
Applications of Genomic Tools in Plant Breeding: Crop Biofortification


Abstract
:1. Introduction
2. Exploring Biodiversity: Searching for Outstanding Material
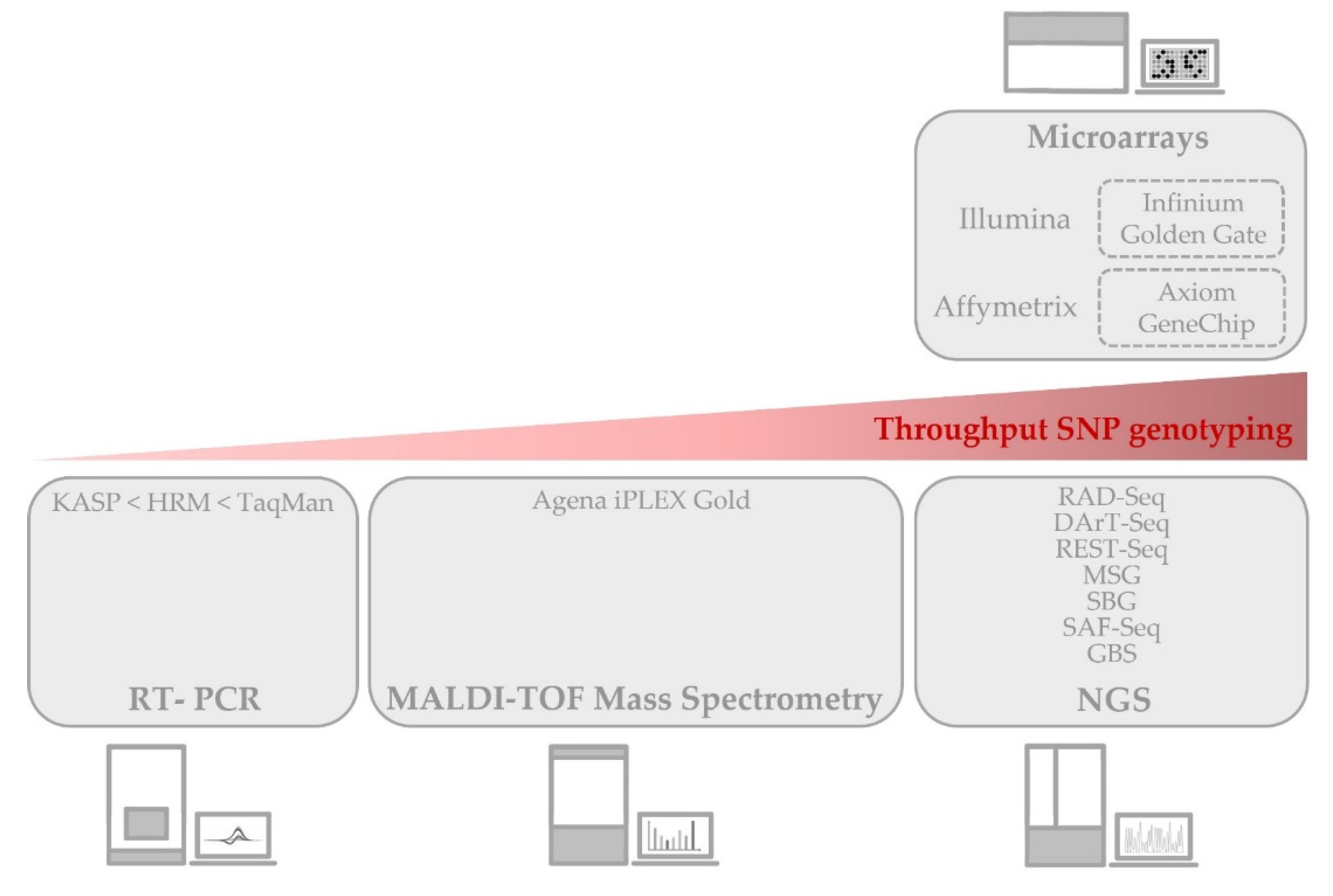
2.1. Genomic Diversity Enquired by SNP Genotyping
2.1.1. SNP Genotyping Microarrays
2.1.2. Real-Time PCR for SNP Genotyping
2.1.3. Mass Spectrometry SNP Genotyping
2.1.4. SNP Analysis by NGS
2.2. Nutritional and Phytochemical Profiles Assessed by Analytical Methods
2.2.1. Mass Spectrometry (MS)
2.2.2. Nuclear Magnetic Resonance (NMR)
3. Association between the Traits of Interest and the Genomic Regions: Fishing for Genes
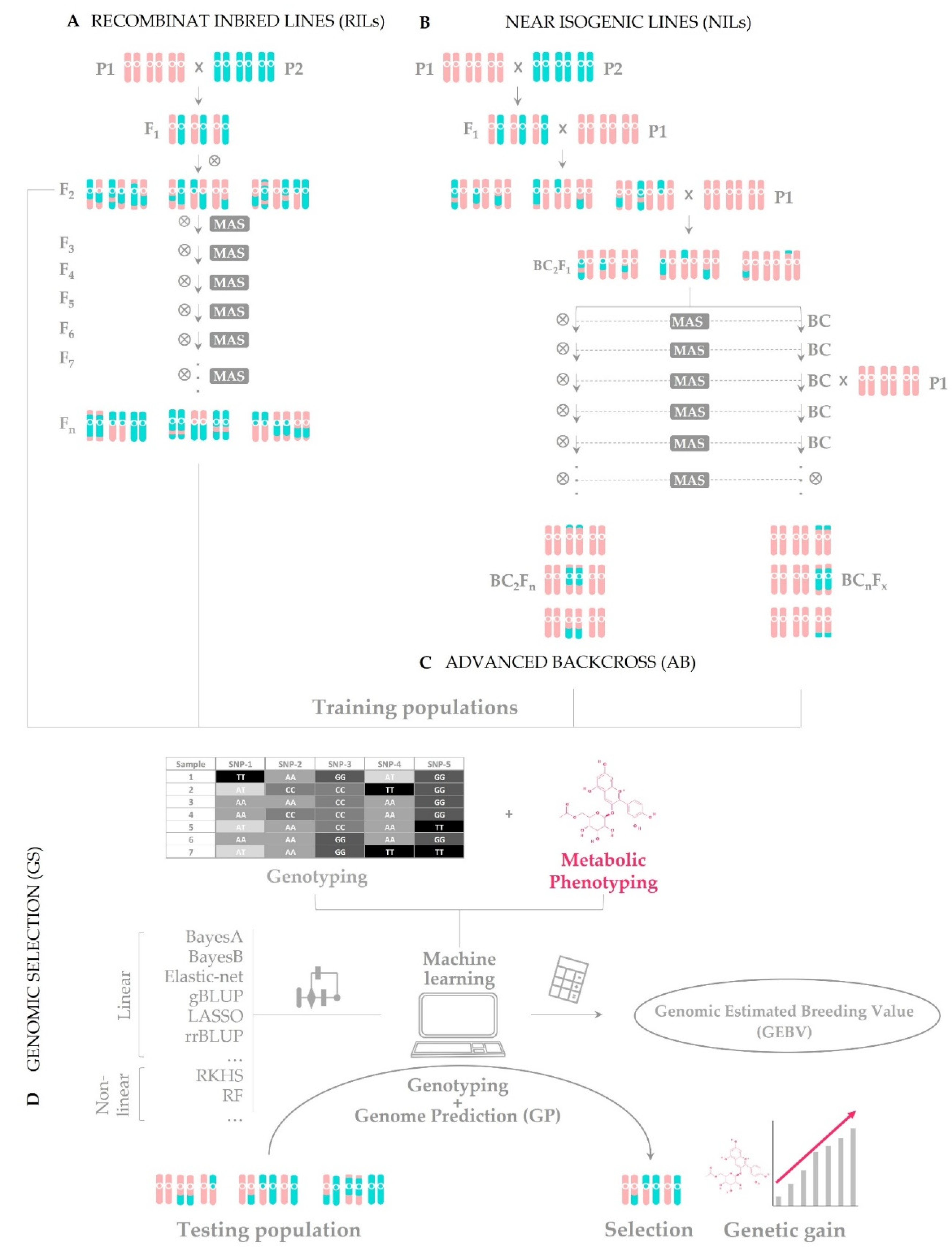
4. Introducing Allelic Variants to Biofortify Crops
4.1. Conventional Breeding Assisted by Genomic Tools
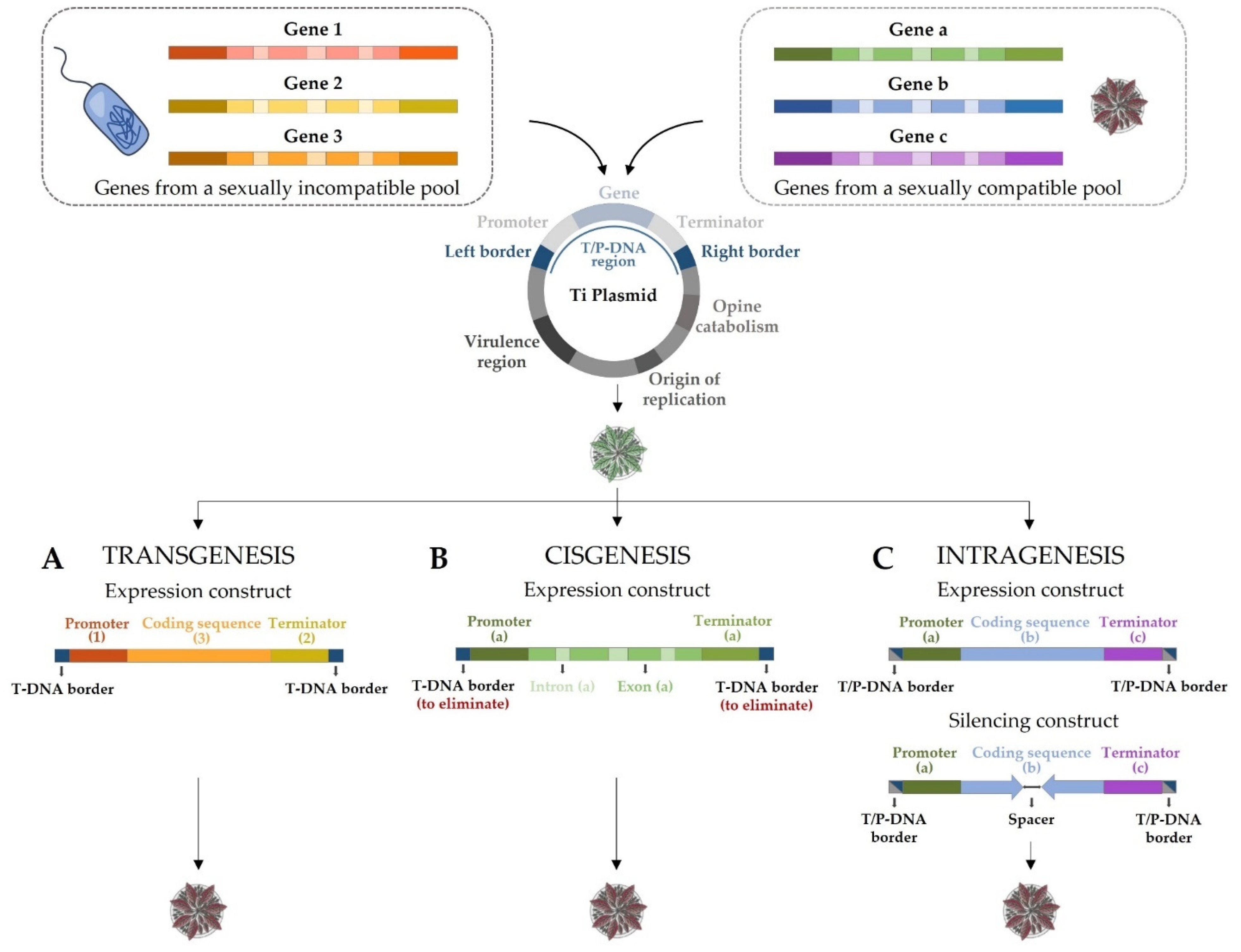
4.2. Modern Biotechnology Techniques
4.2.1. Transgenesis
4.2.2. Cisgenesis and Intragenesis
5. Regulation of Plant Breeding Methods
6. Future Perspectives
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FAO; IFAD; UNICEF; WPF; WHO. The State of Food Security and Nutrition in the World 2021. Trasforming Food System for Food Security, Improved Nutrition and Affordable Healthy Diets for All, 1st ed.; FAO: Rome, Italy, 2021; ISBN 978-92-5-134325-8. [Google Scholar]
- UNICEF. First call for children. In World Declaration and 1990–2000 Plan of Action on the Survival, Protection and Development of Children; UNICEF: New York, NY, USA, 1990. [Google Scholar]
- Medina-Lozano, I.; Díaz, A. Nutritional Value and Phytochemical Content of Crop Landraces and Traditional Varieties. In Landraces—Traditional Variety and Natural Breed; Elkelish, A., Ed.; IntechOpen: Rijeka, Croatia, 2021; pp. 95–116. ISBN 978-1-83968-718-1. [Google Scholar]
- Paul, S.; Ali, N.; Datta, S.K.; Datta, K. Development of an Iron-enriched High-yieldings Indica Rice Cultivar by Introgression of A High-iron Trait from Transgenic Iron-biofortified Rice. Plant Foods Hum. Nutr. 2014, 69, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Neelam, K.; Rawat, N.; Tiwari, V.K.; Kumar, S.; Chhuneja, P.; Singh, K.; Randhawa, G.S.; Dhaliwal, H.S. Introgression of group 4 and 7 chromosomes of Ae. peregrina in wheat enhances grain iron and zinc density. Mol. Breed. 2011, 28, 623–634. [Google Scholar] [CrossRef]
- Haynes, K.G.; Yencho, G.C.; Clough, M.E.; Henninger, M.R.; Sterrett, S.B. Genetic Variation for Potato Tuber Micronutrient Content and Implications for Biofortification of Potatoes to Reduce Micronutrient Malnutrition. Am. J. Potato Res. 2012, 89, 192–198. [Google Scholar] [CrossRef]
- Blair, M.W.; Astudillo, C.; Grusak, M.A.; Graham, R.; Beebe, S.E. Inheritance of seed iron and zinc concentrations in common bean (Phaseolus vulgaris L.). Mol. Breed. 2009, 23, 197–207. [Google Scholar] [CrossRef]
- Aluru, M.R.; Rodermel, S.R.; Reddy, M.B. Genetic modification of low phytic acid 1-1 maize to enhance iron content and bioavailability. J. Agric. Food Chem. 2011, 59, 12954–12962. [Google Scholar] [CrossRef] [PubMed]
- Beasley, J.T.; Bonneau, J.P.; Sánchez-Palacios, J.T.; Moreno-Moyano, L.T.; Callahan, D.L.; Tako, E.; Glahn, R.P.; Lombi, E.; Johnson, A.A.T. Metabolic engineering of bread wheat improves grain iron concentration and bioavailability. Plant Biotechnol. J. 2019, 17, 1514–1526. [Google Scholar] [CrossRef] [PubMed]
- Narayanan, N.; Beyene, G.; Chauhan, R.D.; Gaitán-Solís, E.; Gehan, J.; Butts, P.; Siritunga, D.; Okwuonu, I.; Woll, A.; Jiménez-Aguilar, D.M.; et al. Biofortification of field-grown cassava by engineering expression of an iron transporter and ferritin. Nat. Biotechnol. 2019, 37, 144–151. [Google Scholar] [CrossRef]
- Nozoye, T.; Otani, M.; Senoura, T.; Nakanishi, H.; Nishizawa, N.K. Overexpression of barley nicotianamine synthase 1 confers tolerance in the sweet potato to iron deficiency in calcareous soil. Plant Soil 2017, 418, 75–88. [Google Scholar] [CrossRef]
- Connorton, J.M.; Jones, E.R.; Rodríguez-Ramiro, I.; Fairweather-Tait, S.; Uauy, C.; Balk, J. Wheat Vacuolar Iron Transporter TaVIT2 Transports Fe and Mn and Is Effective for Biofortification. Plant Physiol. 2017, 174, 2434–2444. [Google Scholar] [CrossRef] [Green Version]
- Menkir, A.; Maziya-Dixon, B.; Mengesha, W.; Rocheford, T.; Alamu, E.O. Accruing genetic gain in pro-vitamin A enrichment from harnessing diverse maize germplasm. Euphytica 2017, 213, 105. [Google Scholar] [CrossRef]
- Hossain, F.; Muthusamy, V.; Zunjare, R.U. Molecular Breeding for Development of Biofortified Maize Hybrids in India. In Proceedings of the Extended Summaries: 13th Asian Maize Conference on and Expert Consultation on Maize for Food, Feed, Nutrition and Environmental Security, Ludhiana, India, 8–10 October 2018; pp. 220–230. [Google Scholar]
- Ceballos, H.; Morante, N.; Sanchez, T.; Ortiz, D.; Aragón, I.; Chávez, A.; Pizarro, M.; Calle, F.; Dominique, D. Rapid Cycling Recurrent Selection for Increased Carotenoids Content in Cassava Roots. Crop Sci. 2013, 53, 2342–2351. [Google Scholar] [CrossRef] [Green Version]
- Rezaei, M.; Deokar, A.; Arganosa, G.; Roorkiwal, M.; Pandey, S.; Warkentin, T.; Varshney, R.; Tar’an, B. Mapping Quantitative Trait Loci for Carotenoid Concentration in Three F Populations of Chickpea. Plant Genome 2019, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, X.; Al-Babili, S.; Klöti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broad, R.C.; Bonneau, J.P.; Beasley, J.T.; Roden, S.; Sadowski, P.; Jewell, N.; Brien, C.; Berger, B.; Tako, E.; Glahn, R.P.; et al. Effect of Rice GDP-L-Galactose Phosphorylase Constitutive Overexpression on Ascorbate Concentration, Stress Tolerance, and Iron Bioavailability in Rice. Front. Plant Sci. 2020, 11, 595439. [Google Scholar] [CrossRef] [PubMed]
- Aluru, M.; Xu, Y.; Guo, R.; Wang, Z.; Li, S.; White, W.; Wang, K.; Rodermel, S. Generation of transgenic maize with enhanced provitamin A content. J. Exp. Bot. 2008, 59, 3551–3562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Q.; Wang, K.; Liu, X.; Riaz, B.; Jiang, L.; Wan, X.; Ye, X.; Zhang, C. Improved folate accumulation in genetically modified maize and wheat. J. Exp. Bot. 2019, 70, 1539–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Lepeleire, J.; Strobbe, S.; Verstraete, J.; Blancquaert, D.; Ambach, L.; Visser, R.G.F.; Stove, C.; Van Der Straeten, D. Folate Biofortification of Potato by Tuber-Specific Expression of Four Folate Biosynthesis Genes. Mol. Plant 2018, 11, 175–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yazdani, M.; Sun, Z.; Yuan, H.; Zeng, S.; Thannhauser, T.W.; Vrebalov, J.; Ma, Q.; Xu, Y.; Fei, Z.; Van Eck, J.; et al. Ectopic expression of ORANGE promotes carotenoid accumulation and fruit development in tomato. Plant Biotechnol. J. 2019, 17, 33–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Butelli, E.; Alseekh, S.; Tohge, T.; Rallapalli, G.; Luo, J.; Kawar, P.G.; Hill, L.; Santino, A.; Fernie, A.R.; et al. Multi-level engineering facilitates the production of phenylpropanoid compounds in tomato. Nat. Commun. 2015, 6, 8635. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.-Y.; Khanna, H.; Kleidon, J.; Hoang, P.; Geijskes, J.; Daniells, J.; Zaplin, E.; Rosenberg, Y.; James, A.; Mlalazi, B.; et al. Golden bananas in the field: Elevated fruit pro-vitamin A from the expression of a single banana transgene. Plant Biotechnol. J. 2017, 15, 520–532. [Google Scholar] [CrossRef] [Green Version]
- Ramírez Rivera, N.G.; García-Salinas, C.; Aragão, F.J.L.; Díaz de la Garza, R.I. Metabolic engineering of folate and its precursors in Mexican common bean (Phaseolus vulgaris L.). Plant Biotechnol. J. 2016, 14, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Lydiate, D.J.; Young, L.W.; Schäfer, U.A.; Hannoufa, A. Enhancing the carotenoid content of Brassica napus seeds by downregulating lycopene epsilon cyclase. Transgenic Res. 2008, 17, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Zhu, W.; Tang, R.; Cai, J.; Chen, M.; Yang, Q. Over-expression of StLCYb increases β-carotene accumulation in potato tubers. Plant Biotechnol. Rep. 2016, 10, 95–104. [Google Scholar] [CrossRef]
- Davuluri, G.R.; van Tuinen, A.; Fraser, P.D.; Manfredonia, A.; Newman, R.; Burgess, D.; Brummell, D.A.; King, S.R.; Palys, J.; Uhlig, J.; et al. Fruit-specific RNAi-mediated suppression of DET1 enhances carotenoid and flavonoid content in tomatoes. Nat. Biotechnol. 2005, 23, 890–895. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Gruissem, W.; Bhullar, N.K. Single genetic locus improvement of iron, zinc and β-carotene content in rice grains. Sci. Rep. 2017, 7, 6883. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-López, N.; Haslam, R.P.; Venegas-Calerón, M.; Larson, T.R.; Graham, I.A.; Napier, J.A.; Sayanova, O. The synthesis and accumulation of stearidonic acid in transgenic plants: A novel source of “heart-healthy” omega-3 fatty acids. Plant Biotechnol. J. 2009, 7, 704–716. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Wu, G.; Vrinten, P.; Falk, K.; Bauer, J.; Qiu, X. Towards the production of high levels of eicosapentaenoic acid in transgenic plants: The effects of different host species, genes and promoters. Transgenic Res. 2010, 19, 221–229. [Google Scholar] [CrossRef]
