
The Establishment of a Genetic Transformation System and the Acquisition of Transgenic Plants of Oriental Hybrid Lily (Lilium L.)
Abstract
:1. Introduction
2. Results
2.1. Effects of Different Plant Growth Regulator on Callus Induction of Two Kinds of Lily Stems
2.2. Effects of Different Plant Growth Regulators on Callus Induction of Two Kinds of Lily Filaments
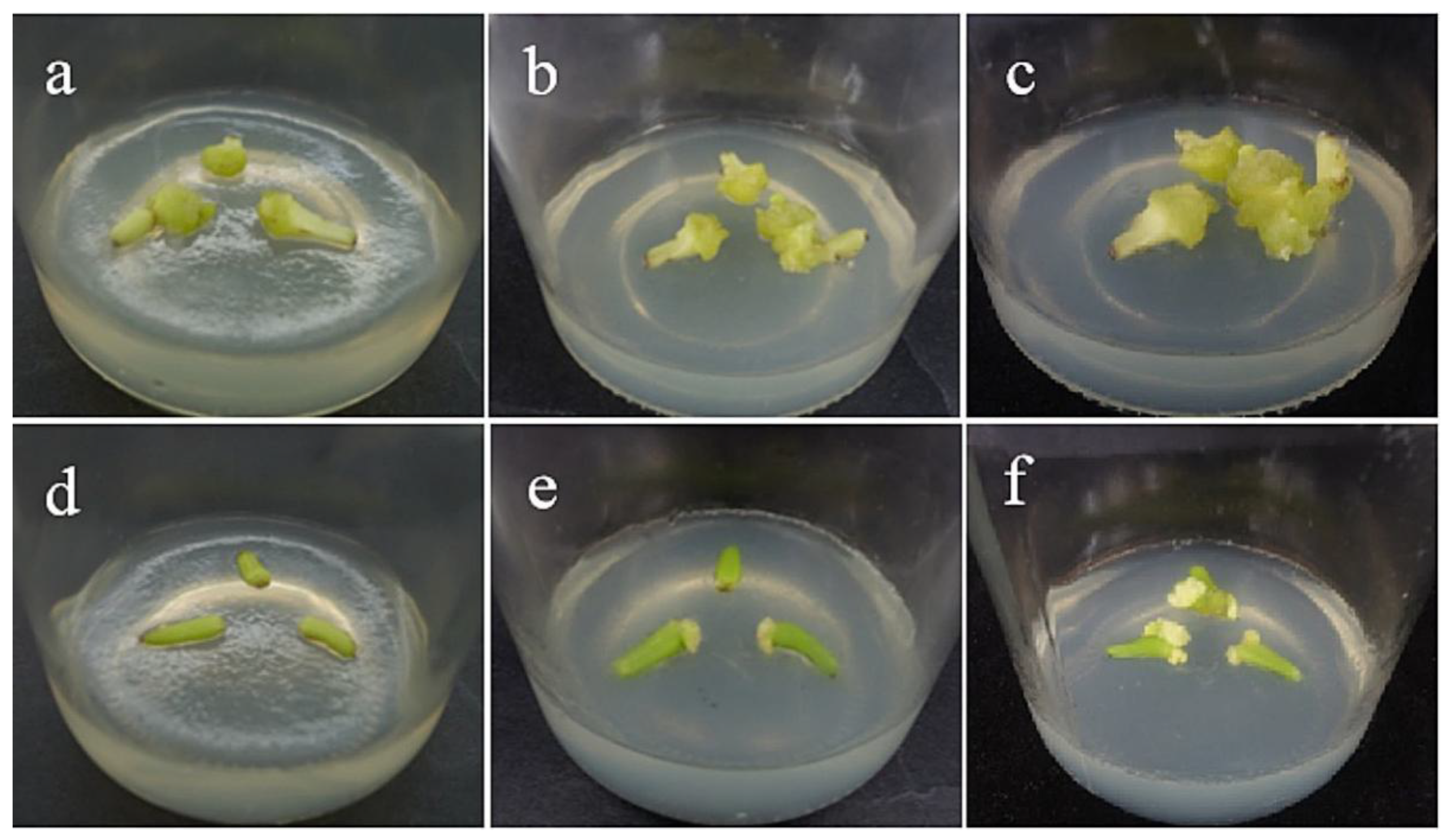
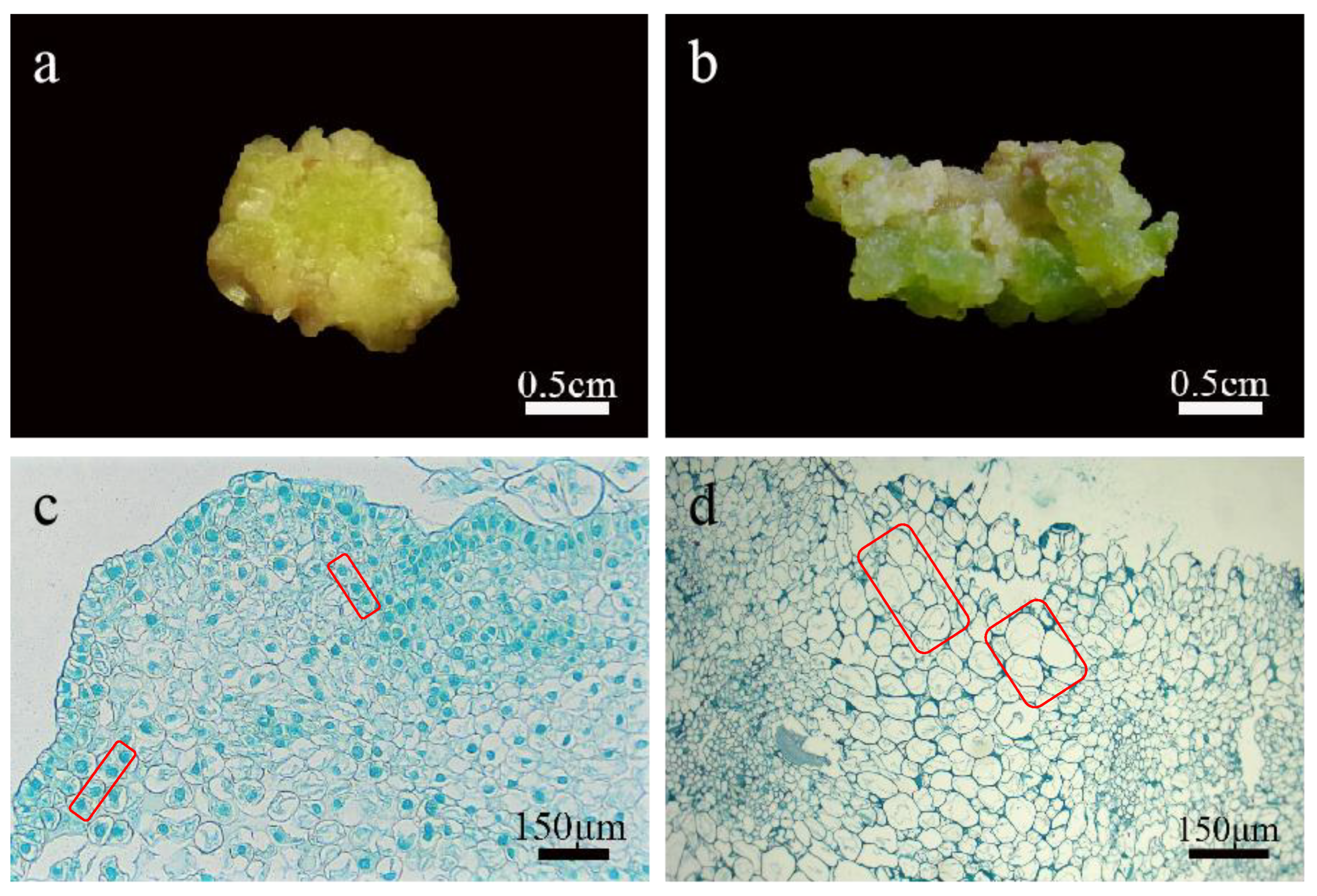
2.3. Morphological and Histological Observation of Callus
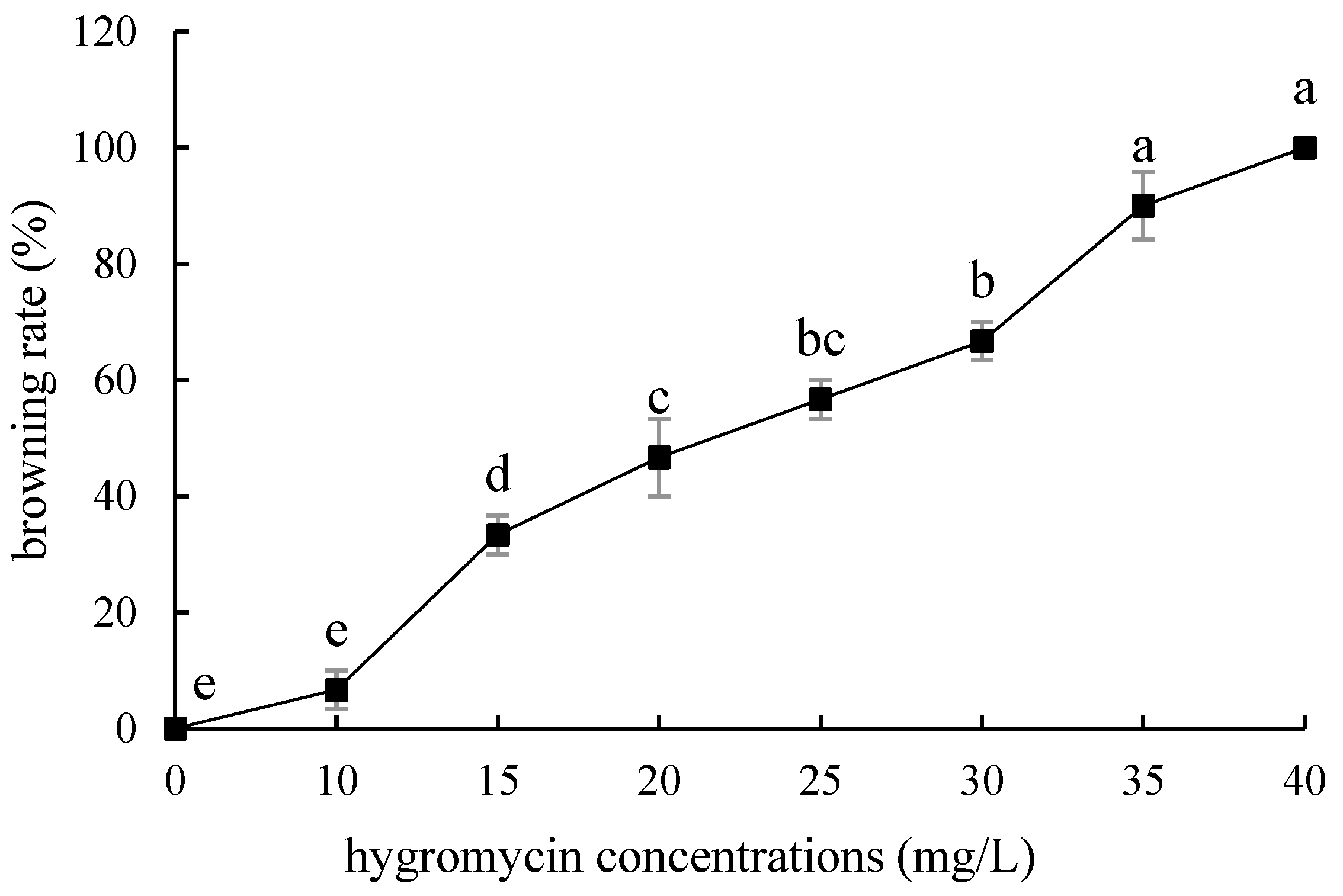
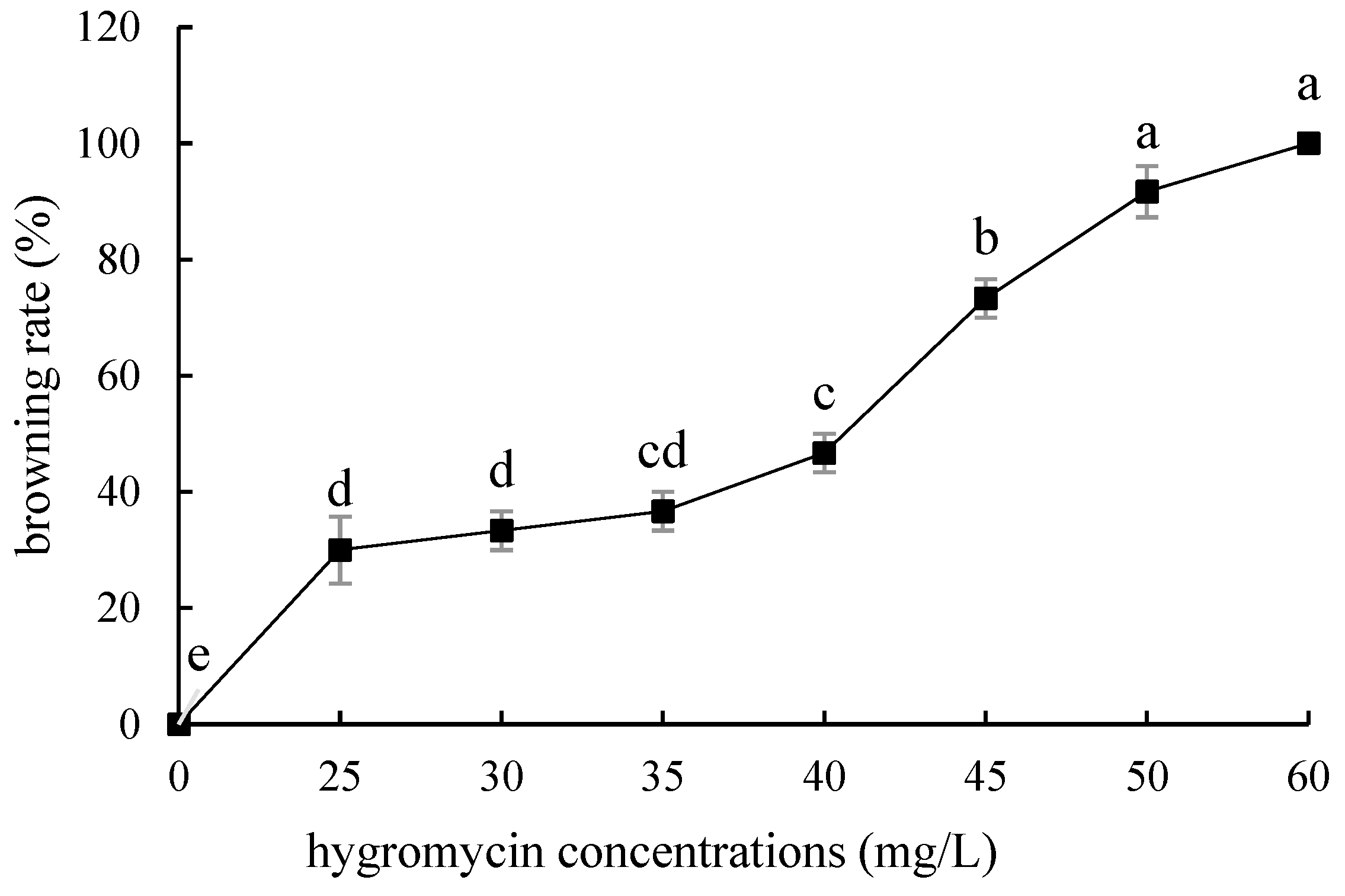
