
Shikimate Kinase Plays Important Roles in Anthocyanin Synthesis in Petunia

Abstract
:1. Introduction
2. Results
2.1. Isolation and Sequence Analyses of PhSK
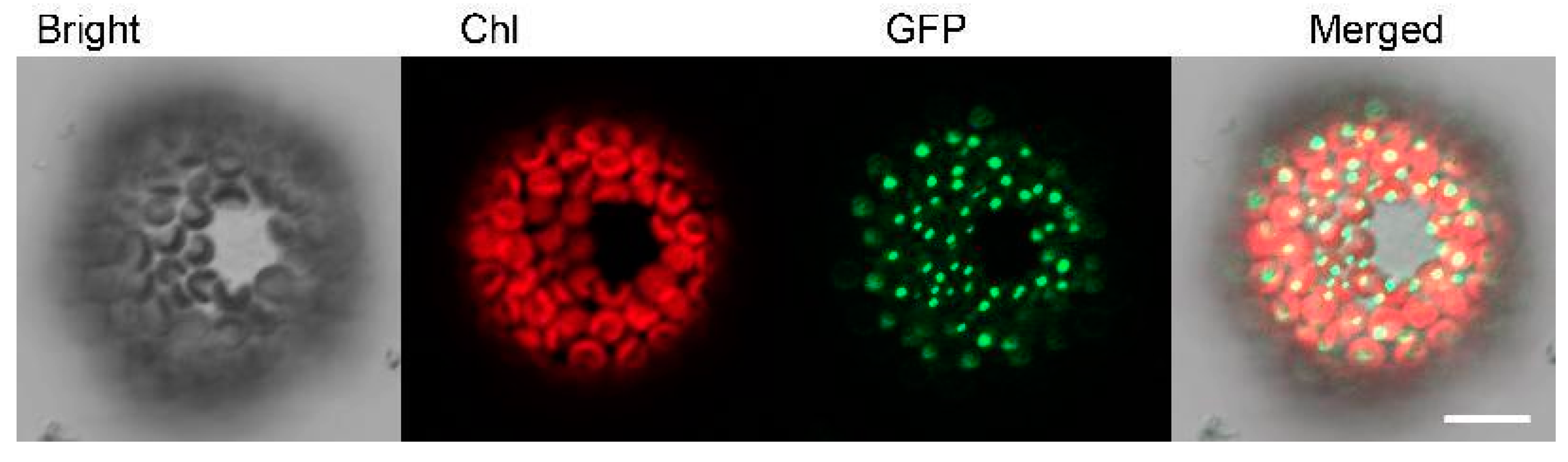
2.2. PhSK Protein Localization in Chloroplasts
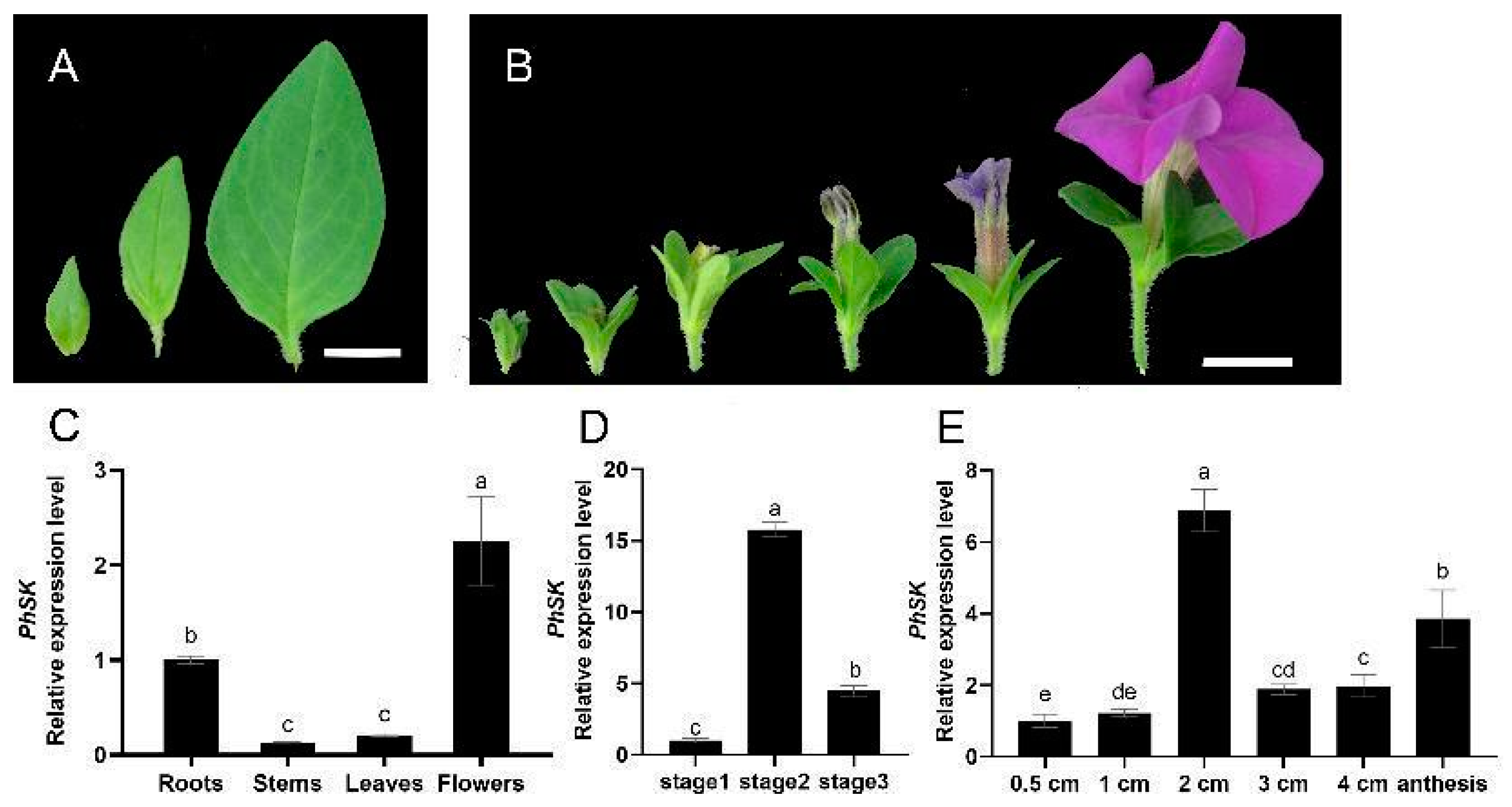
2.3. PhSK Expression
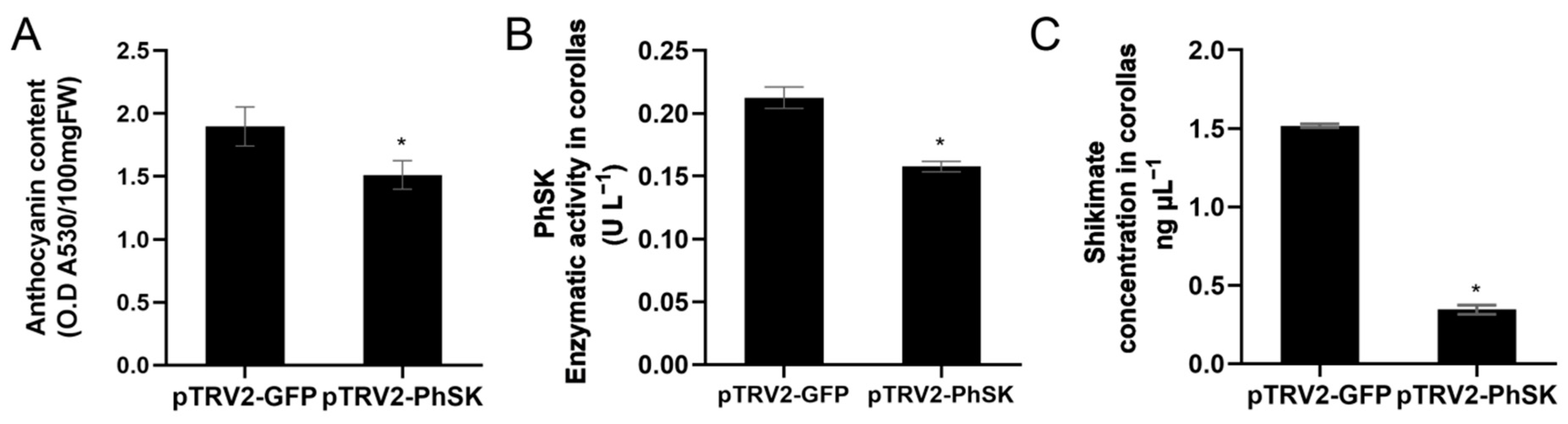
2.4. Phenotype of PhSK-Silenced Petunia Plants
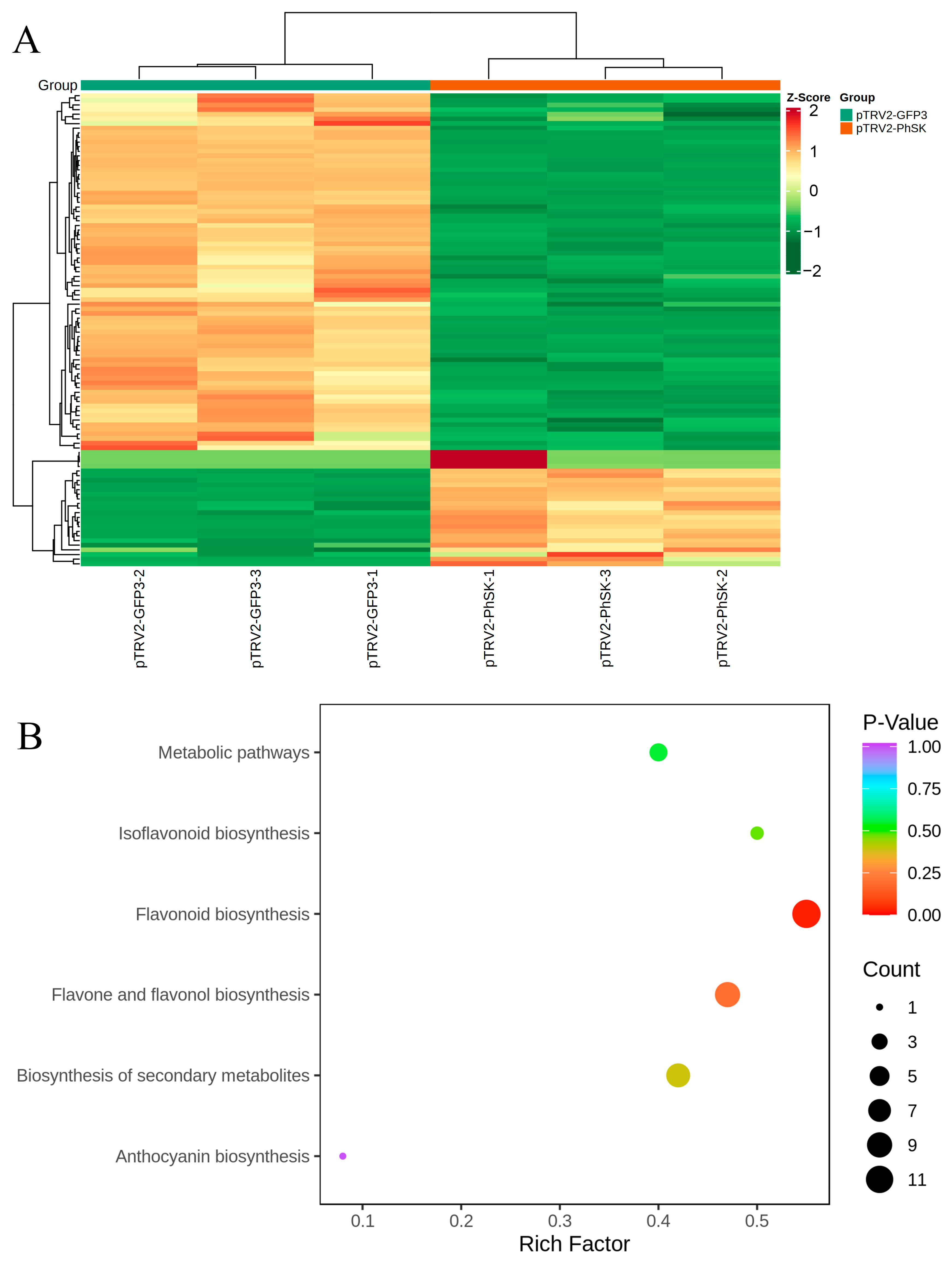
2.5. Changes in the Corolla Flavonoid Metabolome Profile Induced by PhSK Silencing
2.6. PhSK Silencing Reduces the Shikimate Content in Corollas
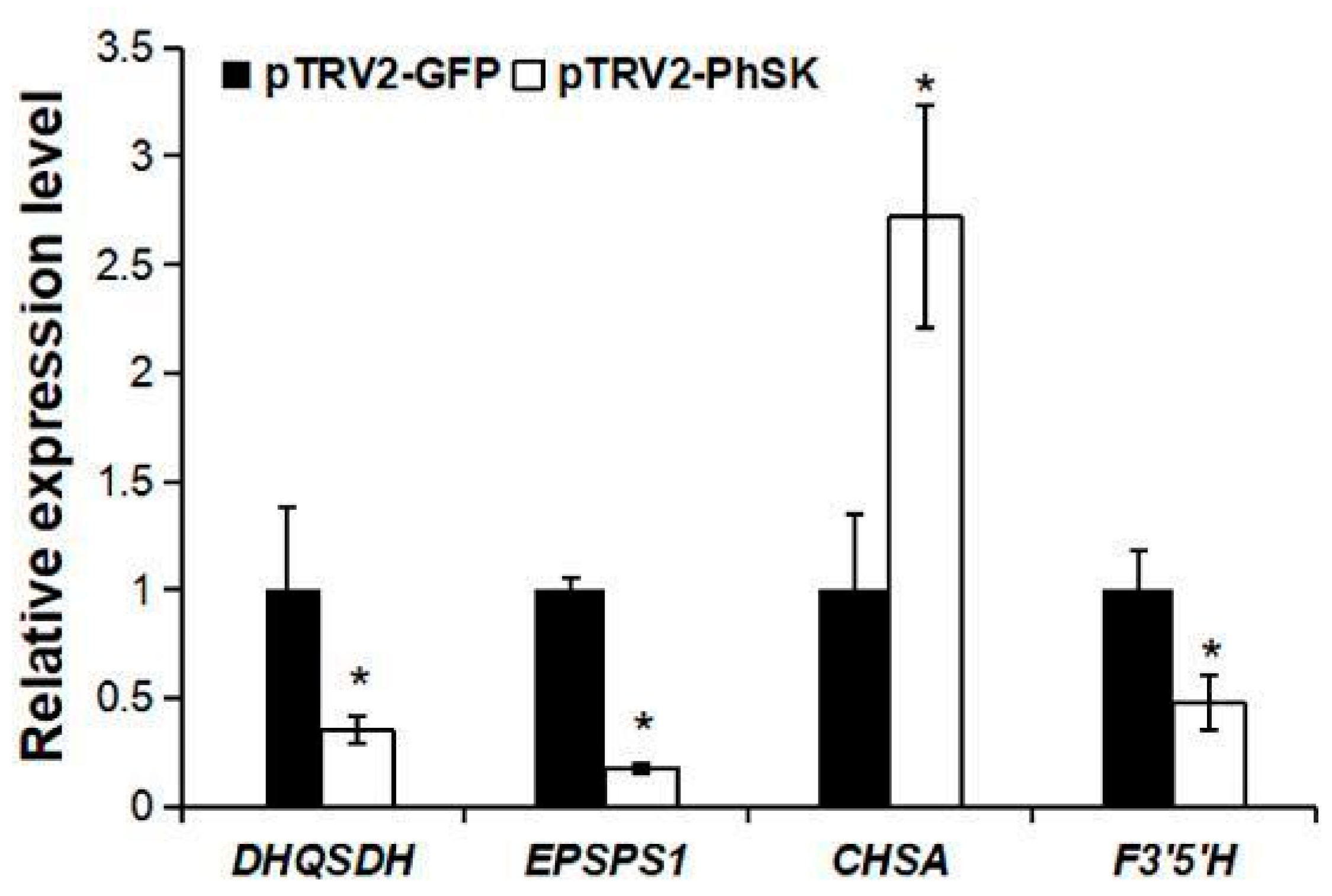
2.7. Effect of PhSK Silencing on the Expression of Some Structural Genes of the Shikimate Pathway and Anthocyanin Synthesis Pathway
3. Discussion
4. Materials and Methods
4.1. RNA Extraction, RT-PCR, and Cloning of the Petunia PhSK Gene
4.2. Sequence Analysis
4.3. Subcellular Localization
4.4. Quantitative Real-Time PCR Assays
4.5. Agroinoculation of pTRV2 Vectors
4.6. Assays of SK Activity
4.7. Anthocyanin Extraction and Measurement
4.8. Widely Targeted Metabolomics Analysis
4.9. Qualitative and Quantitative Determination of Metabolites
4.10. Shikimate Measurement
4.11. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roberts, F.; Roberts, C.; Johnson, J.J.; Kylek, D.E.; Krell, T.; Coggins, J.R.; Coombs, G.H.; Milhousk, W.K.; Tzipori, S.; Ferguson, D.J.P.; et al. Evidence for the shikimate pathway in apicomplexan parasites. Nature 1998, 393, 801–805. [Google Scholar] [CrossRef] [PubMed]
- Bentley, R.; Haslam, E. The shikimate pathway—A metabolic tree with many branche. Crit. Rev. Biochem. Mol. Biol. 1990, 25, 307–384. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, K.M. The shikimate pathway: Early steps in the biosynthesis of aromatic compounds. Plant Cell 1995, 7, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, K.M.; Weaver, L.M. The shikimate pathway. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 473–503. [Google Scholar] [CrossRef]
- Tzin, V.; Gad, G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Mol. Plant 2010, 3, 956–972. [Google Scholar] [CrossRef]