- Jing, G.; Tang, D.; Yao, Y.; Su, Y.; Shen, Y.; Bai, Y.; Jing, W.; Zhang, Q.; Lin, F.; Guo, D.; et al. Seed specifically over-expressing DGAT2A enhances oil and linoleic acid contents in soybean seeds. Biochem. Biophys. Res. Commun. 2021, 568, 143–150. [Google Scholar] [CrossRef]
- Liu, Q.; Guo, Q.; Akbar, S.; Zhi, Y.; El Tahchy, A.; Mitchell, M.; Li, Z.; Shrestha, P.; Vanhercke, T.; Ral, J.P.; et al. Genetic enhancement of oil content in potato tuber (Solanum tuberosum L.) through an integrated metabolic engineering strategy. Plant Biotechnol. J. 2017, 15, 56–67. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.-S.; Sun-Hyung, J.; Oehrle, N.W.; Jez, J.M.; Krishnan, H.B. Overexpression of ATP sulfurylase improves the sulfur amino acid content, enhances the accumulation of Bowman-Birk protease inhibitor and suppresses the accumulation of the β-subunit of β-conglycinin in soybean seeds. Sci. Rep. 2020, 10, 14989. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; He, X.; Luo, Y.; Ma, L.; Tang, X.; Huang, K. Nutritional assessment of transgenic lysine-rich maize compared with conventional quality protein maize. J. Sci. Food Agric. 2013, 93, 1049–1054. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Q.; Zhang, C.Q.; Chan, M.L.; Zhao, D.S.; Chen, J.Z.; Wang, Q.; Li, Q.F.; Yu, H.X.; Gu, M.H.; Sun, S.S.M.; et al. Biofortification of rice with the essential amino acid lysine: Molecular characterization, nutritional evaluation, and field performance. J. Exp. Bot. 2016, 67, 4285–4296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, M.; Zhao, S.; Zhang, Y.; Yin, H.; Peng, X.; Cheng, Z.; Yang, Z.; Zheng, J. Production of Marker-Free Transgenic Rice (Oryza sativa L.) with Improved Nutritive Quality Expressing AmA1. Iran. J. Biotechnol. 2017, 15, 102–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, M.; Chawla, M.; Chunduri, V.; Kumar, R.; Sharma, S.; Sharma, N.K.; Kaur, N.; Kumar, A.; Mundey, J.K.; Saini, M.K.; et al. Transfer of grain colors to elite wheat cultivars and their characterization. J. Cereal Sci. 2016, 71, 138–144. [Google Scholar] [CrossRef]
- Ingallina, C.; Maccelli, A.; Spano, M.; Di Matteo, G.; Di Sotto, A.; Giusti, A.M.; Vinci, G.; Di Giacomo, S.; Rapa, M.; Ciano, S.; et al. Chemico-biological characterization of torpedino di fondi® tomato fruits: A comparison with san marzano cultivar at two ripeness stages. Antioxidants 2020, 9, 1027. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Zeng, D.; Yu, S.; Cui, C.; Li, J.; Li, H.; Chen, J.; Zhang, R.; Zhao, X.; Chen, L.; et al. From Golden Rice to aSTARice: Bioengineering Astaxanthin Biosynthesis in Rice Endosperm. Mol. Plant 2018, 11, 1440–1448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krens, F.A.; Schaart, J.G.; van der Burgh, A.M.; TinnenbroekCapel, I.E.M.; Groenwold, R.; Kodde, L.P.; Broggini, G.A.L.; Gessler, C.; Schouten, H.J. Cisgenic apple trees; development, characterization, and performance. Front. Plant Sci. 2015, 6, 286. [Google Scholar] [CrossRef] [Green Version]
- Hoddinott, J.F.; Rosegrant, M.W.; Torero, M. Investments to reduce hunger and undernutrition. In Global Problems, Smart Solutions; Lomborg, B., Ed.; Cambridge University Press: Cambridge, UK, 2013; pp. 332–367. [Google Scholar]
- Graham, R.D.; Welch, R.M. Breeding for Staple-Food Crops with High Micronutrient Density: Working Papers on Agricultural Strategies for Micronutrients, 3rd ed.; International Food Policy Institute: Washington, DC, USA, 1996. [Google Scholar]
- Graham, R.D.; Welch, R.M.; Bouis, H.E. Addressing Micronutrient Malnutrition Through Enhancing the Nutritional Quality of Staple Foods: Principles, Perspectives and Knowledge Gaps. Adv. Agron. 2001, 70, 77–142. [Google Scholar] [CrossRef]
- United Nations; Department of Economic and Social; Affairs, P.D. World Population Prospects, Medium Prognosis; The 2019 Revision: New York, NY, USA, 2019. [Google Scholar]
- Wortmann, L.; Enneking, U.; Daum, D. German Consumers’ Attitude Towards Selenium-Biofortified Apples and Acceptance of Related Nutrition and Health Claims. Nutrients 2018, 10, 190. [Google Scholar] [CrossRef] [Green Version]
- Welk, A.K.; Kleine-kalmer, R.; Daum, D.; Enneking, U. Consumer Acceptance and Market Potential of Iodine-Biofortified Fruit and Vegetables in Germany. Nutrients 2021, 13, 4198. [Google Scholar] [CrossRef] [PubMed]
- Adekambi, S.A.; Okello, J.J.; Rajendran, S.; Acheremu, K.; Carey, E.E.; Low, J.; Abidin, P.E. Effect of varietal attributes on the adoption of an orange-fleshed sweetpotato variety in Upper East and Northern Ghana. Outlook Agric. 2020, 49, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Gannon, B.; Kaliwile, C.; Arscott, S.A.; Schmaelzle, S.; Chileshe, J.; Kalungwana, N.; Mosonda, M.; Pixley, K.; Masi, C.; Tanumihardjo, S.A. Biofortified orange maize is as efficacious as a vitamin A supplement in Zambian children even in the presence of high liver reserves of vitamin A: A community-based, randomized placebo-controlled trial. Am. J. Clin. Nutr. 2014, 100, 1541–1550. [Google Scholar] [CrossRef] [PubMed]
- Komar, A.A. Single Nucleotide Polymorphisms. Methods Mol. Biol. 2009, 578, 23–39. [Google Scholar]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Unterseer, S.; Bauer, E.; Haberer, G.; Seidel, M.; Knaak, C.; Ouzunova, M.; Meitinger, T.; Strom, T.M.; Fries, R.; Pausch, H.; et al. A powerful tool for genome analysis in maize: Development and evaluation of the high density 600 k SNP genotyping array. BMC Genomics 2014, 15, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, C.; Ren, Y.; Jian, Y.; Guo, Z.; Zhang, Y.; Xie, C.; Fu, J.; Wang, H.; Wang, G.; Xu, Y.; et al. Development of a maize 55 K SNP array with improved genome coverage for molecular breeding. Mol. Breed. 2017, 37, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tung, C.W.; Zhao, K.; Wright, M.H.; Ali, M.L.; Jung, J.; Kimball, J.; Tyagi, W.; Thomson, M.J.; McNally, K.; Leung, H.; et al. Development of a research platform for dissecting phenotype-genotype associations in rice (Oryza spp.). Rice 2010, 3, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Jayaswal, P.K.; Panda, K.; Mandal, P.; Kumar, V.; Singh, B.; Mishra, S.; Singh, Y.; Singh, R.; Rai, V.; et al. Single-copy gene based 50 K SNP chip for genetic studies and molecular breeding in rice. Sci. Rep. 2015, 5, 1–9. [Google Scholar] [CrossRef]
- Bauer, E.; Schmutzer, T.; Barilar, I.; Mascher, M.; Gundlach, H.; Martis, M.M.; Twardziok, S.O.; Hackauf, B.; Gordillo, A.; Wilde, P.; et al. Towards a whole-genome sequence for rye (Secale cereale L.). Plant J. 2017, 89, 853–869. [Google Scholar] [CrossRef] [Green Version]
- Winfield, M.O.; Allen, A.M.; Burridge, A.J.; Barker, G.L.A.; Benbow, H.R.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; Scopes, G.; et al. High-density SNP genotyping array for hexaploid wheat and its secondary and tertiary gene pool. Plant Biotechnol. J. 2016, 14, 1195–1206. [Google Scholar] [CrossRef] [PubMed]
- Allen, A.M.; Winfield, M.O.; Burridge, A.J.; Downie, R.C.; Benbow, H.R.; Barker, G.L.A.; Wilkinson, P.A.; Coghill, J.; Waterfall, C.; Davassi, A.; et al. Characterization of a Wheat Breeders’ Array suitable for high-throughput SNP genotyping of global accessions of hexaploid bread wheat (Triticum aestivum). Plant Biotechnol. J. 2017, 15, 390–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karadi, A.; Samineni, S.; Sajja, S.; Sharma, M.; Thudi, M.; Mallikarjuna, B.P.; Viswanatha, K.P.; Varshney, R.K.; Gaur, P.M. Molecular mapping of dry root rot resistance genes in chickpea (Cicer arietinum L.). Euphytica 2021, 217, 1–13. [Google Scholar] [CrossRef]
- Stoffel, K.; van Leeuwen, H.; Kozik, A.; Caldwell, D.; Ashrafi, H.; Cui, X.; Tan, X.; Hill, T.; Reyes-Chin-Wo, S.; Truco, M.J.; et al. Development and application of a 6.5 million feature Affymetrix Genechip® for massively parallel discovery of single position polymorphisms in lettuce (Lactuca spp.). BMC Genomics 2012, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vos, P.G.; Uitdewilligen, J.G.A.M.L.; Voorrips, R.E.; Visser, R.G.F.; van Eck, H.J. Development and analysis of a 20K SNP array for potato (Solanum tuberosum): An insight into the breeding history. Theor. Appl. Genet. 2015, 128, 2387–2401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.G.; Jeong, N.; Kim, J.H.; Lee, K.; Kim, K.H.; Pirani, A.; Ha, B.K.; Kang, S.T.; Park, B.S.; Moon, J.K.; et al. Development, validation and genetic analysis of a large soybean SNP genotyping array. Plant J. 2015, 81, 625–636. [Google Scholar] [CrossRef]
