
Characteristics of PoVIN3, a Key Gene of Vernalization Pathway, Affects Flowering Time
 ,
,
Abstract
:1. Introduction
2. Results
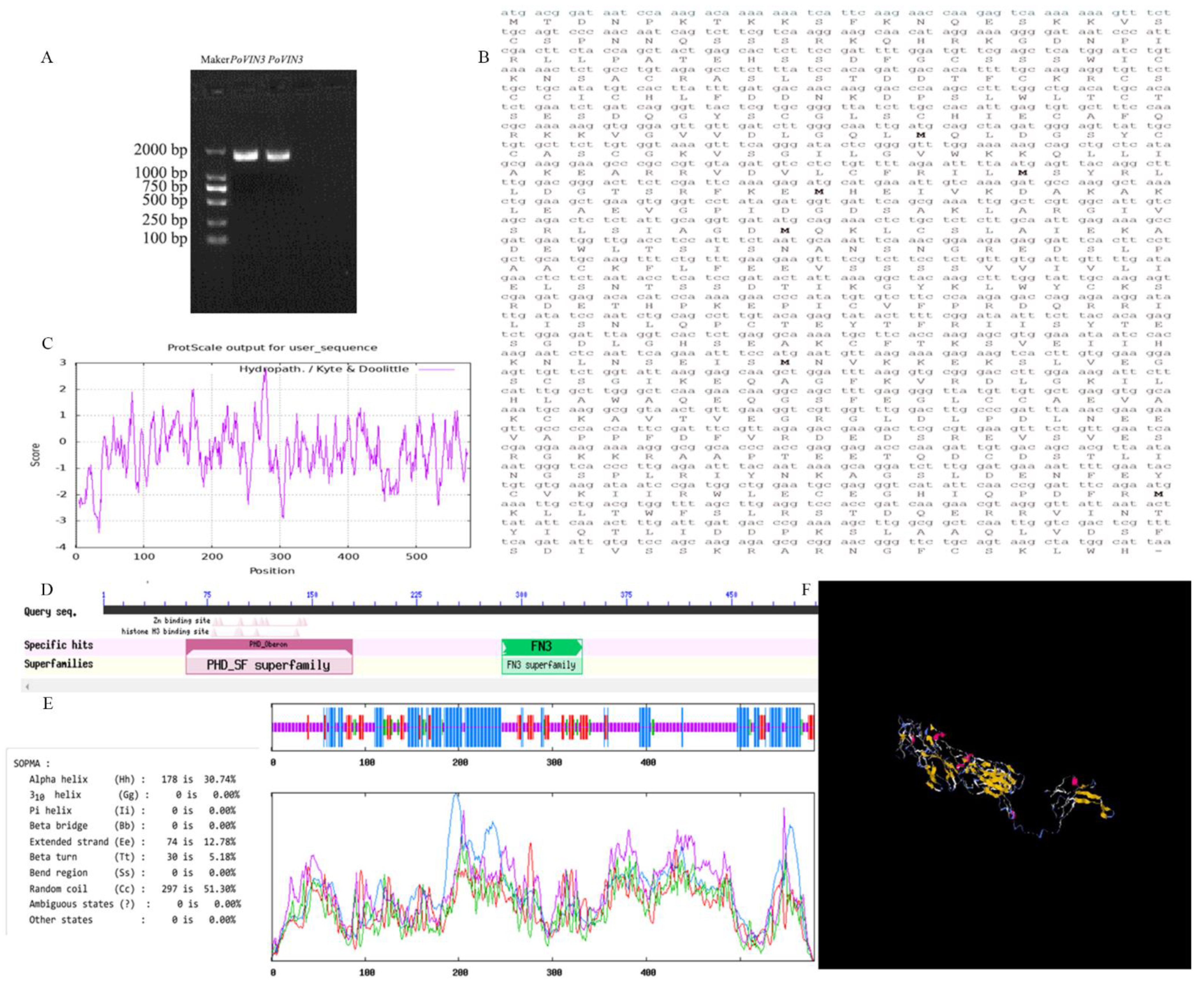
2.1. Cloning and Sequence Analysis of PoVIN3
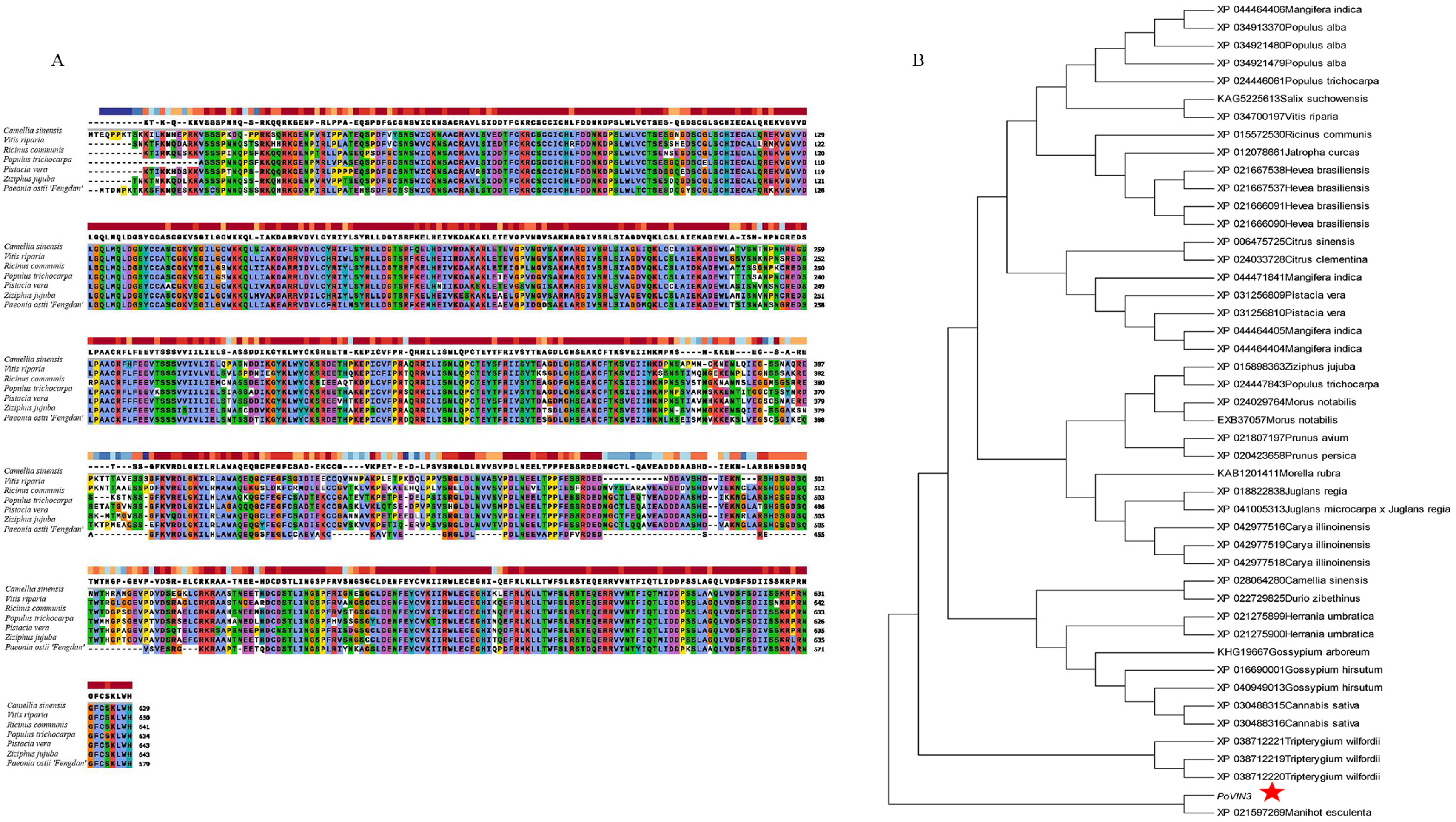
2.2. Phylogenetic Tree Analysis
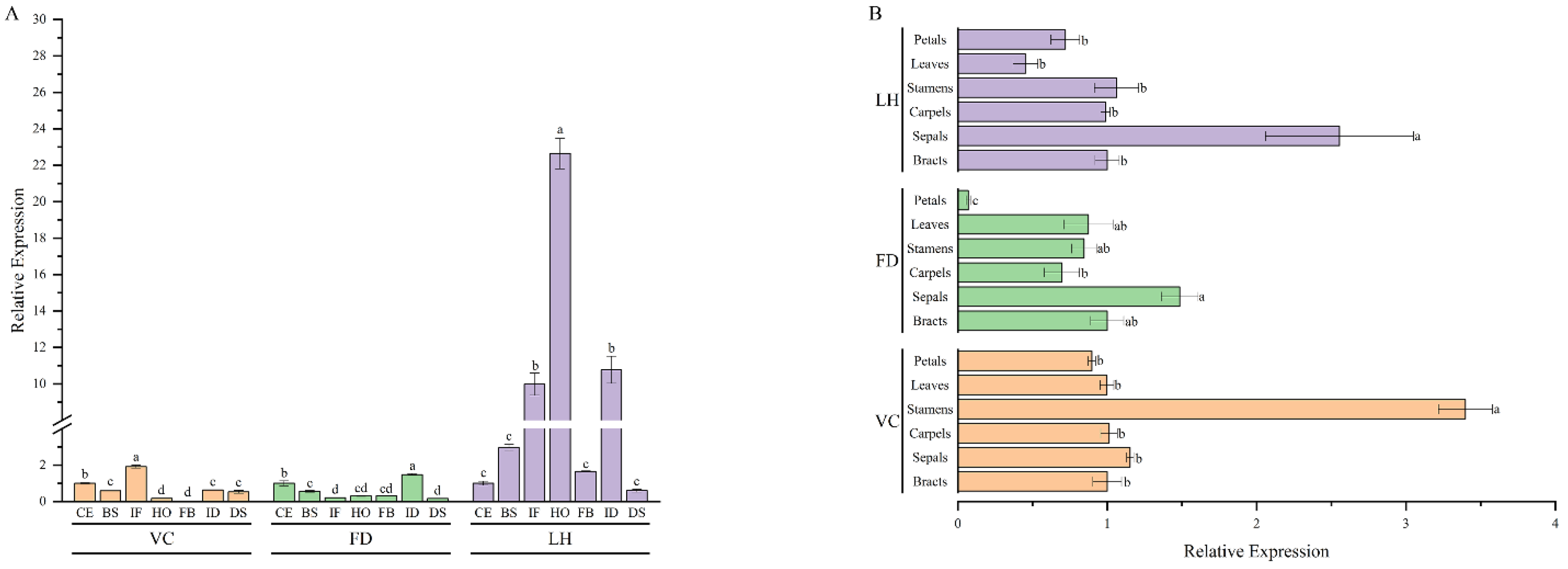
2.3. Expression Pattern of PoVIN3
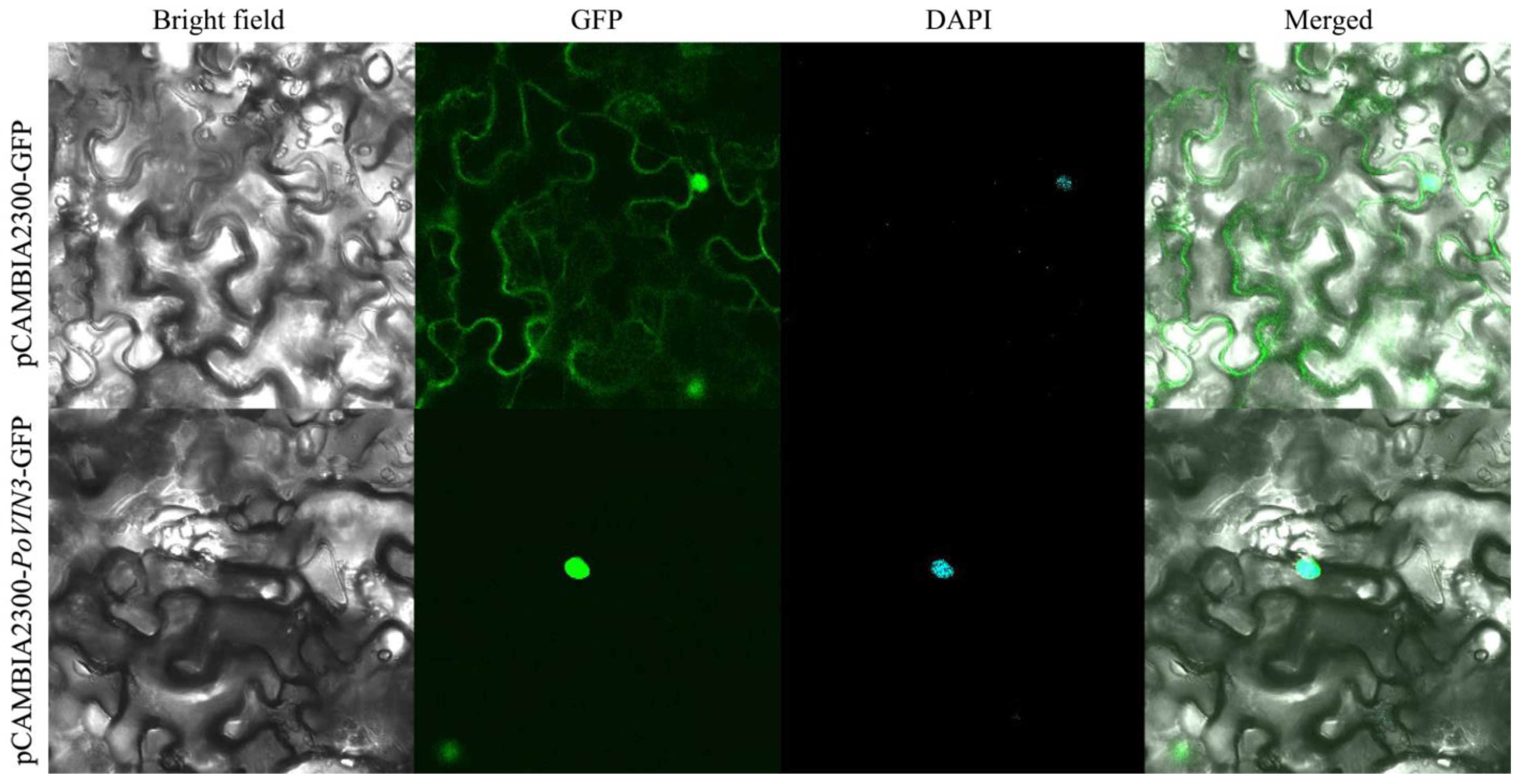
2.4. Subcellular Localization
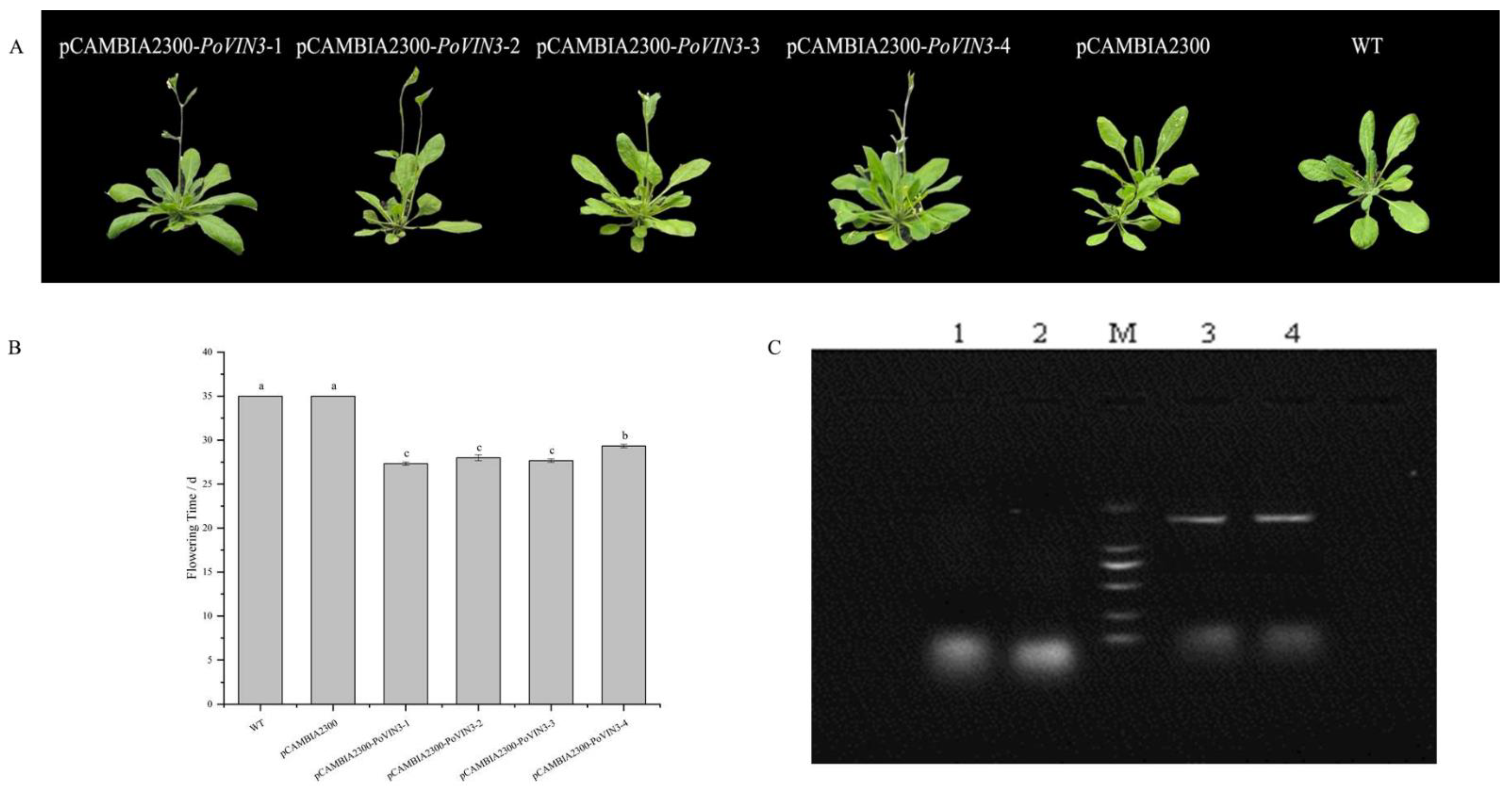
2.5. Ectopic Expression of PoVIN3 Accelerate Flowering in Arabidopsis Thaliana
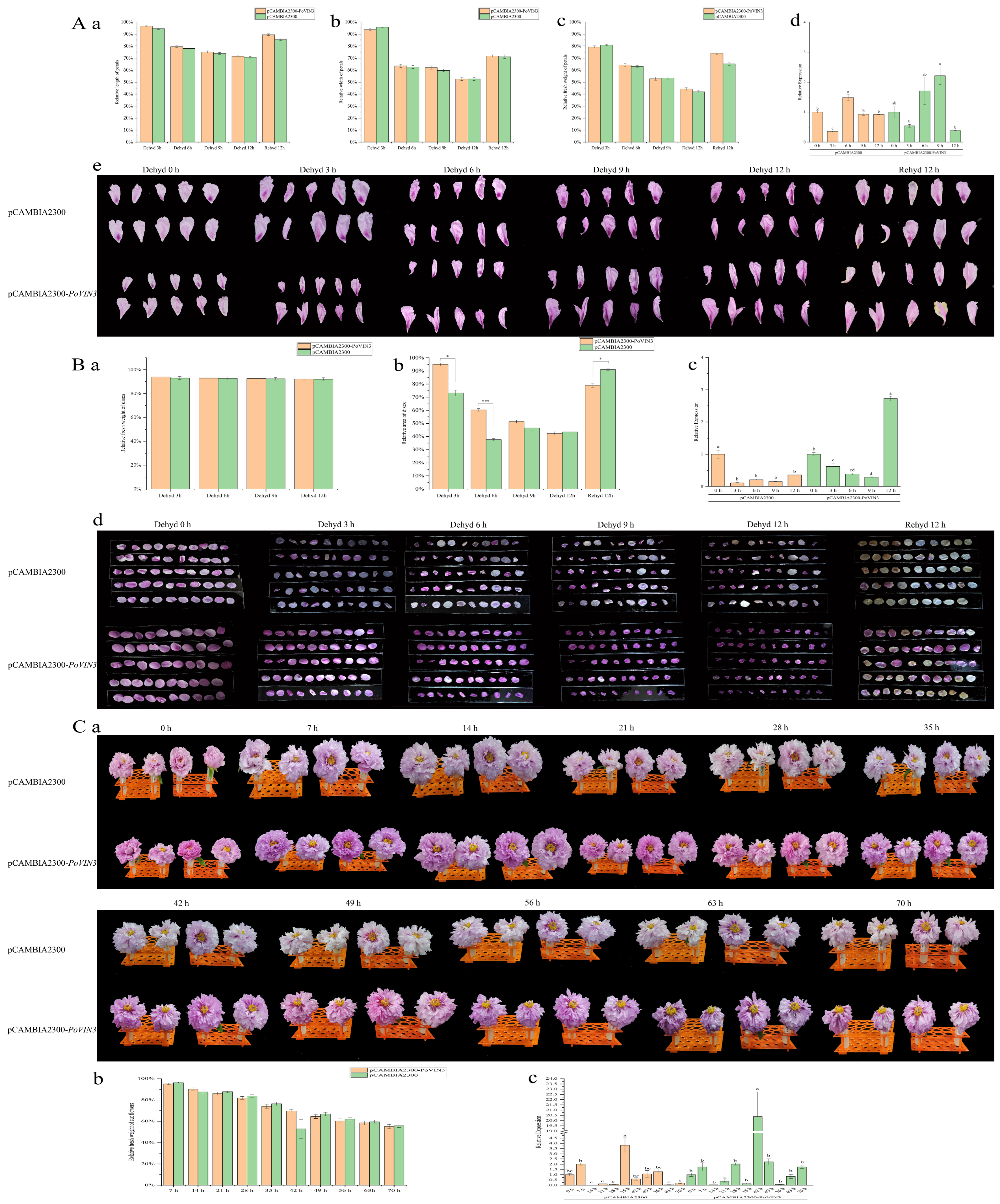
2.6. pCAMBIA2300-PoVIN3 Overexpressed Tree Peony Petals
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Nucleic Acid Extraction and cDNA Synthesis
4.3. Cloning of PoVIN3 and Construction of Expression Vectors
4.4. Bioinformatics Analysis of PoVIN3
4.5. Analysis of Subcellular localization
4.6. Arabidopsis Thaliana Transformation and Analysis of Flowering Time
4.7. Establishment of Transient Expression System of Tree Peony
4.8. Quantitative Real−Time PCR Analysis of PoVIN3
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lv, S.Z.; Cheng, S.; Wang, Z.Y.; Li, S.M.; Jin, X.; Lan, L.; Yang, B.; Yu, K.; Ni, X.M.; Li, N.; et al. Draft genome of the famous ornamental plant Paeonia suffruticosa. Ecol. Evol. 2020, 10, 4518–4530. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Sun, M.; Li, S.S.; Chen, Q.H.; Teixeira da Silva, J.A.; Wang, A.J.; Yu, X.N.; Wang, L.S. Germplasm resources and genetic breeding of Paeonia: A systematic review. Hortic. Res. 2020, 7, 107. [Google Scholar] [CrossRef]
- Li, J.J.; Zhang, X.F.; Zhao, X.Q. Tree Peony of Chinese; Encyclopedia of China Publishing House: Beijing, China, 2011; p. 340. [Google Scholar]