- Keith, B.; Dong, X.; Ausubel, F.M.; Fink, G.R. Differential induction of 3-deoxy-D-arabino-heptulosonate 7-phosphate synthase genes in Arabidopsis thaliana by wounding and pathogenic attack. Proc. Natl. Acad. Sci. USA 1991, 88, 8821–8825. [Google Scholar] [CrossRef] [Green Version]
- Görlach, J.; Rawsecke, H.; Rentsch, D.; Regenass, M.; Roy, P.; Zala, M.; Keel, C.; Boller, T.; Amrhein, N.; Schmid, J. Temporally distinct accumulation of transcripts encoding enzymes of the prechorismate pathway in elicitor-treated, cultured tomato cells. Proc. Natl. Acad. Sci. USA 1995, 92, 3166–3170. [Google Scholar] [CrossRef] [Green Version]
- Maeda, H.; Dudareva, N. The shikimate pathway and aromatic amino Acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef]
- Cao, M.; Li, Y.; An, Z.; Cheng, H.; Hu, Y.; Huang, H. Effects of overexpression of HbDAHPS on stress tolerance in Arabidopsis thaliana. Mol. Plant Breed. 2016, 14, 1107–1114. [Google Scholar] [CrossRef]
- Hamberger, B.; Ehlting, J.; Barbazuk, W.B.; Douglas, C.J. Comparative genomics of the shikimate pathway in Arabidopsis, Populus trichocarpa and Oryza sativa: Shikimate pathway gene family structure and identification of candidates for missing links in phenylalanine biosynthesis. Recent Adv. Phytochem. 2006, 40, 85–113. [Google Scholar] [CrossRef]
- Li, D.; Hofius, D.; Hajirezaei, M.; Fernie, A.R.; Börnke, F.; Sonnewald, U. Functional analysis of the essential bifunctional tobacco enzyme 3-dehydroquinate dehydratase/shikimate dehydrogenase in transgenic tobacco plants. J. Exp. Bot. 2007, 58, 2053–2067. [Google Scholar] [CrossRef] [Green Version]
- Henstrand, J.M.; McCue, K.F.; Brink, K.; Handa, A.K.; Herrmann, K.M.; Conn, E.E. Light and fungal elicitor Induce 3-Deoxy-d-arabino-heptulosonate 7-Phosphate synthase mRNA in suspension cultured cells of parsley (Petroselinum crispum L.). Plant Physiol. 1992, 98, 761–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salas, R.A.; Scott, R.C.; Dayan, F.E.; Burgos, N.R. EPSPS gene amplification in glyphosate-resistant Italian ryegrass (Lolium perenne ssp. multiflorum) from Arkansas. J. Agric. Food Chem. 2012, 68, 1223–1230. [Google Scholar] [CrossRef]