- Bassil, N.V.; Davis, T.M.; Zhang, H.; Ficklin, S.; Mittmann, M.; Webster, T.; Mahoney, L.; Wood, D.; Alperin, E.S.; Rosyara, U.R.; et al. Development and preliminary evaluation of a 90 K Axiom® SNP array for the allo-octoploid cultivated strawberry Fragaria × ananassa. BMC Genomics 2015, 16, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Bianco, L.; Cestaro, A.; Linsmith, G.; Muranty, H.; Denancé, C.; Théron, A.; Poncet, C.; Micheletti, D.; Kerschbamer, E.; Di Pierro, E.A.; et al. Development and validation of the Axiom®Apple480K SNP genotyping array. Plant J. 2016, 86, 62–74. [Google Scholar] [CrossRef] [Green Version]
- Pandey, M.K.; Agarwal, G.; Kale, S.M.; Clevenger, J.; Nayak, S.N.; Sriswathi, M.; Chitikineni, A.; Chavarro, C.; Chen, X.; Upadhyaya, H.D.; et al. Development and Evaluation of a High Density Genotyping “Axiom-Arachis” Array with 58 K SNPs for Accelerating Genetics and Breeding in Groundnut. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Comadran, J.; Kilian, B.; Russell, J.; Ramsay, L.; Stein, N.; Ganal, M.; Shaw, P.; Bayer, M.; Thomas, W.; Marshall, D.; et al. Natural variation in a homolog of Antirrhinum CENTRORADIALIS contributed to spring growth habit and environmental adaptation in cultivated barley. Nat. Genet. 2012, 44, 1388–1391. [Google Scholar] [CrossRef]
- Ganal, M.W.; Durstewitz, G.; Polley, A.; Bérard, A.; Buckler, E.S.; Charcosset, A.; Clarke, J.D.; Graner, E.M.; Hansen, M.; Joets, J.; et al. A Large Maize (Zea mays L.) SNP Genotyping Array: Development and Germplasm Genotyping, and Genetic Mapping to Compare with the B73 Reference Genome. PLoS ONE 2011, 6, e28334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousselle, Y.; Jones, E.; Charcosset, A.; Moreau, P.; Robbins, K.; Stich, B.; Knaak, C.; Flament, P.; Karaman, Z.; Martinant, J.P.; et al. Study on Essential Derivation in Maize: III. Selection and Evaluation of a Panel of Single Nucleotide Polymorphism Loci for Use in European and North American Germplasm. Crop Sci. 2015, 55, 1170–1180. [Google Scholar] [CrossRef]
- Tinker, N.A.; Chao, S.; Lazo, G.R.; Oliver, R.E.; Huang, Y.; Poland, J.A.; Jellen, E.N.; Maughan, P.J.; Kilian, A.; Jackson, E.W. A SNP Genotyping Array for Hexaploid Oat. Plant Genome 2014, 7, 1–8. [Google Scholar] [CrossRef]
- Chen, W.; Gao, Y.; Xie, W.; Gong, L.; Lu, K.; Wang, W.; Li, Y.; Liu, X.; Zhang, H.; Dong, H.; et al. Genome-wide association analyses provide genetic and biochemical insights into natural variation in rice metabolism. Nat. Genet. 2014, 46, 714–721. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Xie, W.; Li, J.; Zhou, F.; Zhang, Q. A whole-genome SNP array (RICE6K) for genomic breeding in rice. Plant Biotechnol. J. 2014, 12, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, C.R.; Chao, S.; Wang, S.; Huang, B.E.; Stephen, S.; Kiani, S.; Forrest, K.; Saintenac, C.; Brown-Guedira, G.L.; Akhunova, A.; et al. Genome-wide comparative diversity uncovers multiple targets of selection for improvement in hexaploid wheat landraces and cultivars. Proc. Natl. Acad. Sci. USA 2013, 110, 8057–8062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, W.E.; Higgins, E.E.; Plieske, J.; Wieseke, R.; Sidebottom, C.; Khedikar, Y.; Batley, J.; Edwards, D.; Meng, J.; Li, R.; et al. A high-density SNP genotyping array for Brassica napus and its ancestral diploid species based on optimised selection of single-locus markers in the allotetraploid genome. Theor. Appl. Genet. 2016, 129, 1887–1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bachlava, E.; Taylor, C.A.; Tang, S.; Bowers, J.E.; Mandel, J.R.; Burke, J.M.; Knapp, S.J. SNP Discovery and Development of a High-Density Genotyping Array for Sunflower. PLoS ONE 2012, 7, e29814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livaja, M.; Unterseer, S.; Erath, W.; Lehermeier, C.; Wieseke, R.; Plieske, J.; Polley, A.; Luerßen, H.; Wieckhorst, S.; Mascher, M.; et al. Diversity analysis and genomic prediction of Sclerotinia resistance in sunflower using a new 25 K SNP genotyping array. Theor. Appl. Genet. 2016, 129, 317–329. [Google Scholar] [CrossRef] [PubMed]
- Close, T.J.; Lucas, M.R.; Muñoz-Amatriain, M.; Mirebrahim, H.; Wanamaker, S.; Barkley, N.A.; Clair, S.S.; Guo, Y.-N.; Lo, S.; Huynh, B.L. A new SNP-genotyping resource for cowpea and its deployment for breeding. In Proceedings of the Plant and Animal Genome Conference, San Diego, CA, USA, 10–14 January 2015; Volume 23, p. P0784. [Google Scholar]
- Hamilton, J.P.; Hansey, C.N.; Whitty, B.R.; Stoffel, K.; Massa, A.N.; Van Deynze, A.; De Jong, W.S.; Douches, D.S.; Buell, C.R. Single nucleotide polymorphism discovery in elite North American potato germplasm. BMC Genomics 2011, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sim, S.C.; Durstewitz, G.; Plieske, J.; Wieseke, R.; Ganal, M.W.; van Deynze, A.; Hamilton, J.P.; Buell, C.R.; Causse, M.; Wijeratne, S.; et al. Development of a Large SNP Genotyping Array and Generation of High-Density Genetic Maps in Tomato. PLoS ONE 2012, 7, e40563. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; Hyten, D.L.; Jia, G.; Quigley, C.V.; Fickus, E.W.; Nelson, R.L.; Cregan, P.B. Development and Evaluation of SoySNP50K, a High-Density Genotyping Array for Soybean. PLoS ONE 2013, 8, e54985. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, L.; Cestaro, A.; Sargent, D.J.; Banchi, E.; Derdak, S.; Di Guardo, M.; Salvi, S.; Jansen, J.; Viola, R.; Gut, I.; et al. Development and validation of a 20K Single Nucleotide Polymorphism (SNP) whole genome genotyping array for apple (Malus × domestica Borkh). PLoS ONE 2014, 9, e110377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagné, D.; Crowhurst, R.N.; Troggio, M.; Davey, M.W.; Gilmore, B.; Lawley, C.; Vanderzande, S.; Hellens, R.P.; Kumar, S.; Cestaro, A.; et al. Genome-Wide SNP Detection, Validation, and Development of an 8K SNP Array for Apple. PLoS ONE 2012, 7, e31745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peace, C.; Bassil, N.; Main, D.; Ficklin, S.; Rosyara, U.R.; Stegmeir, T.; Sebolt, A.; Gilmore, B.; Lawley, C.; Mockler, T.C.; et al. Development and Evaluation of a Genome-Wide 6K SNP Array for Diploid Sweet Cherry and Tetraploid Sour Cherry. PLoS ONE 2012, 7, e48305. [Google Scholar] [CrossRef] [PubMed]
- Verde, I.; Bassil, N.; Scalabrin, S.; Gilmore, B.; Lawley, C.T.; Gasic, K.; Micheletti, D.; Rosyara, U.R.; Cattonaro, F.; Vendramin, E.; et al. Development and Evaluation of a 9k SNP Array for Peach by Internationally Coordinated SNP Detection and Validation in Breeding Germplasm. PLoS ONE 2012, 7, e35668. [Google Scholar] [CrossRef]
- Micheletti, D.; Dettori, M.T.; Micali, S.; Aramini, V.; Pacheco, I.; Da Silva Linge, C.; Foschi, S.; Banchi, E.; Barreneche, T.; Quilot-Turion, B.; et al. Whole-Genome Analysis of Diversity and SNP-Major Gene Association in Peach Germplasm. PLoS ONE 2015, 10, e0136803. [Google Scholar] [CrossRef]
- Mas-Gómez, J.; Cantín, C.M.; Moreno, M.; Prudencio, Á.S.; Gómez-Abajo, M.; Bianco, L.; Troggio, M.; Martínez-Gómez, P.; Rubio, M.; Martínez-García, P.J. Exploring genome-wide diversity in the national peach (Prunus persica) germplasm collection at CITA (Zaragoza, Spain). Agronomy 2021, 11, 481. [Google Scholar] [CrossRef]
- Montanari, S.; Saeed, M.; Knäbel, M.; Kim, Y.K.; Troggio, M.; Malnoy, M.; Velasco, R.; Fontana, P.; Won, K.H.; Durel, C.E.; et al. Identification of Pyrus Single Nucleotide Polymorphisms (SNPs) and Evaluation for Genetic Mapping in European Pear and Interspecific Pyrus Hybrids. PLoS ONE 2013, 8, e77022. [Google Scholar] [CrossRef]
- Myles, S.; Chia, J.M.; Hurwitz, B.; Simon, C.; Zhong, G.Y.; Buckler, E.; Ware, D. Rapid genomic characterization of the genus Vitis. PLoS ONE 2010, 5, e8219. [Google Scholar] [CrossRef] [Green Version]
- Le Paslier, M.-C.; Choisne, N.; Scalabrin, S.; Bacilieri, R.; Berard, A.; Bounon, R.; Boursiquot, J.-M.; Bras, M.; Brunel, D.; Chauveau, A.; et al. The GrapeReSeq 18K Vitis genotyping chip. IX International Symposium on Grapevine Physiology Biotechnology, La Serena, Chile, 2–26 April 2013; p. 18. [Google Scholar]
- De Jong, W.S.; De Jong, D.M.; Bodis, M. A fluorogenic 5′ nuclease (TaqMan) assay to assess dosage of a marker tightly linked to red skin color in autotetraploid potato. Theor. Appl. Genet. 2003, 107, 1384–1390. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Q.; Kang, Z.; Liu, S.; Li, H.; Mu, J.; Dai, M.; Han, D.; Zeng, Q.; Chen, X. Development and validation of KASP-SNP markers for QTL underlying resistance to stripe rust in common wheat cultivar P10057. Plant Dis. 2017, 101, 2079–2087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qureshi, N.; Kandiah, P.; Gessese, M.K.; Nsabiyera, V.; Wells, V.; Babu, P.; Wong, D.; Hayden, M.; Bariana, H.; Bansal, U. Development of co-dominant KASP markers co-segregating with Ug99 effective stem rust resistance gene Sr26 in wheat. Mol. Breed. 2018, 38, 1–9. [Google Scholar] [CrossRef]
- Collins, D.; Emebiri, L.; Tan, M.K.; El Bouhssini, M.; Wildman, O. Association of KASP markers with Hessian fly resistance in wheat of diverse origin. Euphytica 2018, 214, 1–8. [Google Scholar] [CrossRef]
