
Calpain-3 Is Not a Sodium Dependent Protease and Simply Requires Calcium for Activation

Abstract
:1. Introduction
2. Results
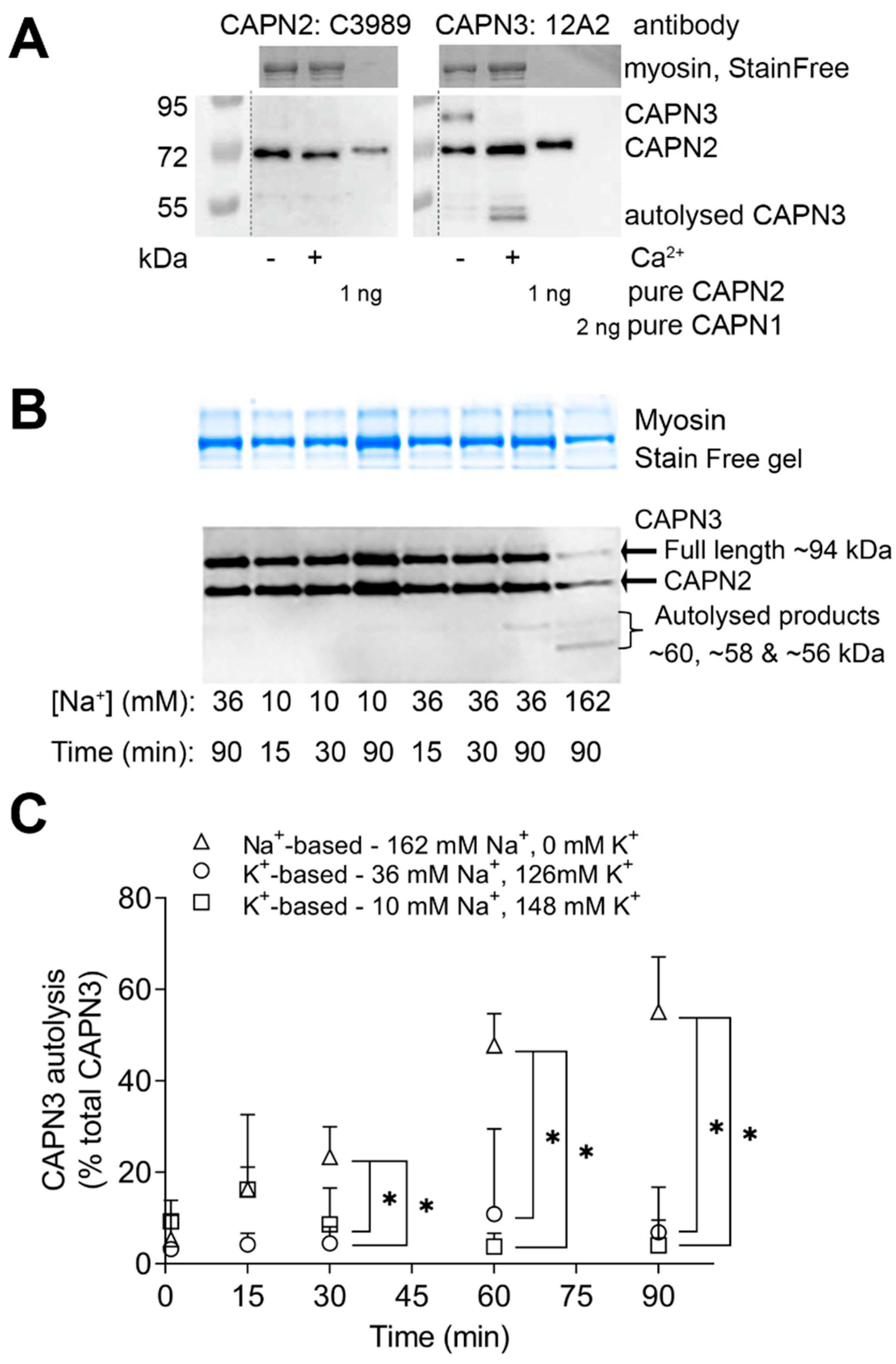
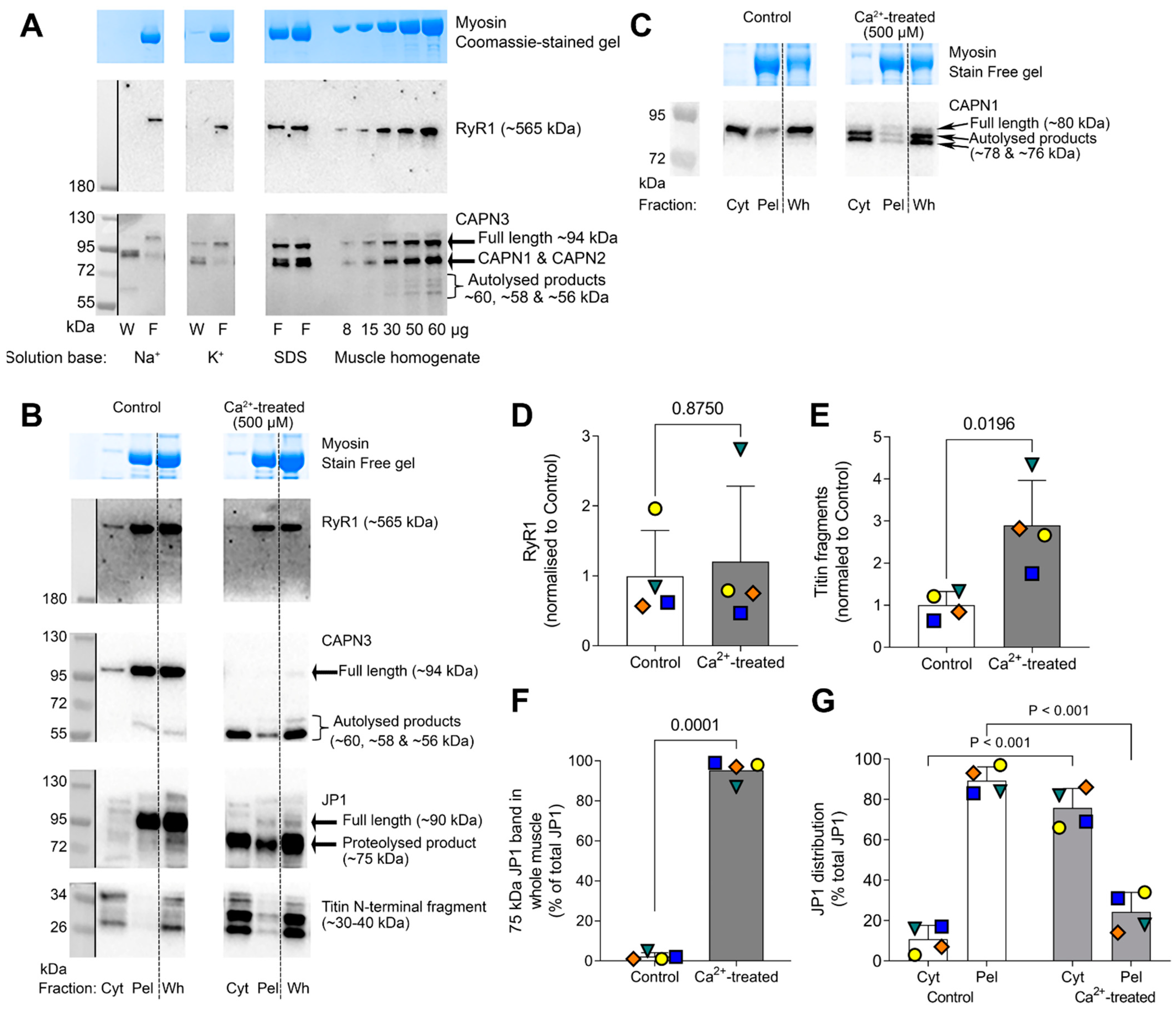
2.1. Effect of Na+ on CAPN3 Autolysis
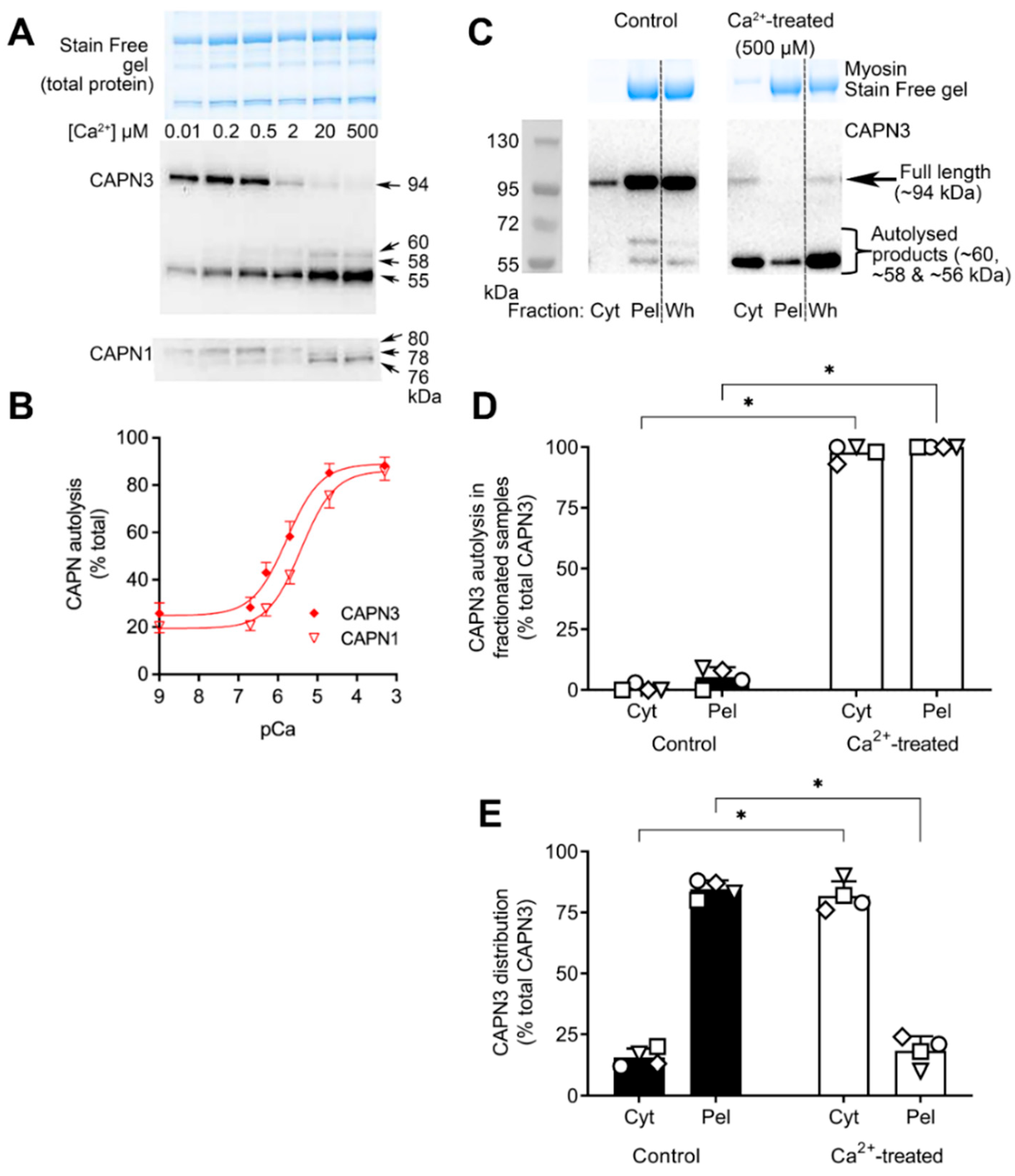
2.2. Ca2+-Dependence of CAPN3 Autolysis, and Redistribution of Autolysed CAPN3 to Cytosol
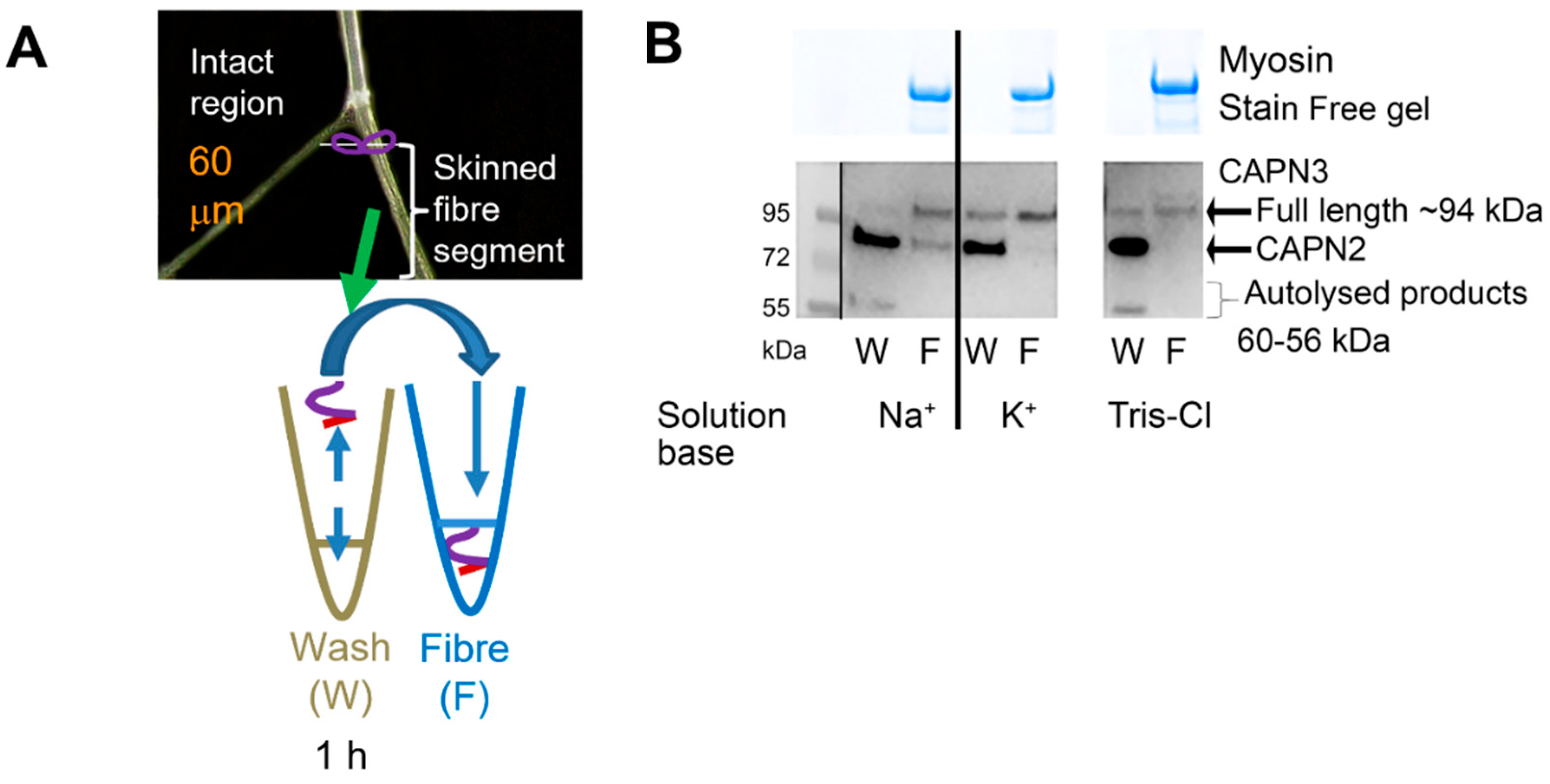
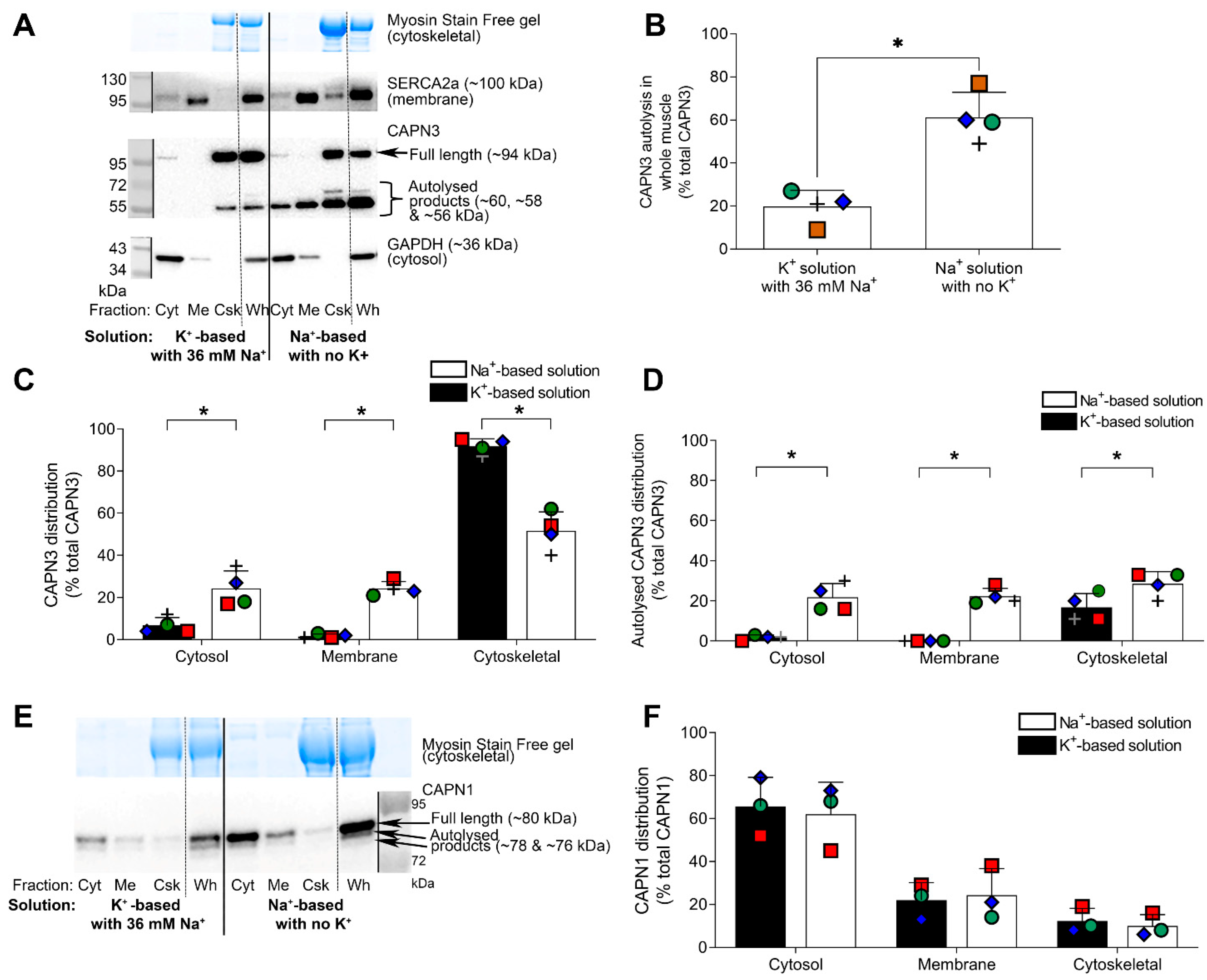
2.3. Further Examination of Effect of Na+ Treatment on CAPN3 Localisation
2.4. RyR1 Protein Is Not Proteolyzed upon CAPN3 Autolysis with Treatment with High [Na+] or 500 µM Ca2+
3. Discussion
3.1. CAPN3 Is Not a Na+-Dependent Protease in Physiological Circumstances
3.2. Activation of CAPN3 in Physiological Circumstances
3.3. Diffusibility of Autolysed CAPN3
3.4. No Evidence of RyR1 Proteolysis upon CAPN3 Activation
4. Materials and Methods
4.1. Chemicals
4.2. Solutions
4.3. Collection of Samples
4.4. Rat Samples and Single Fibre Diffusion Experiments
4.5. Human Samples, Calcium-Treated Muscle Homogenates and Crude Fractionation Experiments
4.6. Western Blotting
4.7. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Diaz, B.G.; Moldoveanu, T.; Kuiper, M.J.; Campbell, R.L.; Davies, P.L. Insertion sequence 1 of muscle-specific calpain, p94, acts as an internal propeptide. J. Biol. Chem. 2004, 279, 27656–27666. [Google Scholar] [CrossRef] [PubMed]