- Huang, L.; Jiang, X.; Li, B.; Li, Y.; Zhang, X. Transformation of EPSP synthetase gene from Allium macrostemon bunge into tobacco and improvement of resistance in transgenic plants to glyphosate. Acta Agron. Sin. 2009, 35, 855–860. [Google Scholar] [CrossRef]
- Zhong, S.; Chen, Z.; Han, J.; Zhao, H.; Liu, J.; Yu, Y. Suppression of chorismate synthase, which is localized in chloroplasts and peroxisomes, results in abnormal flower development and anthocyanin reduction in petunia. Sci. Rep. 2020, 10, 10846. [Google Scholar] [CrossRef]
- Oliveira, J.S.; Pinto, C.A.; Basso, L.A.; Santo, D.S. Cloning and overexpression in soluble form of functional shikimate kinase and 5-enolpyruvylshikimate 3-phosphate synthase enzymes from Mycobacterium tuberculosis. Protein Expr. Purif. 2001, 22, 430–435. [Google Scholar] [CrossRef] [Green Version]
- Whipp, M.J.; Pittard, A.J. A reassessment of the relationship between aroK- and aroL-encoded shikimate kinase enzymes of Escherichia coli. J. Bacteriol. 1995, 177, 1627–1629. [Google Scholar] [CrossRef] [Green Version]
- Minton, N.P.; Whitehead, P.J.; Atkinson, T.; Gilbert, H.J. Nucleotide sequence of an Erwinia chrysanthemi gene encoding shikimate kinase. Nucleic Acids Res. 1989, 17, 1769. [Google Scholar] [CrossRef]
- Defeyter, R.C.; PIittard, J. Purification and properties of shikimate kinase II from Escherichia coli K-12. J. Bacteriol. 1986, 165, 331–333. [Google Scholar] [CrossRef] [Green Version]
- Duncan, K.; Edwards, R.M.; Coggins, J.R. The Saccharomyces cerevisiae ARO1 gene. An example of the co-ordinate regulation of five enzymes on a single biosynthetic pathway. FEBS Lett. 1988, 241, 83–88. [Google Scholar] [CrossRef]
- Kinghorn, J.R.; Hawkins, A.R. Cloning and expression in Escherichia coli K-12 of the biosynthetic dehydroquinase function of the arom cluster gene from the eucaryote, Aspergillus nidulans. Mol. Genet. Genomics 1982, 186, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Schaller, A.; Leibinger, U.; Boll, W.; Amrhein, N. The in-vitro synthesized tomato shikimate kinase precursor is enzymatically active and is imported and processed to the mature enzyme by chloroplasts. Plant