
Molecular and Metabolic Insights into Anthocyanin Biosynthesis for Spot Formation on Lilium leichtlinii var. maximowiczii Flower Petals


Abstract
:1. Introduction
2. Results
2.1. Morphology Analysis of Petal Color Transitions and Spots Formation
2.2. Carotenoid Identification and Quantification in Petals
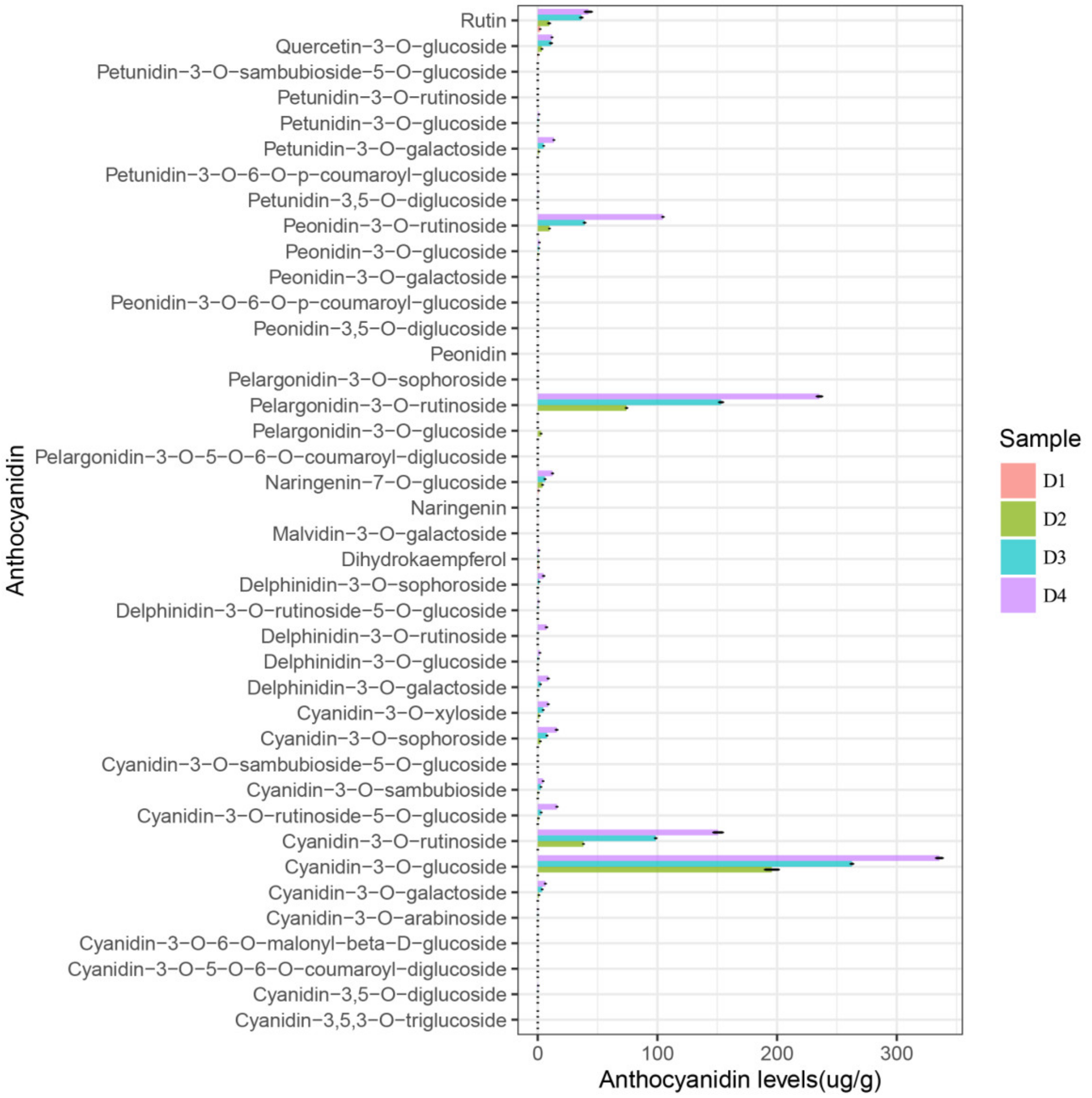
2.3. Anthocyanin Identification and Quantification in Petal Spots
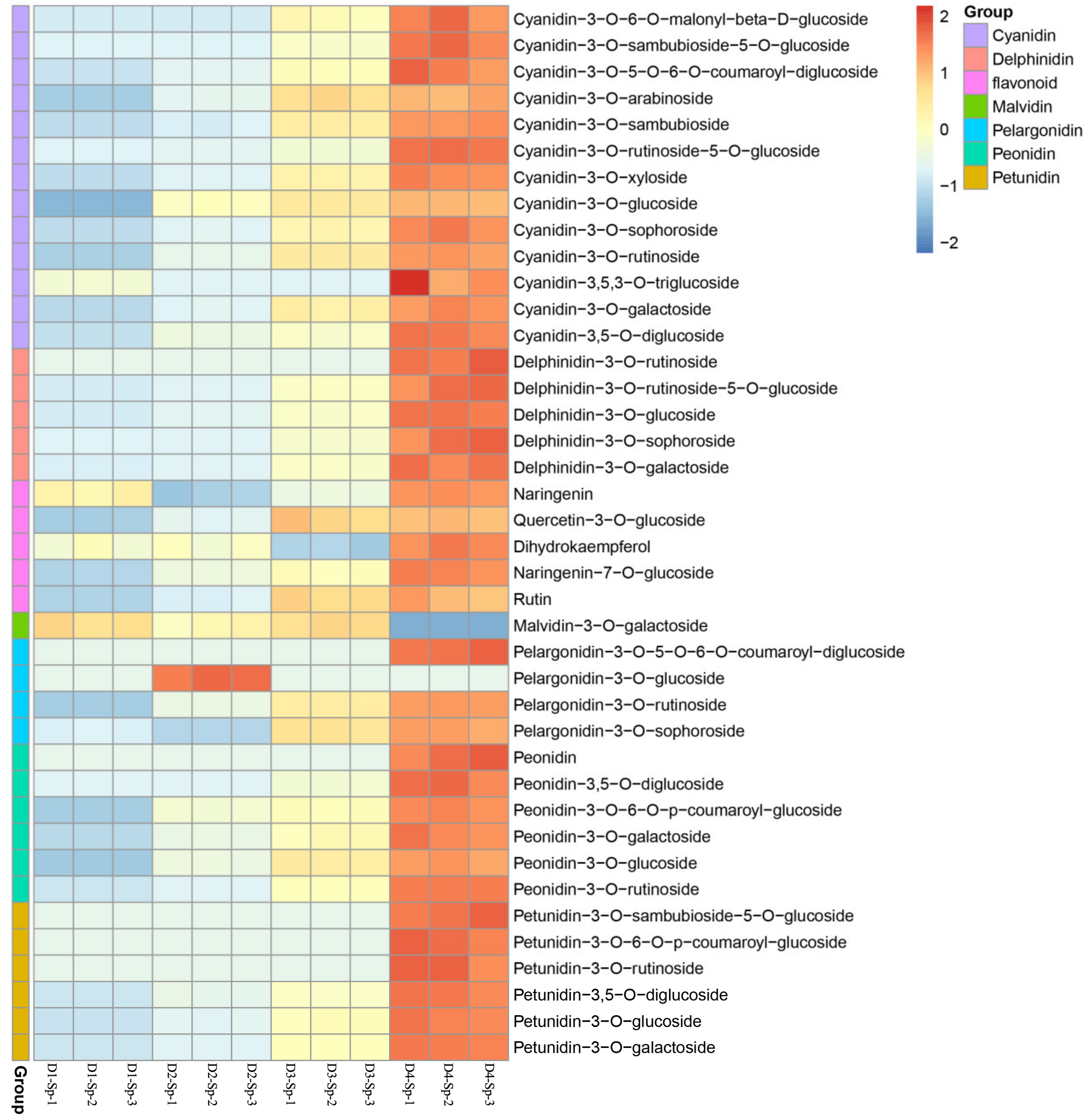
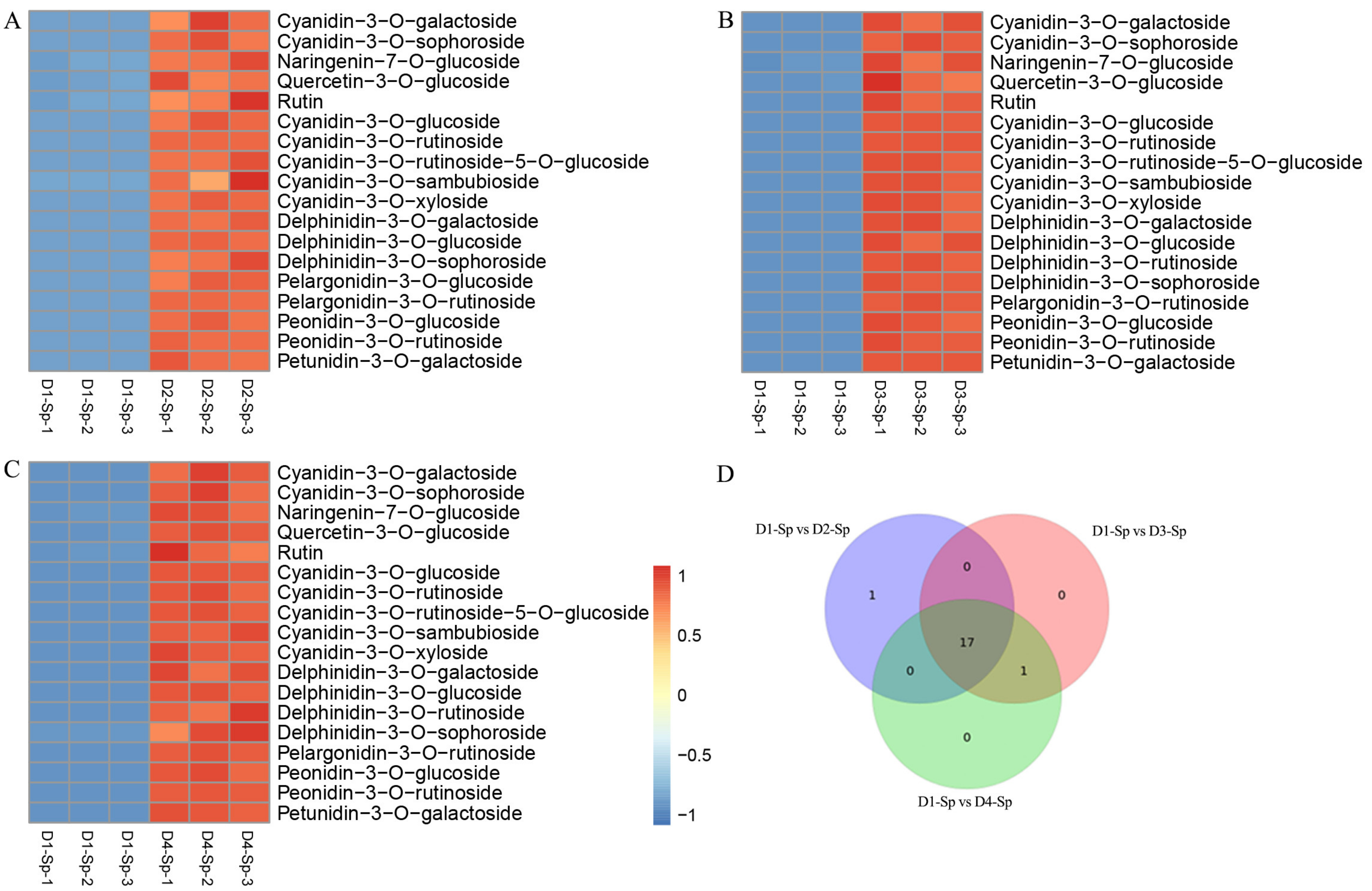
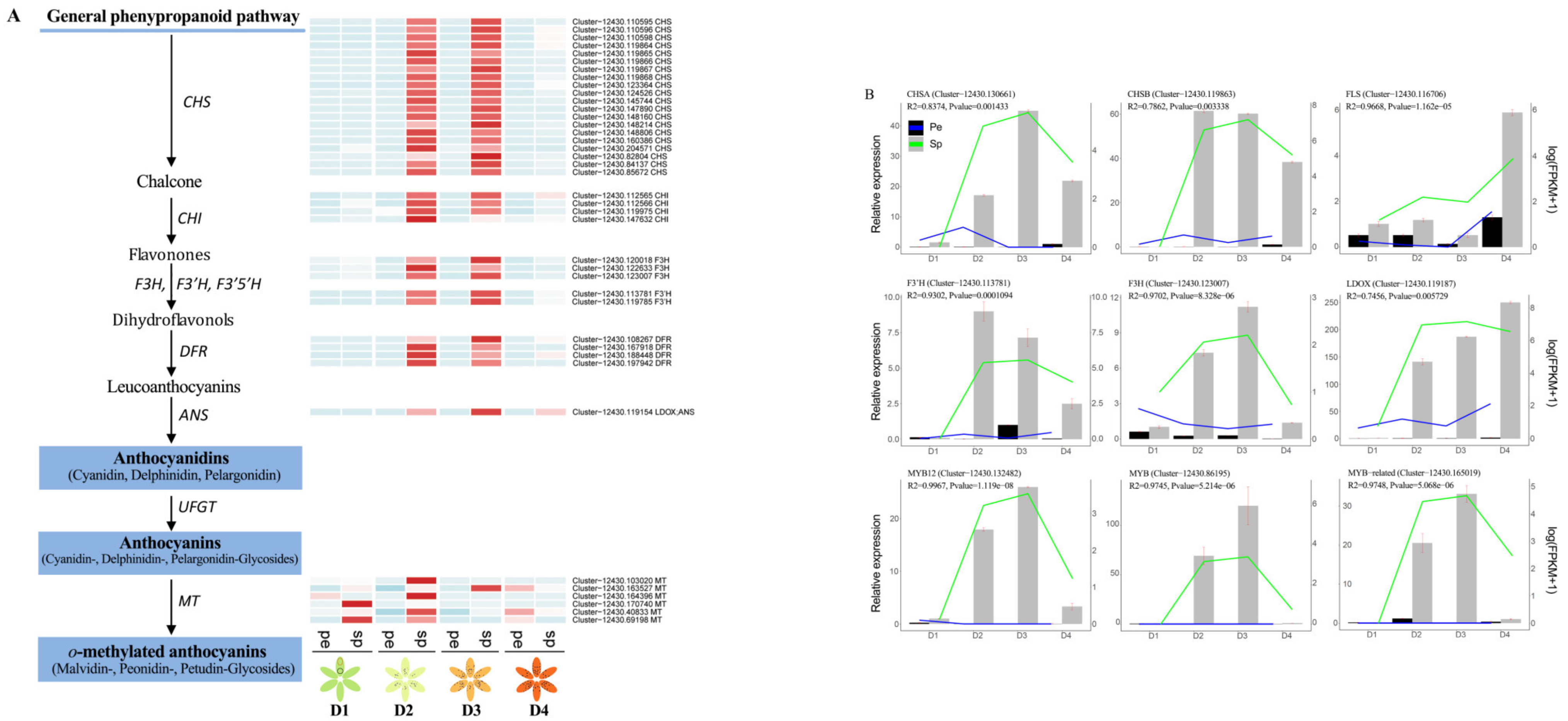
2.4. Differentially Accumulated Anthocyanin Components in Petal Spots
2.5. De Novo Assembly and Gene Function Annotation
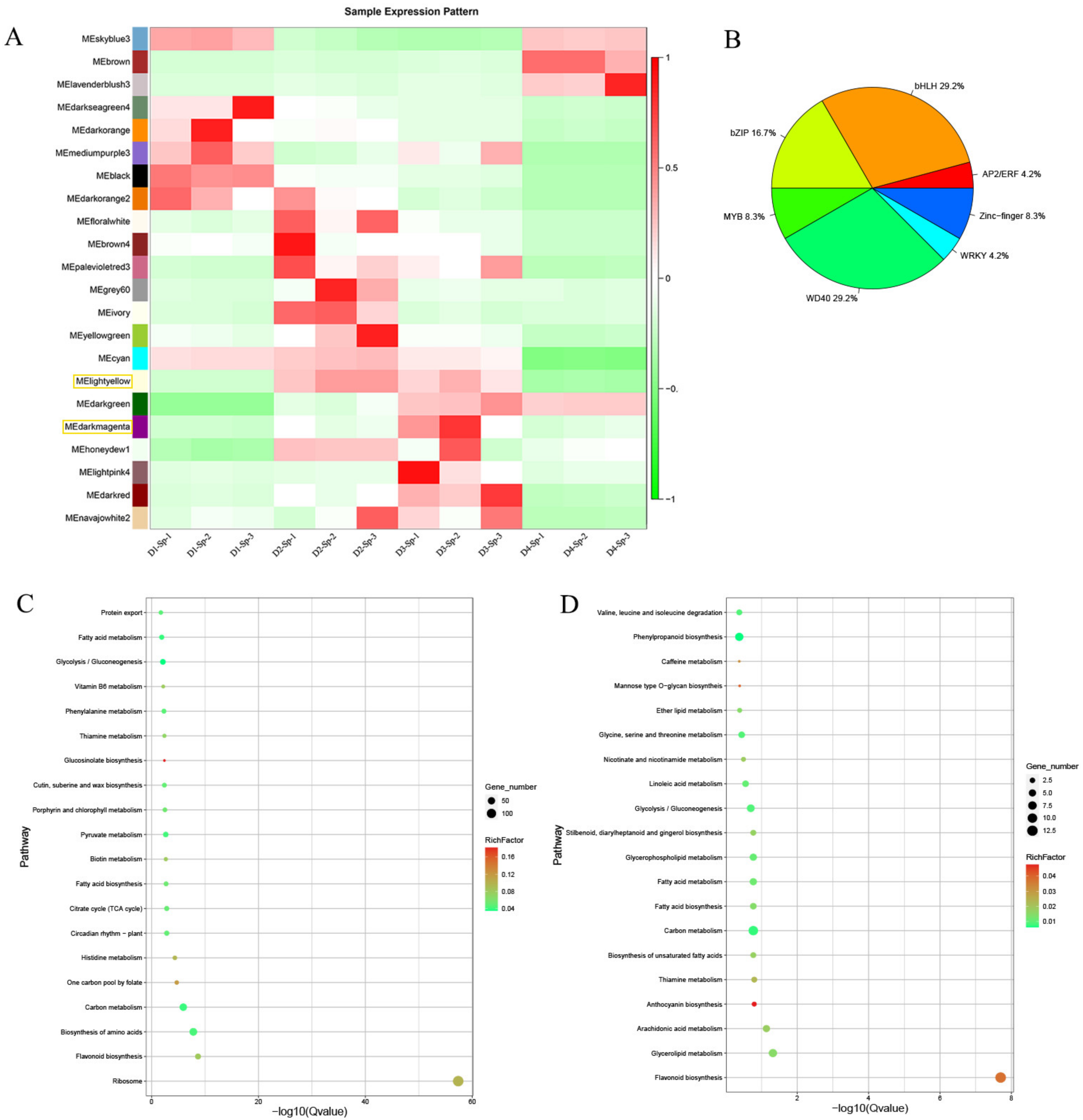
2.6. Identification of DEGs and Co-Expression Network Analysis
2.7. Specifically Expressed Genes in Petal Spots
2.8. Key DEGs Responsible for Anthocyanin Biosynthesis and Quantitative Reverse Transcription PCR (RT-qPCR) Validation
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Sampling
4.2. Sample Preparation and Extraction
4.3. Metabolite Profiling
4.4. RNA Extraction, Quantification, and Sequencing
4.5. Transcriptome Data Analysis
4.6. Petal Surface Analysis Using Scanning Electron Microscopy
4.7. RT-qPCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of Plant Pigments: Anthocyanins, Betalains and Carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Nishihara, M.; Nakatsuka, T. Genetic Engineering of Novel Flower Colors in Floricultural Plants: Recent Advances via Transgenic Approaches. In Protocols for In Vitro Propagation of Ornamental Plants; Methods in Molecular Biology; Jain, S.M., Ochatt, S.J., Eds.; Humana Press: Totowa, NJ, USA, 2010; Volume 589, pp. 325–347. ISBN 978-1-60327-390-9. [Google Scholar]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering Anthocyanin Biosynthesis in Plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Tao, J. Recent Advances on the Development and Regulation of Flower Color in Ornamental Plants. Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, S.; Tao, R.; Yin, L.; Ni, J.; Yang, Q.; Yan, X.; Yang, F.; Guo, X.; Li, H.; Teng, Y. Two B-box Proteins, PpBBX18 and PpBBX21, Antagonistically Regulate Anthocyanin Biosynthesis via Competitive Association with Pyrus Pyrifolia Elongated Hypocotyl 5 in the Peel of Pear Fruit. Plant J. 2019, 100, 1208–1223. [Google Scholar] [CrossRef] [PubMed]
