

SERPINE1 mRNA Binding Protein 1 Is Associated with Ischemic Stroke Risk: A Comprehensive Molecular–Genetic and Bioinformatics Analysis of SERBP1 SNPs
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. Associations of SERBP1 SNPs with the Risk of Ischemic Stroke
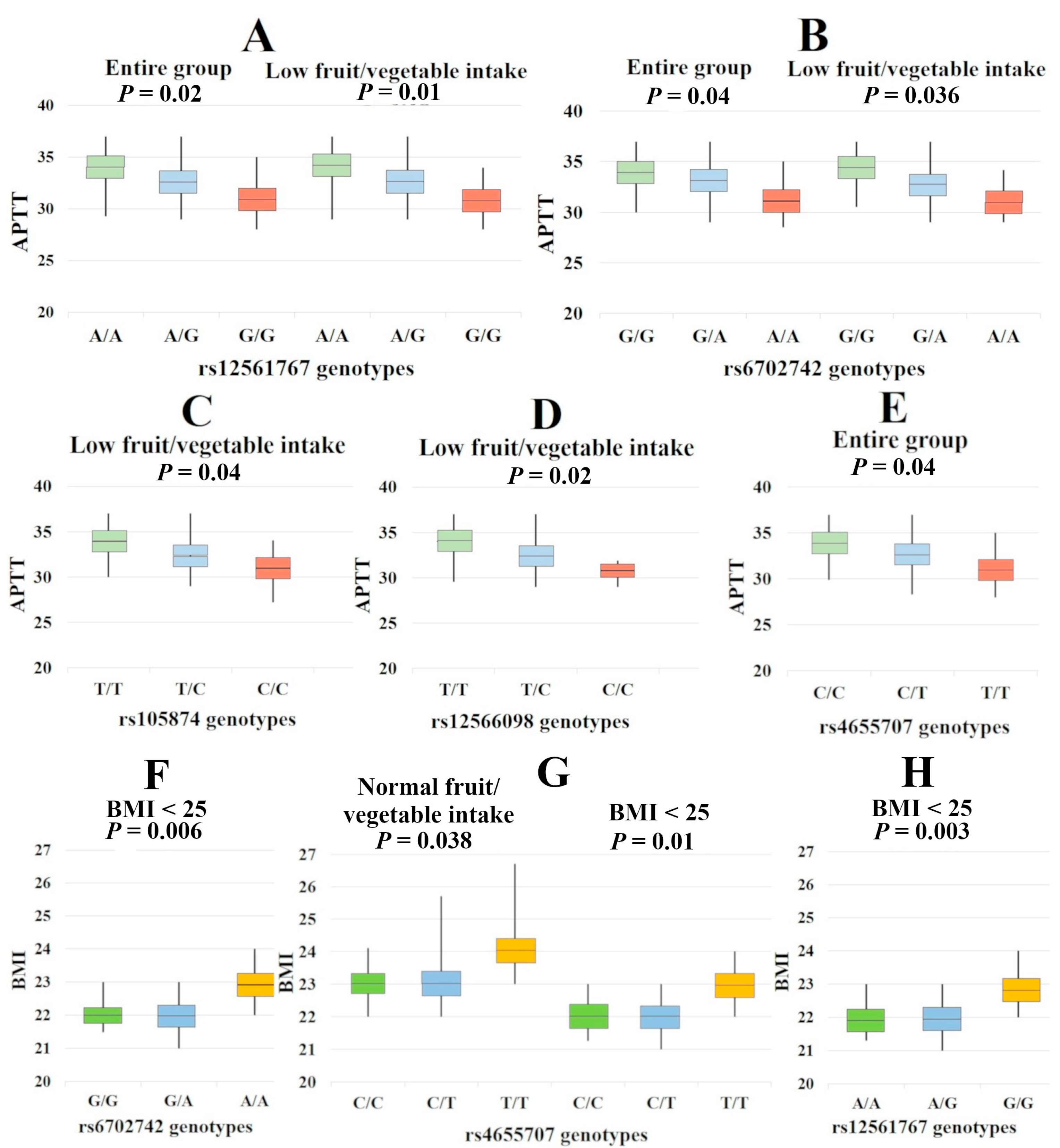
2.2. Associations of SERBP1 with the Clinical Features of IS
2.3. Bioinformatics Analysis
2.3.1. QTL-Effects
2.3.2. Analysis of Transcription Factors
2.3.3. Bioinformatic Analysis of the Associations of SERBP1 SNPs with Cerebral Stroke and Intermediate Phenotypes
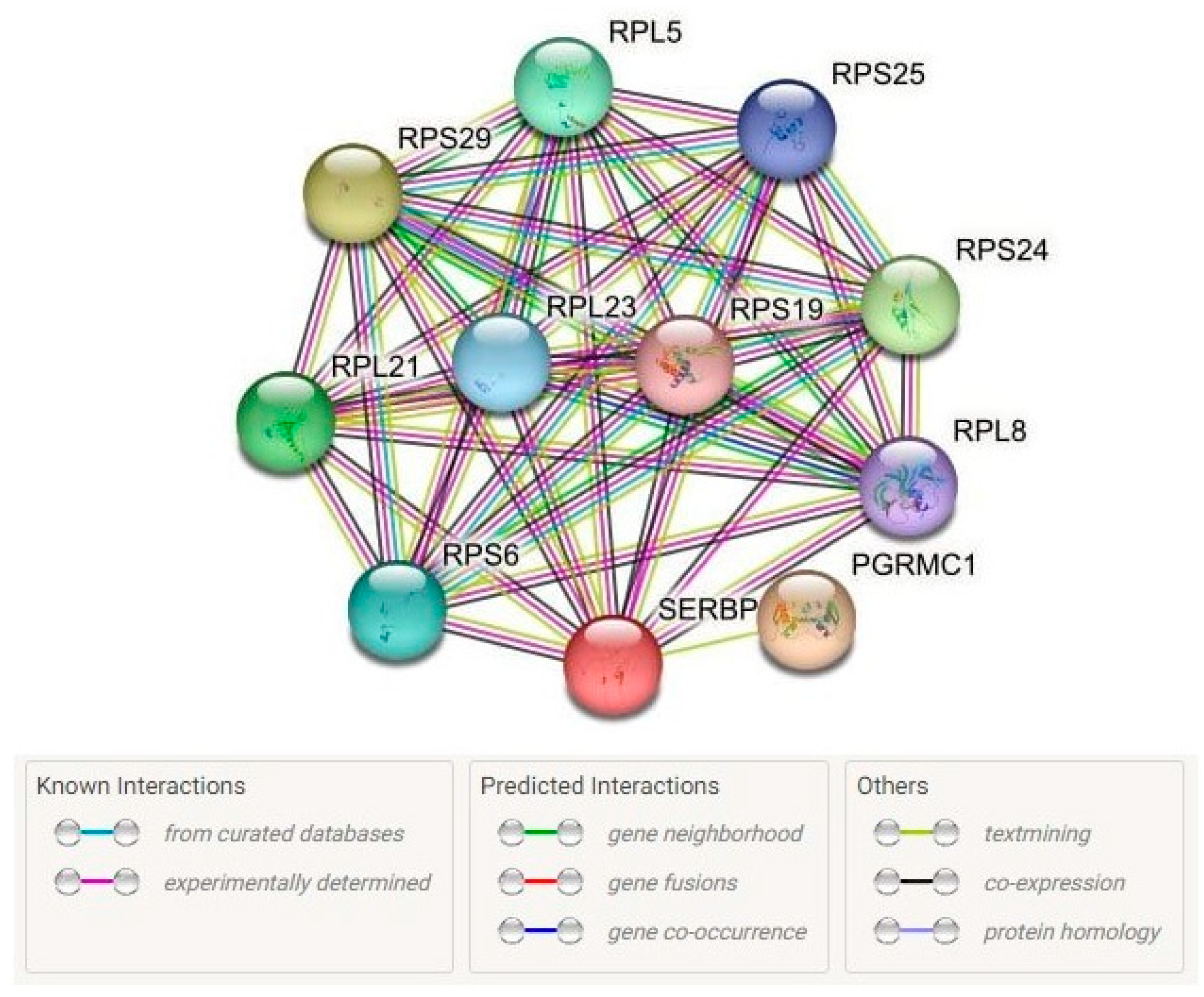
2.3.4. Protein–Protein Interactions
3. Discussion
3.1. SERBP1 SNPs and the IS Risk: Underlying Mechanisms
3.2. Sex-Specific Correlates of SERBP1
3.3. Smoking-Associated Correlates of SERBP1
3.4. Low-Physical-Activity–Associated Correlates of SERBP1
3.5. Low Fruit/Vegetable Intake–Related Correlates of SERBP1
3.6. SERBP1 SNPs and the BMI
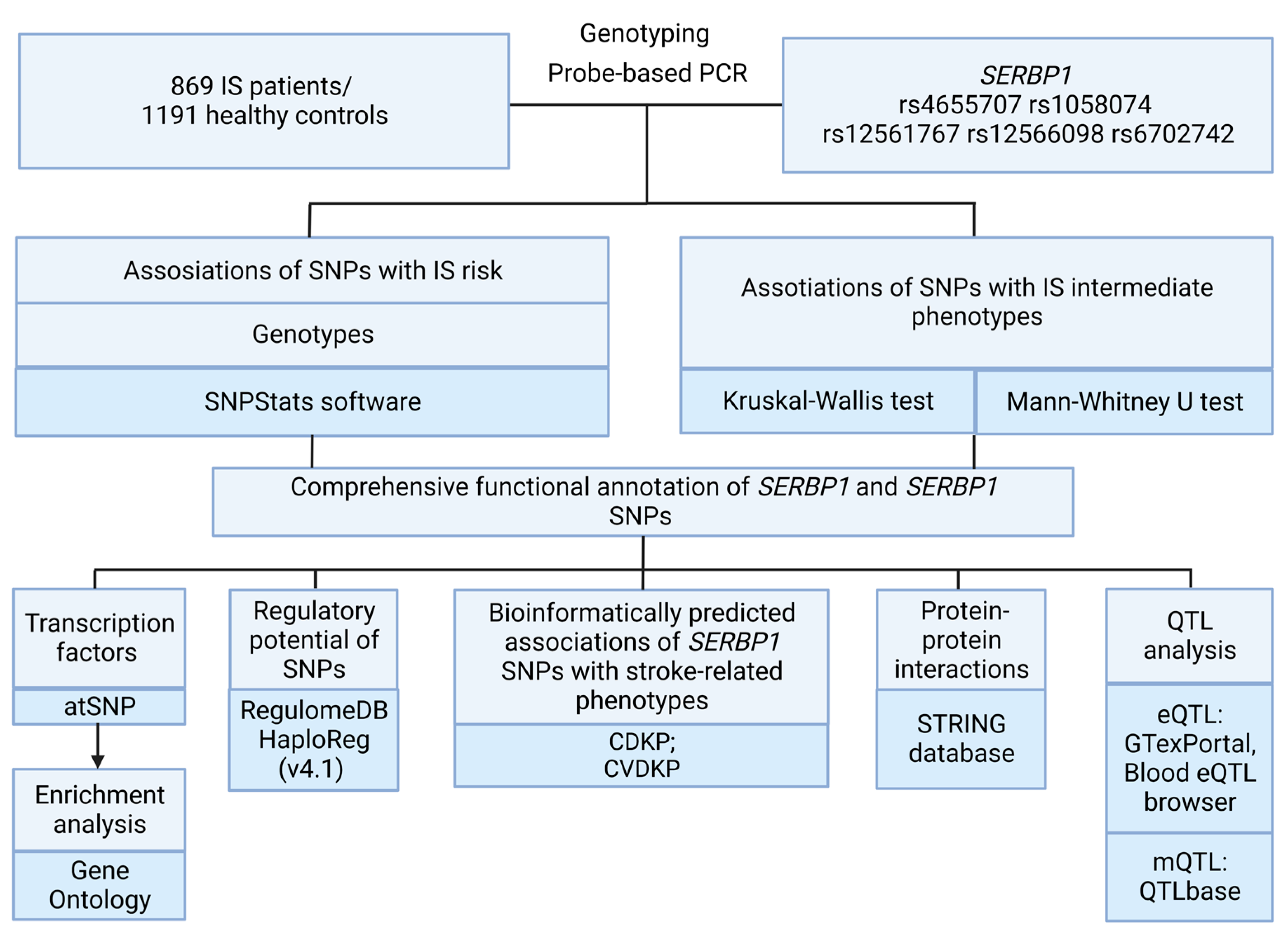
4. Materials and Methods
4.1. Genetic Analysis
4.2. Statistical and Bioinformatics Analysis
- The bioinformatic tool GTExportal (http://www.gtexportal.org/ (accessed on 21 January 2023)) was used to analyze the expression levels of the studied genes in the brain, whole blood, and blood vessels, as well as to analyze expression quantitative trait loci (eQTLs) (The GTEx Consortium, 2020). Additionally, the eQTLGene browser (https://www.eqtlgen.org/ (accessed on 21 January 2023)) was used to analyze the cis-eQTL-mediated effects of SERBP1 SNPs in blood [78];
- The methylation quantitative trait loci (mQTLs) in the brain, whole blood, and blood vessels were examined using QTLbase (http://www.mulinlab.org/qtlbase/index.html (accessed on 21 January 2023)) [79];
