
Transcriptomic and Metabolomic Analyses of the Effects of Exogenous Trehalose on Heat Tolerance in Wheat


Abstract
:1. Introduction
2. Results
2.1. Effects of Exogenous Trehalose on Phenotype and Physiological Indexes of Two Wheat Varieties under Heat Stress
2.2. Effects of Exogenous Trehalose on Chloroplast Ultrastructure of Two Wheat Varieties under Heat Stress
2.3. RNA Sequencing Data Quality Evaluation
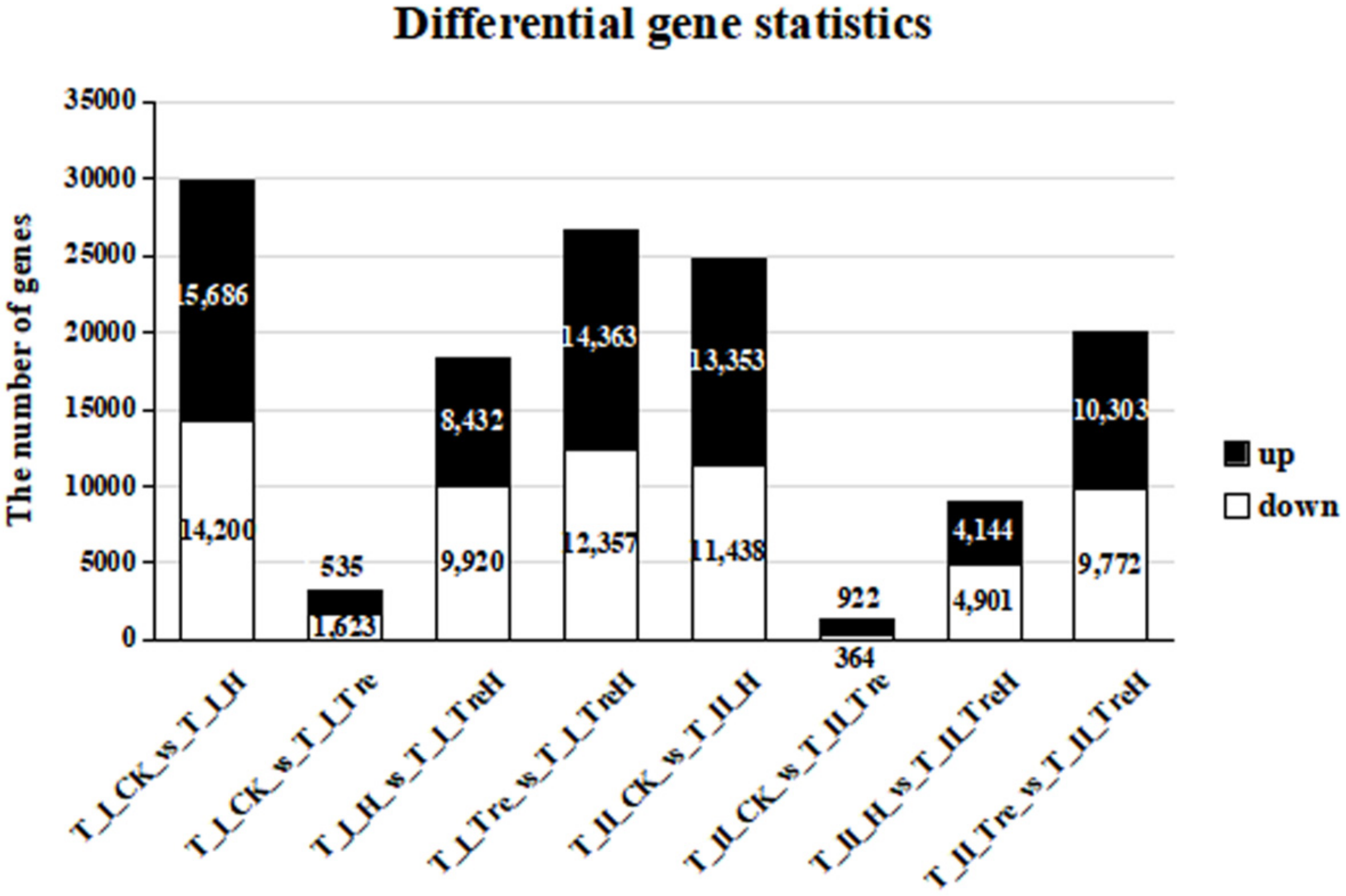
2.4. Statistical Analysis