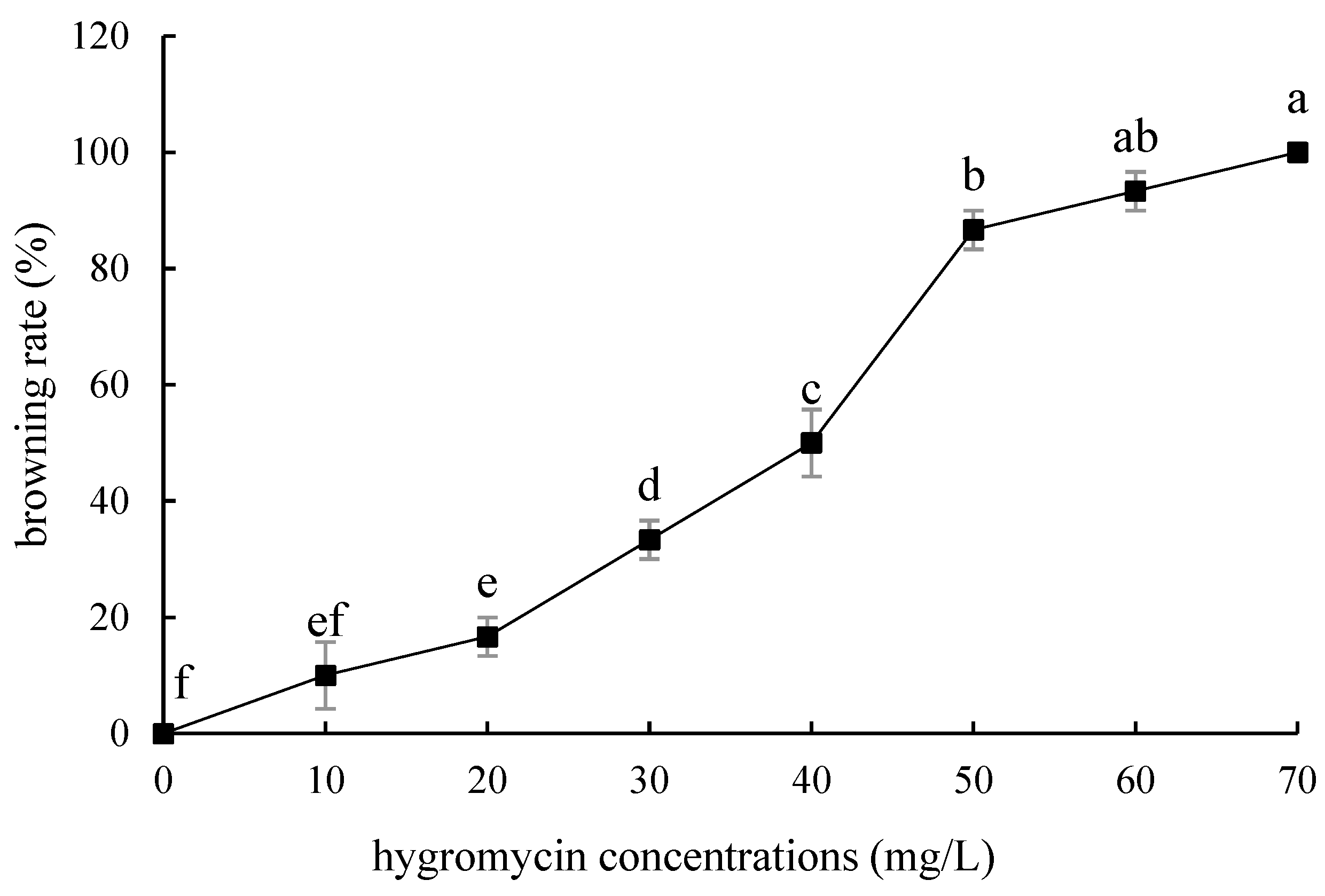
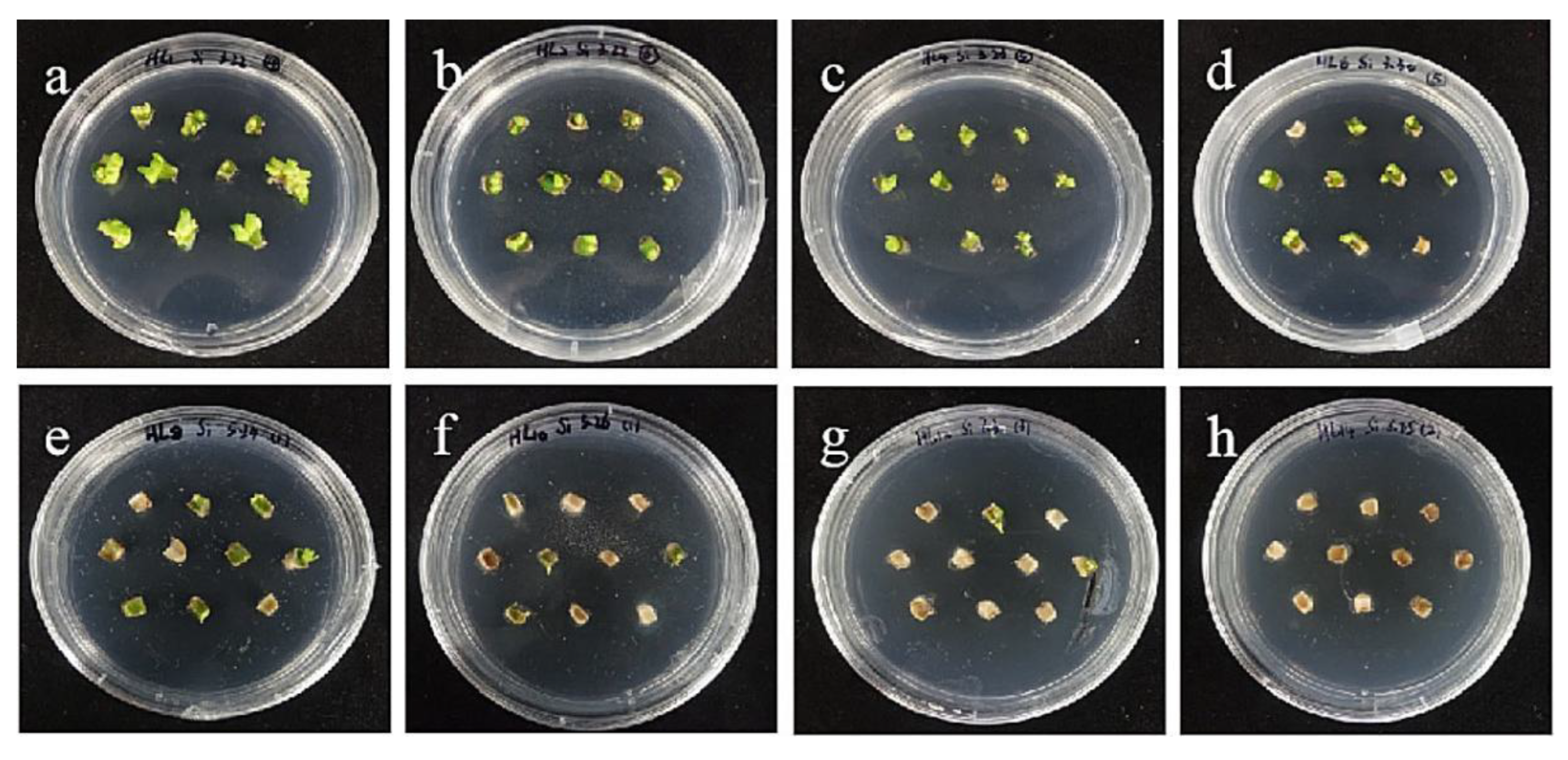
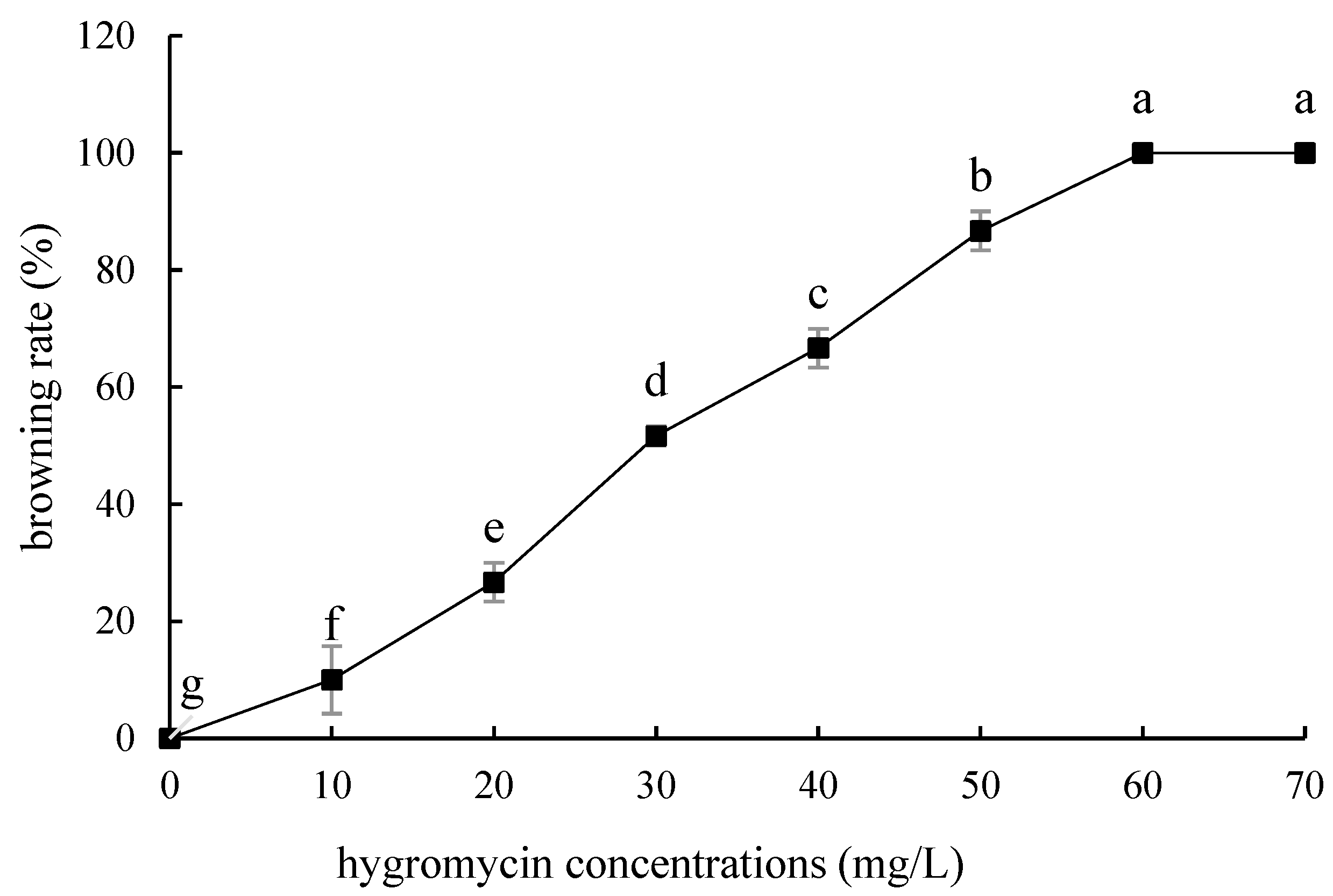
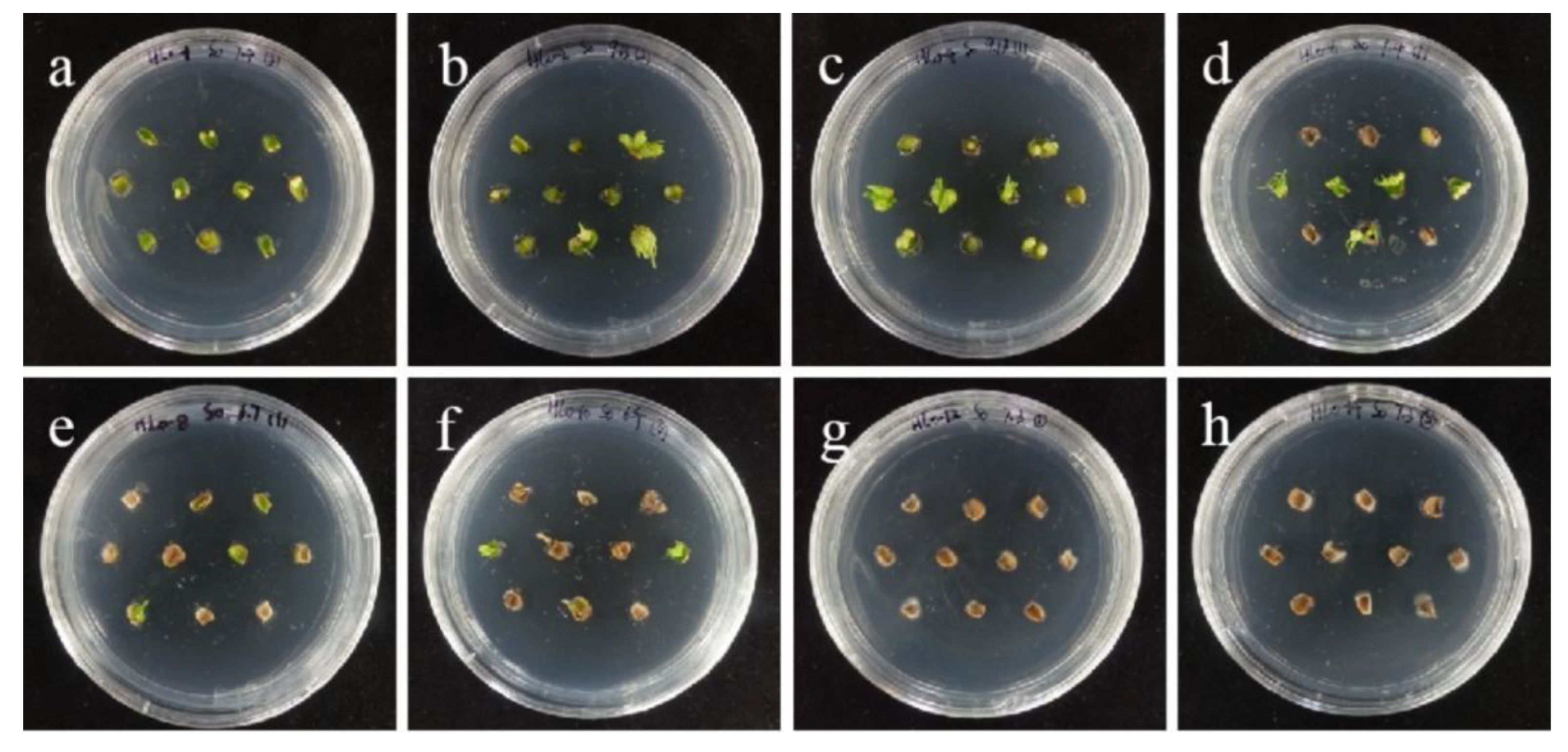
2.4. Hygromycin Concentration Screening
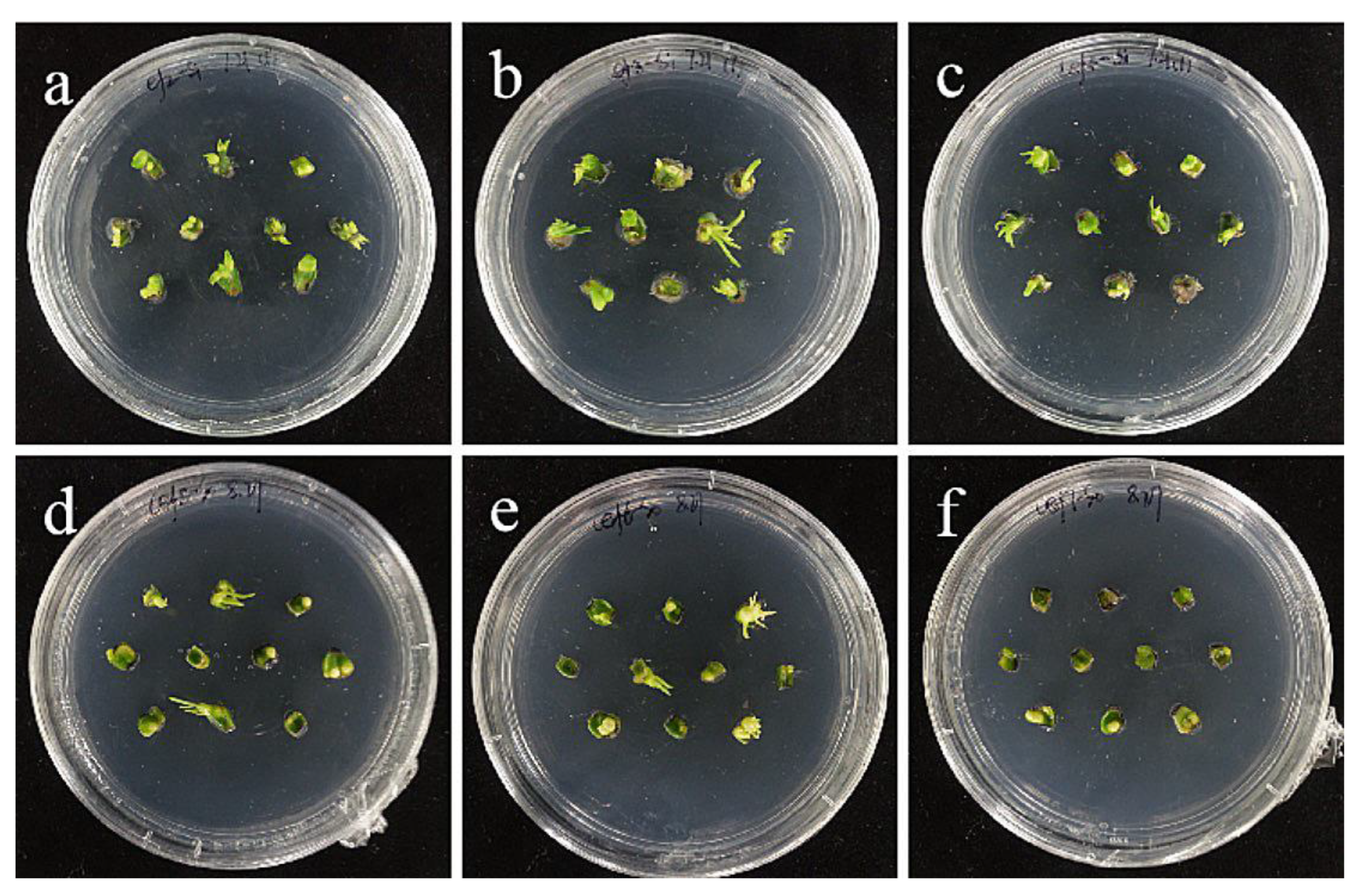
2.5. Cephalosporin Concentration Screening
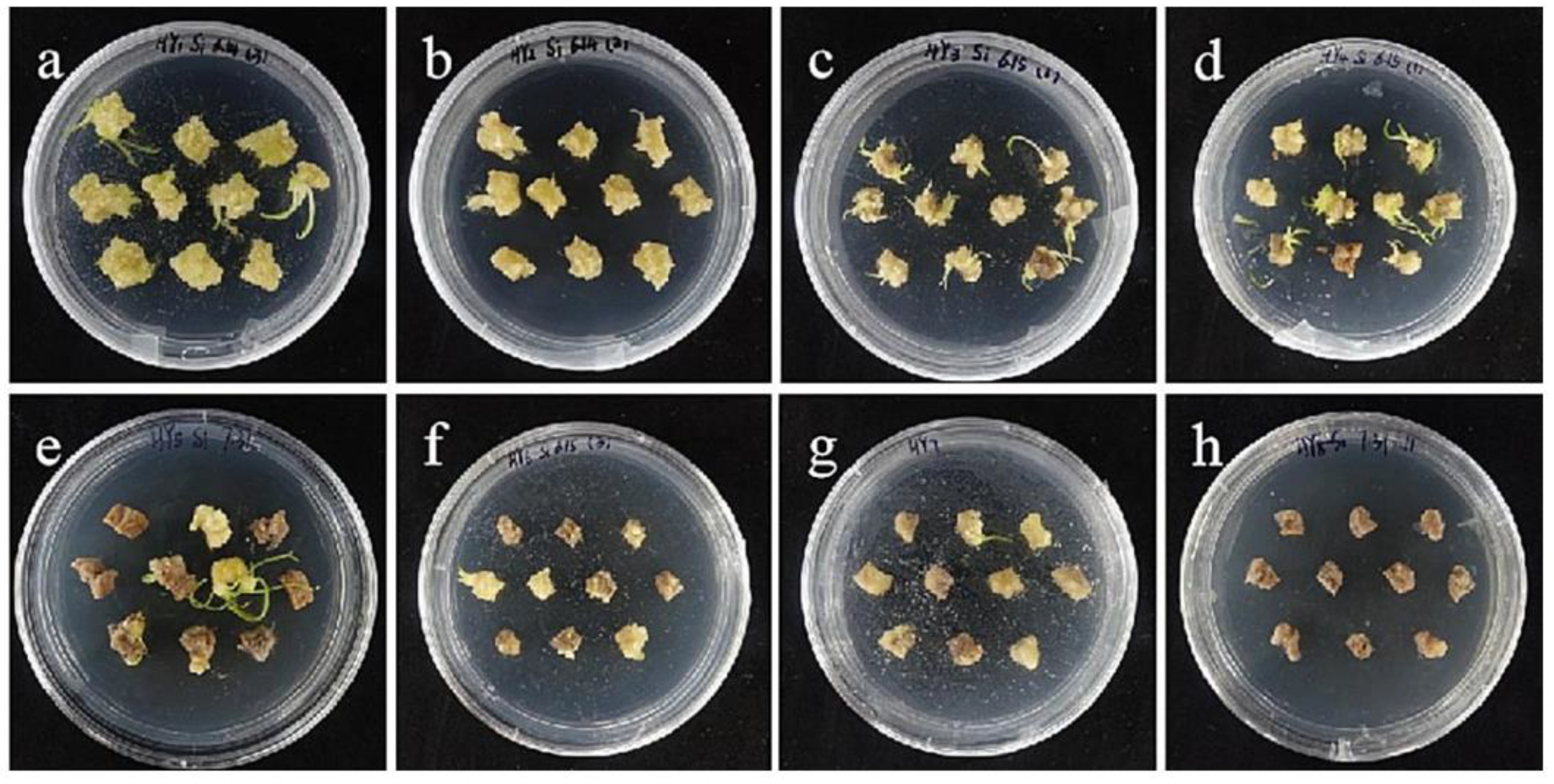
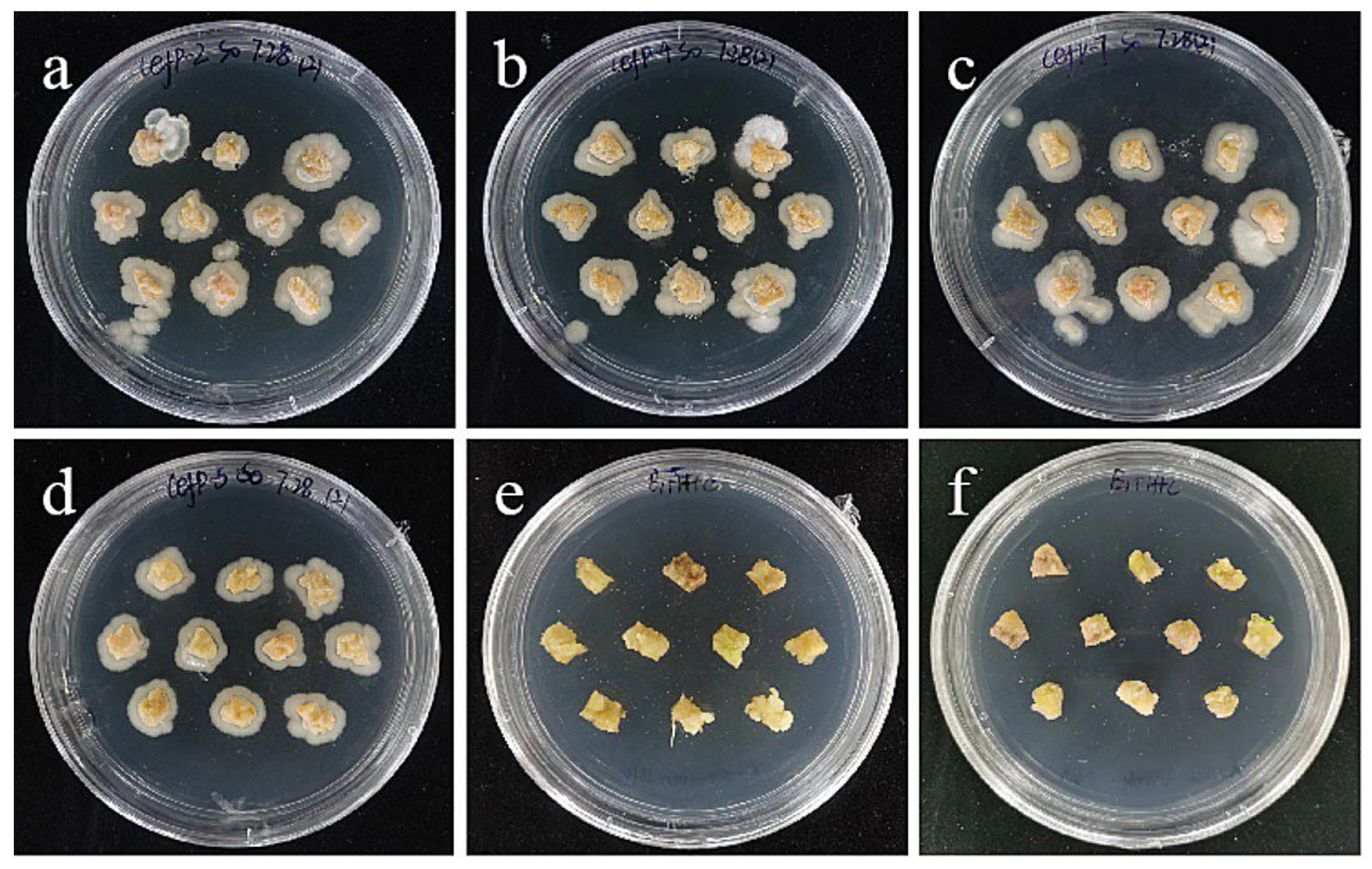
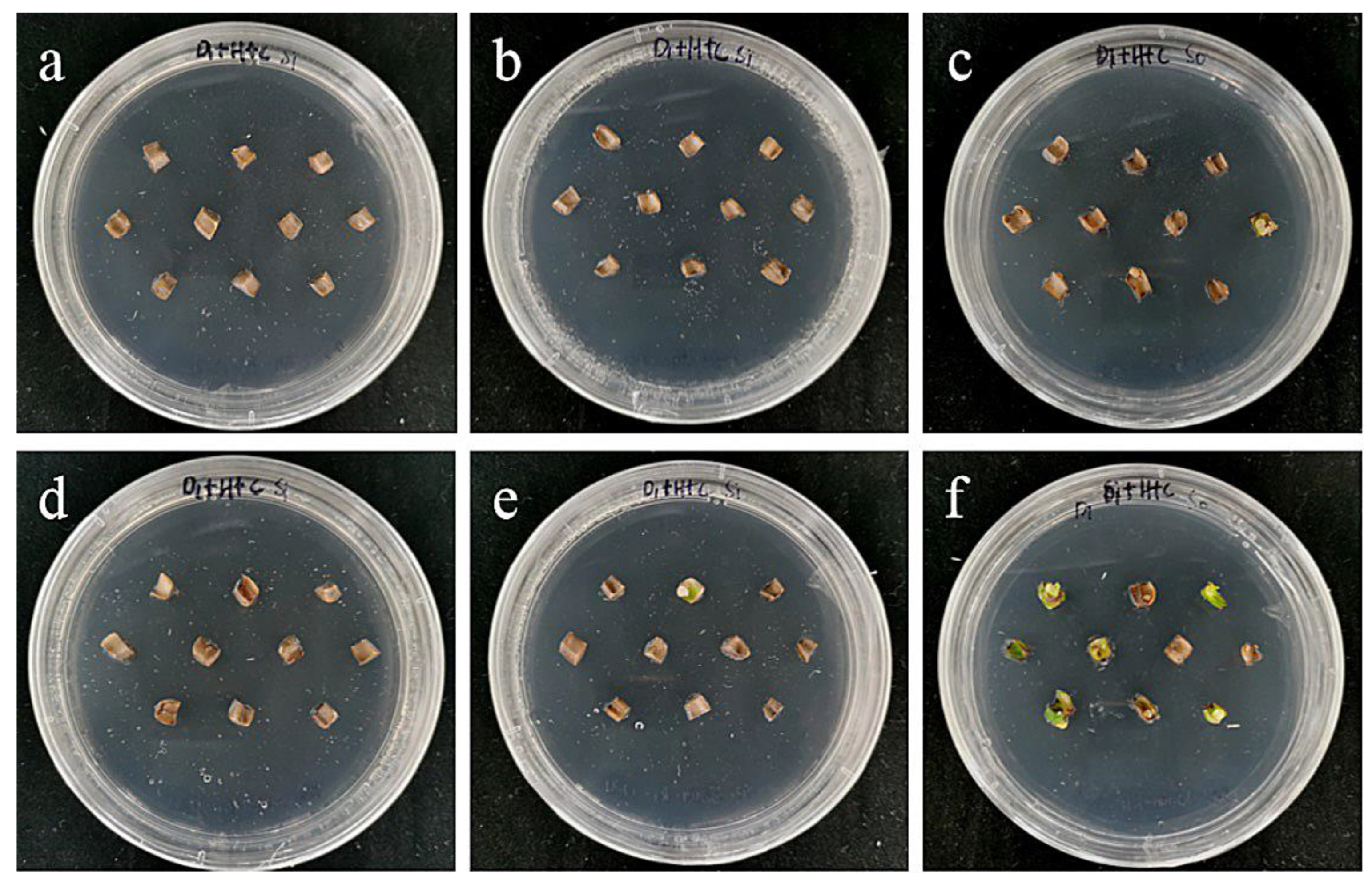
2.6. Effects of Bacterial Concentration and Infection Time on Embryogenic Callus
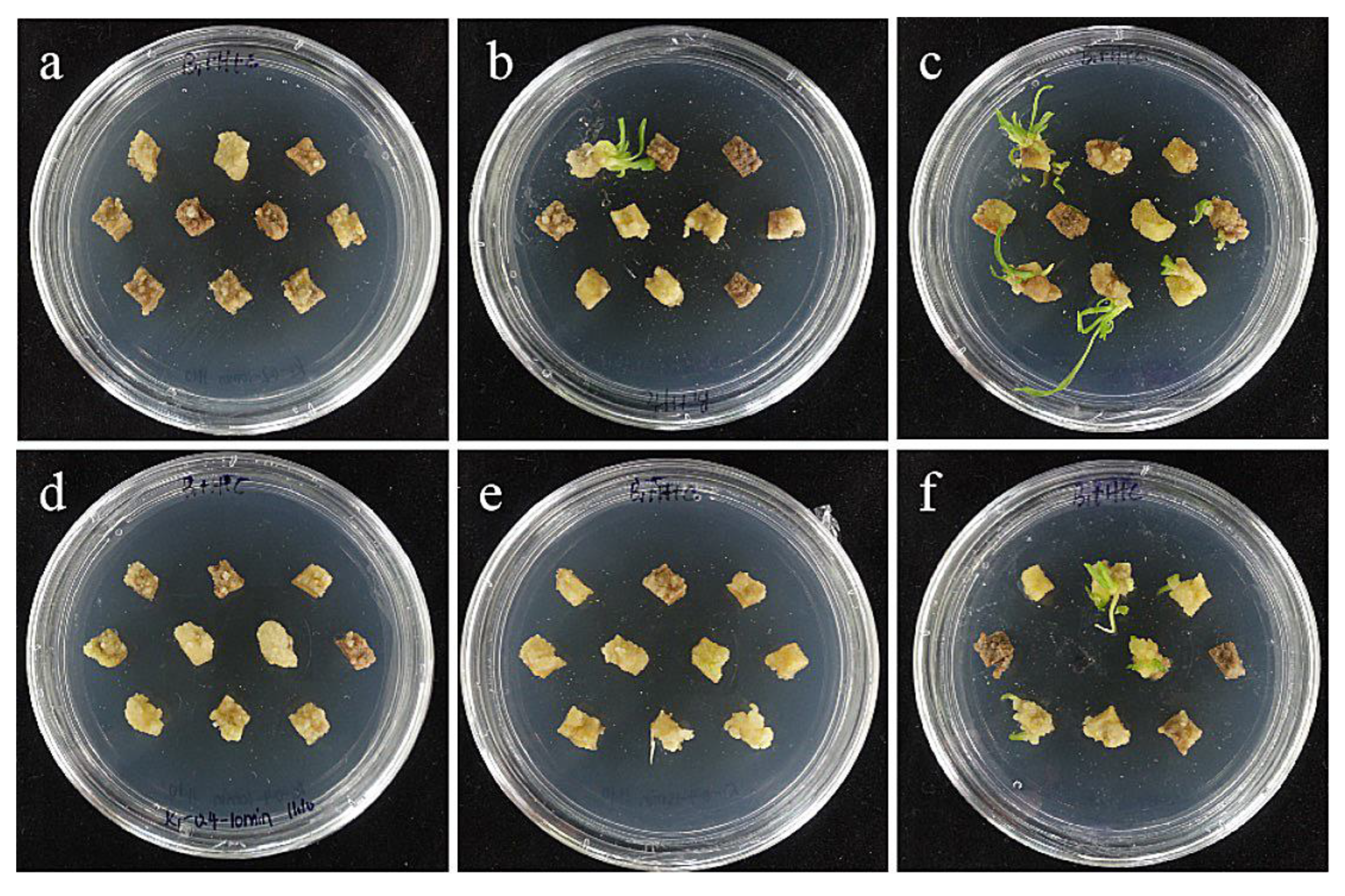
2.7. Effects of Bacterial Concentration and Infection Time on Small Scales of Sterile Seedlings