- Bioinformatic tools of the STRING database (https://string-db.org/ (accessed on 22 January 2023)) were used for the analysis of the main interaction partners of SERBP1 [80]. Analysis of biological processes and molecular functions reflecting interactions with main functionally related proteins also was carried out in STRING database;
- The atSNP Function Prediction online tool (http://atsnp.biostat.wisc.edu/search (accessed on 22 January 2023)) was used to evaluate the impact of SERBP1 SNPs on the binding of transcription factors (TFs) to DNA depending on the carriage of the reference/alternative alleles [81]. TFs were included based on the degree of influence of SNPs on the interaction of TFs with DNA, calculated on the basis of a positional weight matrix;
- Using the Gene Ontology online tool (http://geneontology.org/ (accessed on 23 January 2023)), it was feasible to analyze the joint involvement of TFs linked to the reference/SNP alleles in overrepresented biological processes directly related to the pathogenesis of IS [82]. Biological functions controlled by transcription factors associated with SERBP1 SNPs were used as functional groups;
- The Comparative Toxicogenomics Database (CTD) resource (http://ctdbase.org (accessed on 24 January 2023)) was used for the interpretation of environment-associated correlates of SERBP1. CTD provides the ability to analyze specific interactions between genes and chemicals in vertebrates and invertebrates based on data obtained from published scientific studies worldwide [83]. This tool was used to analyze binary interactions involving one chemical and one gene or protein;
- The Cerebrovascular Disease Knowledge Portal (CDKP) (https://cd.hugeamp.org/ (accessed on 24 January 2023)) and Cardiovascular Disease Knowledge Portal (https://cvd.hugeamp.org/ (accessed on 24 January 2023)) online tools were used for bioinformatic analyses of the associations of SERBP1 SNPs with atherosclerosis-associated diseases, intermediate phenotypes, and risk factors for IS (such as total cholesterol, LDL, BMI, etc.).