- Wang, R.; Liu, Y.; Isham, K.; Zhao, W.; Wheeler, J.; Klassen, N.; Hu, Y.; Bonman, J.M.; Chen, J. QTL identification and KASP marker development for productive tiller and fertile spikelet numbers in two high-yielding hard white spring wheat cultivars. Mol. Breed. 2018, 38, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, L.; Anderson, J.A.; Chen, J.; Gill, B.S.; Tiwari, V.K.; Rawat, N. Development and validation of a perfect KASP marker for fusarium head blight resistance gene Fhb1 in wheat. Plant Pathol. J. 2019, 35, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Lei, L.; Li, G.; Powers, C.; Hunger, R.M.; Carver, B.F.; Yan, L. Development and deployment of KASP markers for multiple alleles of Lr34 in wheat. Theor. Appl. Genet. 2020, 133, 2183–2195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grewal, S.; Othmeni, M.; Walker, J.; Hubbart-Edwards, S.; Yang, C.Y.; Scholefield, D.; Ashling, S.; Isaac, P.; King, I.P.; King, J. Development of Wheat-Aegilops caudata Introgression Lines and Their Characterization Using Genome-Specific KASP Markers. Front. Plant Sci. 2020, 11, 606. [Google Scholar] [CrossRef] [PubMed]
- Makhoul, M.; Rambla, C.; Voss-Fels, K.P.; Hickey, L.T.; Snowdon, R.J.; Obermeier, C. Overcoming polyploidy pitfalls: A user guide for effective SNP conversion into KASP markers in wheat. Theor. Appl. Genet. 2020, 133, 2413–2430. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Fan, C.; Luo, J.; Huang, L.; Xie, D.; Li, Y.; Chen, Z.; Jiang, B.; Ning, S.; Yuan, Z.; et al. KASP markers to detect sub-chromosomal arm translocations between 6VS of Haynaldia villosa and 6AS of wheat. Euphytica 2021, 217, 10. [Google Scholar] [CrossRef]
- Xu, X.; Li, G.; Bai, G.; Bernardo, A.; Carver, B.F.; Amand, P.S.; Armstrong, J.S. Development of KASP markers for wheat greenbug resistance gene Gb5. Crop Sci. 2021, 61, 490–499. [Google Scholar] [CrossRef]
- Sangha, J.; Tucker, J.R.; Legge, W.G.; Badea, A. Use of KASP assays for the analysis of rpg4/Rpg5 gene complex for marker-assisted selection for Ug99 stem rust resistance in barley. Can. J. Plant Pathol. 2017, 39, 578. [Google Scholar]
- Steele, K.A.; Quinton-Tulloch, M.J.; Amgai, R.B.; Dhakal, R.; Khatiwada, S.P.; Vyas, D.; Heine, M.; Witcombe, J.R. Accelerating public sector rice breeding with high-density KASP markers derived from whole genome sequencing of indica rice. Mol. Breed. 2018, 38, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, G.; Chen, S.; Chen, L.; Sun, K.; Huang, C.; Zhou, D.; Huang, Y.; Wang, J.; Liu, Y.; Wang, H.; et al. Development of a core SNP arrays based on the KASP method for molecular breeding of rice. Rice 2019, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Basnet, B.R.; Ibrahim, A.M.H.; Rudd, J.C.; Chen, X.; Bowden, R.L.; Xue, Q.; Wang, S.; Johnson, C.D.; Metz, R.; et al. Developing KASP Markers on a Major Stripe Rust Resistance QTL in a Popular Wheat TAM 111 Using 90K Array and Genotyping-by-Sequencing SNPs. Crop Sci. 2019, 59, 165–175. [Google Scholar] [CrossRef]
- Addison, C.K.; Angira, B.; Kongchum, M.; Harrell, D.L.; Baisakh, N.; Linscombe, S.D.; Famoso, A.N. Characterization of Haplotype Diversity in the BADH2 Aroma Gene and Development of a KASP SNP Assay for Predicting Aroma in U.S. Rice. Rice 2020, 13, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Burow, G.; Chopra, R.; Hughes, H.; Xin, Z.; Burke, J. Marker Assisted Selection in Sorghum Using KASP Assay for the Detection of Single Nucleotide Polymorphism/Insertion Deletion. Methods Mol. Biol. 2019, 1931, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Grimm, K.D.S.; Porter, L.D. Development and Validation of KASP Markers for the Identification of Pea seedborne mosaic virus Pathotype P1 Resistance in Pisum sativum. Plant Dis. 2020, 104, 1824–1830. [Google Scholar] [CrossRef]
- Legendre, R.; McGregor, C. KASP (TM) Markers for Selection for Fruit Shape in Watermelon. Hortscience 2019, 54, S399. [Google Scholar]
- Paudel, L.; Clevenger, J.; McGregor, C. Refining of the egusi locus in watermelon using KASP assays. Sci. Hortic. 2019, 257, 108665. [Google Scholar] [CrossRef]
- Zanotto, S.; Vandenberg, A.; Khazaei, H. Development and validation of a robust KASP marker for zt2 locus in faba bean (Vicia faba). Plant Breed. 2020, 139, 375–380. [Google Scholar] [CrossRef]
- Devran, Z.; Göknur, A.; Mesci, L. Development of molecular markers for the Mi-1 gene in tomato using the KASP genotyping assay. Hortic. Environ. Biotechnol. 2016, 57, 156–160. [Google Scholar] [CrossRef]
- Devran, Z.; Kahveci, E. Development and validation of a user-friendly KASP marker for the Sw-5 locus in tomato. Australas. Plant Pathol. 2019, 48, 503–507. [Google Scholar] [CrossRef]
- Han, F.; Zhang, X.; Yuan, K.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Liu, Y.; Li, Z.; et al. A user-friendly KASP molecular marker developed for the DGMS-based breeding system in Brassica oleracea species. Mol. Breed. 2019, 39, 1–7. [Google Scholar] [CrossRef]
- da Cruz, M.F.A.; Bueno, R.D.; de Souza, F.B.; Moreira, M.A.; de Barros, E.G. Identification of SNPs for fatty acid content in soybean by the HRM technique. Pesqui. Agropecu. Bras. 2013, 48, 1596–1600. [Google Scholar] [CrossRef] [Green Version]
- Rai, V.P.; Singh, A.K.; Jaiswal, H.K.; Singh, S.P.; Singh, R.P.; Waza, S.A. Evaluation of molecular markers linked to fragrance and genetic diversity in Indian aromatic rice. Turk. J. Botany 2015, 39, 209–217. [Google Scholar] [CrossRef]
- Noh, Y.H.; Lee, S.; Whitaker, V.M.; Cearley, K.R.; Cha, J.S. A high-throughput marker-assisted selection system combining rapid DNA extraction and high-resolution melting and simple sequence repeat analysis: Strawberry as a model for fruit crops. J. Berry Res. 2017, 7, 23–31. [Google Scholar] [CrossRef]
- Geng, L.; Li, M.; Xie, S.; Wu, D.; Ye, L.; Zhang, G. Identification of genetic loci and candidate genes related to β-glucan content in barley grain by genome-wide association study in International Barley Core Selected Collection. Mol. Breed. 2021, 41, 1–12. [Google Scholar] [CrossRef]
- Paris, M.; Jones, M.G.K.; Eglinton, J.K. Genotyping Single Nucleotide Polymorphisms for Selection of Barley β-amylase Alleles. Plant Mol. Biol. Report. 2002, 20, 149–159. [Google Scholar] [CrossRef]
- Masouleh, A.K.; Waters, D.L.E.; Reinke, R.F.; Henry, R.J. A high-throughput assay for rapid and simultaneous analysis of perfect markers for important quality and agronomic traits in rice using multiplexed MALDI-TOF mass spectrometry. Plant Biotechnol. J. 2009, 7, 355–363. [Google Scholar] [CrossRef]
- Cheng, P.; Holdsworth, W.; Ma, Y.; Coyne, C.J.; Mazourek, M.; Grusak, M.A.; Fuchs, S.; McGee, R.J. Association mapping of agronomic and quality traits in USDA pea single-plant collection. Mol. Breed. 2015, 35, 1–13. [Google Scholar] [CrossRef]
- Lum, H.-K.; Lee, C.-H.; Butt, Y.K.-C.; Lo, S.C.-L. Sodium nitroprusside affects the level of photosynthetic enzymes and glucose metabolism in Phaseolus aureus (mung bean). Nitric Oxide 2005, 12, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Salazar, J.A.; Pacheco, I.; Shinya, P.; Zapata, P.; Silva, C.; Aradhya, M.; Velasco, D.; Ruiz, D.; Martínez-Gómez, P.; Infante, R. Genotyping by Sequencing for SNP-Based Linkage Analysis and Identification of QTLs Linked to Fruit Quality Traits in Japanese Plum (Prunus salicina Lindl.). Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larsen, B.; Migicovsky, Z.; Jeppesen, A.A.; Gardner, K.M.; Toldam-Andersen, T.B.; Myles, S.; Ørgaard, M.; Petersen, M.A.; Pedersen, C. Genome-Wide Association Studies in Apple Reveal Loci for Aroma Volatiles, Sugar Composition, and Harvest Date. Plant Genome 2019, 12, 180104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, L.; Ruggieri, V.; Pérez, S.; Alexiou, K.G.; Fernández, M.; Jahrmann, T.; Pujol, M.; Garcia-Mas, J. QTL mapping of melon fruit quality traits using a high-density GBS-based genetic map. BMC Plant Biol. 2018, 18, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Dhliwayo, T.; Mageto, E.K.; Palacios-Rojas, N.; Lee, M.; Yu, D.; Ruan, Y.; Zhang, A.; San Vicente, F.; Olsen, M.; et al. Genomic Prediction of Kernel Zinc Concentration in Multiple Maize Populations Using Genotyping-by-Sequencing and Repeat Amplification Sequencing Markers. Front. Plant Sci. 2020, 11, 534. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Coyne, C.J.; Grusak, M.A.; Mazourek, M.; Cheng, P.; Main, D.; McGee, R.J. Genome-wide SNP identification, linkage map construction and QTL mapping for seed mineral concentrations and contents in pea (Pisum sativum L.). BMC Plant Biol. 2017, 17, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Qin, J.; Shi, A.; Mou, B.; Grusak, M.A.; Weng, Y.; Ravelombola, W.; Bhattarai, G.; Dong, L.; Yang, W. Genetic diversity and association mapping of mineral element concentrations in spinach leaves. BMC Genomics 2017, 18, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Tian, H.; Yang, Y.; Wang, R.; Fan, Y.; Yi, H.; Jiang, B.; Wang, L.; Ren, J.; Xu, L.; Zhang, Y.; et al. Screening of 200 core SNPs and the Construction of a Systematic SNP-DNA Standard Fingerprint Database with More Than 20,000 Maize Varieties. Agriculture 2021, 11, 597. [Google Scholar] [CrossRef]
