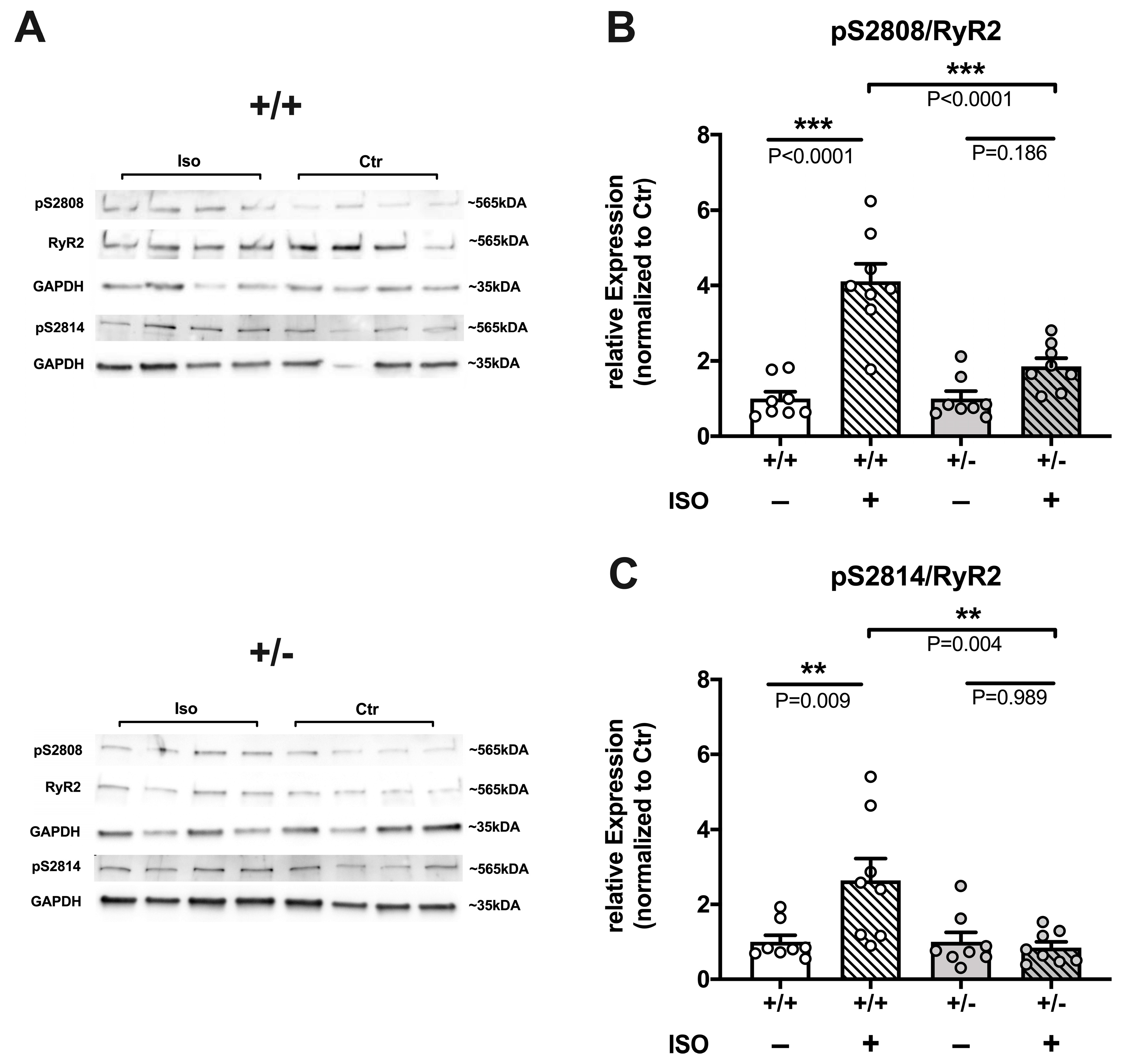
2.3.3. Reduced Efficacy of ISO to Increase RyR2 Phosphorylation in Cacna1c+/− LV Myocardium
2.3.4. Reduced Efficacy of ISO to Increase Sarcolemmal Ca2+ Influx in Cacna1c+/− Ventricular Myocytes
3. Discussion
3.1. Cardiomyocyte Function under Basal Conditions Appears Unaltered in Cacna1c+/− Rats
3.2. Remodeling of Ca2+ Handling Proteins under Basal Conditions in Cacna1c+/− LV Myocardium
3.3. Sympathetic Stress Unmasks a Defective Ca2+ Handling Phenotype in Cacna1c+/− Ventricular Myocytes
3.4. CACNA1C Gene as a Mechanistic Link between Psychiatric Disorders and Cardiac Disease?
3.5. Limitations
3.6. Conclusions
4. Materials and Methods
4.1. Animals
4.2. Left Ventricular Tissue Isolation
4.3. Isolation of Ventricular Myocytes and Treatment of Whole Hearts
4.4. Recording of Isolated Cardiomyocytes
4.5. Stressed Conditions
4.6. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Bhat, S.; Dao, D.T.; Terrillion, C.E.; Arad, M.; Smith, R.J.; Soldatov, N.M.; Gould, T.D. CACNA1C (Cav1.2) in the pathophysiology of psychiatric disease. Prog. Neurobiol. 2012, 99, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rougier, J.S.; Abriel, H. Cardiac voltage-gated calcium channel macromolecular complexes. Biochim. Biophys. Acta 2016, 1863, 1806–1812. [Google Scholar] [CrossRef] [PubMed]
- Kabir, Z.D.; Lee, A.S.; Rajadhyaksha, A.M. L-type Ca2+ channels in mood, cognition and addiction: Integrating human and rodent studies with a focus on behavioural endophenotypes. J. Physiol. 2016, 594, 5823–5837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bers, D.M. Cardiac excitation-contraction coupling. Nature 2002, 415, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Dedic, N.; Pohlmann, M.L.; Richter, J.S.; Mehta, D.; Czamara, D.; Metzger, M.W.; Dine, J.; Bedenk, B.T.; Hartmann, J.; Wagner, K.V.; et al. Cross-disorder risk gene CACNA1C differentially modulates susceptibility to psychiatric disorders during development and adulthood. Mol. Psychiatry 2018, 23, 533–543. [Google Scholar] [CrossRef] [PubMed]
- Splawski, I.; Timothy, K.W.; Sharpe, L.M.; Decher, N.; Kumar, P.; Bloise, R.; Napolitano, C.; Schwartz, P.J.; Joseph, R.M.; Condouris, K.; et al. Ca(V)1.2 calcium channel dysfunction causes a multisystem disorder including arrhythmia and autism. Cell 2004, 119, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wemhoner, K.; Friedrich, C.; Stallmeyer, B.; Coffey, A.J.; Grace, A.; Zumhagen, S.; Seebohm, G.; Ortiz-Bonnin, B.; Rinne, S.; Sachse, F.B.; et al. Gain-of-function mutations in the calcium channel CACNA1C (Cav1.2) cause non-syndromic long-QT but not Timothy syndrome. J. Mol. Cell. Cardiol. 2015, 80, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Nakayama, H.; Zhang, X.; Ai, X.; Harris, D.M.; Tang, M.; Zhang, H.; Szeto, C.; Stockbower, K.; Berretta, R.M.; et al. Calcium influx through Cav1.2 is a proximal signal for pathological cardiomyocyte hypertrophy. J. Mol. Cell. Cardiol. 