- Campbell, R.L.; Davies, P.L. Structure–function relationships in calpains1. Biochem. J. 2012, 447, 335–351. [Google Scholar] [CrossRef]
- Rey, M.A.; Davies, P.L. The protease core of the muscle-specific calpain, p94, undergoes Ca2+-dependent intramolecular autolysis. FEBS Lett. 2002, 532, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Shindo, M.; Doi, N.; Kitamura, F.; Gregorio, C.C.; Sorimachi, H. The N- and C-terminal autolytic fragments of CAPN3/p94/calpain-3 restore proteolytic activity by intermolecular complementation. Proc. Natl. Acad. Sci. USA 2014, 111, E5527–E5536. [Google Scholar] [CrossRef] [PubMed]
- Branca, D.; Gugliucci, A.; Bano, D.; Brini, M.; Carafoli, E. Expression, partial purification and functional properties of themuscle-specific calpain isoform p94. Eur. J. Biochem. 1999, 265, 839–846. [Google Scholar] [CrossRef]
- Murphy, R.M.; Lamb, G.D. Endogenous calpain-3 activation is primarily governed by small increases in resting cytoplasmic [Ca2+] and is not dependent on stretch. J. Biol. Chem. 2009, 284, 7811–7819. [Google Scholar] [CrossRef] [PubMed]
- Keira, Y.; Noguchi, S.; Minami, N.; Hayashi, Y.K.; Nishino, I. Localization of calpain 3 in human skeletal muscle and its alteration in limb-girdle muscular dystrophy 2A muscle. J. Biochem. 2003, 133, 659–664. [Google Scholar] [CrossRef]