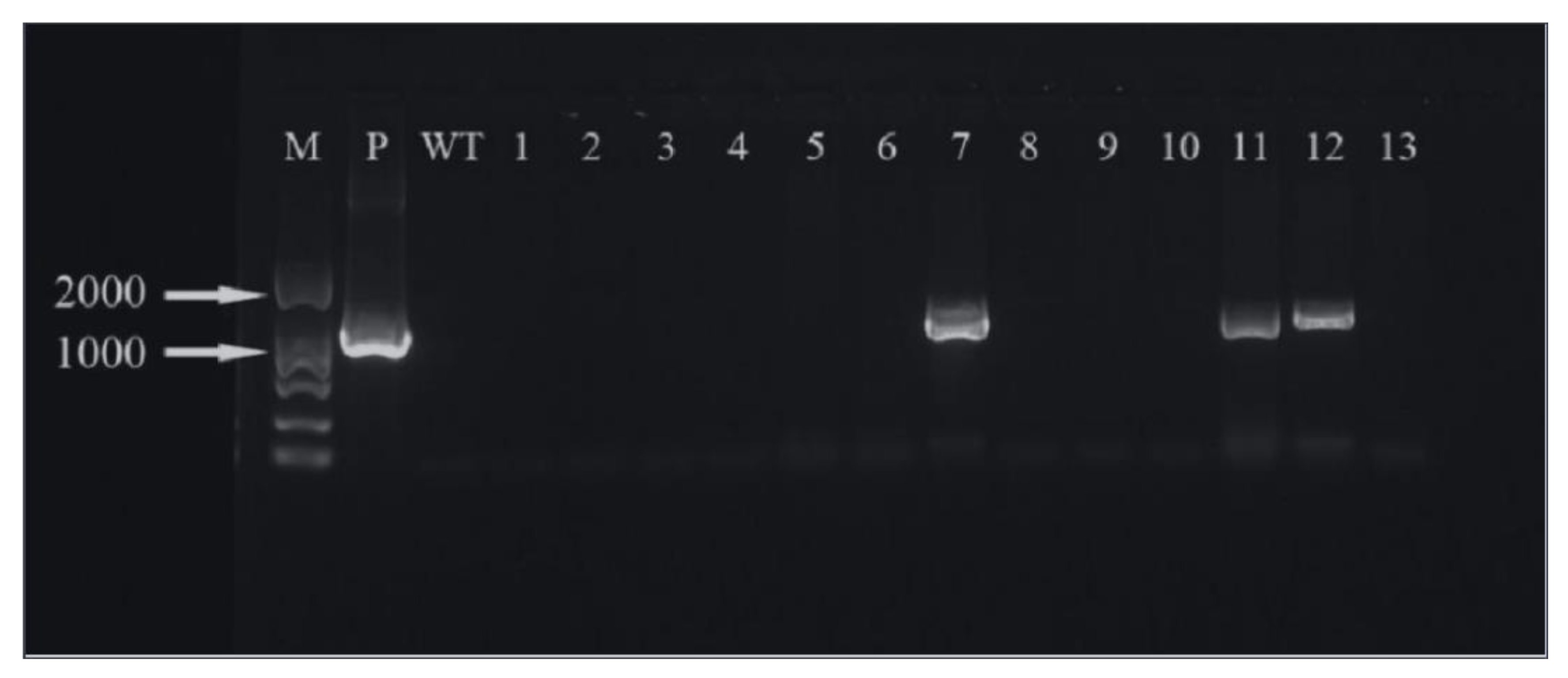
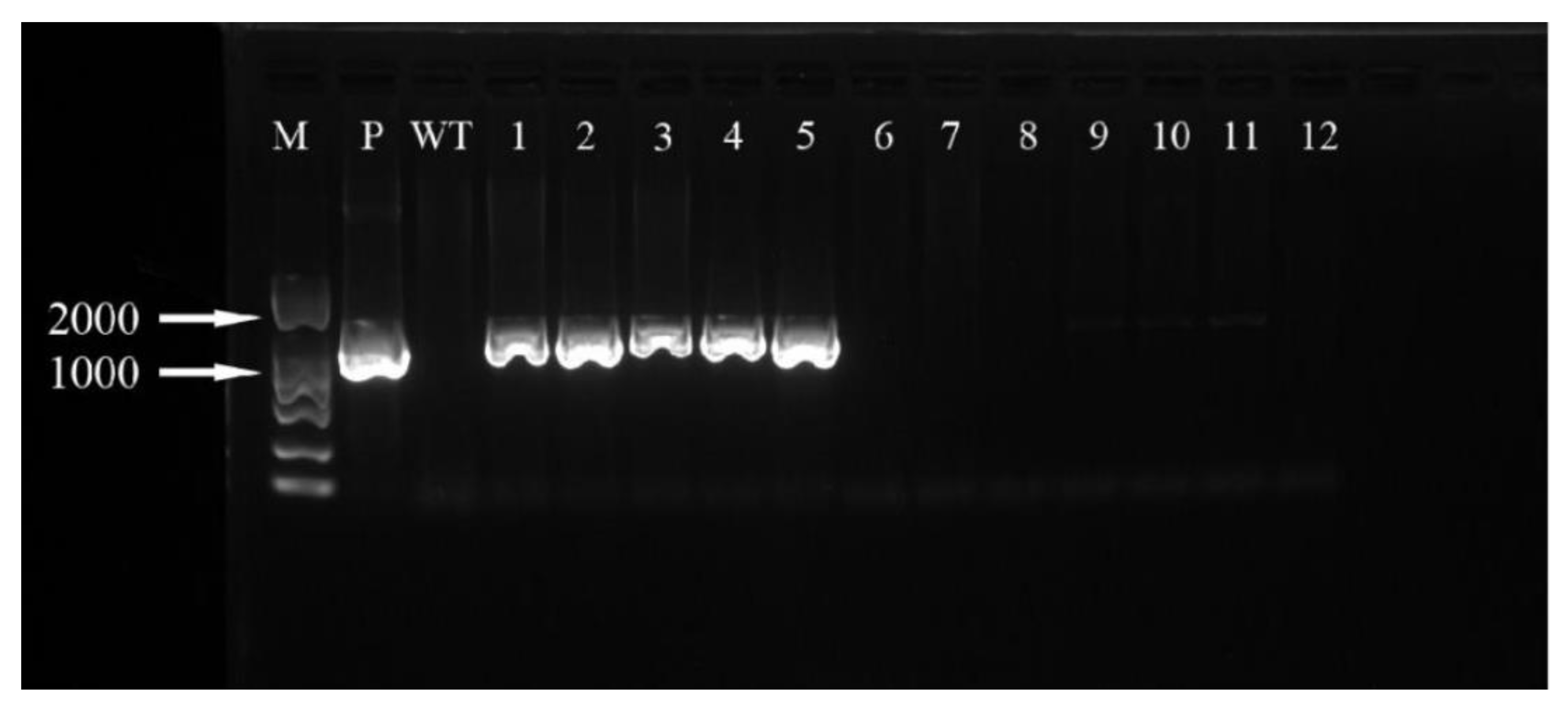
2.8. Transformed Plant Detection
3. Discussion
3.1. Effect of Explants on Embryogenic Callus Induction
3.2. Effect of Plant Growth Regulators on Embryogenic Callus Induction
3.3. Regeneration of Embryogenic Callus
3.4. Antibiotic Concentration Screening and Bacteriostatic Concentration Screening
3.5. Effects of Bacterial Concentration and Infection Time on Genetic Transformation
3.6. Positive Detection of Transformed Plant
4. Materials and Methods