- Jian, W.; Cao, H.; Yuan, S.; Liu, Y.; Lu, J.; Lu, W.; Li, N.; Wang, J.; Zou, J.; Tang, N.; et al. SlMYB75, an MYB-Type Transcription Factor, Promotes Anthocyanin Accumulation and Enhances Volatile Aroma Production in Tomato Fruits. Hortic. Res. 2019, 6, 22. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Dubos, C.; Lepiniec, L. Transcriptional Control of Flavonoid Biosynthesis by MYB–BHLH–WDR Complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef]
- Feng, C.; Ding, D.; Feng, C.; Kang, M. The Identification of an R2R3-MYB Transcription Factor Involved in Regulating Anthocyanin Biosynthesis in Primulina swinglei Flowers. Gene 2020, 752, 144788. [Google Scholar] [CrossRef]
- Lai, Y.-S.; Yamagishi, M.; Suzuki, T. Elevated Temperature Inhibits Anthocyanin Biosynthesis in the Tepals of an Oriental Hybrid Lily via the Suppression of LhMYB12 Transcription. Sci. Hortic. 2011, 132, 59–65. [Google Scholar] [CrossRef]
- An, J.; Wang, X.; Zhang, X.; Bi, S.; You, C.; Hao, Y. Md BBX 22 Regulates UV-B-induced Anthocyanin Biosynthesis through Regulating the Function of Md HY 5 and Is Targeted by Md BT 2 for 26S Proteasome-mediated Degradation. Plant Biotechnol. J. 2019, 17, 2231–2233. [Google Scholar] [CrossRef]
- Plunkett, B.J.; Henry-Kirk, R.; Friend, A.; Diack, R.; Helbig, S.; Mouhu, K.; Tomes, S.; Dare, A.P.; Espley, R.V.; Putterill, J.; et al. Apple B-Box Factors Regulate Light-Responsive Anthocyanin Biosynthesis Genes. Sci. Rep. 2019, 9, 17762. [Google Scholar] [CrossRef]
- Yang, B.; He, S.; Liu, Y.; Liu, B.; Ju, Y.; Kang, D.; Sun, X.; Fang, Y. Transcriptomics Integrated with Metabolomics Reveals the Effect of Regulated Deficit Irrigation on Anthocyanin Biosynthesis in Cabernet Sauvignon Grape Berries. Food Chem. 2020, 314, 126170. [Google Scholar] [CrossRef]
- Lim, K.-B.; Barba-Gonzalez, R.; Zhou, S.; Ramanna, M.S.; van Tuyl, J.M. Interspecific Hybridization in Lily (Lilium): Taxonomic and Commercial Aspects of Using Species Hybrids in Breeding. In Floriculture, Ornamental and Plant Biotechnology; Teixeira da Silva, J.A., Ed.; Global Science Books Ltd.: Carrollton, GA, USA, 2008; pp. 146–150. [Google Scholar]
