
Age- and Lifespan-Dependent Differences in GO Caused DNA Damage in Acheta domesticus
 , , ,
, , ,
Abstract
:1. Introduction
2. Results
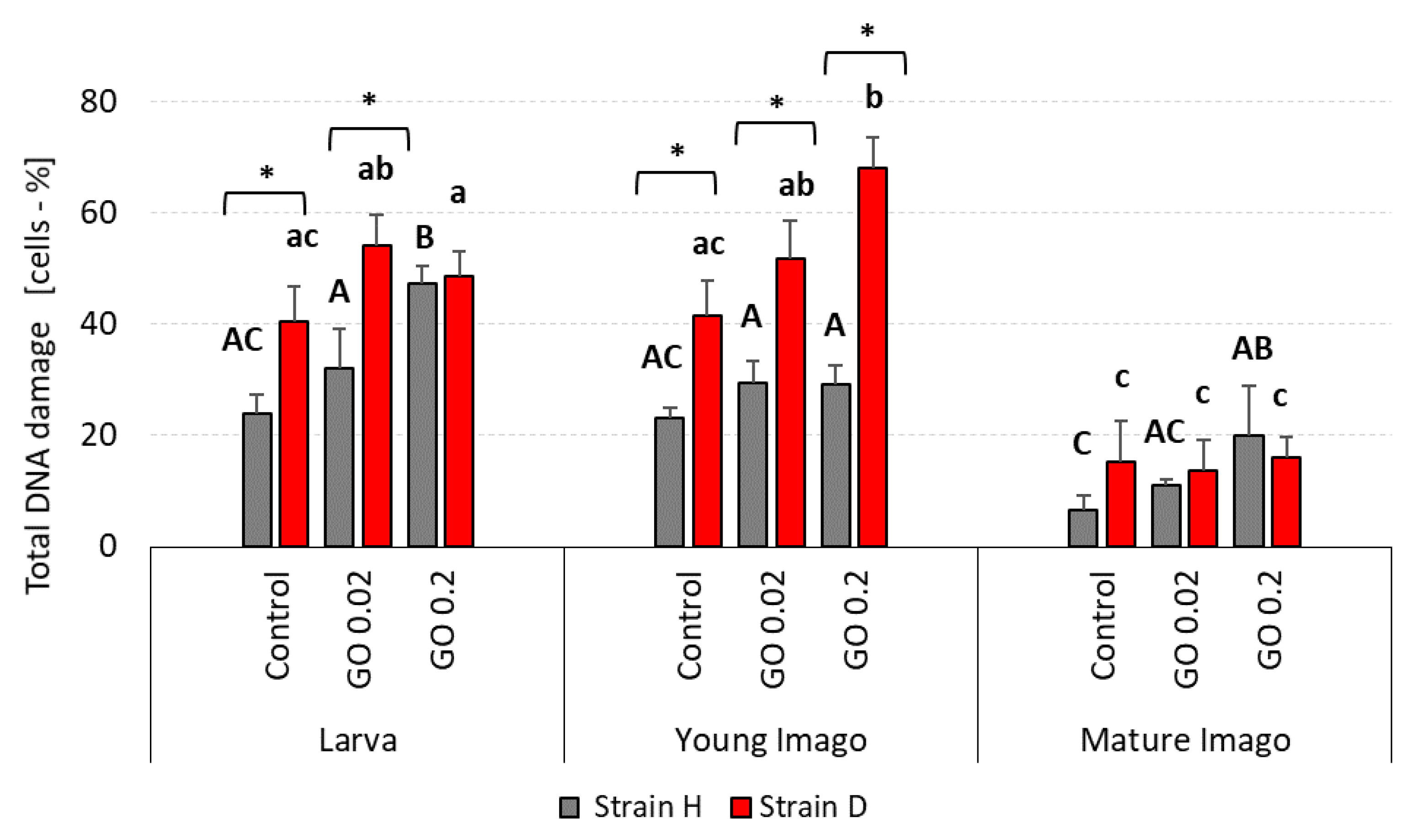
2.1. DNA Damage: Total
2.2. DNA Damage: Double Strand Breaks (DSB)
2.3. DNA Damage: 8-hydroxy-2′-deoxyguanosine (8-OHdG)
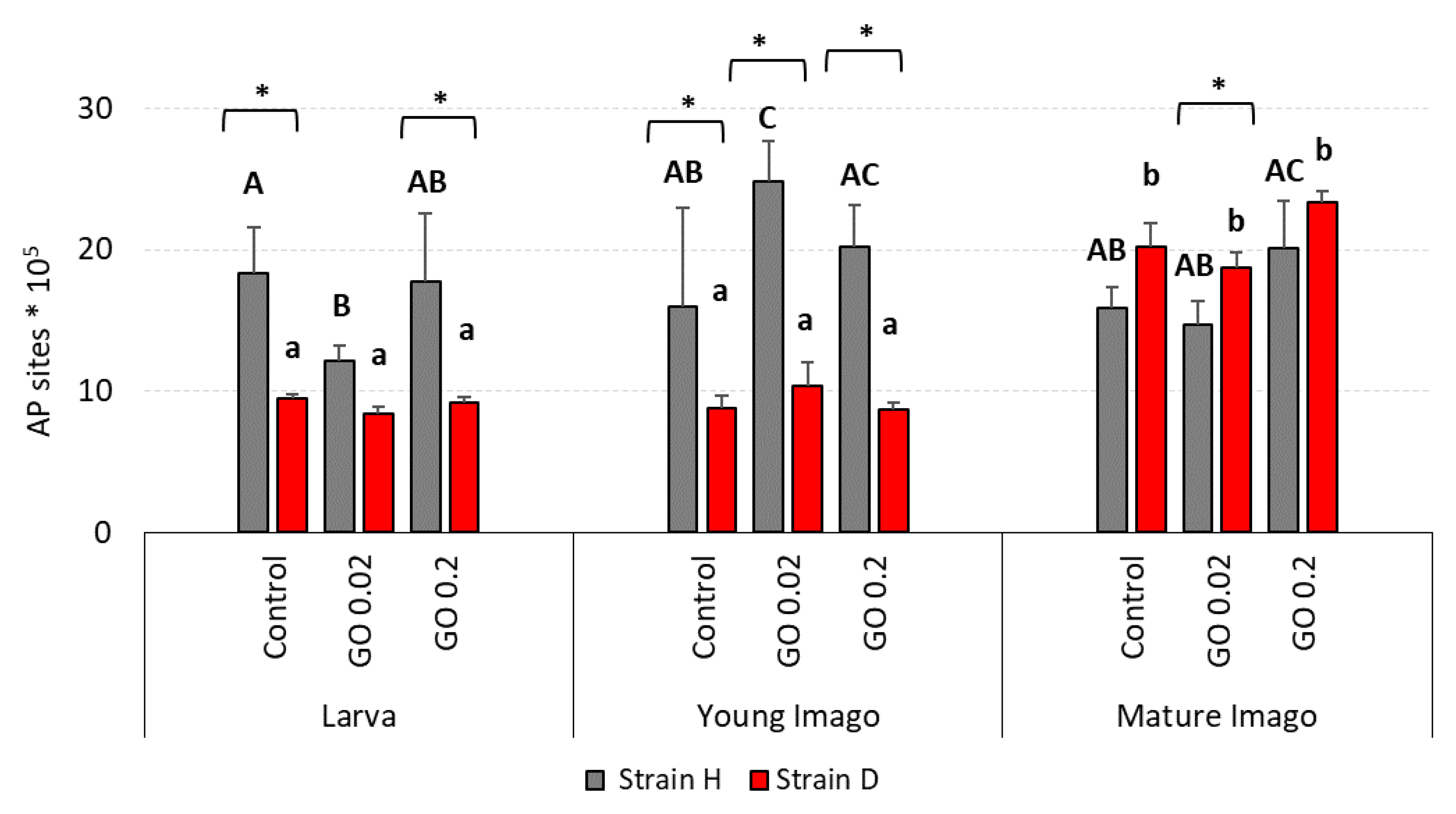
2.4. DNA Damage: Apurinic/Apyrimidinic Sites (AP Sites)
2.5. DNA Damage Response: Ataxia Telangiectasia Mutated (ATM) Kinase Phosphorylation (pATM)
2.6. DNA Damage Response: Phosphorylation of Histone H2A.X (pH2A.X)
2.7. Global DNA Methylation
3. Discussion
3.1. GO-Induced DNA Damage and Developmental Stage

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanoparticle Type | Biological System | Concentration/Dose | Finding/Effect | Reference |
|---|---|---|---|---|
| Graphene oxide; Graphene oxide quantum dots | M. aeruginosa | 49.32 mg/L 22.46 mg/L | Higher ROS; increase in malondialdehyde (MDA) concentration; antioxidant enzymes disruption | [41] |
| Graphene oxide | C. elegans | 0.01; 0.1 mg/L 1; 10; 100 mg/L | GO exposure induced autophagy in a dose dependent manner | [42] |
| Graphene oxide | C. elegans | 1 μg/L | Multi-generational toxicity: shortened life-span; smaller body size; reduced oocytes numbers; impairments of locomotion-related neurons | [43] |
| Graphene oxide Reduced graphene oxide (rGO) | D. rerio hepatocytes | 1–100 µg/mL | Higher ROS in cells exposed to rGO and stopping cell replication. Nevertheless, GO did not stop cell replication, but exposed cells had higher levels of apoptosis and necrosis | [44] |
| Graphene oxide | Mouse | 2 mg/kg 5 mg/kg | Increase in peroxidase activity and MDA concentration; liver inflammation; brain—neuronal cells not affected | [45] |
| Graphene oxide | A. domesticus | 0.2 mg/kg 20 mg/kg | Increase in antioxidative enzyme catalase activity, lower viability of gut cells, DNA damage in hemolymph and changes in the pattern of vitellogenin protein production | [31] |
| Nanodiamonds | A. domesticus | 20 µg/g 200 µg/g | DNA damage; increased activity of oxidative stress enzymes: catalase, glutathione peroxidase; increased total antioxidant capacity, increased level of heat shock protein | [46] |
| Ultrafine carbon black | Mouse | 5–55 µg/mL | <15 μg/mL no significant effect on splenocytes; >15 μg/mL induced ROS and malonaldehyde activity; decreased activity of superoxide dismutase and catalase; formation of protein corona in α-amylase and lipase | [47] |
| Multi-walled carbon nanotubes | Rat | 5000 mg/kg | Mild periarteriolar lymphoid cell depletion in the spleen; mild tubular cell degeneration on the cortex and medulla of both kidneys | [48] |
| Multi-walled carbon nanotubes | X. tropicalis | 0.5 mg/L 2.5 mg/L | Affected the formation of spermatogonia and oocytes; no effect on the heart or liver; changed the microbial community structure and diversity of gut microbiota | [49] |
| Multi-walled carbon nanotubes | C. carpio | 10 μg/L 50 μg/L | Downregulated expression of key steroidogenic and transcription factor genes related to testis and brain; decreases in serum testosterone and 11-ketotestosterone levels; increased activity of glutathione-S-transferases, superoxide dismutase, and catalase in both testis and brain | [50] |
| Fullerenes | M. salmoides | 0.5 ppm | Lipid peroxidation in brain; depletion of total glutathione | [51] |
| Carbon nanofibers | P. expansa | 1 mg/L 10 mg/L | Higher nitrite, hydrogen peroxide and lipid peroxidation levels in liver and brain; increased total glutathione, catalase and superoxide dismutase; damage in erythrocyte DNA; higher apoptosis and necrosis in erythrocytes; increased cerebral and hepatic acetylcholinesterase | [52] |