- Sorimachi, H.; Kinbara, K.; Kimura, S.; Takahashi, M.; Ishiura, S.; Sasagawa, N.; Sorimachi, N.; Shimada, H.; Tagawa, K.; Maruyama, K.; et al. Muscle-specific calpain, p94, responsible for limb girdle muscular dystrophy type 2A, associates with connectin through IS2, a p94-specific sequence. J. Biol. Chem. 1995, 270, 31158–31162. [Google Scholar] [CrossRef]
- Ono, Y.; Ojima, K.; Torii, F.; Takaya, E.; Doi, N.; Nakagawa, K.; Hata, S.; Abe, K.; Sorimachi, H. Skeletal muscle-specific calpain is an intracellular Na+-dependent protease. J. Biol. Chem. 2010, 285, 22986–22998. [Google Scholar] [CrossRef]
- Fitts, R.H. Cellular mechanisms of muscle fatigue. Physiol. Rev. 1994, 74, 49–94. [Google Scholar] [CrossRef]
- Cairns, S.P.; Lindinger, M.I. Do multiple ionic interactions contribute to skeletal muscle fatigue? J. Physiol. 2008, 586, 4039–4054. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, S.; Feng, W.; Varsanyi, M.; Shoshan-Barmatz, V. Identification, characterization and partial purification of a thiol-protease which cleaves specifically the skeletal muscle ryanodine receptor/Ca2+ release channel. J. Membr. Biol. 1998, 161, 33–43. [Google Scholar] [CrossRef]
- Murphy, R.M.; Snow, R.J.; Lamb, G.D. mu-Calpain and calpain-3 are not autolyzed with exhaustive exercise in humans. Am. J. Physiol. Cell Physiol. 2006, 290, C116–C122. [Google Scholar] [CrossRef]