- Matthews, V. The International Lily Register and Checklist; The Royal Horticultural Society: London, UK, 2007. [Google Scholar]
- Yamagishi, M. How Genes Paint Lily Flowers: Regulation of Colouration and Pigmentation Patterning. Sci. Hortic. 2013, 163, 27–36. [Google Scholar] [CrossRef]
- Yamagishi, M.; Akagi, K. Morphology and Heredity of Tepal Spots in Asiatic and Oriental Hybrid Lilies (Lilium spp.). Euphytica 2013, 194, 325–334. [Google Scholar] [CrossRef]
- Yamagishi, M.; Toda, S.; Tasaki, K. The Novel Allele of the Lh MYB 12 Gene Is Involved in Splatter-type Spot Formation on the Flower Tepals of Asiatic Hybrid Lilies (Lilium spp.). New Phytol. 2014, 201, 1009–1020. [Google Scholar] [CrossRef]
- Wang, L.J.; Lian, Y.Q.; Jia, G.X. Hybridization of Lilium leichtlinii var. maximowiczii with Longiflorum × Asiatic and Asiatic cultivars and identification of the hybrid progeny. Acta Hortic. 2017, 1171, 223–230. [Google Scholar] [CrossRef]
- Kim, J.H.; Truong, N.X.; Song, Y.-S.; Kim, N.-S. Natural Triploid Lilium Leichtlinii Var. Maximowiczii Populations in Korea: Natural Triploid Lilium. Plant Species Biol. 2016, 31, 98–106. [Google Scholar] [CrossRef]
- Yamagishi, M.; Kishimoto, S.; Nakayama, M. Carotenoid Composition and Changes in Expression of Carotenoid Biosynthetic Genes in Tepals of Asiatic Hybrid Lily. Plant Breed. 2010, 129, 100–107. [Google Scholar] [CrossRef]
- Hai, N.T.L.; Masuda, J.; Miyajima, I.; Thien, N.Q.; Mojtahedi, N.; Hiramatsu, M.; Kim, J.-H.; Okubo, H. Involvement of Carotenoid Cleavage Dioxygenase 4 Gene in Tepal Color Change in Lilium Brownii Var. Colchesteri. J. Japan. Soc. Hort. Sci. 2012, 81, 366–373. [Google Scholar] [CrossRef] [Green Version]
- Ohmiya, A. Diversity of Carotenoid Composition in Flower Petals. JARQ 2011, 45, 163–171. [Google Scholar] [CrossRef] [Green Version]
- Yamamizo, C.; Kishimoto, S.; Ohmiya, A. Carotenoid Composition and Carotenogenic Gene Expression during Ipomoea Petal Development. J. Exp. Bot. 2010, 61, 709–719. [Google Scholar] [CrossRef]
- Valadon, L.R.G.; Mummery, R.S. Carotenoids of Lilies and of Red Pepper: Biogenesis of Capsanthin and Capsorubin. Z. Pflanzenphysiol. 1977, 82, 407–416. [Google Scholar] [CrossRef]