2011, 50, 460–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goonasekera, S.A.; Hammer, K.; Auger-Messier, M.; Bodi, I.; Chen, X.; Zhang, H.; Reiken, S.; Elrod, J.W.; Correll, R.N.; York, A.J.; et al. Decreased cardiac L-type Ca(2)(+) channel activity induces hypertrophy and heart failure in mice. J. Clin. Investig. 2012, 122, 280–290. [Google Scholar] [CrossRef] [PubMed]
- Pluteanu, F.; Hess, J.; Plackic, J.; Nikonova, Y.; Preisenberger, J.; Bukowska, A.; Schotten, U.; Rinne, A.; Kienitz, M.C.; Schafer, M.K.; et al. Early subcellular Ca2+ remodelling and increased propensity for Ca2+ alternans in left atrial myocytes from hypertensive rats. Cardiovasc. Res. 2015, 106, 87–97. [Google Scholar] [CrossRef]
- Boixel, C.; Gonzalez, W.; Louedec, L.; Hatem, S.N. Mechanisms of L-type Ca2+ current downregulation in rat atrial myocytes during heart failure. Circ. Res. 2001, 89, 607–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schotten, U.; Verheule, S.; Kirchhof, P.; Goette, A. Pathophysiological mechanisms of atrial fibrillation: A translational appraisal. Physiol. Rev. 2011, 91, 265–325. [Google Scholar] [CrossRef] [PubMed]
- Hayes, J.F.; Lundin, A.; Wicks, S.; Lewis, G.; Wong, I.C.K.; Osborn, D.P.J.; Dalman, C. Association of Hydroxylmethyl Glutaryl Coenzyme A Reductase Inhibitors, L-Type Calcium Channel Antagonists, and Biguanides With Rates of Psychiatric Hospitalization and Self-Harm in Individuals With Serious Mental Illness. JAMA Psychiatry 2019, 76, 382–390. [Google Scholar] [CrossRef] [Green Version]
- Kessing, L.V.; Rytgaard, H.C.; Ekstrom, C.T.; Torp-Pedersen, C.; Berk, M.; Gerds, T.A. Antihypertensive Drugs and Risk of Depression: A Nationwide Population-Based Study. Hypertension 2020, 76, 1263–1279. [Google Scholar] [CrossRef] [PubMed]
- Kisko, T.M.; Braun, M.D.; Michels, S.; Witt, S.H.; Rietschel, M.; Culmsee, C.; Schwarting, R.K.W.; Wohr, M. Cacna1c haploinsufficiency leads to pro-social 50-kHz ultrasonic communication deficits in rats. Dis. Model. Mech. 2018, 11, dmm034116. [Google Scholar] [CrossRef] [Green Version]
- Redecker, T.M.; Kisko, T.M.; Schwarting, R.K.W.; Wohr, M. Effects of Cacna1c haploinsufficiency on social interaction behavior and 50-kHz ultrasonic vocalizations in adult female rats. Behav. Brain Res. 2019, 367, 35–52. [Google Scholar] [CrossRef]
- Wohr, M.; Willadsen, M.; Kisko, T.M.; Schwarting, R.K.W.; Fendt, M. Sex-dependent effects of Cacna1c haploinsufficiency on behavioral inhibition evoked by conspecific alarm signals in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 99, 109849. [Google Scholar] [CrossRef]