J. 1992, 2, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Koji, K.; Takuya, K.; Mitsuru, A.; Yasuko, I.; Kyo, W.; Yuzuru, T. Identification of three shikimate kinase genes in rice: Characterization of their differential expression during panicle development and of the enzymatic activities of the encoded proteins. Planta 2005, 222, 438–447. [Google Scholar] [CrossRef]
- Fucile, G.; Falconer, S.; Christendat, D. Evolutionary diversification of plant shikimate kinase gene duplicates. PLoS Genet. 2008, 4, e1000292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, C.L.; Danneel, H.J.; Schultz, G.; Buchanan, B.B. Shikimate kinase from spinach chloroplasts: Purification, characterization, and regulatory function in aromatic amino Acid biosynthesis. Plant Physiol. 1990, 93, 758–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mattioli, R.; Francioso, A.; Mosca, L.; Silva, P. Anthocyanins: A comprehensive review of their chemical properties and health effects on cardiovascular and neurodegenerative diseases. Molecules 2020, 25, 3809. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Alfenito, M.; Rea, P.A.; Walbot, V.; Dixon, R.A. Vacuolar uptake of the phytoalexin medicarpin by the glutathione conjugate pump. Phytochemistry 1997, 45, 689–693. [Google Scholar] [CrossRef]
- Klein, M.; Weissenböck, G.; Dufaud, A.; Gaillard, C.; Kreuz, K.; Martinoia, E. Different energization mechanisms drive the vacuolar uptake of a flavonoidglucoside and a herbicide glucoside. J. Biol. Chem. 1996, 271, 29666–29671. [Google Scholar] [CrossRef] [Green Version]
- Tohge, T.; Watanabe, M.; Hoefgen, R.; Fernie, A.R. Shikimate and phenylalanine biosynthesis in the green lineage. Front. Plant Sci. 2013, 4, 62. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Liu, J.; Wei, Q.; Wang, R.; Yang, W.; Ma, Y.; Chen, G.; Yu, Y. Proteomes and Ubiquitylomes Analysis Reveals the Involvement of Ubiquitination in Protein Degradation in Petunias. Plant Physiol. 2017, 173, 668–687. [Google Scholar] [CrossRef]