- Kuhn, D.N.; Groh, A.; Rahaman, J.; Freeman, B.; Arpaia, M.L.; Van den Berg, N.; Abeysekara, N.; Manosalva, P.; Chambers, A.H. Creation of an avocado unambiguous genotype SNP database for germplasm curation and as an aid to breeders. Tree Genet. Genomes 2019, 15, 71. [Google Scholar] [CrossRef] [Green Version]
- Medina-lozano, I.; Bertolín, J.R.; Zufiaurre, R.; Diaz, A. Improved UPLC-UV Method for the Quantification of Vitamin C in Lettuce Varieties (Lactuca sativa L.) and Crop Wild Relatives (Lactuca spp.). J. Vis. Exp. 2020, 160, e61440. [Google Scholar] [CrossRef] [PubMed]
- Dossou, S.S.K.; Xu, F.; You, J.; Zhou, R.; Li, D.; Wang, L. Widely targeted metabolome profiling of different colored sesame (Sesamum indicum L.) seeds provides new insight into their antioxidant activities. Food Res. Int. 2022, 151, 110850. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.Q.; Huang, X.Q.; Zhang, Y.Z.; Qian, J.; Yang, M.Z.; Wu, C.J.; Liu, J.F. Diversity in the content of some nutritional components in husked seeds of three wild rice species and rice varieties in Yunnan Province of China. J. Integr. Plant Biol. 2005, 47, 1260–1270. [Google Scholar] [CrossRef]
- Esteras, C.; Rambla, J.L.; Sánchez, G.; López-Gresa, M.P.; González-Mas, M.C.; Fernández-Trujillo, J.P.; Bellés, J.M.; Granell, A.; Picó, M.B. Fruit flesh volatile and carotenoid profile analysis within the Cucumis melo L. species reveals unexploited variability for future genetic breeding. J. Sci. Food Agric. 2018, 98, 3915–3925. [Google Scholar] [CrossRef] [PubMed]
- Burbano-Erazo, E.; León-Pacheco, R.I.; Cordero-Cordero, C.C.; López-Hernández, F.; Cortés, A.J.; Tofiño-Rivera, A.P. Multi-Environment Yield Components in Advanced Common Bean (Phaseolus vulgaris L.) × Tepary Bean (P. acutifolius A. Gray) Interspecific Lines for Heat and Drought tolerance. Agronomy 2021, 11, 1978. [Google Scholar] [CrossRef]
- Herraiz, F.J.; Raigón, M.D.; Vilanova, S.; García-Martínez, M.D.; Gramazio, P.; Plazas, M.; Rodríguez-Burruezo, A.; Prohens, J. Fruit composition diversity in land races and modern pepino (Solanum muricatum) varieties and wild related species. Food Chem. 2016, 203, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Lozano, I.; Bertolín, J.R.; Díaz, A. Nutritional value of commercial and traditional lettuce (Lactuca sativa L.) and wild relatives: Vitamin C and anthocyanin content. Food Chem. 2021, 359. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Wang, D.; Gong, L.; Lyu, Y.; Guo, H.; Chen, W.; Jin, C.; Liu, X.; Fang, C.; Luo, J. Comparative analysis of metabolome of rice seeds at three developmental stages using a recombinant inbred line population. Plant J. 2019, 100, 908–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, J.E.; Kim, J.-G.; Fischer, C.R.; Mehta, N.; Dufour-Schroif, C.; Wemmer, K.; Mudgett, M.B.; Sattely, E. Pathogen-responsive gene cluster for highly modified fatty acids in tomato. Cell 2020, 180, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Raza, A. Metabolomics: A systems biology approach for enhancing heat stress tolerance in plants. Plant Cell Rep. 2020. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, D.; Bohra, A.; Thudi, M.; Varshney, R.K. Fine mapping and gene cloning in the post-NGS era: Advances and prospects. Theor. Appl. Genet. 2020, 133, 1791–1810. [Google Scholar] [CrossRef] [Green Version]
- Sharma, V.; Gupta, P.; Priscilla, K.; Sharankumar; Hangargi, B.; Veershetty, A.; Ramrao, D.P.; Suresh, S.; Narasanna, R.; Naik, G.R.; et al. Metabolomics Intervention Towards Better Understanding of Plant Traits. Cells 2021, 10, 346. [Google Scholar] [CrossRef] [PubMed]
- Bilbrey, E.A.; Williamson, K.; Hatzakis, E.; Miller, D.D.; Fresnedo-Ramírez, J.; Cooperstone, J.L. Integrating genomics and multiplatform metabolomics enables metabolite quantitative trait loci detection in breeding-relevant apple germplasm. New Phytol. 2021, 232, 1944–1958. [Google Scholar] [CrossRef] [PubMed]
- Templer, S.E.; Ammon, A.; Pscheidt, D.; Ciobotea, O.; Schuy, C.; McCollum, C.; Sonnewald, U.; Hanemann, A.; Förster, J.; Ordon, F.; et al. Metabolite profiling of barley flag leaves under drought and combined heat and drought stress reveals metabolic QTLs for metabolites associated with antioxidant defense. J. Exp. Bot. 2017, 68, 1697–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matros, A.; Houston, K.; Tucker, M.R.; Schreiber, M.; Berger, B.; Aubert, M.K.; Wilkinson, L.G.; Witzel, K.; Waugh, R.; Seiffert, U.; et al. Genome-wide association study reveals the genetic complexity of fructan accumulation patterns in barley grain. J. Exp. Bot. 2021, 72, 2383–2402. [Google Scholar] [CrossRef] [PubMed]
- Mahalingam, R.; Sallam, A.H.; Steffenson, B.J.; Fiedler, J.D.; Walling, J.G. Genome-wide association analysis of natural variation in seed tocochromanols of barley. Plant Genome 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Peng, M.; Shahzad, R.; Gul, A.; Subthain, H.; Shen, S.; Lei, L.; Zheng, Z.; Zhou, J.; Lu, D.; Wang, S.; et al. Differentially evolved glucosyltransferases determine natural variation of rice flavone accumulation and UV-tolerance. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ferrão, L.F.V.; Johnson, T.S.; Benevenuto, J.; Edger, P.P.; Colquhoun, T.A.; Munoz, P.R. Genome-wide association of volatiles reveals candidate loci for blueberry flavor. New Phytol. 2020, 226, 1725–1737. [Google Scholar] [CrossRef] [PubMed]
- Matros, A.; Liu, G.; Hartmann, A.; Jiang, Y.; Zhao, Y.; Wang, H.; Ebmeyer, E.; Korzun, V.; Schachschneider, R.; Kazman, E.; et al. Genome-metabolite associations revealed low heritability, high genetic complexity, and causal relations for leaf metabolites in winter wheat (Triticum aestivum). J. Exp. Bot. 2017, 68, 415–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Li, S.; Wang, Y.; Wang, B.; Fan, G.; Zeng, Q.; Zhao, F.; Xu, C.; Zhang, X.; Tang, T.; et al. Metabolome-Based Genome-Wide Association Study Provides Genetic Insights Into the Natural Variation of Foxtail Millet. Front. Plant Sci. 2021, 12, 665530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Alseekh, S.; Zhu, X.; Zhang, Q.; Fernie, A.R.; Kuang, H.; Wen, W. Dissection of the domestication-shaped genetic architecture of lettuce primary metabolism. Plant J. 2020, 104, 613–630. [Google Scholar] [CrossRef]
- Wang, Y. A draft genome, resequencing, and metabolomes reveal the genetic background and molecular basis of the nutritional and medicinal properties of loquat (Eriobotrya japonica (Thunb.) Lindl). Hortic. Res. 2021, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Peng, Z.; Yang, X.; Wang, W.; Fu, J.; Wang, J.; Han, Y.; Chai, Y.; Guo, T.; Yang, N.; et al. Genome-wide association study dissects the genetic architecture of oil biosynthesis in maize kernels. Nat. Genet. 2013, 45, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Li, D.; Li, X.; Gao, Y.; Li, W.; Li, H.; Liu, J.; Liu, H.; Chen, W.; Luo, J.; et al. Metabolome-based genome-wide association study of maize kernel leads to novel biochemical insights. Nat. Commun. 2014, 5, 3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riedelsheimer, C.; Lisec, J.; Czedik-Eysenberg, A.; Sulpice, R.; Flis, A.; Grieder, C.; Altmann, T.; Stitt, M.; Willmitzer, L.; Melchinger, A.E. Genome-wide association mapping of leaf metabolic profiles for dissecting complex traits in maize. Proc. Natl. Acad. Sci. USA 2012, 109, 8872–8877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipka, A.E.; Gore, M.A.; Magallanes-Lundback, M.; Mesberg, A.; Lin, H.; Tiede, T.; Chen, H.; Robin Buell, C.; Buckler, E.S.; Rocheford, T.; et al. Genome-Wide Association Study and Pathway-Level Analysis of Tocohromanol Levels in Maize Grain. G3 Genes Genomes Genet. 2013, 3, 1287–1299. [Google Scholar] [CrossRef] [Green Version]
- Owens, B.F.; Gore, M.A.; Magallanes-Lundback, M.; Tiede, T.; Diepenbrock, C.H.; Kandianis, C.B.; Kim, E.; Cepela, J.; Mateos-Hernandez, M.; Robin Buell, C.; et al. A Foundation for Provitamin A Biofortification of Maize: Genome-Wide Association and Genomic Prediction Models of Carotenoid Levels. Genetics 2014, 198, 1699–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Liu, S.; Wang, T.; Li, F.; Cheng, J.; Lai, J.; Qin, F.; Li, Z.; Wang, X.; Jiang, C. Metabolomics-driven gene mining and genetic improvement of tolerance to salt-induced osmotic stress in maize. New Phytol. 2021, 230, 2355–2370. [Google Scholar] [CrossRef]
- Suwarno, W.B.; Pixley, K.V.; Palacios-Rojas, N.; Kaeppler, S.M.; Babu, R. Genome-wide association analysis reveals new targets for carotenoid biofortification in maize. Theor. Appl. Genet. 2015, 128, 851–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, M.; Li, D.; Luo, J.; Xiao, Y.; Liu, H.; Pan, Q.; Zhang, X.; Jin, M.; Zhao, M.; Yan, J. The genetic architecture of amino acids dissection by association and linkage analysis in maize. Plant Biotechnol. J. 2017, 15, 1250–1263. [Google Scholar] [CrossRef] [Green Version]
