
OsNPR1 Enhances Rice Resistance to Xanthomonas oryzae pv. oryzae by Upregulating Rice Defense Genes and Repressing Bacteria Virulence Genes

 and
and 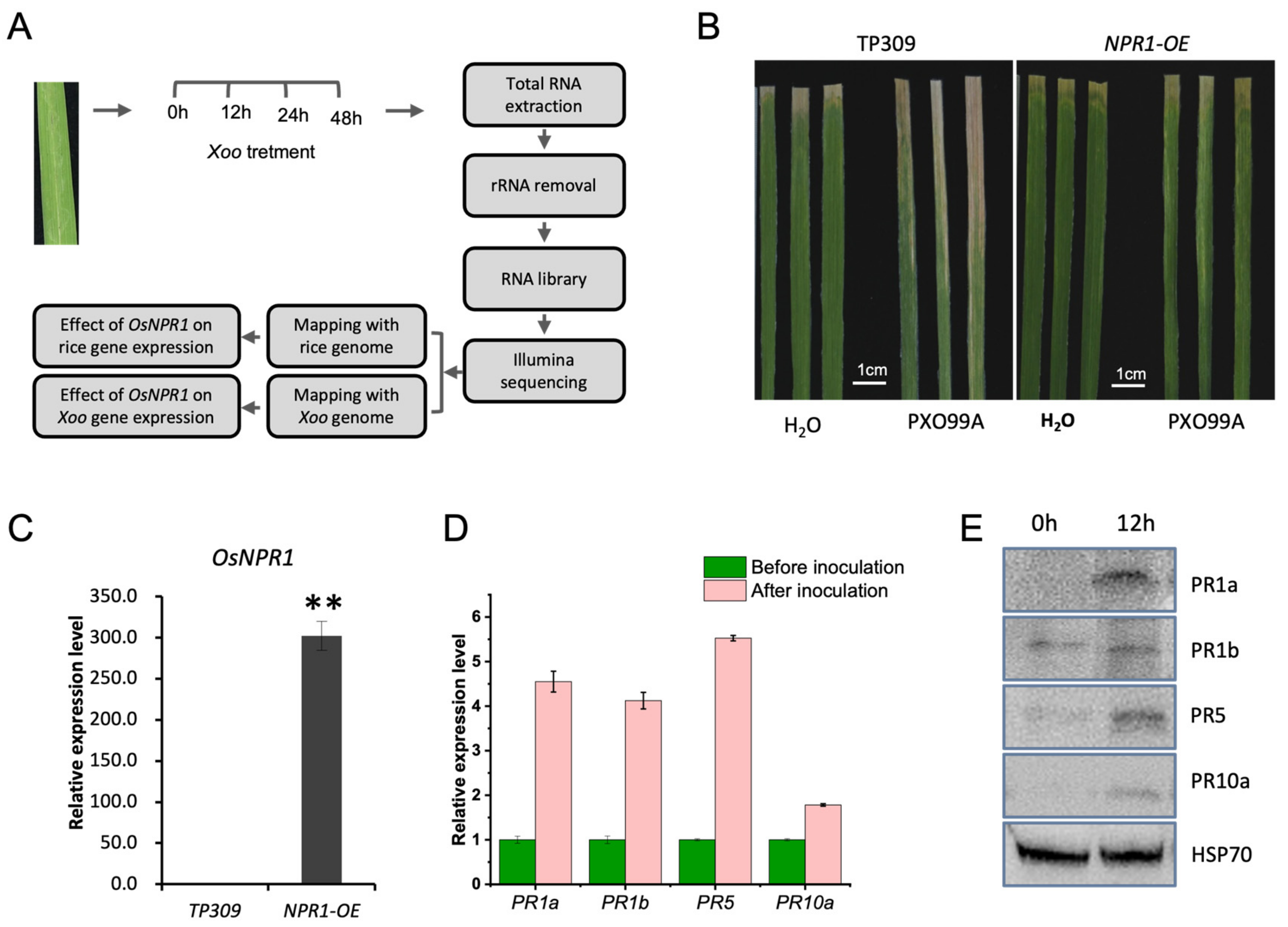{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
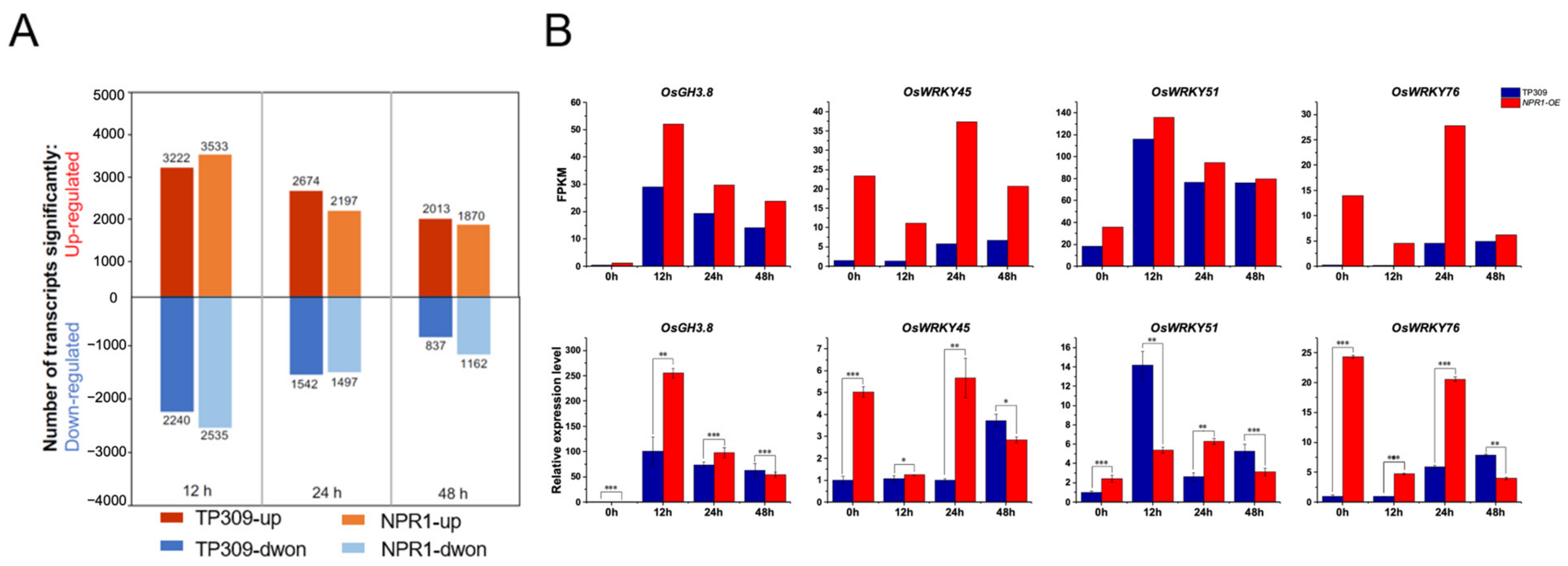
2.1. Dual RNAseq Was Developed to Evaluate the Effects of OsNPR1 on Gene Expression of Rice and Xoo
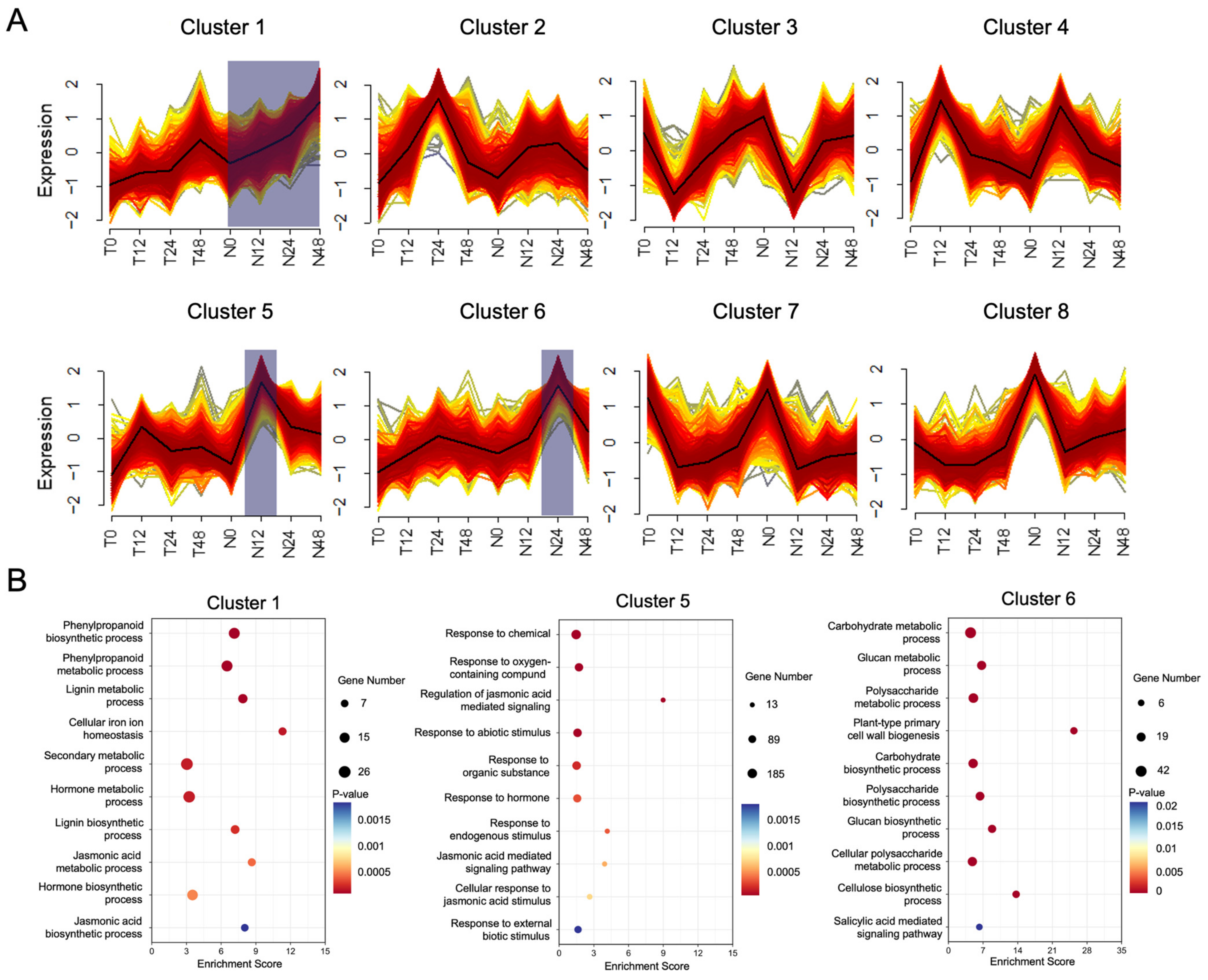
2.2. Genes Involved in Cell Wall Biosynthesis and SA Signaling Pathways Were Upregulated by OsNPR1 Overexpression after PXO99A Infection
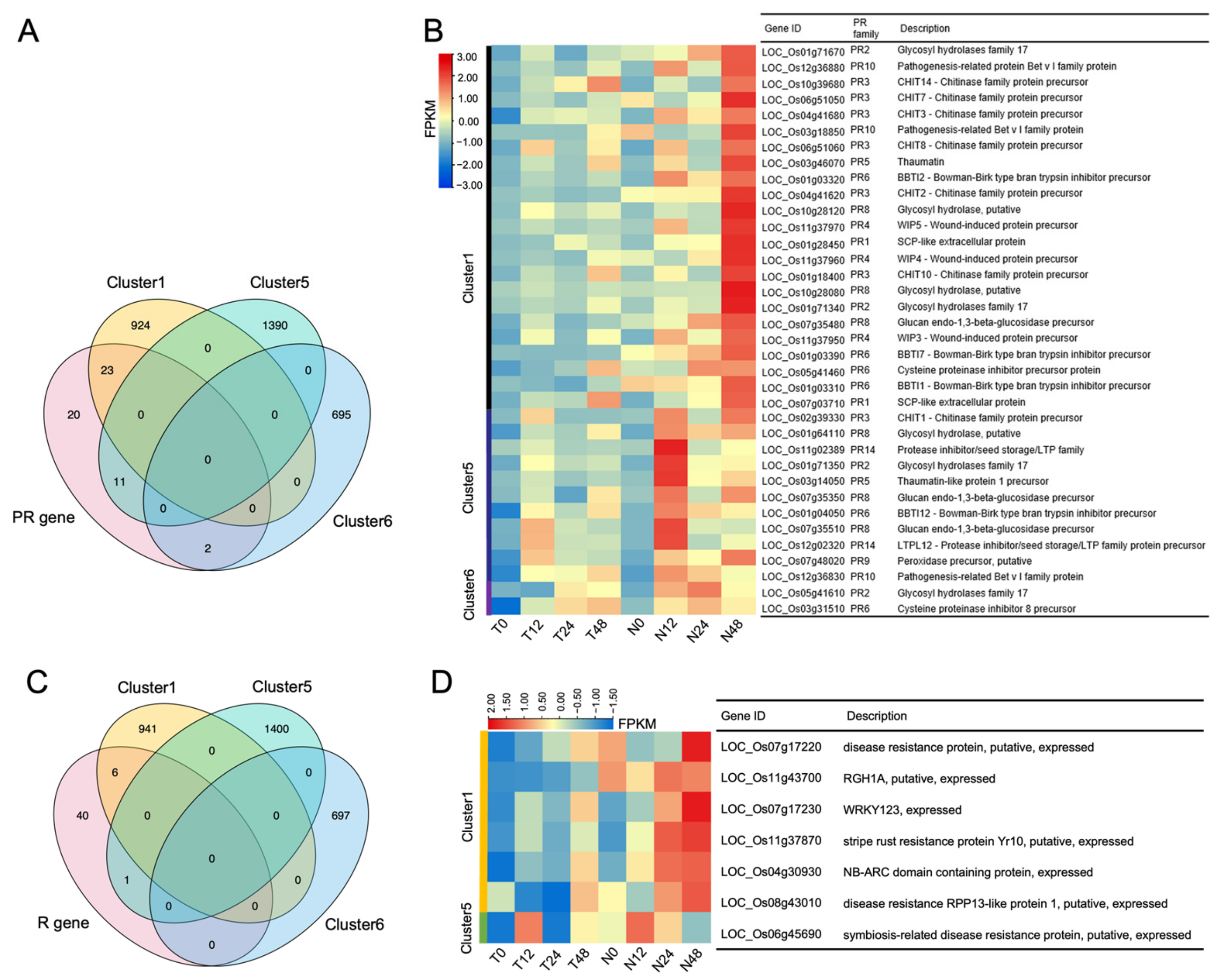
2.3. OsNPR1-Enhanced Expression of PR Genes and R Genes after PXO99A Infection
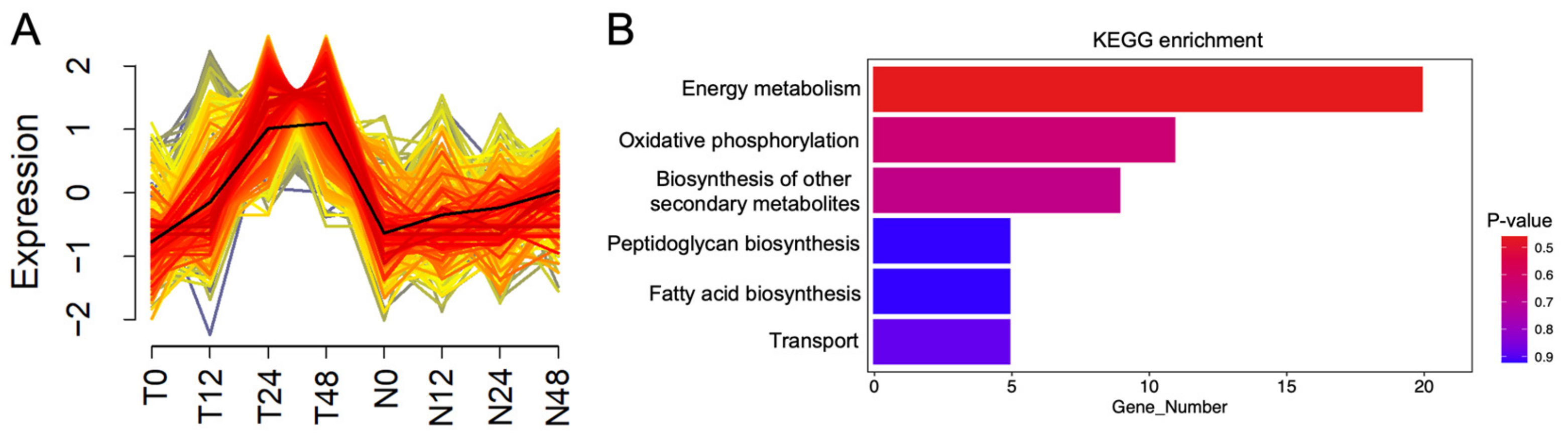
2.4. Expression of Genes Involved in Cellular Metabolisms of PXO99A Was Downregulated by OsNPR1
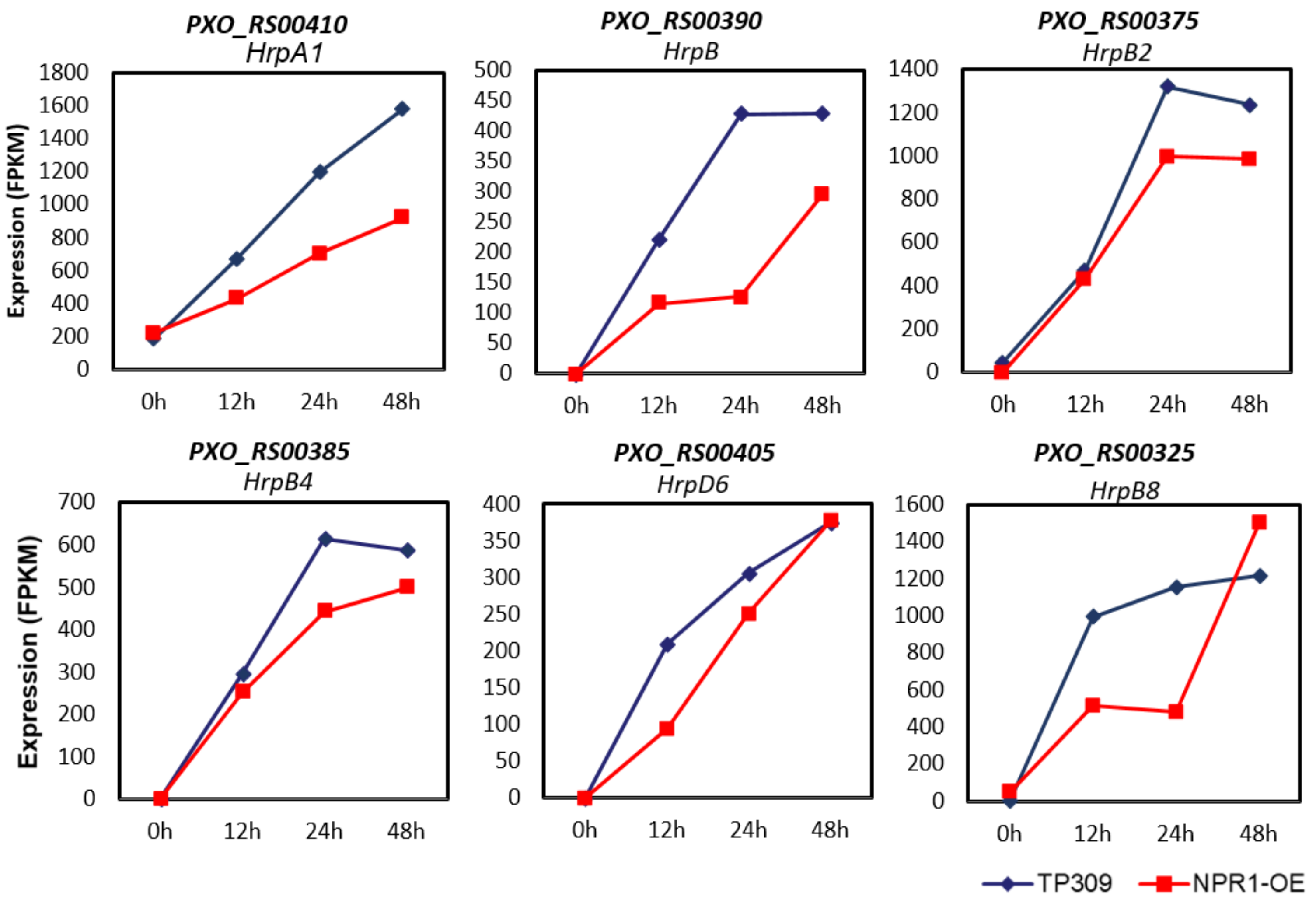
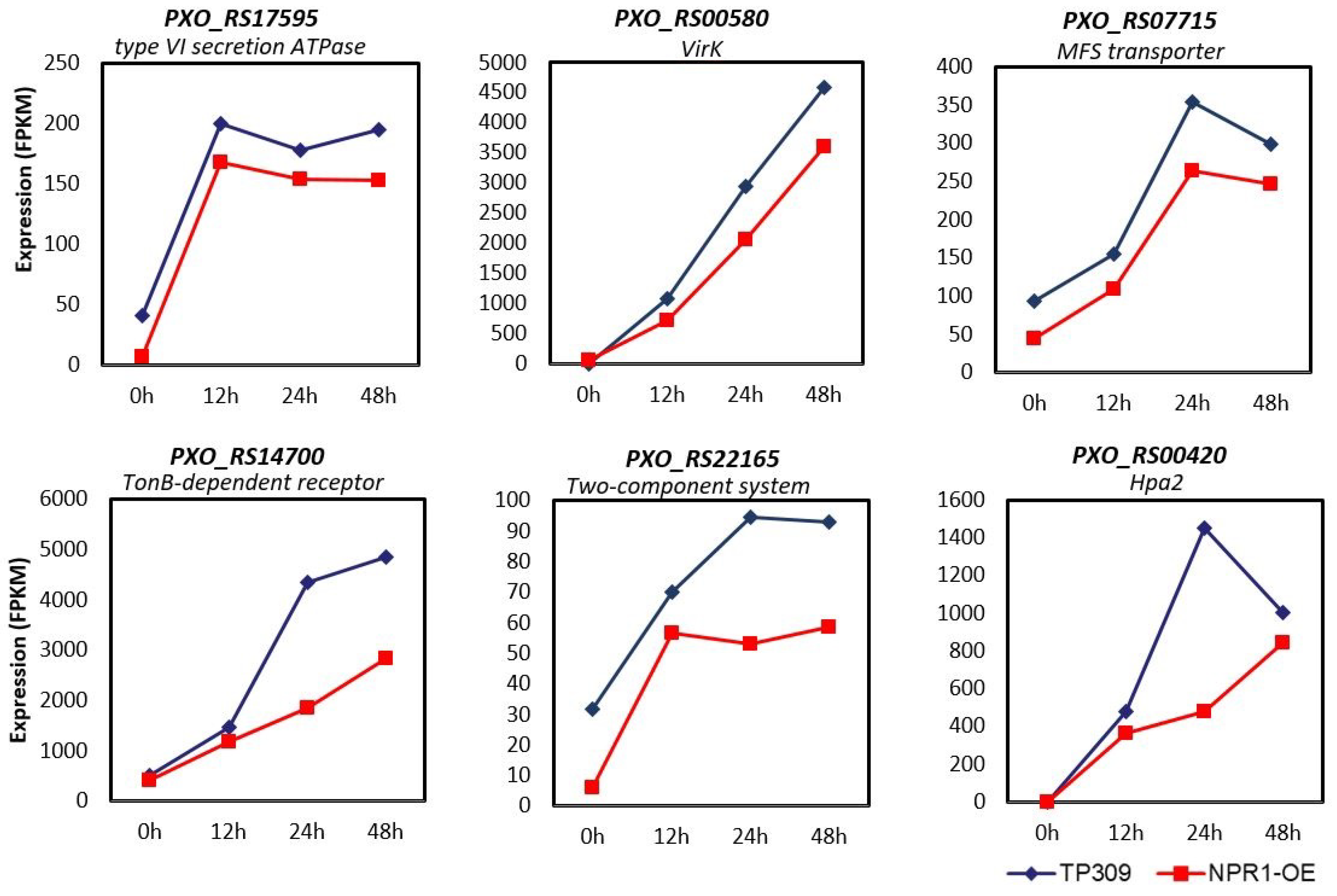
2.5. Expression of PXO99A Virulence Genes Was Repressed by OsNPR1
3. Discussion
3.1. Dual RNAseq Is an Excellent Strategy for Simultaneous Analyses of the Gene Expression of Interactive Organisms
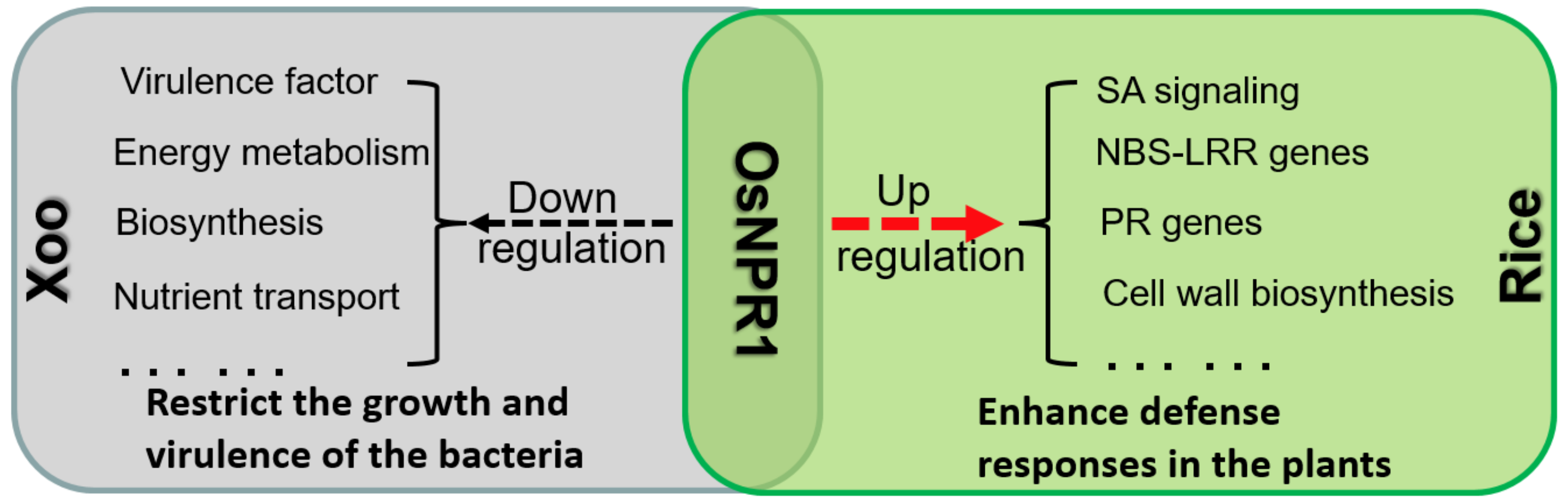
3.2. OsNPR1 Enhances Rice Resistance to POX99A by Upregulating the Expression of Rice Genes Involved in Multi-Layer Defense Responses
3.3. OsNPR1 Enhances Rice Resistance to Xoo by Repressing the Expression of Bacteria Virulence Genes
4. Materials and Methods
4.1. Bacterial Strains, Culture, and Growth Conditions
4.2. Rice Materials and Growth Conditions
4.3. Pathogen Inoculation
4.4. Quantitative Real-Time PCR
4.5. Protein Detection by Western Blot
4.6. RNAseq and Data Process
4.7. Gene Clustering
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, Y.; Kim, S.G.; Wu, J.; Huh, H.H.; Lee, S.J.; Rakwal, R.; Agrawal, G.K.; Park, Z.Y.; Young Kang, K.; Kim, S.T. Secretome analysis of the rice bacterium Xanthomonas oryzae (Xoo) using in vitro and in planta systems. Proteomics 2013, 13, 1901–1912. [Google Scholar] [CrossRef]