4.1. Plant Materials, Agrobacterium Strain, and Plasmid
4.2. Induction of Embryogenic Callus
4.3. Histological Identification of Callus
4.4. Regeneration of Embryogenic Callus
4.5. Scale-Induced Plant Regeneration
4.6. Plant Overexpression Vector Construction
4.7. Screening of Critical Concentrations of Hygromycin and Cephalosporin
4.8. Preparation of Agrobacterium
4.9. Receptor Material Pre-Incubation
4.10. Infection and Co-Culture
4.11. Screening Culture
4.12. Cultivation of Resistant Seedlings
4.13. PCR Detection of Transformed Plants
4.14. Statistical Analysis
5. Conclusions
- (1)
- Induction of embryogenic callus. The stems and filaments of Oriental lilies ‘Siberia’ and ‘Sorbonne’ were used as explants to induce embryogenic callus. The results showed that ‘Siberia’ is more suitable for inducing embryogenic callus with stems, and the best medium was MS + NAA 1 mg·L−1 + TDZ 0.1 mg·L−1 + 30.0 g·L−1 sucrose + 7.0 g·L−1 agar, with an induction rate of 80.7%, while ‘Sorbonne’ is more suitable for inducing embryogenic callus with filaments, and the best medium was MS + PIC 1 mg·L−1 + TDZ 0.1 mg·L−1 + 30.0 g·L−1 sucrose + 7.0 g·L−1 agar, and the induction rate was 91.7%.