| Carbon nanofibers | Diamesa sp., D. cryptomeria; G. suifunensis | 100 mg/L | CNFs accumulated in the intestines; no toxic effects | [53] |
| Carbon nanofibers | E. fetida, D. rerio, O. niloticus | 500 μg/g 10 μg/mL | Erythrocyte nuclear abnormalities; nanoparticle accumulation at trophic levels | [54] |
3.2. Selection for longevity and susceptibility to DNA damage caused by GO
3.3. Global DNA Methylation in Acheta domesticus
4. Materials and Methods
4.1. Graphene Oxide Characteristics
4.2. Characteristics of the Species
4.3. Food Preparation: Graphene Oxide Food, Control Food
4.4. Experimental Model
4.5. DNA isolation
4.6. Tissue Preparation for Flow Cytometry
4.7. Measurements of Selected Parameters
4.7.1. Total DNA Damage, Double Strand Breaks (DSB), pATM and pH2A.X
4.7.2. Apurinic/Apyrimidinic (AP) Sites
4.7.3. 8-hydroxy-2′-deoxyguanosine (8-OHdG)
4.7.4. Global DNA Methylation
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, Y.; Murali, S.; Cai, W.; Li, X.; Suk, J.W.; Potts, J.R.; Ruoff, R.S. Graphene and Graphene Oxide: Synthesis, Properties, and Applications. Adv. Mater. 2010, 22, 3906–3924. [Google Scholar] [CrossRef] [PubMed]
- Chung, C.; Kim, Y.-K.; Shin, D.; Ryoo, S.-R.; Hong, B.H.; Min, D.-H. Biomedical Applications of Graphene and Graphene Oxide. Acc. Chem. Res. 2013, 46, 2211–2224. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, S.; Mohanty, S.; Mukherjee, S.; Basu, S.; Mishra, M. Graphene and Graphene Oxide as Nanomaterials for Medicine and Biology Application. J. Nanostructure Chem. 2018, 8, 123–137. [Google Scholar] [CrossRef] [Green Version]
- Jiříčková, A.; Jankovský, O.; Sofer, Z.; Sedmidubský, D. Synthesis and Applications of Graphene Oxide. Materials 2022, 15, 920. [Google Scholar] [CrossRef] [PubMed]
- Razaq, A.; Bibi, F.; Zheng, X.; Papadakis, R.; Jafri, S.H.M.; Li, H. Review on Graphene-, Graphene Oxide-, Reduced Graphene Oxide-Based Flexible Composites: From Fabrication to Applications. Materials 2022, 15, 1012. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.P.; Herrera, C.E.; Singh, B.; Singh, S.; Singh, R.K.; Kumar, R. Graphene Oxide: An Efficient Material and Recent Approach for Biotechnological and Biomedical Applications. Mater. Sci. Eng. C 2018, 86, 173–197. [Google Scholar] [CrossRef] [PubMed]
- Joshi, D.J.; Koduru, J.R.; Malek, N.I.; Hussain, C.M.; Kailasa, S.K. Surface Modifications and Analytical Applications of Graphene Oxide: A Review. TrAC Trends Anal. Chem. 2021, 144, 116448. [Google Scholar] [CrossRef]
- Nebol’sin, V.A.; Galstyan, V.; Silina, Y.E. Graphene Oxide and Its Chemical Nature: Multi-Stage Interactions between the Oxygen and Graphene. Surf. Interfaces 2020, 21, 100763. [Google Scholar] [CrossRef]
- Dziewięcka, M.; Pawlyta, M.; Majchrzycki, Ł.; Balin, K.; Barteczko, S.; Czerkawska, M.; Augustyniak, M. The Structure–Properties–Cytotoxicity Interplay: A Crucial Pathway to Determining Graphene Oxide Biocompatibility. Int. J. Mol. Sci. 2021, 22, 5401. [Google Scholar] [CrossRef]
- Chang, Y.; Yang, S.-T.; Liu, J.-H.; Dong, E.; Wang, Y.; Cao, A.; Liu, Y.; Wang, H. In Vitro Toxicity Evaluation of Graphene Oxide on A549 Cells. Toxicol Lett. 2011, 200, 201–210. [Google Scholar] [CrossRef]