- Jeknić, Z.; Morré, J.T.; Jeknić, S.; Jevremović, S.; Subotić, A.; Chen, T.H.H. Cloning and Functional Characterization of a Gene for Capsanthin-Capsorubin Synthase from Tiger Lily (Lilium Lancifolium Thunb. ‘Splendens’). Plant Cell Physiol. 2012, 53, 1899–1912. [Google Scholar] [CrossRef] [PubMed]
- Nørbæk, R.; Kondo, T. Anthocyanins from ~owers of Lilium “Liliaceae#. Phytochemistry 1999, 50, 1181–1184. [Google Scholar]
- Yamagishi, M. Oriental Hybrid Lily Sorbonne Homologue of LhMYB12 Regulates Anthocyanin Biosyntheses in Flower Tepals and Tepal Spots. Mol. Breed. 2011, 28, 381–389. [Google Scholar] [CrossRef]
- Yamagishi, M.; Yoshida, Y.; Nakayama, M. The Transcription Factor LhMYB12 Determines Anthocyanin Pigmentation in the Tepals of Asiatic Hybrid Lilies (Lilium spp.) and Regulates Pigment Quantity. Mol. Breed. 2012, 30, 913–925. [Google Scholar] [CrossRef]
- Abe, H.; Nakano, M.; Nakatsuka, A.; Nakayama, M.; Koshioka, M.; Yamagishi, M. Genetic Analysis of Floral Anthocyanin Pigmentation Traits in Asiatic Hybrid Lily Using Molecular Linkage Maps. Theor. Appl. Genet. 2002, 105, 1175–1182. [Google Scholar] [CrossRef]
- Ng, J.; Smith, S.D. Widespread Flower Color Convergence in Solanaceae via Alternate Biochemical Pathways. New Phytol. 2016, 209, 407–417. [Google Scholar] [CrossRef]
- Jaakola, L. New Insights into the Regulation of Anthocyanin Biosynthesis in Fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Petrussa, E.; Braidot, E.; Zancani, M.; Peresson, C.; Bertolini, A.; Patui, S.; Vianello, A. Plant Flavonoids—Biosynthesis, Transport and Involvement in Stress Responses. IJMS 2013, 14, 14950–14973. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Chen, H.; Chen, X.; Lu, J.; Chen, D.; Luo, C.; Cheng, X.; Huang, C. Transcriptomic and Metabolomic Analyses Reveal That MYB Transcription Factors Regulate Anthocyanin Synthesis and Accumulation in the Disc Florets of the Anemone Form of Chrysanthemum Morifolium. Sci. Hortic. 2023, 307, 110847. [Google Scholar] [CrossRef]
- Fatihah, H.N.N.; Wolinska, K. Expression of Anthocyanin Biosynthesis-Related Genes during Flower Development in Lilium spp. Plant Gene 2022, 31, 100372. [Google Scholar] [CrossRef]