- Hou, X.G.; Guo, Q.; Wei, W.Q. Screening of Genes Related to Early and Late Flowering in Tree Peony Based on Bulked Segregant RNA Sequencing and Verification by Quantitative Real-Time PCR. Molecules 2018, 23, 689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, F.Y. Advances in the breeding of tree peonies and a cultivar system for the cultivar group. Int. J. Plant Breed. 2007, 2, 89–104. [Google Scholar]
- Wang, S.L.; Gao, J.; Xue, J.Q. De novo sequencing of tree peony (Paeonia suffruticosa) transcriptome to identify critical genes involved in flowering and floral organ development. BMC Genom. 2019, 20, 572. [Google Scholar] [CrossRef]
- Michaels, S.D.; Amasino, R.M. FLOWERING LOCUS C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell 1999, 11, 949–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hepworth, S.R.; Valverde, F.; Ravenscroft, D. Antagonistic regulation of flowering time gene SOC1 by CONSTANS and FLC via separate promoter motifs. EMBO J. 2002, 21, 4327–4337. [Google Scholar] [CrossRef] [Green Version]
- Helliwell, C.A.; Wood, C.C.; Robertson, M. The Arbidopsis FLC protein interacts directly in vivo with SOC1 and FT chromatin and is a part of a high-molecular-weight protein complex. Plant J. Cell Mol. Biol. 2006, 46, 183–192. [Google Scholar] [CrossRef]
- Fedorenko, O.M.; Topchieva, L.V.; Zaretskaya, M.V. Changes in FLC and VIN3 expression during vernalization of arabidops isthaliana plants from northern natural populations. Russ. J. Genet. 2019, 55, 865–871. [Google Scholar] [CrossRef]
- Kim, S.; Choi, K.; Park, C.; Hwang, H.J.; Lee, I. SUPPRESSOR OF FRIGIDA4, encoding a C2H2-Type zinc finger protein, represses flowering by transcriptional activation of Arabidopsis FLOWERING LOCUS C. Plant Cell 2006, 18, 2985–2998. [Google Scholar] [CrossRef] [Green Version]