- Wang, A.; Pu, K.; Dong, B.; Liu, Y.; Zhang, L.; Zhang, Z.; Duan, W.; Zhu, Y. Role of Surface Charge and Oxidative Stress in Cytotoxicity and Genotoxicity of Graphene Oxide towards Human Lung Fibroblast Cells. J. Appl. Toxicol. 2013, 33, 1156–1164. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Yin, J.; Liang, Y.; Yuan, S.; Wang, F.; Song, M.; Wang, H. Oxidative Stress and Immunotoxicity Induced by Graphene Oxide in Zebrafish. Aquat. Toxicol. 2016, 174, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Dziewięcka, M.; Karpeta-Kaczmarek, J.; Augustyniak, M.; Majchrzycki, Ł.; Augustyniak-Jabłokow, M.A. Evaluation of in Vivo Graphene Oxide Toxicity for Acheta Domesticus in Relation to Nanomaterial Purity and Time Passed from the Exposure. J. Hazard. Mater. 2016, 305, 30–40. [Google Scholar] [CrossRef]
- Dziewięcka, M.; Karpeta-Kaczmarek, J.; Augustyniak, M.; Rost-Roszkowska, M. Short-Term in Vivo Exposure to Graphene Oxide Can Cause Damage to the Gut and Testis. J. Hazard. Mater. 2017, 328, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Dziewięcka, M.; Witas, P.; Karpeta-Kaczmarek, J.; Kwaśniewska, J.; Flasz, B.; Balin, K.; Augustyniak, M. Reduced Fecundity and Cellular Changes in Acheta Domesticus after Multigenerational Exposure to Graphene Oxide Nanoparticles in Food. Sci. Total Environ. 2018, 635, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Dziewięcka, M.; Flasz, B.; Rost - Roszkowska, M.; Kędziorski, A.; Kochanowicz, A.; Augustyniak, M. Graphene Oxide as a New Anthropogenic Stress Factor - Multigenerational Study at the Molecular, Cellular, Individual and Population Level of Acheta Domesticus. J. Hazard. Mater. 2020, 396, 122775. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Yang, Y.; Zhao, L.; Chen, J.; Duan, G.; Yang, Z.; Zhou, R. Graphene Oxide Toxicity in W1118 Flies. Sci. Total Environ. 2022, 805, 150302. [Google Scholar] [CrossRef]
- Lv, X.; Yang, Y.; Tao, Y.; Jiang, Y.; Chen, B.; Zhu, X.; Cai, Z.; Li, B. A Mechanism Study on Toxicity of Graphene Oxide to Daphnia Magna: Direct Link between Bioaccumulation and Oxidative Stress. Environ. Pollut. 2018, 234, 953–959. [Google Scholar] [CrossRef]
- Liu, Y.; Wen, J.; Gao, Y.; Li, T.; Wang, H.; Yan, H.; Niu, B.; Guo, R. Antibacterial Graphene Oxide Coatings on Polymer Substrate. Appl. Surf. Sci. 2018, 436, 624–630. [Google Scholar] [CrossRef]
- Xiong, G.; Deng, Y.; Liao, X.; Zhang, J.; Cheng, B.; Cao, Z.; Lu, H. Graphene Oxide Nanoparticles Induce Hepatic Dysfunction through the Regulation of Innate Immune Signaling in Zebrafish (Danio Rerio). Nanotoxicology 2020, 14, 667–682. [Google Scholar] [CrossRef]
- Ghiasvand Mohammadkhani, L.; Khoshkam, M.; Kompany-Zareh, M.; Amiri, M.; Ramazani, A. Metabolomics Approach to Study in Vivo Toxicity of Graphene Oxide Nanosheets. J. Appl. Toxicol. 2022, 42, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Wang, X.; Zhang, P.; Sun, F.; Chen, Z.; Ma, W.; Meng, F.; Hao, H.; Shang, X. Silica Nanoparticles Cause Spermatogenesis Dysfunction in Mice via Inducing Cell Cycle Arrest and Apoptosis. Ecotoxicol. Env. Saf. 2022, 231, 113210. [Google Scholar] [CrossRef]
- Liu, S.; Zeng, T.H.; Hofmann, M.; Burcombe, E.; Wei, J.; Jiang, R.; Kong, J.; Chen, Y. Antibacterial Activity of Graphite, Graphite Oxide, Graphene Oxide, and Reduced Graphene Oxide: Membrane and Oxidative Stress. ACS Nano 2011, 5, 6971–6980. [Google Scholar] [CrossRef] [PubMed]
- Farid, M.U.; Jeong, S.; Seo, D.H.; Ahmed, R.; Lau, C.; Gali, N.K.; Ning, Z.; An, A.K. Mechanistic Insight into the In-Vitro Toxicity of Graphene Oxide against Biofilm Forming Bacteria Using Laser-Induced Breakdown Spectroscopy. Nanoscale 2018, 10, 4475–4487. [Google Scholar] [CrossRef]
- Wu, K.; Zhou, Q.; Ouyang, S. Direct and Indirect Genotoxicity of Graphene Family Nanomaterials on DNA—A Review. Nanomaterials 2021, 11, 2889. [Google Scholar] [CrossRef] [PubMed]