- Zhao, H.; Zhong, S.; Sang, L.; Zhang, X.; Chen, Z.; Wei, Q.; Chen, G.; Liu, J.; Yu, Y. Corrigendum: PaACL silencing accelerates flower senescence and changes proteome to maintain metabolic homeostasis in Petunia hybrida. J. Exp. Bot. 2020, 71, 5113. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Meng, J.; Liu, J.; Ding, B.; Tan, T.; Wei, Q.; Yu, Y. The N1-Methyladenosine Methylome of Petunia mRNA. Plant Physiol. 2020, 183, 1710–1724. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.E.; Saraste, M.; Runswick, M.J.; Gay, N.J. Distantly related sequences in the alpha- and beta-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. EMBO J. 1982, 1, 945–951. [Google Scholar] [CrossRef]
- Krell, T.; Coggins, J.R.; Lapthorn, A.J. The three-dimensional structure of shikimate kinase. J. Mol. Biol. 1998, 278, 983–997. [Google Scholar] [CrossRef]
- Gu, Y.; Reshetnikova, L.; Li, Y.; Wu, Y.; Yan, H.; Singh, S.; Ji, X. Crystal structure of shikimate kinase from Mycobacterium tuberculosis reveals the dynamic role of the LID domain in catalysis. J. Mol. Biol. 2002, 319, 779–789. [Google Scholar] [CrossRef] [PubMed]
- Della-Cioppa, G.; Bauer, S.C.; Klein, B.K.; Shah, D.M.; Fraley, R.T.; Kishore, G.M. Translocation of the precursor of 5-enolpyruvylshikimate-3-phosphate synthase into chloroplasts of higher plants in vitro. Proc. Natl. Acad. Sci. USA 1986, 83, 6873–6877. [Google Scholar] [CrossRef] [Green Version]
- Colquhoun, T.A.; Schimmel, B.C.; Kim, J.Y.; Reinhardt, D.; Cline, K.; Clark, D.G. A petunia chorismate mutase specialized for the production of floral volatiles. Plant J. 2010, 61, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Dal Cin, V.; Tieman, D.M.; Tohge, T.; McQuinn, R.; de Vos, R.C.; Osorio, S.; Schmelz, E.A.; Taylor, M.G.; Smits-Kroon, M.T.; Schuurink, R.C.; et al. Identification of genes in the phenylalanine metabolic pathway by ectopic expression of a MYB transcription factor in tomato fruit. Plant Cell 2011, 23, 2738–2753. [Google Scholar] [CrossRef]