- Bastow, R.; Mylne, J.S.; Lister, C.; Lippman, Z.; Martienssen, R.A.; Dean, C. Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 2004, 427, 164–167. [Google Scholar] [CrossRef] [PubMed]
- Sung, S.; Amasino, R.M. Vernalization in Arabidopsis thaliana is mediated by the PHD finger protein VIN3. Nature 2004, 427, 159–164. [Google Scholar] [CrossRef]
- Trevaskis, B.; Hemming, M.N.; Dennis, E.S.; James Peacock, W. The molecular basis of vernalization induced flowering in cereals. Trends Plant Sci. 2007, 12, 352–357. [Google Scholar] [CrossRef]
- Fu, D.; Dunbar, M.; Dubcovsky, J. Wheat VIN3-like PHD finger genes are up-regulated by vernalization. Mol. Genet. Genom. 2007, 277, 301–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, S.; Schmitz, R.J.; Amasino, R.M. A PHD finger protein involved in both the vernalization and photoperiod pathways in Arabidopsis. Genes Dev. 2007, 20, 3244–3248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sung, S.; Schmitz, R.; Amasino, R. The Role of VIN3-like genes in environmentally induced epigenetic regulation of flowering. Plant Signal. Behav. 2007, 2, 127–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, Y.; Ma, Q. SKB1-mediated symmetric dimethylation of histone H4R3 controls flowering time in Arabidopsis. EMBO J. 2007, 26, 1934–1941. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, R.J.; Sung, S.; Amasino, R.M. Histone arginine methylation is required for vernalization-induced epigenetic silencing of FLC in winter-annual Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2008, 105, 411–416. [Google Scholar] [CrossRef] [Green Version]