- VALAVANIDIS, A.; VLACHOGIANNI, T.; FIOTAKIS, C. 8-Hydroxy-2′ -Deoxyguanosine (8-OHdG): A Critical Biomarker of Oxidative Stress and Carcinogenesis. J. Environ. Sci. Health Part C 2009, 27, 120–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Firsanov, D.V.; Solovjeva, L.V.; Mikhailov, V.M.; Svetlova, M.P. Methods for the Detection of DNA Damage. In Genome Stability; Elsevier: Amsterdam, The Netherlands, 2016; pp. 635–649. [Google Scholar]
- Kim, J.J.; Lee, S.Y.; Miller, K.M. Preserving Genome Integrity and Function: The DNA Damage Response and Histone Modifications. Crit Rev. Biochem. Mol. Biol. 2019, 54, 208–241. [Google Scholar] [CrossRef]
- Chen, Q.; Bian, C.; Wang, X.; Liu, X.; Ahmad Kassab, M.; Yu, Y.; Yu, X. ADP-ribosylation of Histone Variant H2AX Promotes Base Excision Repair. EMBO J. 2021, 40, e104542. [Google Scholar] [CrossRef]
- Yousefzadeh, M.; Henpita, C.; Vyas, R.; Soto-Palma, C.; Robbins, P.; Niedernhofer, L. DNA Damage—How and Why We Age? Elife 2021, 10, e62852. [Google Scholar] [CrossRef]
- Flasz, B.; Dziewięcka, M.; Kędziorski, A.; Tarnawska, M.; Augustyniak, M. Vitellogenin Expression, DNA Damage, Health Status of Cells and Catalase Activity in Acheta Domesticus Selected According to Their Longevity after Graphene Oxide Treatment. Sci. Total Environ. 2020, 737, 140274. [Google Scholar] [CrossRef]
- Flasz, B.; Dziewięcka, M.; Kędziorski, A.; Tarnawska, M.; Augustyniak, M. Multigenerational Graphene Oxide Intoxication Results in Reproduction Disorders at the Molecular Level of Vitellogenin Protein Expression in Acheta Domesticus. Chemosphere 2021, 280, 130772. [Google Scholar] [CrossRef] [PubMed]
- Flasz, B.; Dziewięcka, M.; Kędziorski, A.; Tarnawska, M.; Augustyniak, J.; Augustyniak, M. Multigenerational Selection towards Longevity Changes the Protective Role of Vitamin C against Graphene Oxide-Induced Oxidative Stress in House Crickets. Environ. Pollut. 2021, 290, 117996. [Google Scholar] [CrossRef] [PubMed]
- Augustyniak, M.; Babczyńska, A.; Dziewięcka, M.; Flasz, B.; Karpeta-Kaczmarek, J.; Kędziorski, A.; Mazur, B.; Rozpędek, K.; Seyed Alian, R.; Skowronek, M.; et al. Does Age Pay off? Effects of Three-Generational Experiments of Nanodiamond Exposure and Withdrawal in Wild and Longevity-Selected Model Animals. Chemosphere 2022, 303, 135129. [Google Scholar] [CrossRef] [PubMed]
- Domenech, J.; Rodríguez-Garraus, A.; López de Cerain, A.; Azqueta, A.; Catalán, J. Genotoxicity of Graphene-Based Materials. Nanomaterials 2022, 12, 1795. [Google Scholar] [CrossRef] [PubMed]
- Cano, A.M.; Maul, J.D.; Saed, M.; Shah, S.A.; Green, M.J.; Cañas-Carrell, J.E. Bioaccumulation, Stress, and Swimming Impairment in Daphnia Magna Exposed to Multiwalled Carbon Nanotubes, Graphene, and Graphene Oxide. Env. Toxicol. Chem. 2017, 36, 2199–2204. [Google Scholar] [CrossRef]
- Hu, C.; Wang, Q.; Zhao, H.; Wang, L.; Guo, S.; Li, X. Ecotoxicological Effects of Graphene Oxide on the Protozoan Euglena Gracilis. Chemosphere 2015, 128, 184–190. [Google Scholar] [CrossRef]
- Boyko, A.; Kovalchuk, I. Genome Instability and Epigenetic Modification—Heritable Responses to Environmental Stress? Curr. Opin. Plant Biol. 2011, 14, 260–266. [Google Scholar] [CrossRef]
- Horne, S.D.; Chowdhury, S.K.; Heng, H.H.Q. Stress, Genomic Adaptation, and the Evolutionary Trade-Off. Front. Genet. 2014, 5, 92. [Google Scholar] [CrossRef] [Green Version]
- Augustyniak, M.; Tarnawska, M.; Dziewięcka, M.; Kafel, A.; Rost-Roszkowska, M.; Babczyńska, A. DNA Damage in Spodoptera Exigua after Multigenerational Cadmium Exposure—A Trade-off between Genome Stability and Adaptation. Sci. Total Environ. 2020, 745, 141048. [Google Scholar] [CrossRef]