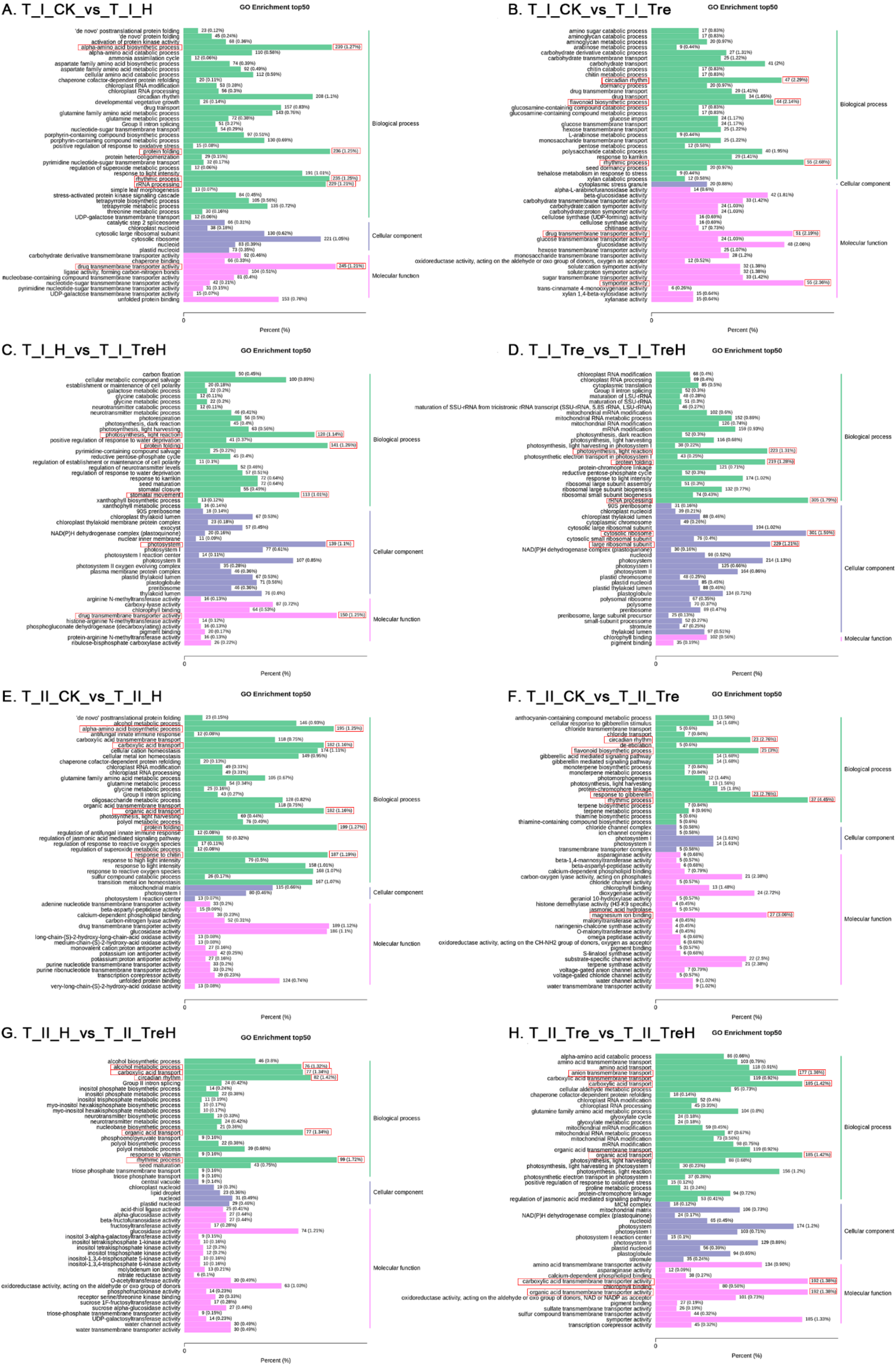
2.5. Gene Ontology (GO) Function Analysis of DEGs
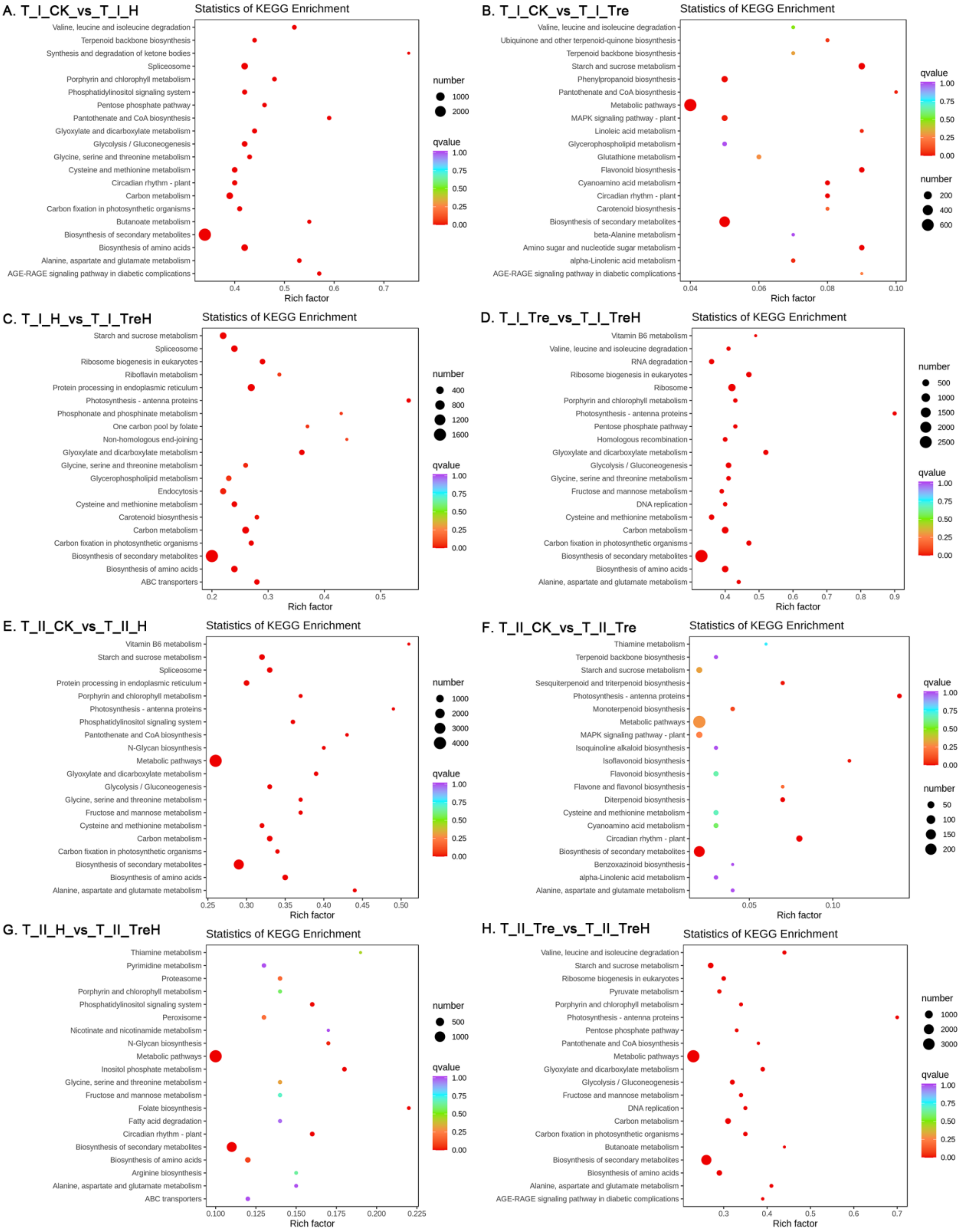
2.6. Enrichment of DEGs by Kyoto Encyclopedia of Genes and Genomes (KEGG) Metabolic Pathway
2.7. Analysis of Trehalose-Responsive Candidate Genes and Transcription Factors
2.8. Differential Metabolite Analysis
2.9. Enrichment Analysis of KEGG Metabolic Pathways of Differential Metabolites
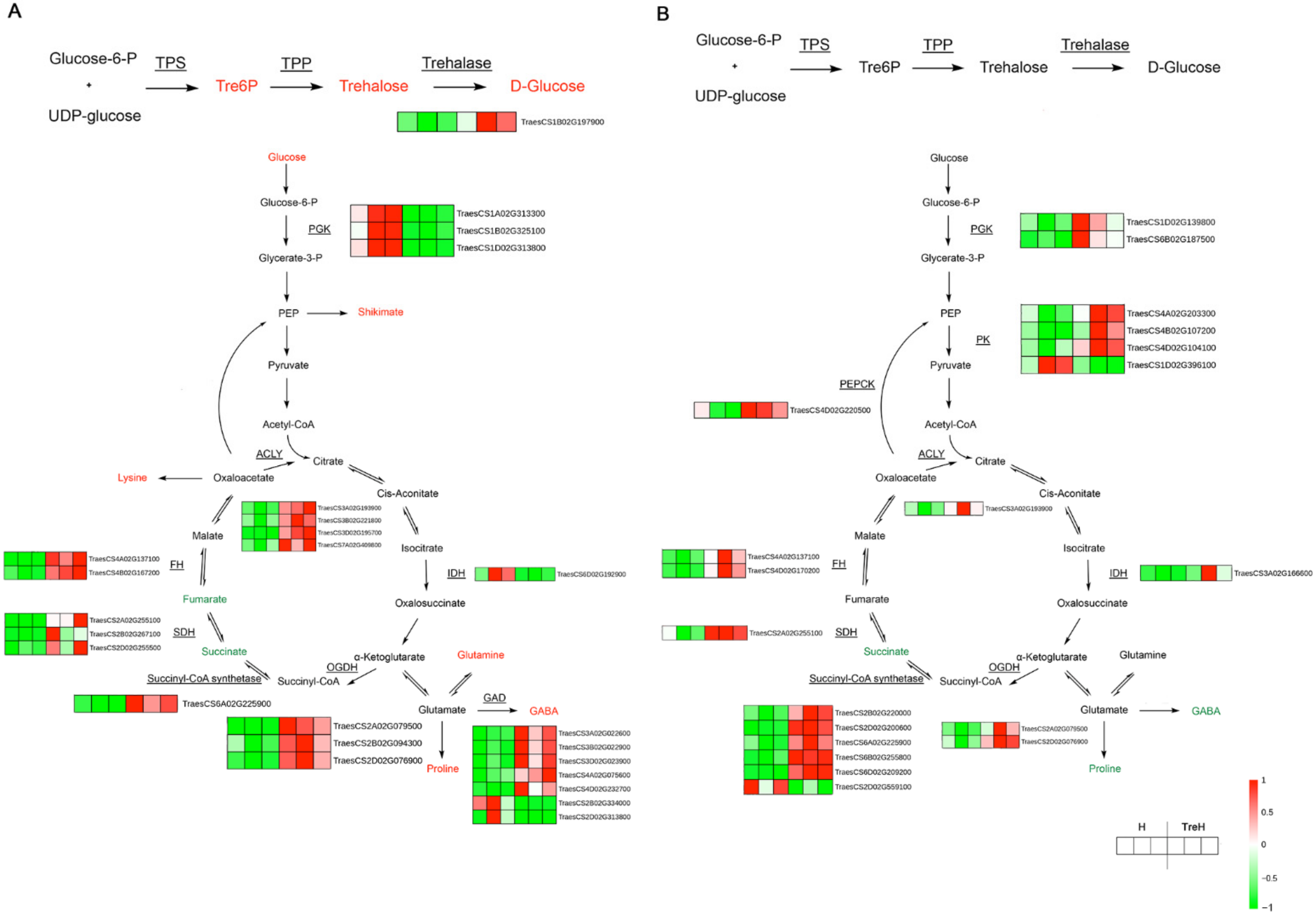
2.10. Integrated Analysis of DEGs and Differential Metabolites
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Stress Treatment
4.2. Phenotypic Analysis
4.3. Physiological Measurement of Wheat Seedlings under High Temperature Stress
4.4. Ultrastructure of Leaves
4.5. RNA Isolation, cDNA Library Construction, and Sequencing
4.6. Analyses of DEGs, KEGG and GO
4.7. Metabolite Extraction and Ultra Performance Liquid Chromatography–Tandem Mass Spectrometry (UPLC-MS/MS) Analysis
- (1)
- Column: Agilent SB-C18 1.8 µm, 2.1 mm × 100 mm;
- (2)
- Mobile phase: Phase A is ultra-pure water (0.1% formic acid added), phase B is acetonitrile (0.1% formic acid added);
- (3)
- Elution gradient: the proportion of phase B was 5% for 0.00 min, and the proportion of phase B linearly increased to 95% within 9.00 min, and maintained at 95% for 1min, 10.00–11.10 min, the proportion of phase B decreased to 5%, and was balanced at 5% for 14 min;
- (4)
- Flow rate 0.35 mL/min; Column temperature 40 °C; injection volume 4 μL.