- Bouché, F.; Woods, D.P.; Amasino, R.M. Winter Memory throughout the Plant Kingdom: Different Paths to Flowering. Plant Physiol. 2017, 173, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Zhao, R.Q.; Hu, Y.; Jiang, X.M.; Yu, X.H. Cloning and Expression Analysis of BoVIN3 from Cabbage. Acta Hortic. Sin. 2012, 39, 1099–1106. [Google Scholar]
- Chen, G.P.; Hu, G.L.; Hu, Z.L.; Li, Y.; Gu, F. Cloning and Expression of Vernalization-related Genes in Purple Flowering Stalk (Brassica compestris ssp. chinensis var. pupurea Hort). Life Sci. Res. 2010, 14, 027–033. [Google Scholar] [CrossRef]
- Kaadige, M.R.; Ayer, D.E. The polybasic region that follows the plant homeodomain zinc finger 1 of Pf1 is necessary and sufficient for specific phosphoinositide binding. J. Biol. Chem. 2006, 281, 28831–28836. [Google Scholar] [CrossRef]
- Ma, H.H.; Fang, C.L.; Cao, P.Y. Plant homeodomain (PHD domain)—The code reading of histone code. Prog. Biochem. Biophys. 2008, 35, 625–630. [Google Scholar]
- Shen, H.R.; Xiao, D.; Hou, X.L. Cloning and expression analysis of flowering time candidate gene BcVIL1 in non-heading Chinese cabbage. J. Nanjing Agric. Univ. 2018, 41, 825–831. [Google Scholar]
- Jeong, G.; Jeon, M.; Shin, J.; Lee, I. HEAT SHOCK TRANSCRIPTION FACTOR B2b acts as a transcriptional repressor of VIN3, a gene induced by long-term cold for flowering. Sci. Rep. 2022, 12, 10963. [Google Scholar] [CrossRef]
- Kim, D.H.; Sung, S. The Binding Specificity of the PHD-Finger Domain of VIN3 Moderates Vernalization Response. Plant Physiol. 2017, 173, 1258–1268. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.S.; Antoniou-Kourounioti, R.L.; Calder, G.; Dean, G.; Howard, M. Temperature-dependent growth contributes to long-term cold sensing. Nature 2020, 583, 825–829. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.J.; Huang, G.D.; Luo, S.X.; Zhou, J.A.; Mo, Y.L.; Li, R.W.; Zhao, Y.; Zhang, Y.; Song, E.L.; Ning, L. Transcriptome of Mangifera indica L. in two different flower bud differentiation stages. J. South. Agric. 2018, 49, 1257–1264. [Google Scholar] [CrossRef]