- Yan, Z.; Yang, X.; Lynch, I.; Cui, F. Comparative Evaluation of the Mechanisms of Toxicity of Graphene Oxide and Graphene Oxide Quantum Dots to Blue-Green Algae Microcystis Aeruginosa in the Aquatic Environment. J. Hazard Mater 2022, 425, 127898. [Google Scholar] [CrossRef]
- Dou, T.; Chen, J.; Wang, R.; Pu, X.; Wu, H.; Zhao, Y. Complementary Protective Effects of Autophagy and Oxidative Response against Graphene Oxide Toxicity in Caenorhabditis Elegans. Ecotoxicol. Env. Saf. 2022, 248, 114289. [Google Scholar] [CrossRef]
- Jin, L.; Dou, T.-T.; Chen, J.-Y.; Duan, M.-X.; Zhen, Q.; Wu, H.-Z.; Zhao, Y.-L. Sublethal Toxicity of Graphene Oxide in Caenorhabditis Elegans under Multi-Generational Exposure. Ecotoxicol. Env. Saf. 2022, 229, 113064. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, P.R.; Souza, J.P.; Estevão, B.M.; Altei, W.F.; Carmo, T.L.L.; Santos, F.A.; Araújo, H.S.S.; Zucolotto, V.; Fernandes, M.N. Concentration- and Time-Dependence Toxicity of Graphene Oxide (GO) and Reduced Graphene Oxide (RGO) Nanosheets upon Zebrafish Liver Cell Line. Aquat. Toxicol. 2022, 248, 106199. [Google Scholar] [CrossRef] [PubMed]
- Rhazouani, A.; Gamrani, H.; Ed-Day, S.; Lafhal, K.; Boulbaroud, S.; Gebrati, L.; Fdil, N.; AZIZ, F. Sub-Acute Toxicity of Graphene Oxide (GO) Nanoparticles in Male Mice after Intraperitoneal Injection: Behavioral Study and Histopathological Evaluation. Food Chem. Toxicol. 2023, 171, 113553. [Google Scholar] [CrossRef] [PubMed]
- Karpeta-Kaczmarek, J.; Dziewięcka, M.; Augustyniak, M.; Rost-Roszkowska, M.; Pawlyta, M. Oxidative Stress and Genotoxic Effects of Diamond Nanoparticles. Env. Res. 2016, 148. [Google Scholar] [CrossRef]
- Gao, S.; Li, T.; Pan, J.; Han, D.; Lin, J.; Niu, Q.; Liu, R. Toxic Effect and Mechanism of Ultrafine Carbon Black on Mouse Primary Splenocytes and Two Digestive Enzymes. Ecotoxicol. Env. Saf. 2021, 212, 111980. [Google Scholar] [CrossRef]
- Reyes, J.P.; Celorico, J.R.; DeYro, P.A.; Ochona, C.N.; Ochona, Z.A.; Visaya, B.A.; Basilia, B.A. Acute Toxicity and 28-Day Repeated Dose Studies of Multi-Walled Carbon Nanotubes. Mater Today Proc. 2022, 66, 3178–3184. [Google Scholar] [CrossRef]
- Zhao, J.; Luo, W.; Xu, Y.; Ling, J.; Deng, L. Potential Reproductive Toxicity of Multi-Walled Carbon Nanotubes and Their Chronic Exposure Effects on the Growth and Development of Xenopus Tropicalis. Sci. Total Environ. 2021, 766, 142652. [Google Scholar] [CrossRef]
- Deepa, S.; Mamta, S.-K.; Anitha, A.; Senthilkumaran, B. Exposure of Carbon Nanotubes Affects Testis and Brain of Common Carp. Env. Toxicol. Pharm. 2022, 95, 103957. [Google Scholar] [CrossRef]
- Eva, O. Manufactured Nanomaterials (Fullerenes, C60) Induce Oxidative Stress in the Brain of Juvenile Largemouth Bass. Env. Health Perspect 2004, 112, 1058–1062. [Google Scholar] [CrossRef]
- Guimarães, A.T.B.; Malafaia, G. Multiple Toxicity Endpoints Induced by Carbon Nanofibers in Amazon Turtle Juveniles: Outspreading Warns about Toxicological Risks to Reptiles. Sci. Total Environ. 2021, 779, 146514. [Google Scholar] [CrossRef] [PubMed]
- Chaika, V.; Pikula, K.; Vshivkova, T.; Zakharenko, A.; Reva, G.; Drozdov, K.; Vardavas, A.I.; Stivaktakis, P.D.; Nikolouzakis, T.K.; Stratidakis, A.K.; et al. The Toxic Influence and Biodegradation of Carbon Nanofibers in Freshwater Invertebrates of the Families Gammaridae, Ephemerellidae, and Chironomidae. Toxicol. Rep. 2020, 7, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.R.; Chagas, T.Q.; Silva, A.M.; Sueli de Lima Rodrigues, A.; Marinho da Luz, T.; Emmanuela de Andrade Vieira, J.; Malafaia, G. Trophic Transfer of Carbon Nanofibers among Eisenia Fetida, Danio Rerio and Oreochromis Niloticus and Their Toxicity at Upper Trophic Level. Chemosphere 2021, 263, 127657. [Google Scholar] [CrossRef]