- Wette, S.G.; Lamb, G.D.; Murphy, R.M. Nuclei isolation methods fail to accurately assess the subcellular localization and behaviour of proteins in skeletal muscle. Acta Physiol. 2021, 233, e13730. [Google Scholar] [CrossRef] [PubMed]
- Baghdiguian, S.; Richard, I.; Martin, M.; Coopman, P.; Beckmann, J.S.; Mangeat, P.; Lefranc, G. Pathophysiology of limb girdle muscular dystrophy type 2A: Hypothesis and new insights into the IkappaBalpha/NF-kappaB survival pathway in skeletal muscle. J. Mol. Med. 2001, 79, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Ono, Y.; Ojima, K.; Shinkai-Ouchi, F.; Hata, S.; Sorimachi, H. An eccentric calpain, CAPN3/p94/calpain-3. Biochimie 2016, 122, 169–187. [Google Scholar] [CrossRef]
- Murphy, R.M.; Dutka, T.L.; Horvath, D.; Bell, J.R.; Delbridge, L.M.; Lamb, G.D. Ca2+-dependent proteolysis of junctophilin-1 and junctophilin-2 in skeletal and cardiac muscle. J. Physiol. 2013, 591, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Meissner, G.; Rios, E.; Tripathy, A.; Pasek, D.A. Regulation of skeletal muscle Ca2+ release channel (ryanodine receptor) by Ca2+ and monovalent cations and anions. J. Biol. Chem. 1997, 272, 1628–1638. [Google Scholar] [CrossRef]
- Baylor, S.M.; Hollingworth, S. Sarcoplasmic reticulum calcium release compared in slow-twitch and fast-twitch fibres of mouse muscle. J. Physiol. 2003, 551, 125–138. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Skeletal muscle fatigue: Cellular mechanisms. Physiol. Rev. 2008, 88, 287–332. [Google Scholar] [CrossRef]