- Nakatsuka, A.; Izumi, Y.; Yamagishi, M. Spatial and Temporal Expression of Chalcone Synthase and Dihydroflavonol 4-Reductase Genes in the Asiatic Hybrid Lily. Plant Sci. 2003, 165, 759–767. [Google Scholar] [CrossRef]
- Lai, Y.-S.; Shimoyamada, Y.; Nakayama, M.; Yamagishi, M. Pigment Accumulation and Transcription of LhMYB12 and Anthocyanin Biosynthesis Genes during Flower Development in the Asiatic Hybrid Lily (Lilium spp.). Plant Sci. 2012, 193–194, 136–147. [Google Scholar] [CrossRef]
- Morita, Y.; Saito, R.; Ban, Y.; Tanikawa, N.; Kuchitsu, K.; Ando, T.; Yoshikawa, M.; Habu, Y.; Ozeki, Y.; Nakayama, M. Tandemly Arranged Chalcone Synthase A Genes Contribute to the Spatially Regulated Expression of SiRNA and the Natural Bicolor Floral Phenotype in Petunia Hybrida: Endogenous SiRNA and Bicolored Flowers of Petunia. Plant J. 2012, 70, 739–749. [Google Scholar] [CrossRef]
- Deng, X.; Bashandy, H.; Ainasoja, M.; Kontturi, J.; Pietiäinen, M.; Laitinen, R.A.E.; Albert, V.A.; Valkonen, J.P.T.; Elomaa, P.; Teeri, T.H. Functional Diversification of Duplicated Chalcone Synthase Genes in Anthocyanin Biosynthesis of Gerbera Hybrida. New Phytol. 2014, 201, 1469–1483. [Google Scholar] [CrossRef]
- Gu, Z.; Zhu, J.; Hao, Q.; Yuan, Y.-W.; Duan, Y.-W.; Men, S.; Wang, Q.; Hou, Q.; Liu, Z.-A.; Shu, Q.; et al. A Novel R2R3-MYB Transcription Factor Contributes to Petal Blotch Formation by Regulating Organ-Specific Expression of PsCHS in Tree Peony (Paeonia Suffruticosa). Plant Cell Physiol. 2019, 60, 599–611. [Google Scholar] [CrossRef]
- Van der Krol, A.R.; Mur, L.A.; de Lange, P.; Gerats, A.G.M.; Mol, J.N.M.; Stuitje, A.R. Antisense Chalcone Synthase Genes in Petunia: Visualization of Variable Transgene Expression. Molec. Gen. Genet. 1990, 220, 204–212. [Google Scholar] [CrossRef]
- Koseki, M.; Goto, K.; Masuta, C.; Kanazawa, A. The Star-Type Color Pattern in Petunia Hybrida ‘Red Star’ Flowers Is Induced by Sequence-Specific Degradation of Chalcone Synthase RNA. Plant Cell Physiol. 2005, 46, 1879–1883. [Google Scholar] [CrossRef] [Green Version]
- Zeng, S.; Wu, M.; Zou, C.; Liu, X.; Shen, X.; Hayward, A.; Liu, C.; Wang, Y. Comparative Analysis of Anthocyanin Biosynthesis during Fruit Development in Two Lycium Species. Physiol. Plant. 2014, 150, 505–516. [Google Scholar] [CrossRef]