- Nino-Liu, D.O.; Ronald, P.C.; Bogdanove, A.J. Xanthomonas oryzae pathovars: Model pathogens of a model crop. Mol. Plant Pathol. 2006, 7, 303–324. [Google Scholar] [CrossRef]
- Gu, K.; Yang, B.; Tian, D.; Wu, L.; Wang, D.; Sreekala, C.; Yang, F.; Chu, Z.; Wang, G.L.; White, F.F.; et al. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature 2005, 435, 1122–1125. [Google Scholar] [CrossRef] [PubMed]
- Cheong, H.; Kim, C.Y.; Jeon, J.S.; Lee, B.M.; Sun Moon, J.; Hwang, I. Xanthomonas oryzae pv. oryzae type III effector XopN targets OsVOZ2 and a putative thiamine synthase as a virulence factor in rice. PLoS ONE 2013, 8, e73346. [Google Scholar] [CrossRef]
- Galan, J.E.; Collmer, A. Type III secretion machines: Bacterial devices for protein delivery into host cells. Science 1999, 284, 1322–1328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notti, R.Q.; Stebbins, C.E. The Structure and Function of Type III Secretion Systems. Microbiol. Spectr. 2016, 4, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; MaGbanua, M.M.; White, F.F. Identification of two novel hrp-associated genes in the hrp gene cluster of Xanthomonas oryzae pv. oryzae. J. Bacteriol. 2000, 182, 1844–1853. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Ronald, P. Molecular determinants of disease and resistance in interactions of Xanthomonas oryzae pv. oryzae and rice. Microbes Infect. 2002, 4, 1361–1367. [Google Scholar] [CrossRef] [Green Version]
- Keen, N.T.; Boyd, C.; Henrissat, B. Cloning and characterization of a xylanase gene from corn strains of Erwinia chrysanthemi. Mol. Plant Microbe Interact. 1996, 9, 651–657. [Google Scholar] [CrossRef]