- Lee, A.S.; Ra, S.; Rajadhyaksha, A.M.; Britt, J.K.; De Jesus-Cortes, H.; Gonzales, K.L.; Lee, A.; Moosmang, S.; Hofmann, F.; Pieper, A.A.; et al. Forebrain elimination of cacna1c mediates anxiety-like behavior in mice. Mol. Psychiatry 2012, 17, 1054–1055. [Google Scholar] [CrossRef] [Green Version]
- Bassani, J.W.; Bassani, R.A.; Bers, D.M. Relaxation in rabbit and rat cardiac cells: Species-dependent differences in cellular mechanisms. J. Physiol. 1994, 476, 279–293. [Google Scholar] [CrossRef]
- Rosati, B.; Yan, Q.; Lee, M.S.; Liou, S.R.; Ingalls, B.; Foell, J.; Kamp, T.J.; McKinnon, D. Robust L-type calcium current expression following heterozygous knockout of the Cav1.2 gene in adult mouse heart. J. Physiol. 2011, 589, 3275–3288. [Google Scholar] [CrossRef]
- Papa, A.; Kushner, J.; Marx, S.O. Adrenergic Regulation of Calcium Channels in the Heart. Annu. Rev. Physiol. 2022, 84, 285–306. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E. Totally Rad? The Long and Winding Road to Understanding CaV1.2 Regulation. Circ. Res. 2021, 128, 89–91. [Google Scholar] [CrossRef] [PubMed]
- Park, N.; Marquez, J.; Pham, T.K.; Ko, T.H.; Youm, J.B.; Kim, M.; Choi, S.H.; Moon, J.; Flores, J.; Ko, K.S.; et al. Cereblon contributes to cardiac dysfunction by degrading Cav1.2alpha. Eur. Heart J. 2022, 43, 1973–1989. [Google Scholar] [CrossRef] [PubMed]
- Marx, S.O.; Reiken, S.; Hisamatsu, Y.; Jayaraman, T.; Burkhoff, D.; Rosemblit, N.; Marks, A.R. PKA phosphorylation dissociates FKBP12.6 from the calcium release channel (ryanodine receptor): Defective regulation in failing hearts. Cell 2000, 101, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Fill, M.; Copello, J.A. Ryanodine receptor calcium release channels. Physiol. Rev. 2002, 82, 893–922. [Google Scholar] [CrossRef] [Green Version]
- Kranias, E.G.; Hajjar, R.J. Modulation of cardiac contractility by the phospholamban/SERCA2a regulatome. Circ. Res. 2012, 110, 1646–1660. [Google Scholar] [CrossRef] [Green Version]
- Bers, D.M. Macromolecular complexes regulating cardiac ryanodine receptor function. J. Mol. Cell. Cardiol. 2004, 37, 417–429. [Google Scholar] [CrossRef]
- Lehnart, S.E.; Wehrens, X.H.T.; Reiken, S.; Warrier, S.; Belevych, A.E.; Harvey, R.D.; Richter, W.; Jin, S.L.C.; Conti, M.; Marks, A.R. Phosphodiesterase 4D Deficiency in the Ryanodine-Receptor Complex Promotes Heart Failure and Arrhythmias. Cell 2005, 123, 25–35. [Google Scholar] [CrossRef] [Green Version]
- Headrick, J.P.; Peart, J.N.; Budiono, B.P.; Shum, D.H.K.; Neumann, D.L.; Stapelberg, N.J.C. The heartbreak of depression: ‘Psycho-cardiac’ coupling in myocardial infarction. J. Mol. Cell. Cardiol. 2017, 106, 14–28. [Google Scholar] [CrossRef] [Green Version]
- Sbolli, M.; Fiuzat, M.; Cani, D.; O’Connor, C.M. Depression and heart failure: The lonely comorbidity. Eur. J. Heart Fail. 2020, 22, 2007–2017. [Google Scholar] [CrossRef]