4.8. RT-qPCR Analysis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2013: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2013. [Google Scholar]
- Zhao, D.Q.; Li, T.T.; Hao, Z.J.; Cheng, M.L.; Tao, J. Exogenous trehalose confers high temperature stress tolerance to herbaceous peony by enhancing antioxidant systems, activating photosynthesis, and protecting cell structure. Cell Stress Chaperones 2019, 24, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Djanaguiraman, M.; Narayanan, S.; Erdayani, E.; Prasad, P. Effects of high temperature stress during anthesis and grain filling periods on photosynthesis, lipids and grain yield in wheat. BMC Plant Biol. 2020, 20, 268. [Google Scholar] [CrossRef] [PubMed]
- Rurek, M.; Woyda-Ploszczyca, A.M.; Jarmuszkiewicz, W. Biogenesis of mitochondria in cauliflower (Brassica oleracea var. botrytis) curds subjected to temperature stress and recovery involves regulation of the complexome, respiratory chain activity, organellar translation and ultrastructure. Biochim. Biophys. Acta 2015, 1847, 399–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jespersen, D.; Huang, B. Proteins associated with heat-induced leaf senescence in creeping bentgrass as affected by foliar application of nitrogen, cytokinins, and an ethylene inhibitor. Proteomics 2015, 15, 798–812. [Google Scholar] [CrossRef]
- Mickelbart, M.V.; Hasegawa, P.M.; Bailey-Serres, J. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat. Rev. Genet. 2015, 16, 237–251. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Parker, J.E.; Ainsworth, E.A.; Oldroyd, G.; Schroeder, J.I. Schroeder, Genetic strategies for improving crop yields. Nature 2019, 575, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Zhang, J.; Gao, W.; Chen, Y.; Li, H.; Lawlor, D.W.; Paul, M.J.; Pan, W. Exogenous trehalose improves growth under limiting nitrogen through upregulation of nitrogen metabolism. BMC Plant Biol. 2017, 17, 247. [Google Scholar] [CrossRef] [Green Version]
- Ali, Q.; Ashraf, M.; Anwar, F.; Al-Qurainy, F. Trehalose-induced changes in seed oil composition and antioxidant potential of maize grown under drought stress. J. Am. Oil Chem. Soc. 2012, 89, 1485–1493. [Google Scholar] [CrossRef]
- Williams, B.; Njaci, I.; Moghaddam, L.; Long, H.; Dickman, M.B.; Zhang, X.; Mundree, S. Trehalose accumulation triggers autophagy during plant desiccation. PLoS Genet. 2015, 11, e1005705. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.S.; Lins, R.D.; Chandrasekhar, I.; Freitas, L.C.; Hünenberger, P.H. Interaction of the disaccharide trehalose with a phospholipid bilayer: A molecular dynamics study. Biophys. J. 2004, 86, 2273–2285. [Google Scholar] [CrossRef] [Green Version]
- Yu, W.; Zhao, R.; Wang, L.; Zhang, S.; Li, R.; Sheng, J.; Shen, L. ABA signaling rather than ABA metabolism is involved in trehalose-induced drought tolerance in tomato plants. Planta 2019, 250, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Iordachescu, M.; Imai, R. Trehalose biosynthesis in response to abiotic stresses. J. Integr. Plant Biol. 2008, 50, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Hespeels, B.; Li, X.; Flot, J.F.; Pigneur, L.M.; Malaisse, J.; Da Silva, C.; Van