- Murphy, R.M.; Goodman, C.A.; McKenna, M.J.; Bennie, J.; Leikis, M.; Lamb, G.D. Calpain-3 is autolyzed and hence activated in human skeletal muscle 24 h following a single bout of eccentric exercise. J. Appl. Physiol. 2007, 103, 926–931. [Google Scholar] [CrossRef]
- Murphy, R.M.; Vissing, K.; Latchman, H.; Lamboley, C.; McKenna, M.J.; Overgaard, K.; Lamb, G.D. Activation of skeletal muscle calpain-3 by eccentric exercise in humans does not result in its translocation to the nucleus or cytosol. J. Appl. Physiol. 2011, 11, 1448–1458. [Google Scholar] [CrossRef] [PubMed]
- Ojima, K.; Kawabata, Y.; Nakao, H.; Nakao, K.; Doi, N.; Kitamura, F.; Ono, Y.; Hata, S.; Suzuki, H.; Kawahara, H.; et al. Dynamic distribution of muscle-specific calpain in mice has a key role in physical-stress adaptation and is impaired in muscular dystrophy. J. Clin. Investig. 2010, 120, 2672–2683. [Google Scholar] [CrossRef]
- Kanzaki, K.; Kuratani, M.; Matsunaga, S.; Yanaka, N.; Wada, M. Three calpain isoforms are autolyzed in rat fast-twitch muscle after eccentric contractions. J. Muscle Res. Cell Motil. 2014, 35, 179–189. [Google Scholar] [CrossRef]
- Lynch, G.S.; Fary, C.J.; Williams, D.A. Quantitative measurement of resting skeletal muscle [Ca2+]i following acute and long-term downhill running exercise in mice. Cell Calcium 1997, 22, 373–383. [Google Scholar] [CrossRef] [PubMed]