- (2)
- Hygromycin and cephalosporin susceptibility tests. The critical concentrations of hygromycin in the embryonic callus of ‘Siberia’ and ‘Sorbonne’ are 35 mg/L and 50 mg/L, respectively. The critical concentrations of hygromycin in the small scales of sterile seedlings are 60 mg/L and 50 mg/L, respectively. Their optimum concentration of cephalosporin is 300 mg·L−1.
- (3)
- Optimization of genetic transformation systems. The most suitable transformation conditions for the ‘Siberia’ embryonic callus were 0.4 OD600 and 15 min of infection time, resulting in a transformation rate of 60.0%. The optimal transformation conditions of the ‘Sorbonne’ small scales of sterile seedlings were 0.6 OD600 and 20 min infection time, resulting in a transformation rate of 60.0%. However, the ‘Sorbonne’ embryonic callus and scales of ‘Siberia’ sterile seedlings are not suitable as transformation receptors.
Author Contributions
Funding
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Media | Composition |
|---|---|
| the stem axis of ‘Siberia’ and ‘Sorbonne’ | |
| induction medium | MS + NAA (1, 2) mg/L + TDZ (0.1, 0.3, 0.5) mg/L + 30.0 g/L sucrose + 7.0 g/L agar |
| regeneration medium | MS + 30.0 g/L sucrose + 7.0 g/L agar |
| expansion medium | MS + NAA 1 mg/L + 90.0 g/L sucrose + 7.0 g/L agar |
| the filament of ‘Siberia’ and ‘Sorbonne’ | |
| induction medium | MS + PIC 1.0 mg/L + TDZ (0.1, 0.2, 0.3, 0.4, 0.5) mg/L + 30.0 g/L sucrose + 7.0 g/L agar |
| regeneration medium | MS + 30.0 g/L sucrose + 7.0 g/L agar |
| expansion medium | MS + NAA 1 mg/L + 90.0 g/L sucrose + 7.0 g/L agar |
| the sterile scales of ‘Siberia’ | |
| induction medium | MS + 6-BA 1 mg/L + NAA 0.4 mg/L + 30.0 g/L sucrose + 7.0 g/L agar |
| regeneration medium | MS + 30.0 g/L sucrose + 7.0 g/L agar |
| expansion medium | MS + NAA 1 mg/L + 90.0 g/L sucrose + 7.0 g/L agar |
| the sterile scales of ‘Sorbonne’ | |
| induction medium | MS + 6-BA 0.5 mg/L + NAA 1 mg/L + 30.0 g/L sucrose + 7.0 g/L agar |
| regeneration medium | MS + 30.0 g/L sucrose + 7.0 g/L agar |
| expansion medium | MS + NAA 1 mg/L + 90.0 g/L sucrose + 7.0 g/L agar |
| the embryogenic callus of ‘Siberia’ | |
| pre-culture medium | MS + NAA 1 mg/L + TDZ 0.1 mg/L + 30.0 g/L sucrose + 7.0 g/L agar |
| co-culture medium | MS + 1 mg/L NAA + 0.1mg/L TDZ + 100 μmol/L AS + 30.0 g/L sucrose + 7.0 g/L agar |
| screening medium | MS + 1 mg/L NAA + 0.1 mg/L TDZ + 30.0 g/L sucrose + 7.0 g/L agar + 35 mg/L Hyg + 300 mg/L Cef |
| the embryogenic callus of ‘Sorbonne’ | |
| pre-culture medium | MS+ 1.0 mg/L PIC + 0.1 mg/L TDZ + 30.0 g/L sucrose + 7.0 g/L agar |
| co-culture medium | MS + 1.0 mg/L PIC + 0.1 mg/L TDZ + 100 μmol/L AS + 30.0 g/L sucrose + 7.0 g/L agar |
| screening medium | MS + 1.0 mg/L PIC + 0.1 mg/L TDZ + 30.0 g/L sucrose + 7.0 g/L agar + 50 mg/L Hyg + 300 mg/L Cef; |
| the sterile seedling small scales of ‘Siberia’ | |
| pre-culture medium | MS + 6-BA 1mg/L + NAA 0.4 mg/L + 30.0g/L sucrose + 7.0 g/L agar |
| co-culture medium | MS + 1 mg/L6-BA + 0.4 mg/L NAA + 100 μmol/L AS + 30.0 g/L sucrose + 7.0 g/L agar |
| screening medium | MS + 1 mg/L 6-BA + 0.4 mg/L NAA + 30.0 g/L sucrose + 7.0 g/L agar + 60 mg/L Hyg + 300 mg/L Cef |
| the sterile seedling small scales of ‘Sorbonne’ | |
| pre-culture medium | MS + 6-BA 0.5 mg/L + NAA 1 mg/L + 30.0 g/L sucrose + 7.0 g/L agar |
| co-culture medium | MS + 0.5 mg/L 6-BA + 1 mg/L NAA + 100 μmol/L AS + 30.0 g/L sucrose + 7.0 g/L agar |
| screening medium | MS + 0.5 mg/L 6-BA + 1 mg/LNAA + 30.0 g/L sucrose + 7.0 g/L agar 50 mg/LHyg + 300 mg/L Cef |
| Primer Name | Primer Sequence (5′-3′) |
|---|---|
| LaKNOX1-F | ATGGATGGCTTCACCCATCTC |
| LaKNOX1-R | CGGGCCCAGGCGGTATGT |
| LlNAC2-F | ATGGGCGGTCCAGATCTTCA |
| LlNAC2-R | GAACGGCTTCAGCAAGTGCA |
| Hyg-F | ATGGATGGCTTCACCCATCTC |
| Hyg-R | CGGGCCCAGGCGGTATGT |
| Reagent | Volume/μL |
|---|---|
| pBI121-LaKNOX1/LlNAC2-GFP | 2 |
| forward primer (10 μM) | 2 |
| Reverse primer (10 μM) | 2 |
| 2× Phanta® Max Master Mix (Dye Plus) | 10 |
| ddH2O | 4 |
| total volume | 20 |
| Reagent | Volume/μL |
|---|---|
| pCAMBIA1300-GUS | 40 |
| Xbal (15 U/μL) | 2.5 |
| SalI (10 U/μL) | 2.5 |
| Cutsmart | 5 |
| total volume | 50 |
| Reagent | Volume/μL |
|---|---|
| pCAMBIA1300-GUS double digestion linear plasmid | >1 |
| target gene | >1 |
| 5× CE II buffer | 4 |
| Exnase II | 2 |
| ddH2O | to 20 |
| total volume | 20 |
References
- Liu, Z.; Zhao, X.; Wang, W.; Ke, Z.; Wang, S. The diversity and application of Garden Lily. Mod. Landsc. Archit. 2014, 11, 100–103. [Google Scholar]
- Wang, J.M.; Ma, S.L.; Li, W.Q.; Wang, Q.; Cao, H.Y.; Gu, J.H.; Lu, Y.M. Genetic variability and diversity of the main resources of lily assessed via phenotypic characters, pollen morphology, and ISSR markers. Genet. Mol. Res. 2016, 4, 15. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yong, Y.; Zhang, Y.; Lyu, Y. Transcriptional regulatory network of GA floral induction pathway in LA Hybrid Lily. Int. J. Mol. Sci. 2019, 20, 2694. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yong, Y.B.; Wang, Q.; Lu, Y.M. Physiological and molecular changes during lily underground stem axillary bulbils formation. Russ. J. Plant Physiol. 2018, 65, 372–383. [Google Scholar] [CrossRef]
- Zambryski, P.; Joos, H.; Genetello, C.; Leemans, J.; Van Montagu, M.; Schell, J. Ti plasmid vector for the introduction of DNA into plant cells without alteration of their normal regeneration capacity. EMBO J. 1983, 2, 2143–2150. [Google Scholar] [CrossRef] [PubMed]
- Rai, G.K.; Rai, N.P.; Kumar, S.; Yadav, A.; Rathaur, S.; Singh, M. Effects of explant age, germination medium, pre-culture parameters, inoculation medium, pH, washing medium, and selection regime on Agrobacterium-mediated transformation of tomato. In Vitro Cell. Dev. Biol.-Plant 2012, 48, 565–578. [Google Scholar] [CrossRef]
- Song, S.; Yan, R.; Wang, C.; Wang, J.; Sun, H. Improvement of a genetic transformation system and preliminary study on the function of LpABCB21 and LpPILS7 based on somatic embryogenesis in Lilium pumilum DC. Fisch. Int. J. Mol. Sci. 2020, 18, 6784. [Google Scholar] [CrossRef]
- Cohen, A.; Meredith, C.P. Agrobactierium-mediated transformation of Lilium. Acta Hortic. 1992, 325, 611–618. [Google Scholar] [CrossRef]
- Dong, W.; Mao, Y.F.; Li, W. Factors influencing T-DNA transferring into pollen of lily in vitro. Russ. J. Plant Physiol. 2007, 54, 420–425. [Google Scholar] [CrossRef]
- Ogaki, M.; Furuichi, Y.; Kuroda, K.; Chin, D.P.; Ogawa, Y.; Mii, M. Importance of co-cultivation medium pH for successful Agrobacterium-mediated transformation of Lilium x formolongi. Plant Cell Rep. 2008, 27, 699–705. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, J.; Xu, B.; Jia, C.; Zhang, J.; Tan, G.; Jin, Z. Regeneration and production of transgenic Lilium longiflorum via Agrobacterium tumefaciens. In Vitro Cell. Dev. Biol.-Plant 2011, 47, 348–356. [Google Scholar] [CrossRef]
- Yong, Y.; Zhang, Y.; Lyu, Y. Functional characterization of Lilium lancifolium cold-responsive Zinc Finger Homeodomain (ZFHD) gene in abscisic acid and osmotic stress tolerance. PeerJ 2021, 9, e11508. [Google Scholar] [CrossRef] [PubMed]
- Yuan, G.; Wu, Z.; Liu, X.; Li, T.; Teng, N. Characterization and functional analysis of LoUDT1, a bHLH transcription factor related to anther development in the lily oriental hybrid Siberia (Lilium spp.). Plant Physiol. Biochem. 2021, 166, 1087–1095. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, Y.; Lan, W.; Zhou, Q.; Li, F.; Chen, F.; Bao, M.; Liu, G. LlDREB1G, a novel DREB subfamily gene from Lilium longiflorum, can enhance transgenic Arabidopsis tolerance to multiple abiotic stresses. Plant Cell Tissue Organ Cult. (PCTOC) 2019, 138, 489–506. [Google Scholar] [CrossRef]
- Li, J.; Chai, M.; Zhu, X.; Zhang, X.; Li, H.; Wang, D.; Xing, Q.; Zhang, J.; Sun, M.; Shi, L. Cloning and expression analysis of LoCCD8 during IAA-induced bulbils outgrowth in lily (Oriental Hybrid ‘Sorbonne’). J. Plant Physiol. 2019, 236, 39–50. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhao, Y.Q.; Gao, X.; Jia, G.X. Analysis of miRNA-mediated regulation of flowering induction in Lilium × formolongi. BMC Plant Biol. 2021, 21, 190. [Google Scholar] [CrossRef]