- Kaku, H.; Nishizawa, Y.; Ishii-Minami, N.; Akimoto-Tomiyama, C.; Dohmae, N.; Takio, K.; Minami, E.; Shibuya, N. Plant cells recognize chitin fragments for defense signaling through a plasma membrane receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 11086–11091. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [Green Version]
- Kaffarnik, F.A.; Jones, A.M.; Rathjen, J.P.; Peck, S.C. Effector proteins of the bacterial pathogen Pseudomonas syringae alter the extracellular proteome of the host plant, Arabidopsis thaliana. Mol. Cell Proteom. 2009, 8, 145–156. [Google Scholar] [CrossRef] [Green Version]
- Hulbert, S.H.; Webb, C.A.; Smith, S.M.; Sun, Q. Resistance gene complexes: Evolution and utilization. Annu. Rev. Phytopathol. 2001, 39, 285–312. [Google Scholar] [CrossRef]
- Brar, D.S.; Khush, G.S. Alien introgression in rice. Plant Mol. Biol. 1997, 35, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.S.; Rasabandith, S.; Angeles, E.R.; Khush, G.S. Inheritance of resistance to bacterial blight in 21 cultivars of rice. Phytopathology 2003, 93, 147–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, F.F.; Yang, B. Host and pathogen factors controlling the rice-Xanthomonas oryzae interaction. Plant Physiol. 2009, 150, 1677–1686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Y.; Yang, J.; Li, X.; Zhang, Y. Salicylic Acid: Biosynthesis and Signaling. Annu. Rev. Plant Biol. 2021, 72, 761–791. [Google Scholar] [CrossRef]
- Gaffney, T.; Friedrich, L.; Vernooij, B.; Negrotto, D.; Nye, G.; Uknes, S.; Ward, E.; Kessmann, H.; Ryals, J. Requirement of salicylic Acid for the induction of systemic acquired resistance. Science 1993, 261, 754–756. [Google Scholar] [CrossRef]
- Mou, Z.; Fan, W.; Dong, X. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef] [Green Version]
- Després, C.; Chubak, C.; Rochon, A.; Clark, R.; Bethune, T.; Desveaux, D.; Fobert, P.R. The Arabidopsis NPR1 disease resistance protein is a novel cofactor that confers redox regulation of DNA binding activity to the basic domain/leucine zipper transcription factor TGA1. Plant Cell 2003, 15, 2181–2191. [Google Scholar] [CrossRef] [Green Version]