- Maeda, H.; Shasany, A.K.; Schnepp, J.; Orlova, I.; Taguchi, G.; Cooper, B.R.; Rhodes, D.; Pichersky, E.; Dudareva, N. RNAi suppression of Arogenate Dehydratase1 reveals that phenylalanine is synthesized predominantly via the arogenate pathway in petunia petals. Plant Cell 2010, 22, 832–849. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, L.; Maury, S.; Martz, F.; Geoffroy, P.; Legrand, M. Purification, cloning, and properties of an acyltransferase controlling shikimate and quinate ester intermediates in phenylpropanoid metabolism. J. Biol. Chem. 2003, 278, 95–103. [Google Scholar] [CrossRef]
- Jones, J.D.; Henstrand, J.M.; Handa, A.K.; Herrmann, K.M.; Weller, S.C. Impaired wound induction of 3-deoxy-D-arabino-heptulosonate-7-phosphate (DAHP) synthase and altered stem development in transgenic potato plants expressing a DAHP synthase antisense construct. Plant Physiol. 1995, 108, 1413–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, N.G.; Yamamoto, E. Lignin: Occurrence, biogenesis and biodegradation. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1990, 41, 455–496. [Google Scholar] [CrossRef] [PubMed]
- Goers, S.K.; Jensen, R.A. The differential allosteric regulation of two chorismate-mutase isoenzymes of Nicotiana silvestris. Planta 1984, 162, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Amrhein, N. Molecular organization of the shikimate pathway in higher plants. Phytochemistry 1995, 39, 737–749. [Google Scholar] [CrossRef]
- Swinney, D.C.; Mak, A.Y.; Barnett, J.; Ramesha, C.S. Differential allosteric regulation of prostaglandin H synthase 1 and 2 by arachidonic acid. J. Biol. Chem. 1997, 272, 12393–12398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, Y.; Liu, J.; Huang, F.; Guan, J.; Zhong, S.; Tang, N.; Zhao, J.; Yang, W.; Yu, Y. PhGRL2 protein, interacting with PhACO1, is Involved in flower senescence in the petunia. Mol. Plant 2014, 7, 1384–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Liu, J.; Tan, Y.; Zhong, S.; Tang, N.; Chen, G.; Yu, Y. Functional characterization of PhGR and PhGRL1 during flower senescence in the petunia. Plant Cell Rep. 2015, 34, 1561–1568. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Liu, J.; Li, J.; Wang, H.; Fu, Z.; Liu, J.; Yu, Y. Identification and expression analysis of ERF transcription factor genes in petunia during flower senescence and in response to hormone treatments. J. Exp. Bot. 2011, 62, 825–840. [Google Scholar] [CrossRef]