- Kramerova, I.; Kudryashova, E.; Wu, B.; Ottenheijm, C.; Granzier, H.; Spencer, M.J. Novel role of calpain-3 in the triad-associated protein complex regulating calcium release in skeletal muscle. Hum. Mol. Genet. 2008, 17, 3271–3280. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, J.S.; Belcastro, A.N.; Katz, S. Intraluminal Ca2+ dependence of Ca2+ and ryanodine-mediated regulation of skeletal muscle sarcoplasmic reticulum Ca2+ release. J. Biol. Chem. 1992, 267, 20850–20856. [Google Scholar] [CrossRef]
- Kinbara, K.; Ishiura, S.; Tomioka, S.; Sorimachi, H.; Jeong, S.Y.; Amano, S.; Kawasaki, H.; Kolmerer, B.; Kimura, S.; Labeit, S.; et al. Purification of native p94, a muscle-specific calpain, and characterization of its autolysis. Biochem. J. 1998, 335 Pt 3, 589–596. [Google Scholar] [CrossRef]
- Ye, Q.; Campbell, R.L.; Davies, P.L. Structures of human calpain-3 protease core with and without bound inhibitor reveal mechanisms of calpain activation. J. Biol. Chem. 2018, 293, 4056–4070. [Google Scholar] [CrossRef]
- Shoshan-Barmatz, V.; Weil, S.; Meyer, H.; Varsanyi, M.; Heilmeyer, L.M. Endogenous, Ca2+-dependent cysteine-protease cleaves specifically the ryanodine receptor/Ca2+ release channel in skeletal muscle. J. Membr. Biol. 1994, 142, 281–288. [Google Scholar] [CrossRef]