- Prestwich, K.N.; Walker, T.J. Energetics of Singing in Crickets: Effect of Temperature in Three Trilling Species (Orthoptera: Gryllidae). J. Comp. Physiol. 1981, 143, 199–212. [Google Scholar] [CrossRef]
- Horch, H.W.; Mito, T.; Popadić, A.; Ohuchi, H.; Noji, S. The Cricket as a Model Organism; Horch, H.W., Mito, T., Popadić, A., Ohuchi, H., Noji, S., Eds.; Springer: Japan, Tokyo, 2017; ISBN 978-4-431-56476-8. [Google Scholar]
- Rogowitz, G.L.; Chappell, M.A. Energy Metabolism of Eucalyptus-Boring Beetles at Rest and during Locomotion: Gender Makes a Difference. J. Exp. Biol. 2000, 203, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Margotta, J.W.; Roberts, S.P.; Elekonich, M.M. Effects of Flight Activity and Age on Oxidative Damage in the Honey Bee, Apis Mellifera. J. Exp. Biol. 2018, 221, jeb183228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, J.; Swenberg, J.A. Endogenous Apurinic/Apyrimidinic Sites in Genomic DNA of Mammalian Tissues. Cancer Res. 1999, 59, 2522–2526. [Google Scholar]
- Krokan, H.E.; Bjoras, M. Base Excision Repair. Cold Spring Harb Perspect. Biol. 2013, 5, a012583. [Google Scholar] [CrossRef]
- Drohat, A.C.; Coey, C.T. Role of Base Excision “Repair” Enzymes in Erasing Epigenetic Marks from DNA. Chem. Rev. 2016, 116, 12711–12729. [Google Scholar] [CrossRef] [Green Version]
- Cai, Y.; Cao, H.; Wang, F.; Zhang, Y.; Kapranov, P. Complex Genomic Patterns of Abasic Sites in Mammalian DNA Revealed by a High-Resolution SSiNGLe-AP Method. Nat. Commun. 2022, 13, 5868. [Google Scholar] [CrossRef]
- Gensler, H.L.; Bernstein, H. DNA Damage as the Primary Cause of Aging. Q. Rev. Biol. 1981, 56, 279–303. [Google Scholar] [CrossRef] [PubMed]
- Garinis, G.A.; van der Horst, G.T.J.; Vijg, J.; H.J. Hoeijmakers, J. DNA Damage and Ageing: New-Age Ideas for an Age-Old Problem. Nat. Cell Biol. 2008, 10, 1241–1247. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, F.; Albertini, M.C.; Orciani, M.; Ceka, A.; Cricca, M.; Procopio, A.D.; Bonafè, M. DNA Damage Response (DDR) and Senescence: Shuttled Inflamma-MiRNAs on the Stage of Inflamm-Aging. Oncotarget 2015, 6, 35509–35521. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, B.; Pothof, J.; Vijg, J.; Hoeijmakers, J.H.J. The Central Role of DNA Damage in the Ageing Process. Nature 2021, 592, 695–703. [Google Scholar] [CrossRef]
- Petr, M.A.; Tulika, T.; Carmona-Marin, L.M.; Scheibye-Knudsen, M. Protecting the Aging Genome. Trends Cell Biol. 2020, 30, 117–132. [Google Scholar] [CrossRef] [Green Version]
- Herrup, K.; Li, J.; Chen, J. The Role of ATM and DNA Damage in Neurons: Upstream and Downstream Connections. DNA Repair 2013, 12, 600–604. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhao, Y.; Ge, W.; Zhang, P.; Liu, X.; Zhang, W.; Hao, Y.; Yu, S.; Li, L.; Chu, M.; et al. Oocyte Exposure to ZnO Nanoparticles Inhibits Early Embryonic Development through the γ-H2AX and NF-ΚB Signaling Pathways. Oncotarget 2017, 8, 42673–42692. [Google Scholar] [CrossRef] [Green Version]
- AshaRani, P.; Sethu, S.; Lim, H.; Balaji, G.; Valiyaveettil, S.; Hande, M.P. Differential Regulation of Intracellular Factors Mediating Cell Cycle, DNA Repair and Inflammation Following Exposure to Silver Nanoparticles in Human Cells. Genome Integr. 2012, 3, 2. [Google Scholar] [CrossRef] [Green Version]