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Study Limitations
References
- Chugh, C. Acute Ischemic Stroke: Management Approach. Indian J. Crit. Care Med. Peer-Rev. Off. Publ. Indian Soc. Crit. Care Med. 2019, 23 (Suppl. S2), S140–S146. [Google Scholar] [CrossRef]
- Johnson, W.; Onuma, O.; Owolabi, M.; Sachdev, S. Stroke: A Global Response Is Needed. Bull. World Health Organ. 2016, 94, 634. [Google Scholar] [CrossRef]
- Qin, C.; Yang, S.; Chu, Y.-H.; Zhang, H.; Pang, X.-W.; Chen, L.; Zhou, L.-Q.; Chen, M.; Tian, D.-S.; Wang, W. Signaling Pathways Involved in Ischemic Stroke: Molecular Mechanisms and Therapeutic Interventions. Signal Transduct. Target. Ther. 2022, 7, 215. [Google Scholar] [CrossRef]
- Lin, J.; Wang, Y.; Wang, Y.; Pan, Y. Inflammatory Biomarkers and Risk of Ischemic Stroke and Subtypes: A 2-Sample Mendelian Randomization Study. Neurol. Res. 2020, 42, 118–125. [Google Scholar] [CrossRef]
- Polonikov, A.; Rymarova, L.; Klyosova, E.; Volkova, A.; Azarova, I.; Bushueva, O.; Bykanova, M.; Bocharova, I.; Zhabin, S.; Churnosov, M.; et al. Matrix Metalloproteinases as Target Genes for Gene Regulatory Networks Driving Molecular and Cellular Pathways Related to a Multistep Pathogenesis of Cerebrovascular Disease. J. Cell. Biochem. 2019, 120, 16467–16482. [Google Scholar] [CrossRef] [PubMed]
- Bushueva, O.; Barysheva, E.; Markov, A.; Belykh, A.; Koroleva, I.; Churkin, E.; Polonikov, A.; Ivanov, V.; Nazarenko, M. DNA Hypomethylation of the MPO Gene in Peripheral Blood Leukocytes Is Associated with Cerebral Stroke in the Acute Phase. J. Mol. Neurosci. 2021, 71, 1914–1932. [Google Scholar] [CrossRef]
- Cui, J.; Li, H.; Chen, Z.; Dong, T.; He, X.; Wei, Y.; Li, Z.; Duan, J.; Cao, T.; Chen, Q.; et al. Thrombo-Inflammation and Immunological Response in Ischemic Stroke: Focusing on Platelet-Tregs Interaction. Front. Cell. Neurosci. 2022, 16, 955385. [Google Scholar] [CrossRef]
- Janardhan, V.; Qureshi, A.I. Mechanisms of Ischemic Brain Injury. Curr. Cardiol. Rep. 2004, 6, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Mishra, V.N.; Chaurasia, R.N.; Joshi, D.; Pandey, V. Modes of Calcium Regulation in Ischemic Neuron. Indian J. Clin. Biochem. 2019, 34, 246–253. [Google Scholar] [CrossRef]
- Yamashita, T.; Abe, K. Pathophysiology of Neuronal Cell Death After Stroke. In Stroke Revisited: Pathophysiology of Stroke: From Bench to Bedside; Lee, S.-H., Ed.; Stroke Revisited; Springer: Singapore, 2020; pp. 235–241. [Google Scholar] [CrossRef]
- Ludhiadch, A.; Sharma, R.; Muriki, A.; Munshi, A. Role of Calcium Homeostasis in Ischemic Stroke: A Review. CNS Neurol. Disord.-Drug Targets-CNS Neurol. Disord. 2022, 21, 52–61. [Google Scholar] [CrossRef]
- Chan, P.H. Reactive Oxygen Radicals in Signaling and Damage in the Ischemic Brain. J. Cereb. Blood Flow Metab. 2001, 21, 2–14. [Google Scholar] [CrossRef]
- Giffard, R.G.; Yenari, M.A. Many Mechanisms for Hsp70 Protection from Cerebral Ischemia. J. Neurosurg. Anesthesiol. 2004, 16, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Frydman, J. Folding of Newly Translated Proteins In Vivo: The Role of Molecular Chaperones. Annu. Rev. Biochem. 2001, 70, 603–647. [Google Scholar] [CrossRef] [PubMed]
- Madrigal-Matute, J.; de Bruijn, J.; van Kuijk, K.; Riascos-Bernal, D.F.; Diaz, A.; Tasset, I.; Martín-Segura, A.; Gijbels, M.J.J.; Sander, B.; Kaushik, S.; et al. Protective Role of Chaperone-Mediated Autophagy against Atherosclerosis. Proc. Natl. Acad. Sci. USA 2022, 119, e2121133119. [Google Scholar] [CrossRef]
- Heaton, J.H.; Dlakic, W.M.; Dlakic, M.; Gelehrter, T.D. Identification and CDNA Cloning of a Novel RNA-Binding Protein That Interacts with the Cyclic Nucleotide-Responsive Sequence in the Type-1 Plasminogen Activator Inhibitor MRNA. J. Biol. Chem. 2001, 276, 3341–3347. [Google Scholar] [CrossRef] [PubMed]
- Griemert, E.-V.; Recarte Pelz, K.; Engelhard, K.; Schäfer, M.K.; Thal, S.C. PAI-1 but Not PAI-2 Gene Deficiency Attenuates Ischemic Brain Injury After Experimental Stroke. Transl. Stroke Res. 2019, 10, 372–380. [Google Scholar] [CrossRef]
- Hu, X.; Zan, X.; Xie, Z.; Li, Y.; Lin, S.; Li, H.; You, C. Association Between Plasminogen Activator Inhibitor-1 Genetic Polymorphisms and Stroke Susceptibility. Mol. Neurobiol. 2017, 54, 328–341. [Google Scholar] [CrossRef]
- Jood, K.; Ladenvall, P.; Tjärnlund-Wolf, A.; Ladenvall, C.; Andersson, M.; Nilsson, S.; Blomstrand, C.; Jern, C. Fibrinolytic Gene Polymorphism and Ischemic Stroke. Stroke 2005, 36, 2077–2081. [Google Scholar] [CrossRef] [PubMed]
- Taraborrelli, S. Physiology, Production and Action of Progesterone. Acta Obstet. Gynecol. Scand. 2015, 94, 8–16. [Google Scholar] [CrossRef]
- Tsuboyama, K.; Osaki, T.; Matsuura-Suzuki, E.; Kozuka-Hata, H.; Okada, Y.; Oyama, M.; Ikeuchi, Y.; Iwasaki, S.; Tomari, Y. A Widespread Family of Heat-Resistant Obscure (Hero) Proteins Protect against Protein Instability and Aggregation. PLoS Biol. 2020, 18, e3000632. [Google Scholar] [CrossRef]
- Kobzeva, K.A.; Shilenok, I.V.; Belykh, A.E.; Gurtovoy, D.E.; Bobyleva, L.A.; Krapiva, A.B.; Stetskaya, T.A.; Bykanova, M.A.; Mezhenskaya, A.A.; Lysikova, E.A.; et al. C9orf16 (BBLN) Gene, Encoding a Member of Hero Proteins, Is a Novel Marker in Ischemic Stroke Risk. Res. Results Biomed. 2022, 8, 278–292. [Google Scholar] [CrossRef]