- Pina, I.L.; Di Palo, K.E.; Ventura, H.O. Psychopharmacology and Cardiovascular Disease. J. Am. Coll. Cardiol. 2018, 71, 2346–2359. [Google Scholar] [CrossRef] [PubMed]
- Vaccarino, V.; Badimon, L.; Bremner, J.D.; Cenko, E.; Cubedo, J.; Dorobantu, M.; Duncker, D.J.; Koller, A.; Manfrini, O.; Milicic, D.; et al. Depression and coronary heart disease: 2018 position paper of the ESC working group on coronary pathophysiology and microcirculation. Eur. Heart J. 2020, 41, 1687–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- von Eisenhart Rothe, A.; Hutt, F.; Baumert, J.; Breithardt, G.; Goette, A.; Kirchhof, P.; Ladwig, K.H. Depressed mood amplifies heart-related symptoms in persistent and paroxysmal atrial fibrillation patients: A longitudinal analysis—Data from the German Competence Network on Atrial Fibrillation. Europace 2015, 17, 1354–1362. [Google Scholar] [CrossRef]
- Studer, R.; Reinecke, H.; Bilger, J.; Eschenhagen, T.; Bohm, M.; Hasenfuss, G.; Just, H.; Holtz, J.; Drexler, H. Gene expression of the cardiac Na(+)-Ca2+ exchanger in end-stage human heart failure. Circ. Res. 1994, 75, 443–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pogwizd, S.M.; Schlotthauer, K.; Li, L.; Yuan, W.; Bers, D.M. Arrhythmogenesis and Contractile Dysfunction in Heart Failure. Circ. Res. 2001, 88, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Reiken, S.; Gaburjakova, M.; Guatimosim, S.; Gomez, A.M.; D’Armiento, J.; Burkhoff, D.; Wang, J.; Vassort, G.; Lederer, W.J.; Marks, A.R. Protein Kinase A Phosphorylation of the Cardiac Calcium Release Channel (Ryanodine Receptor) in Normal and Failing Hearts. J. Biol. Chem. 2003, 278, 444–453. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.S.; Velez Rueda, J.O.; Salas, M.; Becerra, R.; Di Carlo, M.N.; Said, M.; Vittone, L.; Rinaldi, G.; Portiansky, E.L.; Mundina-Weilenmann, C.; et al. Increased Na+/Ca2+ exchanger expression/activity constitutes a point of inflection in the progression to heart failure of hypertensive rats. PLoS ONE 2014, 9, e96400. [Google Scholar] [CrossRef]
- Chen-Izu, Y.; Ward, C.W.; Stark, W.; Banyasz, T.; Sumandea, M.P.; Balke, C.W.; Izu, L.T.; Wehrens, X.H.T. Phosphorylation of RyR2and shortening of RyR2cluster spacing in spontaneously hypertensive rat with heart failure. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2409–H2417. [Google Scholar] [CrossRef] [Green Version]
- Wehrens, X.H.T.; Lehnart, S.E.; Huang, F.; Vest, J.A.; Reiken, S.R.; Mohler, P.J.; Sun, J.; Guatimosim, S.; Song, L.-S.; Rosemblit, N.; et al. FKBP12.6 Deficiency and Defective Calcium Release Channel (Ryanodine Receptor) Function Linked to Exercise-Induced Sudden Cardiac Death. Cell 2003, 113, 829–840. [Google Scholar] [CrossRef] [Green Version]