- Fu, Z.Q.; Dong, X. Systemic acquired resistance: Turning local infection into global defense. Annu. Rev. Plant Biol. 2013, 64, 839–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Sun, T.; Sun, Y.; Zhang, Y.; Radojicic, A.; Ding, Y.; Tian, H.; Huang, X.; Lan, J.; Chen, S.; et al. Diverse Roles of the Salicylic Acid Receptors NPR1 and NPR3/NPR4 in Plant Immunity. Plant Cell 2020, 32, 4002–4016. [Google Scholar] [CrossRef]
- Chern, M.; Fitzgerald, H.A.; Canlas, P.E.; Navarre, D.A.; Ronald, P.C. Overexpression of a rice NPR1 homolog leads to constitutive activation of defense response and hypersensitivity to light. Mol. Plant Microbe Interact. 2005, 18, 511–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugano, S.; Jiang, C.J.; Miyazawa, S.; Masumoto, C.; Yazawa, K.; Hayashi, N.; Shimono, M.; Nakayama, A.; Miyao, M.; Takatsuji, H. Role of OsNPR1 in rice defense program as revealed by genome-wide expression analysis. Plant Mol. Biol. 2010, 74, 549–562. [Google Scholar] [CrossRef]
- Astua-Monge, G.; Freitas-Astua, J.; Bacocina, G.; Roncoletta, J.; Carvalho, S.A.; Machado, M.A. Expression profiling of virulence and pathogenicity genes of Xanthomonas axonopodis pv. citri. J. Bacteriol. 2005, 187, 1201–1205. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Figueiredo, F.; Jones, J.; Wang, N. HrpG and HrpX play global roles in coordinating different virulence traits of Xanthomonas axonopodis pv. citri. Mol. Plant Microbe Interact. 2011, 24, 649–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Cho, Y.J.; Song, E.S.; Lee, S.H.; Kim, J.G.; Kang, L.W. Time-resolved pathogenic gene expression analysis of the plant pathogen Xanthomonas oryzae pv. oryzae. BMC Genom. 2016, 17, 345. [Google Scholar] [CrossRef] [Green Version]