- Zhao, S.Q.; Hu, J.; Guo, L.B.; Qian, Q.; Xue, H.W. Rice leaf inclination2, a VIN3-like protein, regulates leaf angle through modulating cell division of the collar. Cell Res. 2010, 20, 935–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, S.N.; Finnegan, E.J.; Dennis, E.S.; Peacock, W.J.; Trevaskis, B. Vernalization-induced flowering in cereals is associated with changes in histone methylation at the VERNALIZATION1 gene. Proc. Nat. Acad. Sci. USA 2009, 106, 8386–8391. [Google Scholar] [CrossRef] [Green Version]
- Almutairi, Z.M.; Sadder, M.T. Cloning and expression profiling polycomb gene VERNALIZATION INSENSITIVE 3 in tomato. Biol. Plant. 2014, 58, 419–426. [Google Scholar] [CrossRef]
- Zhao, Z.H.; Zeng, Q.; Zhao, S.Q. The Molecular Mechanism of Vernalization in Plants. Chin. Bull. Bot. 2006, 23, 60–67. [Google Scholar] [CrossRef]
- Wang, F.; Bian, X.C.; Liu, C.W.; Xu, R.C.; Wu, C.F. Advances in molecular mechanism of plant vernalization. Xiandai Hortic. 2022, 45, 43–45+48. [Google Scholar] [CrossRef]
- Chang, Y.T.; Hu, T.; Zhang, W.B.; Zhou, L.; Wang, Y.; Jiang, Z.H. Transcriptome profiling for floral development in reblooming cultivar ‘High Noon’ of Paeonia suffruticosa. Sci. Data 2019, 6, 217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Guo, D.L.; Guo, L.L.; Guo, Q.; Wang, H.F.; Hou, X.G. Construction of a high-density genetic map and QTLs mapping with GBS from the interspecific F1 population of P. ostii ‘Fengdan Bai’ and P. suffruticosa ‘Xin Riyuejin. Sci. Hortic. 2019, 246, 190–200. [Google Scholar] [CrossRef]