- Jin, M.; Zhang, X.; Zhao, M.; Deng, M.; Du, Y.; Zhou, Y.; Wang, S.; Tohge, T.; Fernie, A.R.; Willmitzer, L.; et al. Integrated genomics-based mapping reveals the genetics underlying maize flavonoid biosynthesis. BMC Plant Biol. 2017, 17, 17. [Google Scholar] [CrossRef] [Green Version]
- Azmach, G.; Menkir, A.; Spillane, C.; Gedil, M. Genetic Loci Controlling Carotenoid Biosynthesis in Diverse Tropical Maize Lines. G3 Genes Genomes Genet. 2018, 8, 1049–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.; Xu, S.; Fan, Y.; Liu, N.; Zhan, W.; Liu, H.; Xiao, Y.; Li, K.; Pan, Q.; Li, W.; et al. Beyond pathways: Genetic dissection of tocopherol content in maize kernels by combining linkage and association analyses. Plant Biotechnol. J. 2018, 16, 1464–1475. [Google Scholar] [CrossRef] [PubMed]
- Wen, W.; Jin, M.; Li, K.; Liu, H.; Xiao, Y.; Zhao, M.; Alseekh, S.; Li, W.; de Abreu e Lima, F.; Brotman, Y.; et al. An integrated multi-layered analysis of the metabolic networks of different tissues uncovers key genetic components of primary metabolism in maize. Plant J. 2018, 93, 1116–1128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves, M.L.; Bento-Silva, A.; Carbas, B.; Gaspar, D.; Paulo, M.; Brites, C.; Mendes-Moreira, P.; Brites, C.M.; Bronze, M.D.R.; Malosetti, M.; et al. Alleles to Enhance Antioxidant Content in Maize-A Genome-Wide Association Approach. J. Agric. Food Chem. 2020, 68, 4051–4061. [Google Scholar] [CrossRef]
- Baseggio, M.; Murray, M.; Magallanes-Lundback, M.; Kaczmar, N.; Chamness, J.; Buckler, E.S.; Smith, M.E.; DellaPenna, D.; Tracy, W.F.; Gore, M.A. Genome-Wide Association and Genomic Prediction Models of Tocochromanols in Fresh Sweet Corn Kernels. Plant Genome 2019, 12, 180038. [Google Scholar] [CrossRef] [Green Version]
- Diepenbrock, C.H.; Ilut, D.C.; Magallanes-Lundback, M.; Kandianis, C.B.; Lipka, A.E.; Bradbury, P.J.; Holland, J.B.; Hamilton, J.P.; Wooldridge, E.; Vaillancourt, B.; et al. Eleven biosynthetic genes explain the majority of natural variation in carotenoid levels in maize grain. Plant Cell 2021, 33, 882–900. [Google Scholar] [CrossRef]
- Chatham, L.A.; Juvik, J.A. Linking anthocyanin diversity, hue, and genetics in purple corn. G3 Genes Genomes Genet. 2021, 11, jkaa062. [Google Scholar] [CrossRef]
- Levina, A.V.; Hoekenga, O.; Gordin, M.; Broeckling, C.; De Jong, W.S. Genetic analysis of potato tuber metabolite composition: Genome-wide association studies applied to a nontargeted metabolome. Crop. Sci. 2021, 61, 591–603. [Google Scholar] [CrossRef]
- Dong, X.; Gao, Y.; Chen, W.; Wang, W.; Gong, L.; Liu, X.; Luo, J. Spatiotemporal distribution of phenolamides and the genetics of natural variation of hydroxycinnamoyl spermidine in rice. Mol. Plant 2015, 8, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Brotman, Y.; Llorente-Wiegand, C.; Oyong, G.; Badoni, S.; Misra, G.; Anacleto, R.; Parween, S.; Pasion, E.; Tiozon, R.N.; Anonuevo, J.J.; et al. The genetics underlying metabolic signatures in a brown rice diversity panel and their vital role in human nutrition. Plant J. 2021, 106, 507–525. [Google Scholar] [CrossRef]
- Chen, W.; Wang, W.; Peng, M.; Gong, L.; Gao, Y.; Wan, J.; Wang, S.; Shi, L.; Zhou, B.; Li, Z.; et al. Comparative and parallel genome-wide association studies for metabolic and agronomic traits in cereals. Nat. Commun. 2016, 7, 12767. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, F.; Nakabayashi, R.; Yang, Z.; Okazaki, Y.; Yonemaru, J.I.; Ebana, K.; Yano, M.; Saito, K. Metabolome-genome-wide association study dissects genetic architecture for generating natural variation in rice secondary metabolism. Plant J. 2015, 81, 13–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Guo, H.; Huang, J.; Yang, C.; Li, Y.; Wang, X.; Qu, L.; Liu, X.; Luo, J. A UV-B-responsive glycosyltransferase, OsUGT706C2, modulates flavonoid metabolism in rice. Sci. China Life Sci. 2020, 63, 1037–1052. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Tian, R.; Shao, Z.; Zhang, H.; Chu, J.; Li, W.; Kong, Y.; Du, H.; Zhang, C. Genetic loci and causal genes for seed fatty acids accumulation across multiple environments and genetic backgrounds in soybean. Mol. Breed. 2021, 41, 31. [Google Scholar] [CrossRef]
- Wu, D.; Li, D.; Zhao, X.; Zhan, Y.; Teng, W.; Qiu, L.; Zheng, H.; Li, W.; Han, Y. Identification of a candidate gene associated with isoflavone content in soybean seeds using genome-wide association and linkage mapping. Plant J. 2020, 104, 950–963. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Y.; Li, P.; Zhang, Y.W.; Zuo, J.F.; Li, G.; Han, X.; Dunwell, J.M.; Zhang, Y.M. Three-dimensional genetic networks among seed oil-related traits, metabolites and genes reveal the genetic foundations of oil synthesis in soybean. Plant J. 2020, 103, 1103–1124. [Google Scholar] [CrossRef] [PubMed]
- Fang, K.; Xia, Z.; Li, H.; Jiang, X.; Qin, D.; Wang, Q.; Wang, Q.; Pan, C.; Li, B.; Wu, H. Genome-wide association analysis identified molecular markers associated with important tea flavor-related metabolites. Hortic. Res. 2021, 8, 42. [Google Scholar] [CrossRef]
- Yamashita, H.; Uchida, T.; Tanaka, Y.; Katai, H.; Nagano, A.J.; Morita, A.; Ikka, T. Genomic predictions and genome-wide association studies based on RAD-seq of quality-related metabolites for the genomics-assisted breeding of tea plants. Sci. Rep. 2020, 10, 17480. [Google Scholar] [CrossRef]
- Tieman, D.; Zhu, G.; Resende, M.F.R.; Lin, T.; Nguyen, C.; Bies, D.; Rambla, J.L.; Beltran, K.S.O.; Taylor, M.; Zhang, B.; et al. A chemical genetic roadmap to improved tomato flavor. Plant Sci. 2017, 355, 6323. [Google Scholar] [CrossRef]
- Sauvage, C.; Segura, V.; Bauchet, G.; Stevens, R.; Do, P.T.; Nikoloski, Z.; Fernie, A.R.; Causse, M. Genome-Wide Association in Tomato Reveals 44 Candidate Loci for Fruit Metabolic Traits. Plant Physiol. 2014, 165, 1120–1132. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Wang, S.; Huang, Z.; Zhang, S.; Liao, Q.; Zhang, C.; Lin, T.; Qin, M.; Peng, M.; Yang, C.; et al. Rewiring of the Fruit Metabolome in Tomato Breeding. Cell 2018, 172, 249–261.e12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgos, E.; Belen De Luca, M.; Diouf, I.; de Haro, L.A.; Albert, E.; Sauvage, C.; Tao, Z.J.; Bermudez, L.; Asís, R.; Nesi, A.N.; et al. Validated MAGIC and GWAS population mapping reveals the link between vitamin E content and natural variation in chorismate metabolism in tomato. Plant J. 2021, 105, 907–923. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Hu, X.; Shi, T.; Yin, H.; Sun, D.; Hao, Y.; Xia, X.; Luo, J.; Fernie, A.R.; He, Z.; et al. Metabolite-based genome-wide association study enables dissection of the flavonoid decoration pathway of wheat kernels. Plant Biotechnol. J. 2020, 18, 1722–1735. [Google Scholar] [CrossRef] [PubMed]
- Meuwissen, T.H.E.; Hayes, B.J.; Goddard, M.E. Prediction of total genetic value using genome-wide dense marker maps. Genetics 2001, 157, 1819–1829. [Google Scholar] [CrossRef] [PubMed]
- Duangjit, J.; Causse, M.; Sauvage, C. Efficiency of genomic selection for tomato fruit quality. Mol. Breed. 2016, 36, 29. [Google Scholar] [CrossRef]
- Battenfield, S.D.; Guzmán, C.; Gaynor, R.C.; Singh, R.P.; Peña, R.J.; Dreisigacker, S.; Fritz, A.K.; Poland, J.A. Genomic Selection for Processing and End-Use Quality Traits in the CIMMYT Spring Bread Wheat Breeding Program. Plant Genome 2016, 9, plantgenome2016-01. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dempewolf, H.; Baute, G.; Anderson, J.; Kilian, B.; Smith, C.; Guarino, L. Past and Future Use of Wild Relatives in Crop Breeding. Crop. Sci. 2017, 57, 1070–1082. [Google Scholar] [CrossRef]
- Bouis, H.E.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Sec. 2017, 12, 49–58. [Google Scholar] [CrossRef] [PubMed]
- HarvestPlus. Getting Biofortified Food on Everyone’s Plate—2019 Annual Report; HarvestPlus: Washington, DC, USA, 2019. [Google Scholar]
- Sazawal, S.; Dhingra, U.; Dhingra, P.; Dutta, A.; Deb, S.; Kumar, J.; Devi, P.; Prakash, A. Efficacy of high zinc biofortified wheat in improvement of micronutrient status, and prevention of morbidity among preschool children and women—A double masked, randomized, controlled trial. Nutr. J. 2018, 17, 86. [Google Scholar] [CrossRef] [Green Version]
- Palmer, A.C.; Healy, K.; Barffour, M.A.; Siamusantu, W.; Chileshe, J.; Schulze, K.J.; West, K.P.J.; Labrique, A.B. Provitamin A Carotenoid-Biofortified Maize Consumption Increases Pupillary Responsiveness among Zambian Children in a Randomized Controlled Trial. J. Nutr. 2016, 146, 2551–2558. [Google Scholar] [CrossRef] [Green Version]