- Schafer, K.; Müller, K.; Hecke, A.; Mounier, E.; Goebel, J.; Loskutoff, D.J.; Konstantinides, S. Enhanced Thrombosis in Atherosclerosis-Prone Mice Is Associated with Increased Arterial Expression of Plasminogen Activator Inhibitor-1. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 2097–2103. [Google Scholar] [CrossRef] [PubMed]
- de Paula Sabino, A.; Ribeiro, D.D.; Domingueti, C.P.; dos Santos, M.S.; Gadelha, T.; Dusse LM, S.; das Graças Carvalho, M.; Fernandes, A.P. Plasminogen Activator Inhibitor-1 4G/5G Promoter Polymorphism and PAI-1 Plasma Levels in Young Patients with Ischemic Stroke. Mol. Biol. Rep. 2011, 38, 5355–5360. [Google Scholar] [CrossRef]
- Stefansson, S.; Lawrence, D.A.; Argraves, W.S. Plasminogen Activator Inhibitor-1 and Vitronectin Promote the Cellular Clearance of Thrombin by Low Density Lipoprotein Receptor-Related Proteins 1 and 2. J. Biol. Chem. 1996, 271, 8215–8220. [Google Scholar] [CrossRef] [PubMed]
- Chomiki, N.; Henry, M.; Alessi, M.C.; Anfosso, F.; Juhan-Vague, I. Plasminogen Activator Inhibitor-1 Expression in Human Liver and Healthy or Atherosclerotic Vessel Walls. Thromb. Haemost. 1994, 72, 44–53. [Google Scholar] [CrossRef]
- Heaton, J.H.; Dlakic, W.M.; Gelehrter, T.D. Posttranscriptional Regulation of PAI-1 Gene Expression. Thromb. Haemost. 2003, 89, 959–966. [Google Scholar]
- Lijnen, H.R. Pleiotropic Functions of Plasminogen Activator Inhibitor-1. J. Thromb. Haemost. 2005, 3, 35–45. [Google Scholar] [CrossRef]
- Alturfan, A.A.; Eralp, L.; Emekli, N. Investigation of Inflammatory and Hemostatic Parameters in Female Patients Undergoing Total Knee Arthroplasty Surgery. Inflammation 2008, 31, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.J.; Kim, Y.R.; Lee, B.E.; Choi, Y.S.; Kim, J.H.; Shin, J.E.; Rah, H.; Cha, S.H.; Lee, W.S.; Kim, N.K. Genetic Association of Five Plasminogen Activator Inhibitor-1 (PAI-1) Polymorphisms and Idiopathic Recurrent Pregnancy Loss in Korean Women. Thromb. Haemost. 2013, 110, 742–750. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, P.; Yang, W.Y.; Wang, H.; Yang, X. IL-35, as a Newly Proposed Homeostasis-Associated Molecular Pattern, Plays Three Major Functions Including Anti-Inflammatory Initiator, Effector, and Blocker in Cardiovascular Diseases. Cytokine 2019, 122, 154076. [Google Scholar] [CrossRef]
- Fu, H.; Yu, J.; Choi, E.T.; Wang, H.; Yang, X. IL-35 Inhibits Ischemia/Hypoxia-Induced Angiogenesis, Suggesting That This Anti-Inflammatory Cytokine Plays New Roles in the Recovery Stage of Angiogenesis. Circulation 2018, 138 (Suppl. S1), 12666. [Google Scholar]
- Bushueva, O.Y.; Stetskaya, T.A.; Polonikov, A.V.; Ivanov, V.P. The relationship between polymorphism 640A>G of the CYBA gene with the risk of ischemic stroke in the population of the Central Russia. Zhurnal Nevrol. I Psikhiatrii Im. SS Korsakova 2015, 115 Pt 2, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Fava, C.; Montagnana, M.; Danese, E.; Almgren, P.; Hedblad, B.; Engström, G.; Berglund, G.; Minuz, P.; Melander, O. Homozygosity for the EPHX2 K55R Polymorphism Increases the Long-Term Risk of Ischemic Stroke in Men: A Study in Swedes. Pharmacogenet. Genomics 2010, 20, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Vaura, F.; Palmu, J.; Aittokallio, J.; Kauko, A.; Niiranen, T. Genetic, Molecular, and Cellular Determinants of Sex-Specific Cardiovascular Traits. Circ. Res. 2022, 130, 611–631. [Google Scholar] [CrossRef]
- Stetskaia, T.A.; Bushueva, O.I.; Bulgakova, I.V.; Vialykh, E.K.; Shuteeva, T.V.; Biriukov, A.E.; Ivanov, V.P.; Polonikov, A.V. Association of T174M polymorphism of the angiotensinogen gene with the higher risk of cerebral stroke in women. Ter. Arkh. 2014, 86, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Intlekofer, K.A.; Petersen, S.L. 17β-Estradiol and Progesterone Regulate Multiple Progestin Signaling Molecules in the Anteroventral Periventricular Nucleus, Ventromedial Nucleus and Sexually Dimorphic Nucleus of the Preoptic Area in Female Rats. Neuroscience 2011, 176, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Intlekofer, K.A.; Petersen, S.L. Distribution of mRNAs Encoding Classical Progestin Receptor, Progesterone Membrane Components 1 and 2, Serpine mRNA Binding Protein 1, and Progestin and ADIPOQ Receptor Family Members 7 and 8 in Rat Forebrain. Neuroscience 2011, 172, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, S.; Matsumura, K.; Kitamura, N.; Takanami, Y.; Ito, S. Multi-Omics Analysis: Repeated Exposure of a 3D Bronchial Tissue Culture to Whole-Cigarette Smoke. Toxicol. Vitro Int. J. Publ. Assoc. BIBRA 2019, 54, 251–262. [Google Scholar] [CrossRef]
- Panagiotakos, D.B.; Kokkinos, P.; Manios, Y.; Pitsavos, C. Physical Activity and Markers of Inflammation and Thrombosis Related to Coronary Heart Disease. Prev. Cardiol. 2004, 7, 190–194. [Google Scholar] [CrossRef]
- Diteepeng, T.; del Monte, F.; Luciani, M. The Long and Winding Road to Target Protein Misfolding in Cardiovascular Diseases. Eur. J. Clin. Invest. 2021, 51, e13504. [Google Scholar] [CrossRef]