- Ma, J.; Wang, J.; Wang, Q.; Linxue Shang, L.X.; Zhao, Y.; Zhang, G.Z.; Ma, Q.Q.; Hong, S.D.; Gu, G.H. Physiological and transcriptional responses to heat stress and functional analyses of PsHSPs in tree peony (Paeonia suffruticosa). Front. Plant Sci. 2022, 13, 926900. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, T.; Wu, Y.; Tao, J. Identification and functional verification of PlFT gene associated with flowering in herbaceous peony based on transcriptome analysis. Ornam. Plant Res. 2021, 1, 7. [Google Scholar] [CrossRef]
- Duan, Y.H.; Yan, Y.Y.; Chen, L.T.; Li, Q.; Zhang, D.M.; Sun, Z.W.; Zhang, Y.; Ma, C.Y.; Wang, S.F. Cloning and Functional Analysis of GhMYB44 Related to Flowering Time in Gossypium hirsutum. J. Agric. Sci. Technol. 2020, 22, 29–38. [Google Scholar] [CrossRef]
- Shu, Q.Y.; Zhu, J.; Men, S.Q.; Hao, Q.; Wang, Q.Y.; Liu, Z.A.; Zeng, X.L.; Wang, L.S. Establishing virus induced gene silencing (VIGS) system in tree peony using PsUFGT genes. Acta Hortic. Sin. 2018, 45, 168–176. [Google Scholar] [CrossRef]
- Zhang, J.; Tian, J.; Tai, D.Q.; Li, K.T.; Zhu, Y.J.; Yao, Y.C. An optimized TRV-based virus-induced gene silencing protocol for Malus crabapple. Plant Cell Tissue Organ Cult. 2016, 126, 499–509. [Google Scholar] [CrossRef]
- Zhang, Y.; Wu, Z.C.; Feng, M.; Chen, J.W.; Qin, M.Z.; Wang, W.Z. The circadian-controlled PIF8-BBX28 module regulates petal senescence in rose flowers by governing mitochondrial ROS homeostasis at night. Plant Cell 2021, 33, 2716–2735. [Google Scholar] [CrossRef]
- Wang, W.Q.; Moss, S.M.A.; Zeng, L.H.; Espley, R.V.; Wang, T.C.; Wang, K.L.; Fu, B.L.; Schwinn, K.E.; Allan, A.C.; Yin, X.R. The red flesh of kiwifruit is differentially controlled by specific activation–repression systems. New Phytol. 2022, 235, 630–645. [Google Scholar] [CrossRef] [PubMed]