- Hou, M.; Xu, W.; Bai, H.; Liu, Y.; Li, L.; Liu, L.; Liu, B.; Liu, G. Characteristic expression of rice pathogenesis-related proteins in rice leaves during interactions with Xanthomonas oryzae pv. oryzae. Plant Cell Rep. 2012, 31, 895–904. [Google Scholar] [CrossRef]
- Li, X.; Yang, D.L.; Sun, L.; Li, Q.; Mao, B.; He, Z. The Systemic Acquired Resistance Regulator OsNPR1 Attenuates Growth by Repressing Auxin Signaling through Promoting IAA-Amido Synthase Expression. Plant Physiol. 2016, 172, 546–558. [Google Scholar] [CrossRef] [Green Version]
- Shimono, M.; Koga, H.; Akagi, A.; Hayashi, N.; Goto, S.; Sawada, M.; Kurihara, T.; Matsushita, A.; Sugano, S.; Jiang, C.J.; et al. Rice WRKY45 plays important roles in fungal and bacterial disease resistance. Mol. Plant Pathol. 2012, 13, 83–94. [Google Scholar] [CrossRef]
- Hwang, S.H.; Kwon, S.I.; Jang, J.Y.; Fang, I.L.; Lee, H.; Choi, C.; Park, S.; Ahn, I.; Bae, S.C.; Hwang, D.J. OsWRKY51, a rice transcription factor, functions as a positive regulator in defense response against Xanthomonas oryzae pv. oryzae. Plant Cell Rep. 2016, 35, 1975–1985. [Google Scholar] [CrossRef]
- Dou, S.G.; Guan, M.; Li, L.; Liu, G. Pathogenesis-related genes in rice. Chin. Sci. Bull. 2014, 59, 245. (In Chinese) [Google Scholar] [CrossRef]
- Zhang, X.; Yang, S.; Wang, J.; Jia, Y.; Huang, J.; Tan, S.; Zhong, Y.; Wang, L.; Gu, L.; Chen, J.Q.; et al. A genome-wide survey reveals abundant rice blast R genes in resistant cultivars. Plant J. 2015, 84, 20–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genin, S.; Brito, B.; Denny, T.P.; Boucher, C. Control of the Ralstonia solanacearum Type III secretion system (Hrp) genes by the global virulence regulator PhcA. FEBS Lett. 2005, 579, 2077–2081. [Google Scholar] [CrossRef] [Green Version]