- Qi, Y.; Wei, H.; Gu, W.; Shi, W.; Jiang, L.; Deng, L.; Liu, X. Transcriptome Profiling Provides Insights into the Fruit Color Development of Wild Lycium ruthenicum Murr. from Qinghai–Tibet Plateau. Protoplasma 2021, 258, 33–43. [Google Scholar] [CrossRef]
- Li, X.; Li, Y.; Zhao, M.; Hu, Y.; Meng, F.; Song, X.; Tigabu, M.; Chiang, V.L.; Sederoff, R.; Ma, W.; et al. Molecular and Metabolic Insights into Anthocyanin Biosynthesis for Leaf Color Change in Chokecherry (Padus Virginiana). IJMS 2021, 22, 10697. [Google Scholar] [CrossRef] [PubMed]
- Nakatsuka, A.; Yamagishi, M.; Nakano, M.; Tasaki, K.; Kobayashi, N. Light-Induced Expression of Basic Helix-Loop-Helix Genes Involved in Anthocyanin Biosynthesis in Flowers and Leaves of Asiatic Hybrid Lily. Sci. Hortic. 2009, 121, 84–91. [Google Scholar] [CrossRef]
- Yamagishi, M.; Shimoyamada, Y.; Nakatsuka, T.; Masuda, K. Two R2R3-MYB Genes, Homologs of Petunia AN2, Regulate Anthocyanin Biosyntheses in Flower Tepals, Tepal Spots and Leaves of Asiatic Hybrid Lily. Plant Cell Physiol. 2010, 51, 463–474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Tasaki, K.; Yamagishi, M. Two Distinct Spontaneous Mutations Involved in White Flower Development in Lilium speciosum. Mol. Breed. 2015, 35, 193. [Google Scholar] [CrossRef]
- Martins, T.R.; Jiang, P.; Rausher, M.D. How Petals Change Their Spots: Cis-regulatory Re-wiring in Clarkia (Onagraceae). New Phytol. 2017, 216, 510–518. [Google Scholar] [CrossRef]
- Lin, R.; Rausher, M.D. R2R3-MYB Genes Control Petal Pigmentation Patterning in Clarkia gracilis ssp. Sonomensis (Onagraceae). New Phytol. 2021, 229, 1147–1162. [Google Scholar] [CrossRef]
- Yamagishi, M. High Promoter Sequence Variation in Subgroup 6 Members of R2R3-MYB Genes Is Involved in Different Floral Anthocyanin Color Patterns in Lilium spp. Mol. Genet. Genom. 2021, 296, 1005–1015. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness with Single-Copy Orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Salehi, M.; Abdollah, H.; Vahab; Jafarian, V.; Zarre, S.; Cuevas, J. Flower Initiation and Development in Endemic Iranian Lily (Lilium ledebourii Boiss). Acta Sci. Polonorum. Hortorum Cultus Ogrod. 