- Low, J.W.; Mwanga, R.O.M.; Andrade, M.; Carey, E.; Ball, A.-M. Tackling vitamin A deficiency with biofortified sweetpotato in sub-Saharan Africa. Glob. Food Sec. 2017, 14, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, B.M.; Palacios-Rojas, N.; Hossain, F.; Muthusamy, V.; Menkir, A.; Dhliwayo, T.; Ndhlela, T.; San Vicente, F.; Nair, S.K.; Vivek, B.S.; et al. Molecular Breeding for Nutritionally Enriched Maize: Status and Prospects. Front. Genet. 2020, 10, 1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruet-Burgos, C.; Cox, S.; Ioerger, B.P.; Perumal, R.; Hu, Z.; Herald, T.J.; Bean, S.R.; Rhodes, D.H. Advancing provitamin A biofortification in sorghum: Genome-wide association studies of grain carotenoids in global germplasm. Plant Genome 2020, 13, e20013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izquierdo, P.; Astudillo, C.; Blair, M.W.; Iqbal, A.M.; Raatz, B.; Cichy, K.A. Meta-QTL analysis of seed iron and zinc concentration and content in common bean (Phaseolus vulgaris L.). Theor. Appl. Genet. 2018, 131, 1645–1658. [Google Scholar] [CrossRef] [PubMed]
- Krishnappa, G.; Rathan, N.D.; Sehgal, D.; Ahlawat, A.K.; Singh, S.K.; Singh, S.K.; Shukla, R.B.; Jaiswal, J.P.; Solanki, I.S.; Singh, G.P.; et al. Identification of Novel Genomic Regions for Biofortification Traits Using an SNP Marker-Enriched Linkage Map in Wheat (Triticum aestivum L.). Front. Nutr. 2021, 8, 669444. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Sharma, A.; Vats, S.; Tiwari, V.; Kumari, A.; Mishra, V.; Krishania, M. Vitamins in Cereals: A Critical Review of Content, Health Effects, Processing Losses, Bioaccessibility, Fortification, and Biofortification Strategies for Their Improvement. Front. Nutr. 2021, 8, 586815. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, V.; Muthysamy, V.; Jha, S.; Zunjare, R.U.; Baveja, A.; Sosad, S. Development of low phytic acid maize through marker assisted introgression of lpa1-1 and lpa2-1 genes. In Proceedings of the 13th Asian Maize Conference on and Expert Consultation on Maize for Food, Feed, Nutrition and Environmental Security, Ludhiana, India, 8–10 October 2018; pp. 143–144. [Google Scholar]
- Storozhenko, S.; De Brouwer, V.; Volckaert, M.; Navarrete, O.; Blancquaert, D.; Zhang, G.F.; Lambert, W.; Van Der Straeten, D. Folate fortification of rice by metabolic engineering. Nat. Biotechnol. 2007, 25, 1277–1279. [Google Scholar] [CrossRef] [PubMed]
- Díaz de La Garza, R.I.; Gregory, J.F.; Hanson, A.D. Folate biofortification of tomato fruit. Proc. Natl. Acad. Sci. USA 2007, 104, 4218–4222. [Google Scholar] [CrossRef] [Green Version]
- Bulley, S.; Wright, M.; Rommens, C.; Yan, H.; Rassam, M.; Lin-Wang, K.; Andre, C.; Brewster, D.; Karunairetnam, S.; Allan, A.C.; et al. Enhancing ascorbate in fruits and tubers through over-expression of the L-galactose pathway gene GDP-L-galactose phosphorylase. Plant Biotechnol. J. 2012, 10, 390–397. [Google Scholar] [CrossRef]
- Pierce, E.C.; LaFayette, P.R.; Ortega, M.A.; Joyce, B.L.; Kopsell, D.A.; Parrott, W.A. Ketocarotenoid production in soybean seeds through metabolic engineering. PLoS ONE 2015, 10, e0138196. [Google Scholar] [CrossRef] [Green Version]
- Holme, I.B.; Dionisio, G.; Brinch-Pedersen, H.; Wendt, T.; Madsen, C.K.; Vincze, E.; Holm, P.B. Cisgenic barley with improved phytase activity. Plant Biotechnol. J. 2012, 10, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Kamrani, M.; Kohnehrouz, B.B.; Gholizadeh, A. Cisgenic inhibition of the potato cold induced phosphorylase L gene expression and decrease in sugar contents. Afr. J. Biotechnol. 2011, 10, 10076–10082. [Google Scholar] [CrossRef]
- De Vetten, N.; Wolters, A.M.; Raemakers, K.; Van der Meer, I.; Ter Stege, R.; Heeres, E.; Heeres, P.; Visser, R. A transformation method for obtaining marker-free plants of a cross-pollinating and vegetatively propagated crop. Nat. Biotechnol. 2003, 21, 439–442. [Google Scholar] [CrossRef] [PubMed]
- Chawla, R.; Shakya, R.; Rommens, C.M. Tuber-specific silencing of asparagine synthetase-1 reduces the acrylamide-forming potential of potatoes grown in the field without affecting tuber shape and yield. Plant Biotechnol. J. 2012, 10, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Gil-Humanes, J.; Pistón, F.; Hernando, A.; Alvarez, J.B.; Shewry, P.R.; Barro, F. Silencing of γ-gliadins by RNA interference (RNAi) in bread wheat. J. Cereal Sci. 2008, 48, 565–568. [Google Scholar] [CrossRef]
- Brinch-Pedersen, H.; Borg, S.; Tauris, B.; Holm, P.B. Molecular genetic approaches to increasing mineral availability and vitamin content of cereals. J. Cereal Sci. 2007, 46, 308–326. [Google Scholar] [CrossRef]
- Coca, M.; Peñas, G.; Gómez, J.; Campo, S.; Bortolotti, C.; Messeguer, J.; Segundo, B.S. Enhanced resistance to the rice blast fungus Magnaporthe grisea conferred by expression of a cecropin A gene in transgenic rice. Planta 2006, 223, 392–406. [Google Scholar] [CrossRef] [Green Version]
- Girgi, M.; Breese, W.A.; Lörz, H.; Oldach, K.H. Rust and Downy Mildew Resistance in Pearl Millet (Pennisetum glaucum) Mediated by Heterologous Expression of the afp Gene from Aspergillus giganteus. Transgenic Res. 2006, 15, 313–324. [Google Scholar] [CrossRef]
- Yadav, H.; Malik, K.; Kumar, S.; Jaiwal, P.K. Comparative regeneration in six bread wheat (Triticum aestivum L.) varieties from immature and mature scutella for developing efficient and genotype-independent protocol prerequisite for genetic improvement of wheat. Vitr. Cell. Dev. Biol.-Plant 2020, 56, 610–617. [Google Scholar] [CrossRef]
- Sainger, M.; Chaudhary, D.; Dahiya, S.; Jaiwal, R.; Jaiwal, P.K. Development of an efficient in vitro plant regeneration system amenable to Agrobacterium-mediated transformation of a recalcitrant grain legume blackgram (Vigna mungo L. Hepper). Physiol. Mol. Biol. Plants 2015, 21, 505–517. [Google Scholar] [CrossRef] [Green Version]
- Garg, M.; Sharma, N.; Sharma, S.; Kapoor, P.; Kumar, A.; Chunduri, V.; Arora, P. Biofortified crops generated by breeding, agronomy, and transgenic approaches are improving lives of millions of people around the world. Front. Nutr. 2018, 5, 12. [Google Scholar] [CrossRef] [PubMed]
- Tang, G.; Qin, J.; Dolnikowski, G.G.; Russell, R.M.; Grusak, M.A. Golden Rice is an effective source of vitamin A. Am. J. Clin. Nutr. 2009, 89, 1776–1783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, S.J. Genetically modified crops, regulatory delays, and international trade. Food Energy Secur. 2017, 6, 78–86. [Google Scholar] [CrossRef] [Green Version]
- Schouten, H.J.; Krens, F.A.; Jacobsen, E. Do cisgenic plants warrant less stringent oversight? Nat. Biotechnol. 2006, 24, 753. [Google Scholar] [CrossRef] [PubMed]
- Rommens, C.M. All-native DNA transformation: A new approach to plant genetic engineering. Trends Plant Sci. 2004, 9, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Liu, Y.; Liu, Y.; Tan, Y.; Huang, J.; Shu, Q. Mutation of Inositol 1,3,4-trisphosphate 5/6-kinase6 Impairs Plant Growth and Phytic Acid Synthesis in Rice. Plants 2019, 8, 114. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Jiao, G.; Liu, Z.; Zhang, X.; Li, J.; Guo, X.; Du, W.; Du, J.; Francis, F.; Zhao, Y.; et al. Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes. Front. Plant Sci. 2017, 8, 298. [Google Scholar] [CrossRef]
- Schouten, H.J.; Krens, F.A.; Jacobsen, E. Cisgenic plants are similar to traditionally bred plants: International regulations for genetically modified organisms should be altered to exempt cisgenesis. EMBO Rep. 2006, 7, 750–753. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Duwal, A.; Su, Q.; Vossen, J.H.; Visser, R.G.F.; Jacobsen, E. Vector integration in triple R gene transformants and the clustered inheritance of resistance against potato late blight. Transgenic Res. 2013, 22, 315–325. [Google Scholar] [CrossRef]
- Conner, A.J.; Barrell, P.J.; Baldwin, S.J.; Lokerse, A.S.; Cooper, P.A.; Erasmuson, A.K.; Nap, J.P.; Jacobs, J.M.E. Intragenic vectors for gene transfer without foreign DNA. Euphytica 2007, 154, 341–353. [Google Scholar] [CrossRef]
- Schouten, H.J.; Jacobsen, E. Cisgenesis and intragenesis, sisters in innovative plant breeding. Trends Plant Sci. 2008, 13, 260–261. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, M.; Prasad, K.; Bhatnagar-Mathur, P.; Narasu, M.L.; Waliyar, F.; Sharma, K.K. An efficient method for the production of marker-free transgenic plants of peanut (Arachis hypogaea L.). Plant Cell Rep. 2010, 29, 495–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doshi, K.M.; Eudes, F.; Laroche, A.; Gaudet, D. Anthocyanin expression in marker free transgenic wheat and triticale embryos. Vitr. Cell. Dev. Biol.-Plant 2007, 43, 429–435. [Google Scholar] [CrossRef]
- Vanblaere, T.; Szankowski, I.; Schaart, J.; Schouten, H.; Flachowsky, H.; Broggini, G.A.L.; Gessler, C. The development of a cisgenic apple plant. J. Biotechnol. 2011, 154, 304–311. [Google Scholar] [CrossRef]
- Ling, F.; Zhou, F.; Chen, H.; Lin, Y. Development of marker-free insect-resistant indica rice by Agrobacterium tumefaciens-mediated co-transformation. Front. Plant Sci. 2016, 7, 1608. [Google Scholar] [CrossRef] [Green Version]
- Delwaide, A.C.; Nalley, L.L.; Dixon, B.L.; Danforth, D.M.; Nayga, R.M.; Van Loo, E.J.; Verbeke, W. Revisiting GMOs: Are There Differences in European Consumers’ Acceptance and Valuation for Cisgenically vs Transgenically Bred Rice? PLoS ONE 2015, 10, e0126060. [Google Scholar] [CrossRef] [Green Version]