- Santos-Parker, J.R.; Santos-Parker, K.S.; McQueen, M.B.; Martens, C.R.; Seals, D.R. Habitual Aerobic Exercise and Circulating Proteomic Patterns in Healthy Adults: Relation to Indicators of Healthspan. J. Appl. Physiol. 2018, 125, 1646–1659. [Google Scholar] [CrossRef]
- Needham, E.J.; Humphrey, S.J.; Cooke, K.C.; Fazakerley, D.J.; Duan, X.; Parker, B.L.; James, D.E. Phosphoproteomics of Acute Cell Stressors Targeting Exercise Signaling Networks Reveal Drug Interactions Regulating Protein Secretion. Cell Rep. 2019, 29, 1524–1538.e6. [Google Scholar] [CrossRef]
- Yoshizaki, T.; Ishihara, J.; Kotemori, A.; Yamamoto, J.; Kokubo, Y.; Saito, I.; Yatsuya, H.; Yamagishi, K.; Sawada, N.; Iwasaki, M.; et al. Association of Vegetable, Fruit, and Okinawan Vegetable Consumption with Incident Stroke and Coronary Heart Disease. J. Epidemiol. 2020, 30, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, J.; Liu, F.; Huang, K.; Yang, X.; Liu, X.; Cao, J.; Chen, S.; Shen, C.; Yu, L.; et al. Genetic Predisposition, Fruit Intake and Incident Stroke: A Prospective Chinese Cohort Study. Nutrients 2022, 14, 5056. [Google Scholar] [CrossRef] [PubMed]
- Polonikov, A.; Vialykh, E.; Vasil’eva, O.; Bulgakova, I.; Bushueva, O.; Illig, T.; Solodilova, M. Genetic Variation in Glutathione S-Transferase Genes and Risk of Nonfatal Cerebral Stroke in Patients Suffering from Essential Hypertension. J. Mol. Neurosci. 2012, 47, 511–513. [Google Scholar] [CrossRef]
- Bushueva, O.Y.U.; Bulgakova, I.V.; Ivanov, V.P.; Polonikov, A.V. Association of Flavin Monooxygenase Gene E158K Polymorphism with Chronic Heart Disease Risk. Bull. Exp. Biol. Med. 2015, 159, 776–778. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A.V.; Kotani, K.; Bushueva, O.Y.; Polonikov, A.V. Antioxidant-Related Gene Polymorphisms Associated with the Cardio-Ankle Vascular Index in Young Russians. Cardiol. Young 2016, 26, 677–682. [Google Scholar] [CrossRef] [PubMed]
- Polonikov, A.; Bocharova, I.; Azarova, I.; Klyosova, E.; Bykanova, M.; Bushueva, O.; Polonikova, A.; Churnosov, M.; Solodilova, M. The Impact of Genetic Polymorphisms in Glutamate-Cysteine Ligase, a Key Enzyme of Glutathione Biosynthesis, on Ischemic Stroke Risk and Brain Infarct Size. Life 2022, 12, 602. [Google Scholar] [CrossRef]
- Sorokin, A.V.; Kotani, K.; Bushueva, O.Y. Association of Matrix Metalloproteinase 3 and γ-Glutamyltransferase 1 Gene Polymorphisms with the Cardio-Ankle Vascular Index in Young Russians. Cardiol. Young 2016, 26, 1238–1240. [Google Scholar] [CrossRef]
- Guan, R.; Van Le, Q.; Yang, H.; Zhang, D.; Gu, H.; Yang, Y.; Sonne, C.; Lam, S.S.; Zhong, J.; Jianguang, Z.; et al. Review of Dietary Phytochemicals and Their Relation to Oxidative Stress and Human Diseases. Chemosphere 2021, 271, 129499. [Google Scholar] [CrossRef]
- Pacifici, F.; Rovella, V.; Pastore, D.; Bellia, A.; Abete, P.; Donadel, G.; Santini, S.; Beck, H.; Ricordi, C.; Daniele, N.D.; et al. Polyphenols and Ischemic Stroke: Insight into One of the Best Strategies for Prevention and Treatment. Nutrients 2021, 13, 1967. [Google Scholar] [CrossRef]
- Vita, J.A. Polyphenols and Cardiovascular Disease: Effects on Endothelial and Platelet Function. Am. J. Clin. Nutr. 2005, 81, 292S–297S. [Google Scholar] [CrossRef] [PubMed]
- Bazzano, L.A.; He, J.; Ogden, L.G.; Loria, C.; Vupputuri, S.; Myers, L.; Whelton, P.K. Dietary Intake of Folate and Risk of Stroke in US Men and Women: NHANES I Epidemiologic Follow-up Study. National Health and Nutrition Examination Survey. Stroke 2002, 33, 1183–1188. [Google Scholar] [CrossRef]
- Margetic, S.; Gazzola, C.; Pegg, G.G.; Hill, R.A. Leptin: A Review of Its Peripheral Actions and Interactions. Int. J. Obes. 2002, 26, 1407–1433. [Google Scholar] [CrossRef] [PubMed]
- Funahashi, H.; Yada, T.; Suzuki, R.; Shioda, S. Distribution, Function, and Properties of Leptin Receptors in the Brain. Int. Rev. Cytol. 2003, 224, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Liu, F.; Li, R.W.; Yang, R.; Wang, Y.; Xue, C.; Tang, Q. Triacylglycerol Rich in Docosahexaenoic Acid Regulated Appetite via the Mediation of Leptin and Intestinal Epithelial Functions in High-Fat, High-Sugar Diet-Fed Mice. J. Nutr. Biochem. 2022, 99, 108856. [Google Scholar] [CrossRef]
- Bungau, S.; Behl, T.; Tit, D.M.; Banica, F.; Bratu, O.G.; Diaconu, C.C.; Nistor-Cseppento, C.D.; Bustea, C.; Aron, R.A.C.; Vesa, C.M. Interactions between Leptin and Insulin Resistance in Patients with Prediabetes, with and without NAFLD. Exp. Ther. Med. 2020, 20, 197. [Google Scholar] [CrossRef]
- Gruzdeva, O.; Borodkina, D.; Uchasova, E.; Dyleva, Y.; Barbarash, O. Leptin Resistance: Underlying Mechanisms and Diagnosis. Diabetes Metab. Syndr. Obes. 2019, 12, 191–198. [Google Scholar] [CrossRef]
- Izquierdo, A.G.; Crujeiras, A.B.; Casanueva, F.F.; Carreira, M.C. Leptin, Obesity, and Leptin Resistance: Where Are We 25 Years Later? Nutrients 2019, 11, 2704. [Google Scholar] [CrossRef]
- Loskutoff, D.J.; Samad, F. The Adipocyte and Hemostatic Balance in Obesity: Studies of PAI-1. Arterioscler. Thromb. Vasc. Biol. 1998, 18, 1–6. [Google Scholar] [CrossRef]
- Kershaw, E.E.; Flier, J.S. Adipose Tissue as an Endocrine Organ. J. Clin. Endocrinol. Metab. 2004, 89, 2548–2556. [Google Scholar] [CrossRef] [PubMed]