- Furutani, A.; Tsuge, S.; Ohnishi, K.; Hikichi, Y.; Oku, T.; Tsuno, K.; Inoue, Y.; Ochiai, H.; Kaku, H.; Kubo, Y. Evidence for HrpXo-dependent expression of type II secretory proteins in Xanthomonas oryzae pv. oryzae. J. Bacteriol. 2004, 186, 1374–1380. [Google Scholar] [CrossRef] [Green Version]
- Schulte, R.; Bonas, U. A Xanthomonas Pathogenicity Locus Is Induced by Sucrose and Sulfur-Containing Amino Acids. Plant Cell 1992, 4, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Diehl, B.G.; Brown, N.R. Lignin cross-links with cysteine- and tyrosine-containing peptides under biomimetic conditions. J. Agric. Food Chem. 2014, 62, 10312–10319. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Jeon, H.S.; Kim, S.H.; Chung, J.H.; Roppolo, D.; Lee, H.J.; Cho, H.J.; Tobimatsu, Y.; Ralph, J.; Park, O.K. Lignin-based barrier restricts pathogens to the infection site and confers resistance in plants. EMBO J. 2019, 38, e101948. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.; Wang, L. The phenylpropanoid pathway and plant defence-a genomics perspective. Mol. Plant Pathol. 2002, 3, 371–390. [Google Scholar] [CrossRef]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid Pathway Engineering: An Emerging Approach towards Plant Defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef] [Green Version]
- Tonnessen, B.W.; Manosalva, P.; Lang, J.M.; Baraoidan, M.; Bordeos, A.; Mauleon, R.; Oard, J.; Hulbert, S.; Leung, H.; Leach, J.E. Rice phenylalanine ammonia-lyase gene OsPAL4 is associated with broad spectrum disease resistance. Plant Mol. Biol. 2015, 87, 273–286. [Google Scholar] [CrossRef]
- Liu, L.; Shang-Guan, K.; Zhang, B.; Liu, X.; Yan, M.; Zhang, L.; Shi, Y.; Zhang, M.; Qian, Q.; Li, J.; et al. Brittle Culm1, a COBRA-like protein, functions in cellulose assembly through binding cellulose microfibrils. PLoS Genet. 2013, 9, e1003704. [Google Scholar] [CrossRef] [PubMed]