- Tammineni, E.R.; Figueroa, L.; Manno, C.; Varma, D.; Kraeva, N.; Ibarra, C.A.; Klip, A.; Riazi, S.; Rios, E. Muscle calcium stress cleaves junctophilin1, unleashing a gene regulatory program predicted to correct glucose dysregulation. eLife 2023, 12, e78874. [Google Scholar] [CrossRef] [PubMed]
- Verburg, E.; Murphy, R.M.; Richard, I.; Lamb, G.D. Involvement of calpains in Ca2+-induced disruption of excitation-contraction coupling in mammalian skeletal muscle fibers. Am. J. Physiol. Cell Physiol. 2009, 296, C1115–C1122. [Google Scholar] [CrossRef] [PubMed]
- Lamb, G.D.; Stephenson, D.G. Measurement of force and calcium release using mechanically skinned fibers from mammalian skeletal muscle. J. Appl. Physiol. 2018, 125, 1105–1127. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, J. Muscle electrolytes in man. Scand. J. Clin. Lab. Investig. 1962, 14, 1–110. [Google Scholar]
- Evans, W.J.; Phinney, S.D.; Young, V.R. Suction applied to a muscle biopsy maximizes sample size. Med. Sci. Sport. Exerc. 1982, 14, 101–102. [Google Scholar]
- Murphy, R.M.; Mollica, J.P.; Lamb, G.D. Plasma membrane removal in rat skeletal muscle fibers reveals caveolin-3 hot-spots at the necks of transverse tubules. Exp. Cell Res. 2009, 315, 1015–1028. [Google Scholar] [CrossRef]
- Mollica, J.P.; Oakhill, J.S.; Lamb, G.D.; Murphy, R.M. Are genuine changes in protein expression being overlooked? Reassessing Western blotting. Anal. Biochem. 2009, 386, 270–275. [Google Scholar] [CrossRef]