Doninck, K. Against all odds: Trehalose-6-phosphate synthase and trehalase genes in the bdelloid rotifer Adineta vaga were acquired by horizontal gene transfer and are upregulated during desiccation. PLoS ONE 2015, 10, e0131313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debast, S.; Nunes-Nesi, A.; Hajirezaei, M.R.; Hofmann, J.; Sonnewald, U.; Fernie, A.R.; Börnke, F. Altering trehalose-6-phosphate content in transgenic potato tubers affects tuber growth and alters responsiveness to hormones during sprouting. Plant Physiol. 2011, 156, 1754–1771. [Google Scholar] [CrossRef] [Green Version]
- Shi, Y.; Sun, H.; Wang, X.; Jin, W.; Chen, Q.; Yuan, Z.; Yu, H. Physiological and transcriptomic analyses reveal the molecular networks of responses induced by exogenous trehalose in plant. PLoS ONE 2019, 14, e0217204. [Google Scholar] [CrossRef] [Green Version]
- Asseng, S.; Ewert, F.; Martre, P.; Rötter, R.P.; Lobell, D.B.; Cammarano, D.; Kimball, B.A.; Ottman, M.J.; Wall, G.W.; White, J.W.; et al. Rising temperatures reduce global wheat production. Nat. Clim. Chang. 2015, 5, 143–147. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Burritt, D.J.; Gupta, A.; Tsujimoto, H.; Tran, L.P. Heat stress effects on source-sink relationships and metabolome dynamics in wheat. J. Exp. Bot. 2020, 71, 543–554. [Google Scholar] [CrossRef]
- Che-Othman, M.H.; Millar, A.H.; Taylor, N.L. Connecting salt stress signalling pathways with salinity-induced changes in mitochondrial metabolic processes in C3 plants. Plant Cell Environ. 2017, 40, 2875–2905. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, S.; Matsumoto, M.; Hakomori, Y.; Takagi, H.; Shimada, H.; Sakamoto, A. The purine metabolite allantoin enhances abiotic stress tolerance through synergistic activation of abscisic acid metabolism. Plant Cell Environ. 2014, 37, 1022–1036. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Wang, W.; Bittner, F.; Schmidt, N.; Berkey, R.; Zhang, L.; King, H.; Zhang, Y.; Feng, J.; Wen, Y.; et al. Dual and opposing roles of xanthine dehydrogenase in defense-associated reactive oxygen species metabolism in Arabidopsis. Plant Cell 2016, 28, 1108–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, R.; He, X.; Pan, X.; Shi, Q.; Wu, Z. Enhancing xanthine dehydrogenase activity is an effective way to delay leaf senescence and increase rice yield. Rice 2020, 13, 16. [Google Scholar] [CrossRef] [PubMed]
- Sukrong, S.; Yun, K.Y.; Stadler, P.; Kumar, C.; Facciuolo, T.; Moffatt, B.A.; Falcone, D.L. Improved growth and stress tolerance in the Arabidopsis oxt1 mutant triggered by altered adenine metabolism. Mol. Plant 2012, 5, 1310–1332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, H.J.; Baek, D.; Cho, H.M.; Lee, S.H.; Jin, B.J.; Yun, D.J.; Hong, Y.S.; Kim, M.C. Lignin biosynthesis genes play critical roles in the adaptation of Arabidopsis plants to high-salt stress. Plant Signal. Behav. 2019, 14, 1625697. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Wang, Y.; Xie, H.; Qiu, C.; Zhang, S.; Xiao, J.; Li, H.; Chen, L.; Li, X.; Ding, Z. Drought stress triggers proteomic changes involving lignin, flavonoids and fatty acids in tea plants. Sci. Rep. 2020, 10, 15504. [Google Scholar] [CrossRef] [PubMed]