- Eulgem, T.; Somssich, I.E. Networks of WRKY transcription factors in defense signaling. Curr. Opin. Plant Biol. 2007, 10, 366–371. [Google Scholar] [CrossRef] [Green Version]
- Vlot, A.C.; Dempsey, D.A.; Klessig, D.F. Salicylic Acid, a multifaceted hormone to combat disease. Annu. Rev. Phytopathol. 2009, 47, 177–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndamukong, I.; Abdallat, A.A.; Thurow, C.; Fode, B.; Zander, M.; Weigel, R.; Gatz, C. SA-inducible Arabidopsis glutaredoxin interacts with TGA factors and suppresses JA-responsive PDF1.2 transcription. Plant J. 2007, 50, 128–139. [Google Scholar] [CrossRef] [PubMed]
- Pajerowska-Mukhtar, K.M.; Emerine, D.K.; Mukhtar, M.S. Tell me more: Roles of NPRs in plant immunity. Trends. Plant Sci. 2013, 18, 402–411. [Google Scholar] [CrossRef]
- Wasternack, C.; Song, S. Jasmonates: Biosynthesis, metabolism, and signaling by proteins activating and repressing transcription. J. Exp. Bot. 2017, 68, 1303–1321. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.C.; Rep, M.; Pieterse, C.M. Significance of inducible defense-related proteins in infected plants. Annu. Rev. Phytopathol. 2006, 44, 135–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oku, T.; Tanaka, K.; Iwamoto, M.; Inoue, Y.; Ochiai, H.; Kaku, H.; Tsuge, S.; Tsuno, K. Structural conservation of the hrp gene cluster in Xanthomons oryzae pv. oryzae. J. Gen. Plant Pathol. 2004, 70, 159–167. [Google Scholar] [CrossRef]
- Costa, T.R.; Felisberto-Rodrigues, C.; Meir, A.; Prevost, M.S.; Redzej, A.; Trokter, M.; Waksman, G. Secretion systems in Gram-negative bacteria: Structural and mechanistic insights. Nat. Rev. Microbiol. 2015, 13, 343–359. [Google Scholar] [CrossRef]
- Chen, T.; Chen, Z.; Li, Y.; Zeng, B.; Zhang, Z. A Novel Major Facilitator Superfamily Transporter Gene Aokap4 near the Kojic Acid Gene Cluster Is Involved in Growth and Kojic Acid Production in Aspergillus oryzae. J. Fungi. 2022, 8, 885. [Google Scholar] [CrossRef] [PubMed]