- Boknik, P.; Heinroth-Hoffmann, I.; Kirchhefer, U.; Knapp, J.; Linck, B.; Luss, H.; Muller, T.; Schmitz, W.; Brodde, O.; Neumann, J. Enhanced protein phosphorylation in hypertensive hypertrophy. Cardiovasc. Res. 2001, 51, 717–728. [Google Scholar] [CrossRef] [Green Version]
- Power, A.S.; Hickey, A.J.; Crossman, D.J.; Loiselle, D.S.; Ward, M.L. Calcium mishandling impairs contraction in right ventricular hypertrophy prior to overt heart failure. Pflug. Arch. 2018, 470, 1115–1126. [Google Scholar] [CrossRef] [PubMed]
- Bristow, M.R.; Ginsburg, R.; Minobe, W.; Cubicciotti, R.S.; Sageman, W.S.; Lurie, K.; Billingham, M.E.; Harrison, D.C.; Stinson, E.B. Decreased catecholamine sensitivity and beta-adrenergic-receptor density in failing human hearts. N. Engl. J. Med. 1982, 307, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Schillinger, W.; Pieske, B.; Holubarsch, C.; Heilmann, C.; Posival, H.; Kuwajima, G.; Mikoshiba, K.; Just, H.; Hasenfuss, G.; et al. Alterations of sarcoplasmic reticulum proteins in failing human dilated cardiomyopathy. Circulation 1995, 92, 778–784. [Google Scholar] [CrossRef]
- Ai, X.; Curran, J.W.; Shannon, T.R.; Bers, D.M.; Pogwizd, S.M. Ca2+/calmodulin-dependent protein kinase modulates cardiac ryanodine receptor phosphorylation and sarcoplasmic reticulum Ca2+ leak in heart failure. Circ. Res. 2005, 97, 1314–1322. [Google Scholar] [CrossRef] [Green Version]
- Fischer, T.H.; Herting, J.; Tirilomis, T.; Renner, A.; Neef, S.; Toischer, K.; Ellenberger, D.; Forster, A.; Schmitto, J.D.; Gummert, J.; et al. Ca2+/calmodulin-dependent protein kinase II and protein kinase A differentially regulate sarcoplasmic reticulum Ca2+ leak in human cardiac pathology. Circulation 2013, 128, 970–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Zhang, T.; Mestril, R.; Bers, D.M. Ca/Calmodulin-Dependent Protein Kinase II Phosphorylation of Ryanodine Receptor Does Affect Calcium Sparks in Mouse Ventricular Myocytes. Circ. Res. 2006, 99, 398–406. [Google Scholar] [CrossRef] [Green Version]
- Kockskamper, J.; Pieske, B. Phosphorylation of the cardiac ryanodine receptor by Ca2+/calmodulin-dependent protein kinase II: The dominating twin of protein kinase A? Circ. Res. 2006, 99, 333–335. [Google Scholar] [CrossRef] [Green Version]
- Sossalla, S.; Fluschnik, N.; Schotola, H.; Ort, K.R.; Neef, S.; Schulte, T.; Wittköpper, K.; Renner, A.; Schmitto, J.D.; Gummert, J.; et al. Inhibition of Elevated Ca2+/Calmodulin-Dependent Protein Kinase II Improves Contractility in Human Failing Myocardium. Circ. Res. 2010, 107, 1150–1161. [Google Scholar] [CrossRef] [Green Version]
- Fischer, T.H.; Neef, S.; Maier, L.S. The Ca-calmodulin dependent kinase II: A promising target for future antiarrhythmic therapies? J. Mol. Cell. Cardiol. 2013, 58, 182–187. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Larsson, H.P. Insights into Cardiac IKs (KCNQ1/KCNE1) Channels Regulation. Int. J. Mol. Sci. 2020, 21, 9440. [Google Scholar] [CrossRef]