- Le Gall, H.; Philippe, F.; Domon, J.M.; Gillet, F.; Pelloux, J.; Rayon, C. Cell wall metabolism in response to abiotic stress. Plants 2015, 4, 112–166. [Google Scholar] [CrossRef]
- Han, F.; Chen, H.; Li, X.J.; Yang, M.F.; Liu, G.S.; Shen, S.H. A comparative proteomic analysis of rice seedlings under various high-temperature stresses. Biochim. Biophys. Acta 2009, 1794, 1625–1634. [Google Scholar] [CrossRef]
- Gulen, H.; Eris, A. Effect of heat stress on peroxidase activity and total protein content in strawberry plants. Plant Sci. 2004, 166, 739–744. [Google Scholar] [CrossRef]
- Buer, C.S.; Kordbacheh, F.; Truong, T.T.; Hocart, C.H.; Djordjevic, M.A. Alteration of flavonoid accumulation patterns in transparent testa mutants disturbs auxin transport, gravity responses, and imparts long-term effects on root and shoot architecture. Planta 2013, 238, 171–189. [Google Scholar] [CrossRef]
- Gielwanowska, I.; Szczuka, E.; Bednara, J.; Górecki, R. Anatomical features and ultrastructure of Deschampsia antarctica (Poaceae) leaves from different growing habitats. Ann. Bot. 2005, 96, 1109–1119. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Y.; Smyth, G.K.; Shi, W. FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [PubMed] [Green Version]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Total | Down | Up |
|---|---|---|---|
| T_I_CK_vs_T_I_H | 29,886 | 14,200 | 15,686 |
| T_I_CK_vs_T_I_Tre | 3158 | 1623 | 1535 |
| T_I_H_vs_T_I_TreH | 18,352 | 9920 | 8432 |
| T_I_Tre_vs_T_I_TreH | 26,720 | 12,357 | 14,363 |
| T_II_CK_vs_T_II_H | 24,791 | 11,438 | 13,353 |
| T_II_CK_vs_T_II_Tre | 1286 | 364 | 922 |
| T_II_H_vs_T_II_TreH | 9045 | 4901 | 4144 |
| T_II_Tre_vs_T_II_TreH | 20,075 | 9772 | 10,303 |
| Group | Total | Down | Up |
|---|---|---|---|
| M_I_CK_vs_M_I_H | 256 | 113 | 143 |
| M_I_CK_vs_M_I_Tre | 53 | 34 | 19 |
| M_I_H_vs_M_I_TreH | 183 | 96 | 87 |
| M_I_Tre_vs_M_I_TreH | 248 | 109 | 139 |
| M_II_CK_vs_M_II_H | 218 | 105 | 113 |
| M_II_CK_vs_M_II_Tre | 117 | 65 | 52 |
| M_II_H_vs_M_II_TreH | 77 | 50 | 27 |
| M_II_Tre_vs_M_II_TreH | 240 | 123 | 117 |
| Group Name | Pretreatment with 0.5 mM Trehalose (Tre) | High-Temperature Stress Treatment (H) |
|---|---|---|
| Ⅰ_CK | ||
| Ⅰ_Tre | ✓ | |
| Ⅰ_H | ✓ | |
| Ⅰ_TreH | ✓ | ✓ |
| Ⅱ_CK | ||
| Ⅱ_Tre | ✓ | |
| Ⅱ_H | ✓ | |
| Ⅱ_TreH | ✓ | ✓ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, Y.; Wang, Y.; Xie, Y.; Gao, Y.; Li, W.; Lang, S. Transcriptomic and Metabolomic Analyses of the Effects of Exogenous Trehalose on Heat Tolerance in Wheat. Int. J. Mol. Sci. 2022, 23, 5194. https://doi.org/10.3390/ijms23095194
Luo Y, Wang Y, Xie Y, Gao Y, Li W, Lang S. Transcriptomic and Metabolomic Analyses of the Effects of Exogenous Trehalose on Heat Tolerance in Wheat. International Journal of Molecular Sciences. 2022; 23(9):5194. https://doi.org/10.3390/ijms23095194
Chicago/Turabian StyleLuo, Yin, Yue Wang, Yanyang Xie, Yamin Gao, Weiqiang Li, and Shuping Lang. 2022. "Transcriptomic and Metabolomic Analyses of the Effects of Exogenous Trehalose on Heat Tolerance in Wheat" International Journal of Molecular Sciences 23, no. 9: 5194. https://doi.org/10.3390/ijms23095194