- Locatelli, F.; Vannini, C.; Magnani, E.; Coraggio, I.; Bracale, M. Efficiency of transient transformation in tobacco protoplasts is independent of plasmid amount. Plant Cell Rep. 2003, 21, 865–871. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, E45. [Google Scholar] [CrossRef]
- Mallona, I.; Lischewski, S.; Weiss, J.; Hause, B.; Egea-Cortines, M. Validation of reference genes for quantitative real-time PCR during leaf and flower development in Petunia hybrida. BMC Plant Biol. 2010, 10, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broderick, S.R.; Jones, M.L. An optimized protocol to increase virus-induced gene silencing efficiency and minimize viral symptoms in petunia. Plant Mol. Biol. Rep. 2014, 32, 219–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spitzer-Rimon, B.; Farhi, M.; Albo, B.; Cna’Ani, A.; Zvi, M.M.B.; Masci, T.; Edelbaum, O.; Yu, Y.; Shklarman, E.; Ovadis, M.; et al. The R2R3-MYB-like regulatory factor EOBI, acting downstream of EOBII, regulates scent production by activating ODO1 and structural scent-related genes in petunia. Plant Cell 2012, 24, 5089–5105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambrook, J.; Maniatis, T.; Fritsch, E. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratories: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Ai, T.N.; Naing, A.H.; Arun, M. Sucrose-induced anthocyanin accumulation in vegetative tissue of Petunia plants requires anthocyanin regulatory transcription factors. Plant Sci. 2016, 252, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Mehrtens, F.; Kranz, H.; Bednarek, P.; Weisshaar, B. The Arabidopsis transcription factor MYB12 is a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol. 2005, 138, 1083–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Song, J. Analysis of widely targeted metabolites of the euhalophyte Suaeda salsa under saline conditions provides new insights into salt tolerance and nutritional value in halophytic species. BMC Plant Biol. 2019, 19, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Gong, L.; Guo, Z.; Wang, W.; Zhang, H.; Liu, X.; Yu, S.; Xiong, L.; Luo, J. A novel Integrated method for large-scale detection, Identification, and quantification of widely targeted metabolites: Application in the study of rice metabolomics. Mol. Plant 2013, 6, 1769–1780. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pTRV2-GFP | pTRV2-PhSK | pTRV2-PhSK/ pTRV2-GFP (%) | |