2018, 17, 105–113. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing Real-Time PCR Data by the Comparative CT Method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Compounds | Molecular Weight (Da) | Q1 (Da) | Q3 (Da) | Rt (Min) | D1-Pe | D2-Pe | D3-Pe | D4-Pe | Trend |
|---|---|---|---|---|---|---|---|---|---|---|
| Carotenes | β-carotene | 536.44 | 537.60 | 177.10 | 6.23 | 12.85 ± 0.31 a | 11.82 ± 0.07 b | 4.93 ± 0.10 c | 1.22 ± 0.06 d | down |
| (E/Z)-phytoene | 544.50 | 545.30 | 81.00 | 4.83 | 0.00 c | 1.92 ± 0.15 b | 0.00 c | 4.81 ± 0.80 a | - | |
| Xanthophylls | capsanthin | 584.87 | 585.50 | 109.10 | 4.47 | 1.22 ± 0.07 c | 2.27 ± 0.15 c | 406.27 ± 33.40 b | 940.63 ± 98.30 a | up |
| capsorubin | 600.42 | 601.40 | 109.00 | 4.36 | 0.25 ± 0.01 c | 1.65 ± 0.04 c | 215.28 ± 11.42 b | 493.43 ± 51.11 a | up | |
| zeaxanthin | 568.43 | 569.40 | 477.50 | 4.61 | 0.85 ± 0.02 c | 2.12 ± 0.16 c | 10.73 ± 0.46 b | 31.24 ± 1.81 a | up | |
| violaxanthin | 600.42 | 601.40 | 221.00 | 1.59 | 11.66 ± 0.32 c | 6.89 ± 0.36 d | 13.11 ± 0.58 b | 29.07 ± 0.72 a | - | |
| β-cryptoxanthin | 552.43 | 553.50 | 177.40 | 5.49 | 1.66 ± 0.34 c | 1.73 ± 0.34 c | 2.29 ± 0.22 b | 3.74 ± 0.10 a | up | |
| lutein | 568.43 | 551.50 | 175.40 | 4.00 | 158.63 ± 9.98 a | 114.62 ± 1.51 b | 32.10 ± 1.36 c | 1.27 ± 0.09 d | down | |
| neoxanthin | 600.42 | 601.40 | 565.50 | 1.96 | 13.47 ± 0.10 a | 11.45 ± 0.84 b | 8.25 ± 0.76 c | 5.55 ± 0.43 d | down | |
| antheraxanthin | 584.42 | 585.50 | 175.40 | 2.87 | 2.05 ± 0.07 b | 2.20 ± 0.13 b | 10.99 ± 1.07 a | 0.00 c | - | |
| α-cryptoxanthin | 552.43 | 553.50 | 123.10 | 5.04 | 0.16 ± 0.04 ab | 0.00 c | 0.11 ± 0.01 b | 0.19 ± 0.03 a | - | |
| 8′-apo-beta-carotenal | 416.64 | 417.30 | 325.30 | 4.46 | 0.00 b | 0.00 b | 0.00 b | 0.04 ± 0.00 a | - | |
| echinenone | 550.90 | 551.60 | 203.10 | 5.51 | 0.01 ± 0.00 a | 0.01 ± 0.00 a | 0.00 ± 0.00 b | 0.01 ± 0.00 a | - | |
| β-citraurin | 432.60 | 433.30 | 341.10 | 2.79 | 0.00 b | 0.00 b | 0.05 ± 0.00 b | 0.61 ± 0.07 a | - |
| Compounds | Molecular Weight (Da) | Q1(Da) | Q3 (Da) | Rt (Min) | D1-Sp | D2-Sp | D3-Sp | D4-Sp | Trend |