- Ogaki, M.; Furuichi, Y.; Kuroda, K.; Chin, P.D.; Mii, M.; Ogawa, Y. Importance of co-cultivation medium pH for successful Agrobacterium-mediated transformation of Lilium × formolongi. Plant Cell Rep. 2008, 27, 699–705. [Google Scholar] [CrossRef]
- Ben-Amar, A.; Cobanov, P.; Buchholz, G.; Mliki, A.; Reustle, G. In planta agro-infiltration system for transient gene expression in grapevine (Vitis spp.). Acta Physiol. Plant. 2013, 35, 3147–3156. [Google Scholar] [CrossRef]
- Carvalho, F.R.; Carvalho, D.S.; O’Grady, K.; Folta, M.K. Agroinfiltration of strawberry fruit-a powerful transient expression system for gene validation. Curr. Plant Biol. 2016, 6, 19–37. [Google Scholar] [CrossRef] [Green Version]
- Verweij, W.; Di Sansebastiano, G.P.; Quattrocchio, F.; Dalessandro, G. Agrobacterium-mediated transient expression of vacuolar GFPs in Petunia leaves and petals. Plant Biosyst. 2008, 142, 343–347. [Google Scholar] [CrossRef]
- Jalal, A.; Ali, Q.; Manghwar, H.; Zhu, D.C. Identification, phylogeny, divergence, structure, and expression analysis of A20/AN1 zinc finger domain containing Stress-Associated Proteins (SAPs) genes in Jatropha curcas L. Genes 2022, 13, 1766. [Google Scholar] [CrossRef]
- Jalal, A.; Sun, J.R.; Chen, Y.Q.; Fan, C.G.; Liu, J.Y.; Wang, C.Q. Evolutionary analysis and functional identification of clock-associated PSEUDO-RESPONSE REGULATOR (PRRs) genes in the flowering regulation of Roses. Int. J. Mol. Sci. 2022, 23, 7335. [Google Scholar] [CrossRef]
- Liu, J.Y.; Wu, S.L.; Sun, J.J.; Sun, J.R.; Wang, H.L.; Cao, X.; Lu, J.; Jalal, A.; Wang, C.Q. Genome-wide analysis reveals widespread roles for RcREM genes in floral organ development in Rosa chinensis. Genomics 2021, 113, 3881–3894. [Google Scholar] [CrossRef]