|---|---|---|---|
| Length of pedicels (cm) | 2.71 ± 0.67 | 1.58 ± 0.47 * | 58.30 |
| Length of sepals (cm) | 2.05 ± 0.20 | 1.69 ± 0.20 * | 82.43 |
| Width of sepals (cm) | 0.58 ± 0.09 | 0.56 ± 0.16 | 96.55 |
| Length–width ratio of sepals (cm) | 26.57 ± 2.88 | 35.11 ± 4.49 * | 132.14 |
| Circumference of corolla tube (cm) | 2.24 ± 0.19 | 1.85 ± 0.25 * | 82.59 |
| Length of corolla tube (cm) | 3.12 ± 0.09 | 3.23 ± 0.27 | 103.53 |
| Diameter of corollas (cm) | 6.73 ± 0.37 | 5.76 ± 0.59 * | 85.59 |
| Length of internodes (cm) | 2.61 ± 0.55 | 1.52 ± 0.37 * | 58.24 |
| Compounds | VIP | Fold Change | Type |
|---|---|---|---|
| Cyanidin-3-O-(2″-O-glucosyl)rutinoside | 1.09 | 0.46 | down |
| Cyanidin-3-O-(6″-O-malonyl)sophoroside-5-O-glucoside | 1.04 | 0.47 | down |
| Cyanidin-3,3′-di-O-glucoside-7-O-(6″-O-caffeoyl)glucoside | 1.14 | 0.48 | down |
| Cyanidin-3-O-glucoside (Kuromanin) | 1.09 | 0.48 | down |
| Cyanidin-3-O-sophoroside-5-O-glucoside | 1.12 | 0.48 | down |
| Delphinidin-3-O-rutinoside-7-O-glucoside | 1.14 | 0.49 | down |
| Petunidin-3-O-(6″-O-feruloyl)rutinoside-5-O-glucoside | 1.09 | 2.02 | up |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, J.; Zhong, S.; Long, Y.; Guo, J.; Yu, Y.; Liu, J. Shikimate Kinase Plays Important Roles in Anthocyanin Synthesis in Petunia. Int. J. Mol. Sci. 2022, 23, 15964. https://doi.org/10.3390/ijms232415964
Yuan J, Zhong S, Long Y, Guo J, Yu Y, Liu J. Shikimate Kinase Plays Important Roles in Anthocyanin Synthesis in Petunia. International Journal of Molecular Sciences. 2022; 23(24):15964. https://doi.org/10.3390/ijms232415964
Chicago/Turabian StyleYuan, Junwei, Shiwei Zhong, Yu Long, Jingling Guo, Yixun Yu, and Juanxu Liu. 2022. "Shikimate Kinase Plays Important Roles in Anthocyanin Synthesis in Petunia" International Journal of Molecular Sciences 23, no. 24: 15964. https://doi.org/10.3390/ijms232415964