|---|---|---|---|---|---|---|---|---|---|
| Cyanidin-3-O-glucoside | 449.11 | 449.10 | 287.10 | 5.54 | 0.00 d | 195.67 ± 7.04 c | 262.56 ± 1.27 b | 335.65 ± 3.37 a | up |
| Pelargonidin-3-O-rutinoside | 579.17 | 579.06 | 271.10 | 7.25 | 0.00 d | 74.23 ± 0.55 c | 153.24 ± 1.96 b | 235.45 ± 2.71 a | up |
| Cyanidin-3-O-rutinoside | 595.17 | 595.17 | 287.10 | 6.16 | 0.00 d | 38.23 ± 0.13 c | 98.59 ± 0.61 b | 150.75 ± 4.84 a | up |
| Peonidin-3-O-rutinoside | 609.19 | 609.50 | 301.10 | 7.87 | 0.00 d | 9.84 ± 0.14 c | 39.20 ± 0.69 b | 104.65 ± 0.58 a | up |
| Rutin | 610.15 | 611.20 | 303.10 | 8.88 | 2.07 ± 0.09 d | 9.57 ± 0.81 c | 36.45 ± 1.08 b | 42.36 ± 3.48 a | up |
| Cyanidin-3-O-rutinoside-5-O-glucoside | 757.22 | 757.22 | 287.10 | 4.06 | 0.00 d | 0.67 ± 0.03 c | 2.95 ± 0.07 b | 16.06 ± 0.34 a | up |
| Cyanidin-3-O-sophoroside | 611.16 | 611.20 | 287.15 | 4.90 | 0.00 d | 2.17 ± 0.10 c | 7.76 ± 0.19 b | 15.83 ± 0.75 a | up |
| Petunidin-3-O-galactoside | 479.12 | 479.10 | 317.10 | 5.88 | 0.00 d | 1.06 ± 0.04 c | 5.03 ± 0.02 b | 13.52 ± 0.22 a | up |
| Naringenin-7-O-glucoside | 434.12 | 435.10 | 273.10 | 9.39 | 0.79 ± 0.06 d | 4.04 ± 0.18 c | 6.11 ± 0.28 b | 12.38 ± 0.43 a | up |
| Quercetin-3-O-glucoside | 464.10 | 465.10 | 303.10 | 8.79 | 0.46 ± 0.03 c | 3.50 ± 0.20 b | 11.07 ± 0.87 a | 11.94 ± 0.19 a | up |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Li, X.; Chen, M.; Yang, L.; Zhang, Y. Molecular and Metabolic Insights into Anthocyanin Biosynthesis for Spot Formation on Lilium leichtlinii var. maximowiczii Flower Petals. Int. J. Mol. Sci. 2023, 24, 1844. https://doi.org/10.3390/ijms24031844
Wang Z, Li X, Chen M, Yang L, Zhang Y. Molecular and Metabolic Insights into Anthocyanin Biosynthesis for Spot Formation on Lilium leichtlinii var. maximowiczii Flower Petals. International Journal of Molecular Sciences. 2023; 24(3):1844. https://doi.org/10.3390/ijms24031844
Chicago/Turabian StyleWang, Zhen, Xin Li, Minmin Chen, Liuyan Yang, and Yongchun Zhang. 2023. "Molecular and Metabolic Insights into Anthocyanin Biosynthesis for Spot Formation on Lilium leichtlinii var. maximowiczii Flower Petals" International Journal of Molecular Sciences 24, no. 3: 1844. https://doi.org/10.3390/ijms24031844