- Li, Y.Y.; Guo, L.L.; Sun, G.R.; Guo, D.L.; Fan, M.Y.; Zhang, X.X.; Hou, X.G. Analysis of DNA methylation during seed development of Paeonia ostii ‘Feng Dan’ using methylation sensitive amplification polymorphism (MSAP). J. Hortic. Sci. Biotechnol. 2022, 97, 437–455. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′-3′) | Use |
|---|---|---|
| PoVIN3 | F: ATGACGGATAATCCAAAGAC | Cloning the full−length of ORF |
| R: TTAATGCCATAGCTTACTGC | ||
| PoVIN3−qRT | F: CAGGGATACTCGGGGTTT | Quantitative Real−time PCR |
| R: TCTGCTGACAATGCCACG | ||
| Tbulin-α | F: CCGTCAACTTTTCCACCCTG | |
| R: CCTCACTCGGTCAAGGCAGA | ||
| pCAMBIA2300−PoVIN3−GFP | F: GGAGAGGACAGGGTACCATGACGGATAATCCAAAGAC | Subcellular localization |
| R: GATCCCCGGGTACCATGCCATAGCTTACTGCAGAAC | ||
| pCAMBIA2300−PoVIN3 | F: GGAGAGGACAGGGTACCATGACGGATAATCCAAAGAC | |
| R: GGATCCCCGGGTACCTTAATGCCATAGCTTACTGC | ||
| AtVIN3 | F: CCGTAAAGACTGGCGAACAG | Functional verification |
| R: CCGCACGAGTAACCCTGAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Wang, C.; Guo, Q.; Song, C.; Wang, X.; Guo, L.; Hou, X. Characteristics of PoVIN3, a Key Gene of Vernalization Pathway, Affects Flowering Time. Int. J. Mol. Sci. 2022, 23, 14003. https://doi.org/10.3390/ijms232214003
Li Y, Wang C, Guo Q, Song C, Wang X, Guo L, Hou X. Characteristics of PoVIN3, a Key Gene of Vernalization Pathway, Affects Flowering Time. International Journal of Molecular Sciences. 2022; 23(22):14003. https://doi.org/10.3390/ijms232214003
Chicago/Turabian StyleLi, Yuying, Can Wang, Qi Guo, Chengwei Song, Xiaohui Wang, Lili Guo, and Xiaogai Hou. 2022. "Characteristics of PoVIN3, a Key Gene of Vernalization Pathway, Affects Flowering Time" International Journal of Molecular Sciences 23, no. 22: 14003. https://doi.org/10.3390/ijms232214003