- Cao, L.; Liu, D.; Jiang, F.; Wang, B.; Wu, Y.; Che, D.; Fan, J. Heterologous Expression of LiSEP3 from Oriental Lilium Hybrid ‘Sorbonne’ Promotes the Flowering of Arabidopsis thaliana L. Mol. Biotechnol. 2022, 64, 1120–1129. [Google Scholar] [CrossRef]
- Kurokawa, K.; Kobayashi, J.; Nemoto, K.; Nozawa, A.; Sawasaki, T.; Nakatsuka, T. Expression of LhFT1, the Flowering Inducer of Asiatic Hybrid Lily, in the Bulb Scales. Front. Plant Sci. 2020, 11, 570915. [Google Scholar] [CrossRef]
- Yang, J.; Meng, J.; Liu, X.; Hu, J.; Zhu, Y.; Zhao, Y.; Jia, G.; He, H.; Yuan, T. Integrated mRNA and small RNA sequencing reveals a regulatory network associated with flower color in oriental hybrid lily. Plant Physiol. Biochem. 2021, 166, 103–114. [Google Scholar] [CrossRef]
- Cao, Y.; Xu, L.; Xu, H.; Yang, P.; He, G.; Tang, Y.; Qi, X.; Song, M.; Ming, J. LhGST is an anthocyanin-related glutathione S-transferase gene in Asiatic hybrid lilies (Lilium spp.). Plant Cell Rep. 2021, 40, 85–95. [Google Scholar] [CrossRef]
- Zhang, T.; Guo, Y.; Shi, X.; Yang, Y.; Chen, J.; Zhang, Q.; Sun, M. Overexpression of LiTPS2 from a cultivar of lily (Lilium ‘Siberia’) enhances the monoterpenoids content in tobacco flowers. Plant Physiol. Biochem. 2020, 151, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Abbas, F.; Ke, Y.; Zhou, Y.; Ashraf, U.; Li, X.; Yu, Y.; Yue, Y.; Ahmad, K.W.; Yu, R.; Fan, Y. Molecular cloning, characterization and expression analysis of LoTPS2 and LoTPS4 involved in floral scent formation in oriental hybrid Lilium variety ‘Siberia’. Phytochemistry 2020, 173, 112294. [Google Scholar] [CrossRef] [PubMed]
- Yan, R.; Wang, Z.; Ren, Y.; Li, H.; Liu, N.; Sun, H. Establishment of efficient genetic transformation systems and application of CRISPR/Cas9 genome editing technology in Lilium pumilum DC. Fisch. And Lilium longiflorum White Heaven. Int. J. Mol. Sci. 2019, 20, 2920. [Google Scholar] [CrossRef] [Green Version]
- Bakhshaie, M.; Khosravi, S.; Azadi, P.; Bagheri, H.; van Tuyl, J.M. Biotechnological advances in Lilium. Plant Cell Rep. 2016, 35, 1799–1826. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Retuerto, R. Small-scale Heterogeneity in Soil Quality Influences Photosynthetic Efficiency and Habitat Selection in a Clonal Plant. Ann. Bot. 2006, 98, 1043–1052. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, Y.; Liu, X.; Huang, J.; Wang, Q.; Gu, J.; Lu, Y. Transcriptome profiling of the cold response and signaling pathways in Lilium lancifolium. BMC Genom. 2014, 15, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Wang, Q.; Yang, Y.; Liu, X.; Gu, J.; Li, W.; Ma, S.; Lu, Y. De novo assembly and characterization of stress transcriptome and regulatory networks under temperature, salt and hormone stresses in Lilium lancifolium. Mol. Biol. Rep. 2014, 41, 8231–8245. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter, R.; Vollbrecht, E.; Lowe, B.; Veit, B.; Yamaguchi, J.; Hake, S. Sequence analysis and expression patterns divide the maize knotted1-like homeobox genes into two classes. Plant Cell 1994, 6, 1877–1887. [Google Scholar]
- Reiser, L.; Sánchez-Baracaldo, P.; Hake, S. Knots in the family tree: Evolutionary relationships and functions of knox homeobox genes. Plant Mol. Biol. 2000, 42, 151–166. [Google Scholar] [CrossRef]
- Sinha, N.R.; Williams, R.E.; Hake, S. Overexpression of the maize homeo box gene, KNOTTED-1, causes a switch from determinate to indeterminate cell fates. Genes Dev. 1993, 7, 787–795. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.E.; Jacobsen, S.E.; Levin, J.Z.; Meyerowitz, E.M. The clavata and shoot meristemless loci competitively regulate meristem activity in Arabidopsis. Development 1996, 122, 1567–1575. [Google Scholar] [CrossRef] [PubMed]
- Kerstetter, R.A.; Laudencia-Chingcuanco, D.; Smith, L.G.; Hake, S. Loss-of-function mutations in the maize homeobox gene, knotted1, are defective in shoot meristem maintenance. Development 1997, 124, 3045–3054. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.L.; Gago, G.M.; Palena, C.M.; Gonzalez, D.H. Homeoboxes in plant development. Biochim. Biophys. Acta 1998, 1442, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Smith, L.G.; Greene, B.; Veit, B.; Hake, S. A dominant mutation in the maize homeobox gene, Knotted-1, causes its ectopic expression in leaf cells with altered fates. Development 1992, 116, 21–30. [Google Scholar] [CrossRef]
- Hay, A.; Tsiantis, M. KNOX genes: Versatile regulators of plant development and diversity. Development 2010, 137, 3153–3165. [Google Scholar] [CrossRef] [Green Version]
- Hake, S.; Smith, H.M.; Holtan, H.; Magnani, E.; Mele, G.; Ramirez, J. The role of knox genes in plant development. Annu. Rev. Cell Dev. Biol. 2004, 20, 125–151. [Google Scholar] [CrossRef]
- Frugis, G.; Giannino, D.; Mele, G.; Nicolodi, C.; Chiappetta, A.; Bitonti, M.B.; Innocenti, A.M.; Dewitte, W.; Van Onckelen, H.; Mariotti, D. Overexpression of KNAT1 in lettuce shifts leaf determinate growth to a shoot-like indeterminate growth associated with an accumulation of isopentenyl-type cytokinins. Plant Physiol. 2001, 126, 1370–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.; Cho, Y.H.; Ryu, H.; Kim, Y.; Kim, T.H.; Hwang, I. BLH1 and KNAT3 modulate ABA responses during germination and early seedling development in Arabidopsis. Plant J. 2013, 75, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Zhong, R.; Lee, C.; Zhou, J.; McCarthy, R.L.; Ye, Z.H. A battery of transcription factors Involved in the regulation of secondary cell wall biosynthesis in Arabidopsis. Plant Cell. 2008, 20, 2763–2782. [Google Scholar] [CrossRef] [Green Version]
- Truernit, E.; Haseloff, J. A Role for KNAT Class II Genes in Root Development. Plant Signal Behav. 2007, 2, 10. [Google Scholar] [CrossRef] [Green Version]
- Bhargava, A.; Mansfield, S.D.; Hall, H.C.; Douglas, C.J.; Ellis, B.E.; Bhargava, A.; Mansfield, S.D.; Hall, H.C. MYB75 Functions in Regulation of Secondary Cell Wall Formation in the Arabidopsis Inflorescence Stem. Plant Physiol. 2010, 154, 1428–1438. [Google Scholar] [PubMed] [Green Version]
- Li, E.; Wang, S.; Liu, Y.; Chen, J.G.; Douglas, C.J. OVATE FAMILY PROTEIN4 (OFP4) interaction with KNAT7 regulates secondary cell wall formation in Arabidopsis thaliana. Plant J. 2011, 67, 328–341. [Google Scholar] [CrossRef]
- Li, E.; Bhargava, A.; Qiang, W.; Friedmann, M.C.; Forneris, N.; Savidge, R.A.; Johnson, L.A.; Mansfield, S.D.; Ellis, B.E.; Douglas, C.J. The Class II KNOX gene KNAT7 negatively regulates secondary wall formation in Arabidopsis and is functionally conserved in Populus. New Phytol. 2012, 194, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Scofield, S.; Murray, J.A. KNOX Gene Function in Plant Stem Cell Niches. Plant Mol. Biol. 2006, 60, 929–946. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Meilan, R.; Woeste, K. A KNAT3-like homeobox gene from Juglans nigra L. JnKNAT3-like, highly expressed during heartwood formation. Plant Cell Rep. 2009, 28, 1717–1724. [Google Scholar]
- Moreno-Pachón, N.M. Mechanisms of Vegetative Propagation in Bulbs: A Molecular Approach. Ph.D. Thesis, Wageningen University, Wageningen, The Netherlands, 2017. [Google Scholar]
- Chung, P.J.; Jung, H.; Choi, Y.D.; Kim, J.K. Genome-wide analyses of direct target genes of four rice NAC-domain transcription factors involved in drought tolerance. BMC Genom. 2018, 19, 40. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Deng, Z.; Lai, J.; Zhang, Y.; Yang, C.; Yin, B.; Zhao, Q.; Zhang, L.; Li, Y.; Yang, C.; et al. Dual function of Arabidopsis ATAF1 in abiotic and biotic stress responses. Cell Res. 2009, 19, 1279–1290. [Google Scholar] [CrossRef] [Green Version]