- Wan, R.; Mo, Y.; Feng, L.; Chien, S.; Tollerud, D.J.; Zhang, Q. DNA Damage Caused by Metal Nanoparticles: Involvement of Oxidative Stress and Activation of ATM. Chem. Res. Toxicol. 2012, 25, 1402–1411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kansara, K.; Patel, P.; Shah, D.; Shukla, R.K.; Singh, S.; Kumar, A.; Dhawan, A. TiO2 Nanoparticles Induce DNA Double Strand Breaks and Cell Cycle Arrest in Human Alveolar Cells. Env. Mol. Mutagen 2015, 56, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Krasteva, N.; Staneva, D.; Vasileva, B.; Miloshev, G.; Georgieva, M. Bioactivity of PEGylated Graphene Oxide Nanoparticles Combined with Near-Infrared Laser Irradiation Studied in Colorectal Carcinoma Cells. Nanomaterials 2021, 11, 3061. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T. Life-History Evolution and the Genetics of Fitness Components in Drosophila Melanogaster. Genetics 2020, 214, 3–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Promislow, D.E.L.; Flatt, T.; Bonduriansky, R. The Biology of Aging in Insects: From Drosophila to Other Insects and Back. Annu. Rev. Entomol 2022, 67, 83–103. [Google Scholar] [CrossRef]
- Jemielity, S.; Chapuisat, M.; Parker, J.D.; Keller, L. Long Live the Queen: Studying Aging in Social Insects. Age 2005, 27, 241–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasaki, E.; Kobayashi, K.; Matsuura, K.; Iuchi, Y. An Efficient Antioxidant System in a Long-Lived Termite Queen. PLoS ONE 2017, 12, e0167412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, H.; Finkel, T. Key Proteins and Pathways That Regulate Lifespan. J. Biol. Chem. 2017, 292, 6452–6460. [Google Scholar] [CrossRef] [Green Version]
- Li, W.-J.; Wang, C.-W.; Tao, L.; Yan, Y.-H.; Zhang, M.-J.; Liu, Z.-X.; Li, Y.-X.; Zhao, H.-Q.; Li, X.-M.; He, X.-D.; et al. Insulin Signaling Regulates Longevity through Protein Phosphorylation in Caenorhabditis Elegans. Nat. Commun. 2021, 12, 4568. [Google Scholar] [CrossRef]
- Ehrlich, M.; Gama-Sosa, M.A.; Huang, L.-H.; Midgett, R.M.; Kuo, K.C.; McCune, R.A.; Gehrke, C. Amount and Distribution of 5-Methylcytosine in Human DNA from Different Types of Tissues or Cells. Nucleic Acids Res. 1982, 10, 2709–2721. [Google Scholar] [CrossRef]
- Lister, R.; Pelizzola, M.; Dowen, R.H.; Hawkins, R.D.; Hon, G.; Tonti-Filippini, J.; Nery, J.R.; Lee, L.; Ye, Z.; Ngo, Q.-M.; et al. Human DNA Methylomes at Base Resolution Show Widespread Epigenomic Differences. Nature 2009, 462, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Glastad, K.M.; Hunt, B.G.; Yi, S.v.; Goodisman, M.A.D. DNA Methylation in Insects: On the Brink of the Epigenomic Era. Insect Mol. Biol. 2011, 20, 553–565. [Google Scholar] [CrossRef]
- Field, L.M.; Lyko, F.; Mandrioli, M.; Prantera, G. DNA Methylation in Insects. Insect Mol. Biol. 2004, 13, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Lyko, F.; Maleszka, R. Insects as Innovative Models for Functional Studies of DNA Methylation. Trends Genet. 2011, 27, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Rao, S.R.v.; Gupta, V.S.; Hendre, R.R. 5-Methylcytosine Content in Gryllotalpa Fossor (Orthoptera). Genome 1992, 35, 163–166. [Google Scholar] [CrossRef]
- Cabej, N.R. The Epigenetic System of Inheritance. In Epigenetic Principles of Evolution; Elsevier: Amsterdam, The Netherlands, 2019; pp. 215–258. [Google Scholar]
- Tweedie, S.; Ng, H.-H.; Barlow, A.L.; Turner, B.M.; Hendrich, B.; Bird, A. Vestiges of a DNA Methylation System in Drosophila Melanogaster? Nat. Genet 1999, 23, 389–390. [Google Scholar] [CrossRef] [PubMed]
- Duncan, E.J.; Cunningham, C.B.; Dearden, P.K. Phenotypic Plasticity: What Has DNA Methylation Got to Do with It? Insects 2022, 13, 110. [Google Scholar] [CrossRef]