- Song, W.Y.; Wang, G.L.; Chen, L.L.; Kim, H.S.; Pi, L.Y.; Holsten, T.; Gardner, J.; Wang, B.; Zhai, W.X.; Zhu, L.H.; et al. A receptor kinase-like protein encoded by the rice disease resistance gene, Xa21. Science 1995, 270, 1804–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, Y.; Liu, H.; Li, X.; Xiao, J.; Wang, S. Rice OsPAD4 functions differently from Arabidopsis AtPAD4 in host-pathogen interactions. Plant J. 2014, 78, 619–631. [Google Scholar] [CrossRef]
- Salzberg, S.L.; Sommer, D.D.; Schatz, M.C.; Phillippy, A.M.; Rabinowicz, P.D.; Tsuge, S.; Furutani, A.; Ochiai, H.; Delcher, A.L.; Kelley, D.; et al. Genome sequence and rapid evolution of the rice pathogen Xanthomonas oryzae pv. oryzae PXO99A. BMC Genom. 2008, 9, 204. [Google Scholar] [CrossRef]
- Kumar, L.; Futschik, M.E. Mfuzz: A software package for soft clustering of microarray data. Bioinformation 2007, 2, 5–7. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dai, X.; Wang, Y.; Yu, K.; Zhao, Y.; Xiong, L.; Wang, R.; Li, S. OsNPR1 Enhances Rice Resistance to Xanthomonas oryzae pv. oryzae by Upregulating Rice Defense Genes and Repressing Bacteria Virulence Genes. Int. J. Mol. Sci. 2023, 24, 8687. https://doi.org/10.3390/ijms24108687
Dai X, Wang Y, Yu K, Zhao Y, Xiong L, Wang R, Li S. OsNPR1 Enhances Rice Resistance to Xanthomonas oryzae pv. oryzae by Upregulating Rice Defense Genes and Repressing Bacteria Virulence Genes. International Journal of Molecular Sciences. 2023; 24(10):8687. https://doi.org/10.3390/ijms24108687
Chicago/Turabian StyleDai, Xing, Yankai Wang, Kaili Yu, Yonghui Zhao, Langyu Xiong, Ruozhong Wang, and Shengben Li. 2023. "OsNPR1 Enhances Rice Resistance to Xanthomonas oryzae pv. oryzae by Upregulating Rice Defense Genes and Repressing Bacteria Virulence Genes" International Journal of Molecular Sciences 24, no. 10: 8687. https://doi.org/10.3390/ijms24108687