- Yong, Y.; Zhang, Y.; Lyu, Y. A Stress-Responsive NAC Transcription Factor from Tiger Lily (LlNAC2) Interacts with LlDREB1 and LlZHFD4 and Enhances Various Abiotic Stress Tolerance in Arabidopsis. Int. J. Mol. Sci. 2019, 20, 3225. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Wang, L.; Nawaz, M.A.; Niu, M.; Sun, J.; Xie, J.; Kong, Q.; Huang, Y.; Cheng, F.; Bie, Z. Ectopic expression of Pumpkin NAC transcription factor CmNAC1 improves multiple abiotic stress tolerance in Arabidopsis. Front. Plant Sci. 2017, 8, 2052. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Sun, Y.; Wang, H.; Shi, H.; Su, M.; Shan, H.; Li, T.; Li, Q. The SlNAC8 gene of the halophyte Suaeda liaotungensis enhances drought and salt stress tolerance in transgenic Arabidopsis thaliana. Gene 2018, 662, 10–20. [Google Scholar] [CrossRef]
- Wang, L.; Li, Z.; Lu, M.; Wang, Y. ThNAC13, a NAC transcription factor from Tamarix hispida, confers salt and osmotic stress tolerance to transgenic Tamarix and Arabidopsis. Front. Plant Sci. 2017, 8, 635. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Zhang, X.; Duan, H.; Lian, C.; Liu, C.; Yin, W.; Xia, X. Three stress-responsive NAC transcription factors from Populus euphratica differentially regulate salt and drought tolerance in transgenic plants. Physiol. Plant. 2018, 162, 73–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.Y.; Niu, L.X.; Zhang, Y.L.; Zhang, X.G.; Li, S.H. Study on callus induction and rooting of wild lily organs. Res. Prog. Ornam. Hortic. China 2016, 6, 448–453. [Google Scholar]
- Liu, X.; Gu, J.; Wang, J.; Lu, Y. Lily breeding by using molecular tools and transformation systems. Mol. Biol. Rep. 2014, 41, 6899–6908. [Google Scholar] [CrossRef]
- Qi, Y.; Du, L.; Quan, Y.; Tian, F.; Liu, Y.; Wang, Y. Agrobacterium-mediated transformation of embryogenic cell suspension cultures and plant regeneration in Lilium tenuifolium Oriental × trumpet ‘Robina’. Acta Physiol. Plant. 2014, 36, 2047–2057. [Google Scholar] [CrossRef]
- Mori, S.; Adachi, Y.; Horimoto, S.; Suzuki, S.; Nakano, M. Callus formation and plant regeneration in various Lilium species and cultivars. In Vitro Cell. Dev. Biol. Plant 2005, 41, 783–788. [Google Scholar] [CrossRef]
- Zhang, J.; Gai, M.; Li, X.; Li, T.; Sun, H. Somatic embryogenesis and direct as well as indirect organogenesis in Lilium pumilum DC. Fisch., an endangered ornamental and medicinal plant. Biosci. Biotechnol. Biochem. 2016, 80, 1898–1906. [Google Scholar] [CrossRef] [Green Version]
- Loyola-Vargas, V.M.; Ochoa-Alejo, N. The Many Ways of Somatic Embryo Initiation; Springer: Berlin/Heidelberg, Germany, 2016; Volume 3, pp. 23–37. [Google Scholar]
- Gnanaraj, M.; Sneka, C.; Sisubalan, N.; Baburajan, R.; Manikandan, R.; Muneeswaran, T. Polyethylene glycol induced somatic embryogenesis and plant regeneration from cotyledons of Vigna radiata (L.) Wilczek. South Afr. J. Bot. 2022, 150, 721–730. [Google Scholar] [CrossRef]
- Nakano, M.; Otani, M. Plant regeneration and Agrobacterium-mediated genetic transformation systems in liliaceous ornamental plants. Plant Biotechnol. 2020, 37, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Oota, M.; Nakano, M. Embryogenic callus induction from leaf explants of the Liliaceous ornamental plant, Agapanthus praecox ssp. orientalis (Leighton) Leighton-Histological study and response to selective agents. Sci. Hortic. 2002, 95, 123–132. [Google Scholar] [CrossRef]
- Ahmed, R.I.; Ding, A.; Xie, M.; Kong, Y. Progress in Optimization of Agrobacterium-Mediated Transformation in Sorghum (Sorghum bicolor). Int. J. Mol. Sci. 2018, 19, 2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pushyami, B.; Beena, M.R.; Sinha, M.K.; Kirti, P.B. In vitro regeneration and optimization of conditions for Agrobacterium mediated transformation in jute, Corchorus capsularis. J. Plant Biochem. Biotechnol. 2011, 20, 39–46. [Google Scholar] [CrossRef]
- Li, S.; Yang, N.; Chen, L. Paraffin section observation of flower bud differentiation of Chimonanthus praecox in Kunming and comparison of the differentiation processes in different regions, China. Hortic. Plant J. 2022, 8, 221–229. [Google Scholar] [CrossRef]
- Zhou, N.F.; Zhang, J.P.; Liu, H.; Weiwei, Z.; Pei, D. New protocols for paraffin sections of heterogeneous tissues of woody plants. Chin. Bull. Bot. 2018, 53, 653–660. [Google Scholar]
- Zhang, X.W.; Liu, L.P. Research on dandelion paraffin sectioning technology. Spec. Econ. Anim. Plants 2022, 25, 18–20. [Google Scholar]




















| Medium Number | NAA Concentration (mg·L−1) | TDZ Concentration (mg·L−1) | Callus Induction Rate (%) | Browning Rate (%) |
|---|---|---|---|---|
| N1 | 1 | 0.1 | 80.7 ± 4.38 a | 19.3 ± 4.38 d |
| N2 | 1 | 0.3 | 61.1 ± 3.20 b | 38.9 ± 3.20 c |
| N3 | 1 | 0.5 | 27.8 ± 3.20 d | 72.2 ± 3.20 a |
| N4 | 2 | 0.1 | 42.6 ± 4.91 c | 57.4 ± 4.91 b |
| N5 | 2 | 0.3 | 57.4 ± 1.83 b | 42.6 ± 1.83 c |
| N6 | 2 | 0.5 | 63.0 ± 4.88 b | 37.0 ± 4.88 c |
| Medium Number | NAA Concentration (mg·L−1) | TDZ Concentration (mg·L−1) | Callus Induction Rate (%) | Browning Rate (%) |
|---|---|---|---|---|
| N1 | 1 | 0.1 | 13.0 ± 1.87 a | 87.0 ± 1.83 c |
| N2 | 1 | 0.3 | 5.6 ± 0.00 bc | 94.4 ± 0.00 ab |
| N3 | 1 | 0.5 | 5.6 ± 3.2 bc | 94.4 ± 3.20 ab |
| N4 | 2 | 0.1 | 3.7 ± 1.87 c | 96.3 ± 1.87 a |
| N5 | 2 | 0.3 | 11.1 ± 0.00 bc | 88.9 ± 0.00 bc |
| N6 | 2 | 0.5 | 7.4 ± 1.83 abc | 92.6 ± 1.83 abc |
| Medium Number | PIC Concentration (mg·L−1) | TDZ Concentration (mg·L−1) | Callus Induction Rate (%) | Browning Rate (%) |
|---|---|---|---|---|
| P1 | 1 | 0.0 | 38.9 ± 2.78 b | 15.3 ± 1.39 d |
| P2 | 1 | 0.1 | 47.2 ± 2.78 a | 23.6 ± 1.40 c |
| P3 | 1 | 0.2 | 11.1 ± 1.39 c | 55.6 ± 2.78 a |
| P4 | 1 | 0.3 | 33.3 ± 0.00 b | 33.3 ± 4.81 b |
| P5 | 1 | 0.4 | 33.3 ± 4.81 b | 33.3 ± 0.00 b |
| P6 | 1 | 0.5 | 36.1 ± 1.39 b | 13.9 ± 1.39 d |
| Medium Number | PIC Concentration (mg·L−1) | TDZ Concentration (mg·L−1) | Callus Induction Rate (%) | Browning Rate (%) |
|---|---|---|---|---|
| P1 | 1 | 0.0 | 49.5 ± 2.82 d | 0.00 |
| P2 | 1 | 0.1 | 91.7 ± 4.81 a | 0.00 |
| P3 | 1 | 0.2 | 77.8 ± 2.78 bc | 0.00 |
| P4 | 1 | 0.3 | 83.3 ± 0.00 abc | 0.00 |
| P5 | 1 | 0.4 | 86.1 ± 1.39 ab | 0.00 |
| P6 | 1 | 0.5 | 75.0 ± 2.41 c | 0.00 |
| OD 600 Infection Concentration | Infection Time (min) | Infected Embryogenic Callus Number | Resistant Embryogenic Callus Number | Embryogenic Callus Transformation Rate (%) |
|---|---|---|---|---|
| 0.2 | 10 | 30 | 9 | 30.0% |
| 0.2 | 15 | 30 | 11 | 36.7% |
| 0.2 | 20 | 30 | 12 | 40.0% |
| 0.4 | 10 | 30 | 14 | 46.7% |
| 0.4 | 15 | 30 | 18 | 60.0% |
| 0.4 | 20 | 30 | 13 | 43.3% |
| OD 600 Infection Concentration | Infection Time (min) | Infected Sterile Seedling Small Scales Number | Induced Number of Resistant Buds | Resistant Transformation Rate (%) |
|---|---|---|---|---|
| 0.6 | 10 | 30 | 7 | 23.3% |
| 0.6 | 15 | 30 | 10 | 33.3% |
| 0.6 | 20 | 30 | 18 | 60.0% |
| 0.8 | 10 | 30 | 16 | 53.3% |
| 0.8 | 15 | 30 | 13 | 43.3% |
| 0.8 | 20 | 30 | 7 | 23.3% |
| OD 600 Infection Concentration | Infection Time (min) | Infected Sterile Seedling Small Scales Number | Induced Number of Resistant Buds | Resistant Transformation Rate (%) |
|---|---|---|---|---|
| 0.6 | 10 | 30 | 0 | 0 |
| 0.6 | 15 | 30 | 0 | 0 |
| 0.6 | 20 | 30 | 1 | 3.3 |
| 0.8 | 10 | 30 | 0 | 0 |
| 0.8 | 15 | 30 | 1 | 3.3 |
| 0.8 | 20 | 30 | 4 | 13.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Y.; Hou, X.; Zheng, Y.; Lyu, Y. The Establishment of a Genetic Transformation System and the Acquisition of Transgenic Plants of Oriental Hybrid Lily (Lilium L.). Int. J. Mol. Sci. 2023, 24, 782. https://doi.org/10.3390/ijms24010782
Chen Y, Hou X, Zheng Y, Lyu Y. The Establishment of a Genetic Transformation System and the Acquisition of Transgenic Plants of Oriental Hybrid Lily (Lilium L.). International Journal of Molecular Sciences. 2023; 24(1):782. https://doi.org/10.3390/ijms24010782
Chicago/Turabian StyleChen, Yue, Xinru Hou, Yupin Zheng, and Yingmin Lyu. 2023. "The Establishment of a Genetic Transformation System and the Acquisition of Transgenic Plants of Oriental Hybrid Lily (Lilium L.)" International Journal of Molecular Sciences 24, no. 1: 782. https://doi.org/10.3390/ijms24010782