- Klose, R.J.; Bird, A.P. Genomic DNA Methylation: The Mark and Its Mediators. Trends Biochem. Sci. 2006, 31, 89–97. [Google Scholar] [CrossRef]
- Patel, T.N.; R., P.; Vashi, Y.; Bhattacharya, P. Toxic Impacts and Industrial Potential of Graphene. J. Environ. Sci. Health Part C 2020, 38, 269–297. [Google Scholar] [CrossRef]
- Chatterjee, N.; Kim, Y.; Yang, J.; Roca, C.P.; Joo, S.-W.; Choi, J. A Systems Toxicology Approach Reveals the Wnt-MAPK Crosstalk Pathway Mediated Reproductive Failure in Caenorhabditis Elegans Exposed to Graphene Oxide (GO) but Not to Reduced Graphene Oxide (RGO). Nanotoxicology 2017, 11, 76–86. [Google Scholar] [CrossRef]
- Bicho, R.C.; Roelofs, D.; Mariën, J.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Epigenetic Effects of (Nano)Materials in Environmental Species—Cu Case Study in Enchytraeus Crypticus. Env. Int. 2020, 136, 105447. [Google Scholar] [CrossRef]
- Bewick, A.J.; Vogel, K.J.; Moore, A.J.; Schmitz, R.J. Evolution of DNA Methylation across Insects. Mol. Biol. Evol. 2016, 34, 654–665. [Google Scholar] [CrossRef] [Green Version]
- Kausar, S.; Abbas, M.N.; Cui, H. A Review on the DNA Methyltransferase Family of Insects: Aspect and Prospects. Int. J. Biol. Macromol 2021, 186, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Lyko, F.; Ramsahoye, B.H.; Kashevsky, H.; Tudor, M.; Mastrangelo, M.-A.; Orr-Weaver, T.L.; Jaenisch, R. Mammalian (Cytosine-5) Methyltransferases Cause Genomic DNA Methylation and Lethality in Drosophila. Nat. Genet 1999, 23, 363–366. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-J.; Tang, L.-Y.; Reddy, M.N.; Shen, C.-K.J. DNA Methyltransferase Gene DDnmt2 and Longevity of Drosophila. J. Biol. Chem. 2005, 280, 861–864. [Google Scholar] [CrossRef] [PubMed]








| Effects 1 | DNA Damage Parameters 2 | |
|---|---|---|
| F | p | |
| Strain (1) | 9.715 | <0.0001 |
| Treatment (2) | 2.514 | 0.0152 |
| Stage (3) | 7.858 | <0.0001 |
| (1) × (2) | 1.203 | 0.3042 |
| (1) × (3) | 4.583 | <0.0001 |
| (2) × (3) | 1.298 | 0.2041 |
| (1) × (2) × (3) | 1.826 | 0.0317 |
| Effects 1 | DNA Damage Response Parameters 2 | |||
|---|---|---|---|---|
| pATM | pH2A.X | |||
| F | p | F | p | |
| Strain (1) | 0.237 | 0.6284 | 1.863 | 0.1774 |
| Treatment (2) | 0.147 | 0.8638 | 3.049 | 0.0548 |
| Stage (3) | 0.726 | 0.4882 | 3.020 | 0.0563 |
| (1) × (2) | 0.117 | 0.8900 | 3.300 | 0.0437 |
| (1) × (3) | 2.570 | 0.0849 | 3.583 | 0.0339 |
| (2) × (3) | 0.665 | 0.6190 | 0.935 | 0.4496 |
| (1) × (2) × (3) | 2.010 | 0.1044 | 0.957 | 0.4378 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flasz, B.; Dziewięcka, M.; Ajay, A.K.; Tarnawska, M.; Babczyńska, A.; Kędziorski, A.; Napora-Rutkowski, Ł.; Ziętara, P.; Świerczek, E.; Augustyniak, M. Age- and Lifespan-Dependent Differences in GO Caused DNA Damage in Acheta domesticus. Int. J. Mol. Sci. 2023, 24, 290. https://doi.org/10.3390/ijms24010290
Flasz B, Dziewięcka M, Ajay AK, Tarnawska M, Babczyńska A, Kędziorski A, Napora-Rutkowski Ł, Ziętara P, Świerczek E, Augustyniak M. Age- and Lifespan-Dependent Differences in GO Caused DNA Damage in Acheta domesticus. International Journal of Molecular Sciences. 2023; 24(1):290. https://doi.org/10.3390/ijms24010290
Chicago/Turabian StyleFlasz, Barbara, Marta Dziewięcka, Amrendra K. Ajay, Monika Tarnawska, Agnieszka Babczyńska, Andrzej Kędziorski, Łukasz Napora-Rutkowski, Patrycja Ziętara, Ewa Świerczek, and Maria Augustyniak. 2023. "Age- and Lifespan-Dependent Differences in GO Caused DNA Damage in Acheta domesticus" International Journal of Molecular Sciences 24, no. 1: 290. https://doi.org/10.3390/ijms24010290