- Murphy, R.M.; Lamb, G.D. Important considerations for protein analyses using antibody based techniques: Down-sizing Western blotting up-sizes outcomes. J. Physiol. 2013, 591, 5823–5831. [Google Scholar] [CrossRef]
- Kanda, K.; Sakuma, J.; Akimoto, T.; Kawakami, Y.; Suzuki, K. Detection of titin fragments in urine in response to exercise-induced muscle damage. PLoS ONE 2017, 12, e0181623. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ionic Condition: | Ca2+ with or without K+ | Na+ without K+ Present | Na+ with K+ Present |
|---|---|---|---|
| CAPN1 autolysis | ✓ | X | X |
| CAPN3 autolysis | ✓ | ✓ | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wette, S.G.; Lamb, G.D.; Murphy, R.M. Calpain-3 Is Not a Sodium Dependent Protease and Simply Requires Calcium for Activation. Int. J. Mol. Sci. 2023, 24, 9405. https://doi.org/10.3390/ijms24119405
Wette SG, Lamb GD, Murphy RM. Calpain-3 Is Not a Sodium Dependent Protease and Simply Requires Calcium for Activation. International Journal of Molecular Sciences. 2023; 24(11):9405. https://doi.org/10.3390/ijms24119405
Chicago/Turabian StyleWette, Stefan G., Graham D. Lamb, and Robyn M. Murphy. 2023. "Calpain-3 Is Not a Sodium Dependent Protease and Simply Requires Calcium for Activation" International Journal of Molecular Sciences 24, no. 11: 9405. https://doi.org/10.3390/ijms24119405