- Sergeeva, C.N.; Sokorev, S.N.; Efremova, O.A.; Sorokina, I.N. Analysis of the level of population endogamia as the basis of population-genetic and medical-genetic studies. Res. Results Biomed. 2021, 7, 375–387. [Google Scholar] [CrossRef]
- Vialykh, E.K.; Solidolova, M.A.; Bushueva, O.I.; Bulgakova, I.V.; Polonikov, A.V. [Catalase gene polymorphism is associated with increased risk of cerebral stroke in hypertensive patients]. Zhurnal Nevrol. I Psikhiatrii Im. SS Korsakova 2012, 112 Pt 2, 3–7. [Google Scholar]
- World Health Organization. Diet, Nutrition, and the Prevention of Chronic Diseases: Report of a Joint WHO/FAO Expert Consultation; World Health Organization: Geneva, Switzerland, 2003; Volume 916. [Google Scholar]
- Gutin, B.; Owens, S. The Influence of Physical Activity on Cardiometabolic Biomarkers in Youths: A Review. Pediatr. Exerc. Sci. 2011, 23, 169–185. [Google Scholar] [CrossRef]
- Fernandes, R.A.; Zanesco, A. Early Physical Activity Promotes Lower Prevalence of Chronic Diseases in Adulthood. Hypertens. Res. Off. J. Jpn. Soc. Hypertens. 2010, 33, 926–931. [Google Scholar] [CrossRef]
- Colbert, L.H.; Visser, M.; Simonsick, E.M.; Tracy, R.P.; Newman, A.B.; Kritchevsky, S.B.; Pahor, M.; Taaffe, D.R.; Brach, J.; Rubin, S.; et al. Physical Activity, Exercise, and Inflammatory Markers in Older Adults: Findings from The Health, Aging and Body Composition Study. J. Am. Geriatr. Soc. 2004, 52, 1098–1104. [Google Scholar] [CrossRef]
- World Health Organization. WHO European Regional Obesity Report 2022; World Health Organization. Regional Office for Europe: Geneva, Switzerland, 2022. [Google Scholar]
- Bushueva, O.; Solodilova, M.; Ivanov, V.; Polonikov, A. Gender-Specific Protective Effect of the −463G>A Polymorphism of Myeloperoxidase Gene against the Risk of Essential Hypertension in Russians. J. Am. Soc. Hypertens. 2015, 9, 902–906. [Google Scholar] [CrossRef] [PubMed]
- Bushueva, O. Single Nucleotide Polymorphisms in Genes Encoding Xenobiotic Metabolizing Enzymes Are Associated with Predisposition to Arterial Hypertension. Res. Results Biomed. 2020, 6, 447–456. [Google Scholar] [CrossRef]
- Dong, S.; Boyle, A.P. Predicting Functional Variants in Enhancer and Promoter Elements Using RegulomeDB. Hum. Mutat. 2019, 40, 1292–1298. [Google Scholar] [CrossRef]
- Ward, L.D.; Kellis, M. HaploReg: A Resource for Exploring Chromatin States, Conservation, and Regulatory Motif Alterations within Sets of Genetically Linked Variants. Nucleic Acids Res. 2012, 40, D930–D934. [Google Scholar] [CrossRef]
- Koressaar, T.; Remm, M. Enhancements and Modifications of Primer Design Program Primer3. Bioinforma. Oxf. Engl. 2007, 23, 1289–1291. [Google Scholar] [CrossRef]
- Strimmer, K. A Unified Approach to False Discovery Rate Estimation. BMC Bioinform. 2008, 9, 303. [Google Scholar] [CrossRef]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A Web Tool for the Analysis of Association Studies. Bioinforma. Oxf. Engl. 2006, 22, 1928–1929. [Google Scholar] [CrossRef] [PubMed]
- Andrade, C. Multiple Testing and Protection Against a Type 1 (False Positive) Error Using the Bonferroni and Hochberg Corrections. Indian J. Psychol. Med. 2019, 41, 99–100. [Google Scholar] [CrossRef] [PubMed]
- Võsa, U.; Claringbould, A.; Westra, H.-J.; Bonder, M.J.; Deelen, P.; Zeng, B.; Kirsten, H.; Saha, A.; Kreuzhuber, R.; Kasela, S.; et al. Unraveling the Polygenic Architecture of Complex Traits Using Blood EQTL Metaanalysis. bioRxiv 2018. [Google Scholar] [CrossRef]
- Zheng, Z.; Huang, D.; Wang, J.; Zhao, K.; Zhou, Y.; Guo, Z.; Zhai, S.; Xu, H.; Cui, H.; Yao, H.; et al. QTLbase: An Integrative Resource for Quantitative Trait Loci across Multiple Human Molecular Phenotypes. Nucleic Acids Res. 2020, 48, D983–D991. [Google Scholar] [CrossRef]
- von Mering, C.; Jensen, L.J.; Snel, B.; Hooper, S.D.; Krupp, M.; Foglierini, M.; Jouffre, N.; Huynen, M.A.; Bork, P. STRING: Known and Predicted Protein–Protein Associations, Integrated and Transferred across Organisms. Nucleic Acids Res. 2005, 33, D433–D437. [Google Scholar] [CrossRef]
- Shin, S.; Hudson, R.; Harrison, C.; Craven, M.; Keleş, S. AtSNP Search: A Web Resource for Statistically Evaluating Influence of Human Genetic Variation on Transcription Factor Binding. Bioinforma. Oxf. Engl. 2019, 35, 2657–2659. [Google Scholar] [CrossRef] [PubMed]
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 Years and Still GOing Strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): Update 2021. Nucleic Acids Res. 2021, 49, D1138–D1143. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Genetic Variant | Effect Allele | Other Allele | N | OR (95% CI) 1 | p 2 |
|---|---|---|---|---|---|
| rs4655707 | T | C | 1883 | 1.07 [0.93; 1.23] | 0.35 |
| rs1058074 | C | T | 1915 | 1.15 [0.99; 1.35] | 0.071 |
| rs12561767 * | G | A | 1857 | 1.11 [0.96; 1.27] | 0.16 |
| rs12566098 * | C | G | 2060 | 1.28 [1.10; 1.49] | 0.001 |
| rs6702742 | A | G | 1898 | 1.04 [0.91; 1.20] | 0.54 |
| Genetic Variant | Effect Allele | Other Allele | N | OR (95% CI) 1 | p 2 (Pbonf) | N | OR (95% CI) 1 | p 2 (Pbonf) |
|---|---|---|---|---|---|---|---|---|
| Males | Females | |||||||
| rs4655707 | T | C | 929 | 1.08 [0.90; 1.31] | 0.39 | 954 | 1.07 [0.88; 1.29] | 0.51 |
| rs1058074 | C | T | 950 | 1.12 [0.91; 1.38] | 0.29 | 965 | 1.29 [1.05; 1.58] | 0.02 |