- Dai, L.; Zang, Y.; Zheng, D.; Xia, L.; Gong, Y. Role of CaMKII and PKA in Early Afterdepolarization of Human Ventricular Myocardium Cell: A Computational Model Study. Comput. Math Methods Med. 2016, 2016, 4576313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zang, Y.; Dai, L.; Zhan, H.; Dou, J.; Xia, L.; Zhang, H. Theoretical investigation of the mechanism of heart failure using a canine ventricular cell model: Especially the role of up-regulated CaMKII and SR Ca2+ leak. J. Mol. Cell. Cardiol. 2013, 56, 34–43. [Google Scholar] [CrossRef] [PubMed]
- Geurts, A.M.; Cost, G.J.; Freyvert, Y.; Zeitler, B.; Miller, J.C.; Choi, V.M.; Jenkins, S.S.; Wood, A.; Cui, X.; Meng, X.; et al. Knockout rats via embryo microinjection of zinc-finger nucleases. Science 2009, 325, 433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seisenberger, C.; Specht, V.; Welling, A.; Platzer, J.; Pfeifer, A.; Kuhbandner, S.; Striessnig, J.; Klugbauer, N.; Feil, R.; Hofmann, F. Functional embryonic cardiomyocytes after disruption of the L-type alpha1C (Cav1.2) calcium channel gene in the mouse. J. Biol. Chem. 2000, 275, 39193–39199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plackic, J.; Preissl, S.; Nikonova, Y.; Pluteanu, F.; Hein, L.; Kockskamper, J. Enhanced nucleoplasmic Ca2+ signaling in ventricular myocytes from young hypertensive rats. J. Mol. Cell. Cardiol. 2016, 101, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Plackic, J.; Kockskamper, J. Isolation of Atrial and Ventricular Cardiomyocytes for In Vitro Studies. Methods Mol. Biol. 2018, 1816, 39–54. [Google Scholar] [CrossRef] [PubMed]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fender, H.; Walter, K.; Kiper, A.K.; Plačkić, J.; Kisko, T.M.; Braun, M.D.; Schwarting, R.K.W.; Rohrbach, S.; Wöhr, M.; Decher, N.; et al. Calcium Handling Remodeling Underlies Impaired Sympathetic Stress Response in Ventricular Myocardium from Cacna1c Haploinsufficient Rats. Int. J. Mol. Sci. 2023, 24, 9795. https://doi.org/10.3390/ijms24129795
Fender H, Walter K, Kiper AK, Plačkić J, Kisko TM, Braun MD, Schwarting RKW, Rohrbach S, Wöhr M, Decher N, et al. Calcium Handling Remodeling Underlies Impaired Sympathetic Stress Response in Ventricular Myocardium from Cacna1c Haploinsufficient Rats. International Journal of Molecular Sciences. 2023; 24(12):9795. https://doi.org/10.3390/ijms24129795
Chicago/Turabian StyleFender, Hauke, Kim Walter, Aytug K. Kiper, Jelena Plačkić, Theresa M. Kisko, Moria D. Braun, Rainer K. W. Schwarting, Susanne Rohrbach, Markus Wöhr, Niels Decher, and et al. 2023. "Calcium Handling Remodeling Underlies Impaired Sympathetic Stress Response in Ventricular Myocardium from Cacna1c Haploinsufficient Rats" International Journal of Molecular Sciences 24, no. 12: 9795. https://doi.org/10.3390/ijms24129795