| rs12561767 * | G | A | 918 | 1.12 [0.93; 1.35] | 0.23 | 939 | 1.09 [0.90; 1.32] | 0.39 |
| rs12566098 * | C | G | 1026 | 1.25 [1.02; 1.53] | 0.03 | 1124 | 1.39 [1.14; 1.71] | 0.002 |
| rs6702742 | A | G | 939 | 1.10 [0.92; 1.32] | 0.29 | 959 | 1.00 [0.82; 1.20] | 0.97 |
| Nonsmokers | Smokers | |||||||
| rs4655707 | T | C | 1281 | 1.11 [0.94; 1.31] | 0.22 | 602 | 0.93 [0.73; 1.20] | 0.59 |
| rs1058074 | C | T | 1317 | 1.32 [1.10; 1.59] | 0.003 | 598 | 0.87 [0.67; 1.14] | 0.32 |
| rs12561767 * | G | A | 1277 | 1.18 [1.00; 1.39] | 0.054 | 580 | 0.94 [0.73; 1.21] | 0.62 |
| rs12566098 * | C | G | 1400 | 1.43 [1.20; 1.72] | 1 × 10−4 | 660 | 1.02 [0.79; 1.32] | 0.89 |
| rs6702742 | A | G | 1305 | 1.09 [0.93; 1.29] | 0.29 | 593 | 0.92 [0.72; 0.72] | 0.49 |
| Low physical activity (f+) | Normal physical activity level (f-) | |||||||
| rs4655707 | T | C | 1387 | 1.07 [0.89; 1.28] | 0.46 (0.92) | 1546 | 1.06 [0.90; 1.24] | 0.48 (0.96) |
| rs1058074 | C | T | 1400 | 1.26 [1.04; 1.54] | 0.02 (0.04) | 1567 | 1.14 [0.96; 1.36] | 0.14 (0.28) |
| rs12561767 * | G | A | 1364 | 1.17 [0.98; 1.40] | 0.086 (0.172) | 1526 | 1.05 [0.90; 1.23] | 0.54 (1.0) |
| rs12566098 * | C | G | 1525 | 1.39 [1.15; 1.68] | 9 × 10−4 (0.002) | 1692 | 1.26 [1.06; 1.49] | 0.009 (0.02) |
| rs6702742 | A | G | 1411 | 1.10 [0.93; 1.31] | 0.3 (0.6) | 1564 | 1.00 [0.85; 1.17] | 0.99 (1.0) |
| Low fruit/vegetable intake (f+) | Normal fruit/vegetable intake (f-) | |||||||
| rs4655707 | T | C | 1492 | 1.08 [0.92; 1.28] | 0.32 (0.64) | 1441 | 1.04 [0.87; 1.23] | 0.68 (1.0) |
| rs1058074 | C | T | 1514 | 1.24 [1.04; 1.48] | 0.02 (0.04) | 1453 | 1.13 [0.93; 1.37] | 0.21 (0.42) |
| rs12561767 * | G | A | 1468 | 1.08 [0.92; 1.28] | 0.33 (0.66) | 1422 | 1.11 [0.94; 1.32] | 0.22 (0.44) |
| rs12566098* | C | G | 1638 | 1.37 [1.15; 1.63] | 4 × 10−4 (8 × 10−4) | 1579 | 1.24 [1.02; 1.49] | 0.03 (0.06) |
| rs6702742 | A | G | 1510 | 1.04 [0.89; 1.22] | 0.62 (1.0) | 1465 | 1.04 [0.87; 1.23] | 0.67 (1.0) |
| BMI ≥ 25 | BMI < 25 | |||||||
| rs4655707 | T | C | 1234 | 1.20 [0.95; 1.53] | 0.13 (0.26) | 1443 | 1.01 [0.85; 1.20] | 0.91 (1.0) |
| rs1058074 | C | T | 1252 | 1.47 [1.14; 1.89] | 0.0045 (0.007) | 1456 | 1.03 [0.85; 1.25] | 0.74 (1.0) |
| rs12561767 * | G | A | 1213 | 1.25 [0.98; 1.59] | 0.071 (0.142) | 1430 | 1.03 [0.87; 1.22] | 0.71 (1.0) |
| rs12566098 * | C | G | 1366 | 1.52 [1.19; 1.95] | 0.001 (0.002) | 1579 | 1.17 [0.97; 1.41] | 0.096 (0.192) |
| rs6702742 | A | G | 1272 | 1.22 [0.97; 1.54] | 0.091 (0.182) | 1478 | 1.00 [0.84; 1.18] | 0.99 (1.0) |
| SNP | GTEx Portal Data (https://gtexportal.org (Accessed on 21 January 2023)) | eQTLGene Browser Data (https://www.eqtlgen.org/ (Accessed on 21 January 2023)) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Effect Allele | Gene Expressed | p-Value | Effect (NES) | Tissue | Effect Allele | Gene Expressed | Z-Score | p-Value | |
| rs4655707 SERBP1 (T/C) | C | IL12RB2 | 6.2 × 10−10 | ↑(0.28) | Artery—Tibial | T | IL12RB2 | ↑(25.93) | 2.93 × 10−148 |
| SERBP1 | 1.8 × 10−6 | ↓(−0.091) | Artery—Tibial | ||||||
| IL12RB2 | 3.8 × 10−8 | ↓(−0.13) | Whole Blood | ||||||
| rs1058074 SERBP1 (C/T) | T | IL12RB2 | 1.5 × 10−11 | ↑(0.30) | Artery—Tibial | C | IL12RB2 | ↑(16.31) | 9.01 × 10−60 |
| SERBP1 | 8.7 × 10−10 | ↓(−0.12) | Artery—Tibial | ||||||
| IL12RB2 | 5.6 × 10−7 | ↓(−0.12) | Whole Blood | ||||||
| IL12RB2 | 3.6 × 10−5 | ↑(0.25) | Artery—Aorta | ||||||
| rs12561767 * SERBP1 (G/A) | A | IL12RB2 | 2.8 × 10−10 | ↑(0.28) | Artery—Tibial | G | IL12RB2 | ↑(26.20) | 2.92 × 10−151 |
| IL12RB2 | 9.1 × 10−9 | ↓(−0.14) | Whole Blood | ||||||
| SERBP1 | 2.3 × 10−6 | ↓(−0.09) | Artery—Tibial | ||||||
| rs12566098 * SERBP1 (C/G) | G | SERBP1 | 1.3 × 10−9 | ↓(−0.12) | Artery—Tibial | C | IL12RB2 | ↑(17.40) | 8.60 × 10−68 |
| IL12RB2 | 3.7 × 10−7 | ↓(−0.13) | Whole Blood | ||||||
| IL12RB2 | 2.3 × 10−11 | ↑(0.30) | Artery—Tibial | ||||||
| rs6702742 SERBP1 (A/G) | G | IL12RB2 | 3.9 × 10−10 | ↑(0.27) | Artery—Tibial | A | IL12RB2 | ↑(26.29) | 2.42 × 10−152 |
| IL12RB2 | 1.0 × 10−9 | ↓(−0.14) | Whole Blood | ||||||
| SERBP1 | 9.5 × 10−7 | ↓(−0.092) | Artery—Tibial | ||||||
| Trait | Effect Allele | Tissue | Effect Size (Beta) | FDR |
|---|---|---|---|---|
| rs4655707 SERBP1 | ||||
| cg24364144 (chr1:68102479) | C | Brain-Prefrontal Cortex | 0.02 | 9.5 × 10−14 |
| rs1058074 SERBP1 | ||||
| cg24364144 (chr1:68102479) | C | Brain-Prefrontal Cortex | −0.01 | 1.6 × 10−10 |
| rs12561767 SERBP1 * | ||||
| cg24364144 (chr1:68102479) | A | Brain-Prefrontal Cortex | 0.02 | 9.5 × 10−14 |
| rs12566098 SERBP1 * | ||||
| cg24364144 (chr1:68102479) | C | Brain-Prefrontal Cortex | −0.01 | 5.1 × 10−11 |
| rs6702742 SERBP1 | ||||
| cg24364144 (chr1:68102479) | G | Brain-Prefrontal Cortex | 0.01 | 6.5 × 10−13 |
| № | SNPs | Phenotype | p-Value | Beta (OR) | Sample Size |
|---|---|---|---|---|---|
| 1. | rs4655707 (T/C) | 1 TOAST other undetermined | 0.01 | OR▼ 0.7757 | 9487 |
| 2. | 1 TOAST cardio-aortic embolism | 0.02 | OR▼ 0.7946 | 9470 | |
| 3. | 1 Lacunar stroke | 0.043 | OR▼ 0.9611 | 28,530 | |
| 4. | 2 LDL cholesterol | 0.015 | Beta▼ −0.0031 | 2,028,070 | |
| 5. | 2 Leptin | 0.038 | Beta▲ 0.0097 | 36,525 | |
| 6. | rs1058074 (C/T) | 1 Lacunar stroke | 0.020 | OR▼ 0.9546 | 28,530 |
| 7. | 1 TOAST other undetermined | 0.021 | OR▼ 0.9546 | 9487 | |
| 8. | 2 Arm fat ratio | 0.007 | Beta▼ −0.0013 | 232,276 | |
| 9. | 2 Dyslipidemia | 0.020 | OR▲ 1.0322 | 56,375 | |
| 10. | 2 LDL cholesterol | 0.020 | Beta▼ −0.0029 | 2,010,200 | |
| 11. | 2 Leptin | 0.037 | Beta▲ 0.0108 | 32,169 | |
| 12. | 2 Non-HDL cholesterol | 0.042 | Beta▼ −0.0032 | 1,096,010 | |
| 13. | 2 Total cholesterol | 0.049 | Beta▼ −0.0026 | 1,921,240 | |
| 14. | rs12561767 * (G/A) | 1 TOAST other undetermined | 0.010 | OR▼ 0.7775 | 9487 |
| 15. | 2 LDL cholesterol | 0.010 | Beta▼ −0.0032 | 2,014,850 | |
| 16. | 2 Dyslipidemia | 0.040 | OR▲ 1.0264 | 56,375 | |
| 17. | 2 Leptin | 0.036 | Beta▲ 0.0098 | 36,521 | |
| 18. | rs12566098 * (C/G) | 1 TOAST small artery occlusion | 0.047 | OR▲ 2.5943 | 254,558 |
| 19. | 2 LDL cholesterol | 0.010 | Beta▼ −0.0032 | 2,021,740 | |
| 20. | 2 Arm fat ratio | 0.010 | Beta▼ −0.0112 | 232,276 | |
| 21. | 2 Dyslipidemia | 0.020 | OR▲ 1.0315 | 56,375 | |
| 22. | 2 Leptin | 0.020 | Beta▲ 0.0118 | 34,005 | |
| 23. | 2 Non-HDL cholesterol | 0.030 | Beta▼ −0.0035 | 1,108,320 | |
| 24. | rs6702742 (A/G) | 1 TOAST other undetermined | 0.009 | OR▼ 0.7726 | 9487 |
| 25. | 1 TOAST cardio-aortic embolism | 0.025 | OR▼ 0.7952 | 9470 | |
| 26. | 2 Leptin | 0.002 | Beta▲ 0.0188 | 29,651 | |
| 27. | 2 HDL3 cholesterol | 0.021 | Beta▲ 0.0336 | 10,984 | |
| 28. | 2 LDL cholesterol | 0.024 | Beta▼ −0.0027 | 2,144,190 | |
| 29. | 2 Leptin adj BMI | 0.028 | Beta▲ 0.0170 | 22,924 | |
| 30. | 2 Dyslipidemia | 0.044 | OR▲ 1.0264 | 56,375 |
| Baseline and Clinical Characteristics | IS Patients (n = 869) | Controls (n = 1191) | p-Value | |
|---|---|---|---|---|
| Age, Me [Q1; Q3] | 62 [55; 70] | 59 [53; 66] | <0.001 | |
| Gender | Males, N (%) | 482 (55.5%) | 544 (45.7%) | <0.001 |
| Females, N (%) | 387 (44.5%) | 647 (54.3%) | ||
| Smoking | Yes, N (%) | 419 (48.2%) | 241 (20.2%) | <0.001 |
| No, N (%) | 450 (51.8%) | 950 (79.8%) | ||
| Low physical activity | Yes, N (%) | 334 (40.0%) | ND | |
| No, N (%) | 501 (60.0%) | |||
| Low fruit/vegetable consumption | Yes, N (%) | 447 (53.53%) | ND | |
| No, N (%) | 388 (46.47%) | |||
| Coronary artery disease | Yes, N (%) | 264 (26.72%) | - | |
| No, N (%) | 724 (73.28%) | - | ||
| Type 2 diabetes mellitus | Yes, N (%) | 101 (12.11%) | - | |
| No, N (%) | 733 (87.89%) | - | ||
| Body mass index, Me [Q1; Q3] | 23 [22; 26] (n = 563) | - | ||
| Overweight | Normal weight (BMI = 18.5–24.9), N (%) | 388 (68.92%) | ND | |
| Overweight (BMI of 25–29.9), N (%) | 118 (20.96%) | |||
| Obesity (BMI of 30 or greater), N (%) | 57 (10.12%) | |||
| Family history of cerebrovascular diseases | Yes, N (%) | 311 (34.44%) | ND | |
| No, N (%) | 592 (65.56%) | ND | ||
| Age at onset of stroke, Me [Q1; Q3] | 61 [53; 69] (n = 851) | - | ||
| Number of strokes including event in question | 1, N (%) | 751 (88.35%) | - | |
| 2, N (%) | 86 (10.12%) | - | ||
| 3, N (%) | 13 (1.53%) | - | ||
| Type of stroke | Atherothrombotic, N (%) | 616 (70.89%) | - | |
| Cardioembolic, N (%) | 160 (18.41%) | - | ||
| Unspecified, N (%) | 93 (10.70%) | - | ||
| Stroke localization | Right/left middle cerebral artery basin, N (%) | 705 (83.04%) | - | |
| Vertebrobasilar basin, N (%) | 144 (16.96%) | - | ||
| Area of lesion in stroke, mm2, Me [Q1; Q3] | 99.50 [30; 461] (n = 776) | - | ||
| Total cholesterol, mmol/L, Me [Q1; Q3] | 5.2 [4.4; 5.8] (n = 577) | ND | ||
| Triglycerides, mmol/L, Me [Q1; Q3] | 1.3 [1.09; 1.80] (n = 571) | ND | ||
| Glucose level, mmol/L, Me [Q1; Q3] | 4.8 [4.3; 5.5] (n = 840) | ND | ||
| Prothrombin time, seconds, Me [Q1; Q3] | 10.79 [10.14; 11.70] (n = 827) | ND | ||
| International normalized ratio, Me [Q1; Q3] | 1 [0.93; 1.09] (n = 566) | ND | ||
| Activated partial thromboplastin time, seconds, Me [Q1; Q3] | 32.6 [29; 37] (n = 569) | ND | ||
| Alanin aminotransferase, IU/L | 21.9 [18; 31.2] (n = 646) | ND | ||
| Aspartate aminotransferase, IU/L | 28.2 [20.5; 37.4] (n = 646) | ND | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shilenok, I.; Kobzeva, K.; Stetskaya, T.; Freidin, M.; Soldatova, M.; Deykin, A.; Soldatov, V.; Churnosov, M.; Polonikov, A.; Bushueva, O. SERPINE1 mRNA Binding Protein 1 Is Associated with Ischemic Stroke Risk: A Comprehensive Molecular–Genetic and Bioinformatics Analysis of SERBP1 SNPs. Int. J. Mol. Sci. 2023, 24, 8716. https://doi.org/10.3390/ijms24108716
Shilenok I, Kobzeva K, Stetskaya T, Freidin M, Soldatova M, Deykin A, Soldatov V, Churnosov M, Polonikov A, Bushueva O. SERPINE1 mRNA Binding Protein 1 Is Associated with Ischemic Stroke Risk: A Comprehensive Molecular–Genetic and Bioinformatics Analysis of SERBP1 SNPs. International Journal of Molecular Sciences. 2023; 24(10):8716. https://doi.org/10.3390/ijms24108716
Chicago/Turabian StyleShilenok, Irina, Ksenia Kobzeva, Tatiana Stetskaya, Maxim Freidin, Maria Soldatova, Alexey Deykin, Vladislav Soldatov, Mikhail Churnosov, Alexey Polonikov, and Olga Bushueva. 2023. "SERPINE1 mRNA Binding Protein 1 Is Associated with Ischemic Stroke Risk: A Comprehensive Molecular–Genetic and Bioinformatics Analysis of SERBP1 SNPs" International Journal of Molecular Sciences 24, no. 10: 8716. https://doi.org/10.3390/ijms24108716