- Shew, A.M.; Nalley, L.L.; Danforth, D.M.; Dixon, B.L.; Nayga, R.M.; Delwaide, A.C.; Valent, B. Are all GMOs the same? Consumer acceptance of cisgenic rice in India. Plant Biotechnol. J. 2016, 14, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Edenbrandt, A.K.; Gamborg, C.; Thorsen, B.J. Consumers’ Preferences for Bread: Transgenic, Cisgenic, Organic or Pesticide-free? J. Agric. Econ. 2018, 69, 121–141. [Google Scholar] [CrossRef]
- Raj, R.S.; Singh, C.; Modi, A.; Subhash, N. Genetic transformation of lowland rice variety GR11 for drought tolerance and its ratification for upland paddy cultivation. Indian J. Genet. Plant Breed. 2015, 75, 30–40. [Google Scholar] [CrossRef]
- Roberts, R.J. The Nobel Laureates’ Campaign Supporting GMOs. J. Innov. Knowl. 2018, 3, 61–65. [Google Scholar] [CrossRef]
- June, M.K. International Service for the Acquisition of Agri-Biotech (ISAAA). Global Status of Commercialized Biotech/GM Crops: 2019; ISAAA: Ithaca, NY, USA, 2020; ISBN 978-1-892456-69-9. [Google Scholar]
- Purchase, I.F.H. What determines the acceptability of genetically modified food that can improve human nutrition? Toxicol. Appl. Pharmacol. 2005, 207, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.; Ammann, K. New GMO regulations for old: Determining a new future for EU crop biotechnology. GM Crop. Food 2017, 8, 13–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougall, P. R&D Trends for Chemical Crop Protection Products and the Position of the European Market; Phillips McDougall Ltd.: Pathhead, UK, 2013. [Google Scholar]
- Holme, I.B.; Wendt, T.; Holm, P.B. Intragenesis and cisgenesis as alternatives to transgenic crop development. Plant Biotechnol. J. 2013, 11, 395–407. [Google Scholar] [CrossRef] [PubMed]
- Hou, H.; Atlihan, N.; Lu, Z.X. New biotechnology enhances the application of cisgenesis in plant breeding. Front. Plant Sci. 2014, 5, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russell, A.W.; Sparrow, R. The case for regulating intragenic GMOs. J. Agric. Environ. Ethics 2008, 21, 153–181. [Google Scholar] [CrossRef]
- EFSA Panel on Genetically Modified Organisms (GMO). Scientific opinion addressing the safety assessment of plants developed through cisgenesis and intragenesis. EFSA J. 2012, 10, 2561. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA). International Scientific Workshop ‘Risk Assessment Considerations for RNAi-Based GM Plants’; EFSA Supporting Publication: Brussels, Belgium, 2014; Volume EN-705. [Google Scholar]
- Environmental Protection Agency (EPA). Pesticides; Data Requirements for Plant.-Incorporated Protectants (PIPs) and Certain Exemptions for PIPs; EPA: Whasington, DC, USA, 2011; Volume 76.
- De Steur, H.; Wesana, J.; Blancquaert, D.; Van Der Straeten, D.; Gellynck, X. The socioeconomics of genetically modified biofortified crops: A systematic review and meta-analysis. Ann. N. Y. Acad. Sci. 2017, 1390, 14–33. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Li, P.; Zou, C.; Lu, Y.; Xie, C.; Zhang, X.; Prasanna, B.M.; Olsen, M.S. Enhancing genetic gain in the era of molecular breeding. J. Exp. Bot. 2017, 68, 2641–2666. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Database Name | Url | Crop ‡ |
|---|---|---|
| CerealsDB | https://www.cerealsdb.uk.net/cerealgenomics/CerealsDB/indexNEW.php, accessed on 17 January 2022 | Bread wheat (Triticum eastivum L.) |
| Chickpea SNP-InDel Database (CicArVarDB) | https://cegresources.icrisat.org/cicarvardb, accessed on the 17 of January 2022 | Chickpea (Cicer arietinum L.) |
| CropSNPdb | http://snpdb.appliedbioinformatics.com.au/, accessed on 17 January 2022 | Bread wheat (T. eastivum L.) Cabbage (Brassica rapa L.) Cauliflower (Brassica oleracea L.) Indian mustard (Brassica juncea L.) Oilseed rape (Brassica napus L.) |
| Cucurbit Genomics Database (CuGeDG) | http://cucurbitgenomics.org/, accessed on 17 January 2022 | Cucumber (Cucumis sativus L.) Melon (Cucumis melo L.) Pumpkin (Cucurbita spp.) Watermelon (Citrullus lanatus Thumb.) |
| Genome Database for Rosaceae (GDR) | https://www.rosaceae.org, accessed on 17 January 2022 | Apple tree (Malus spp.) Blackberry (Rubus spp.) Peach tree (Prunus spp.) Pear tree (Pyrus spp.) Strawberry (Fragaria spp.) |
| Gramene | https://www.gramene.org, accessed on 17 January 2022 | African rice (Oryza galberrina Steud) Asian rice (Oryza sativa L.) Barley (Hordeum vulgare L.) Foxtail millet (Setaria italica (L.) Beauv.) Maize (Zea mays L.) Sorghum (Sorghum bicolor (L.) Moench) Wheat (Triticum spp.) |
| Kazusa Tomato Genomics Database (KaTomicsDB) | https://www.kazusa.or.jp/tomato/, accessed on 17 January 2022 | Tomato (Solanum lycopersicum L.) |
| Lettuce Genome Database (LettuceGDB) | https://www.lettucegdb.com, accessed on 17 January 2022 | Lettuce (Lactuca sativa L.) |
| Maize Genetics and Genomics Database (MaizeGDB) | https://www.maizegdb.org/, accessed on 17 January 2022 | Maize (Z. mays L.) |
| Maize SNP-DNA Fingerprint Database | http://doi.org/10.3390/agriculture11070597 (Tables S1 and S2; [127]), accessed on 18 January 2022 | Maize (Z. mays L.) |
| Q-TARO (QTL Annotation Rice Online) database | http://qtaro.abr.affrc.go.jp/index.html, accessed on 18 January 2022 | Asian rice (O. sativa L.) |
| SNP genotype database for avocado | https://doi.org/10.1007/s11295-019-1374-1 (Table S2; [128]), accessed on 18 January 2022 | Avocado (Persea americana Mill.) |
| Sol Genomics Network | https://solgenomics.net, accessed on 18 January 2022 | Tomato (S. lycopersicum L.) |
| SorGSD | https://ngdc.cncb.ac.cn/sorgsd, accessed on 18 January 2022 | Sorghum (S. bicolor (L.) Moench) |
| SpinachBase | http://www.spinachbase.org, accessed on 19 January 2022 | Spinach (Spinacia oleracea L.) |
| Rice SNP-Seek Database | https://snp-seek.irri.org, accessed on 19 January 2022 | Asian rice (O. sativa L.) |
| The IPK Crop EST Database (CR-EST) | http://pgrc.ipk-gatersleben.de/cr-est, accessed on 19 January 2022 | Barley (H. vulgare L.) Bread wheat (T. eastivum L.) Pea (Pisum sativum L.) Potato (Solanum tuberosum L.) |
| The Tomato Integrated Database (Tomatonics) | http://plantomics.mind.meiji.ac.jp/tomatomics, accessed on 19 January 2022 | Tomato (S. lycopersicum L.) |
| TropGENE-DB | http://tropgenedb.cirad.fr/tropgene/JSP/index.jsp, accessed on 19 January 2022 | Asian rice (O. sativa L.) Banana (Musa acuminata Juss.) Bread fruit (Artocarpus altilis (Parkinson) Fosberg) Cassava (Manihot esculenta Crantz) Clemetine (Citrus clementina L.) Cocoa (Theobroma cacao L.) Coconut (Cocos nucifera L.) Coffee (Coffea canephora L.) Cupuassu (Theobroma grandiflorum Schum.) Oil palm (Elaeis guineensis Jacq.) Pummelo (Citrus grandis (L.) Osbeck) Sorghum (S. bicolor L. Moench) Sugarcane (Saccharum officinarus L.) Sweet orange (Citrus sinensis Osbeck) |
| Vitis International Variety Catalogue (VIVC) | https://www.vivc.de/index.php?r=site%2Findex, accessed on 19 January 2022 | Grapevine (Vitis spp.) |
| Crop | Species | Analytical Technique ‡ | Metabolite | Reference |
|---|---|---|---|---|
| Apple tree | Malus × domestica Borkh. | UHPLC–ESI-QTOF-MS, NMR | Flavonoids, polyphenols, sugars, terpenoids | [141] |
| Barley | H. vulgare | HPLC-FL, HPLC-MS, IC-MS/MS | Amino acids, glutathione, organic acids, starch, sugars, vitamin E (tocopherol) | [142] |
| HPAEC-PAD, HPLC-ELSD, HPLC-MALDITOF-MS | Sugars | [143] | ||
| HPLC-Fluorescence detection | Carotenoids (i.e., tocopherols and tocotrienols: vitamin E) | [144] | ||
| Barley Bread wheat Maize Potato Rice Sweet orange tree | H. vulgare T. aestivum Z.mays S. tuberosum O.sativa Citrus xsinensis (L.) Osbeck | GC-TOF-MS | Flavonoids | [145] |
| Blueberry | Vaccinium spp. | GC-MS | Fatty acids, phenylpropanoids, terpenoids | [146] |
| Bread wheat | T.aestivum | GC-MS | Amino acids, organic acid ‡‡ sugars | [147] |
| Foxtail millet | S.italica | HPLC-ESI-QTRAP-MS/MS | Alkaloids, amino acids, fatty acids, organic acids, phenolamides, polyphenols (i.e., flavonoids, anthocyanins...), sugars, vitamins | [148] |
| Lettuce | L.sativa | GC-TOF-MS | Alkaloids, amino acids, organic acids, polyamines, polyphenols, sugars, vitamins, etc. | [149] |
| Loquat | Eriobotryajaponica (Thunb.) Lindl. | UPLC-ESI-MS/MS | Alkaloids, flavonoids, phenolic acids, polysaccharides, terpenoids | [150] |
| Maize | Z.mays | LC-MS/MS | Fatty acids | [151] |
| LC-ESI-(QTRAP or QqTOF)-MS/MS | Amino acids, fatty acids, flavonoids | [152] | ||
| GC-MS | Amino acids, organic acids, phenylpropanoids | [153] | ||
| HPLC-Fluorescence detection | Tocochromanols (tocopherols and tocotrienols) | [154] | ||
| HPLC-PDA | Carotenoids | [155] | ||
| UPLC-HRMS | Amino acids, fatty acids, flavonoids, benzoxazinoids, terpenoids | [156] | ||
| HPLC, UPLC | Carotenoids | [157] | ||
| CEC | Amino acids | [158] | ||
| LC-ESI-QqTOF-MS/MS | Flavonoids | [159] | ||
| HPLC | Carotenoids | [160] | ||
| UPLC-PDA | Tocopherol (part of vitamin E) | [161] | ||
| GC-TOF-MS | Amino acids, (poly)amines, organic acids, sugars, vitamin E (tocopherol) | [162] | ||
| HPLC-PDA, HPLC-fluorescence detection | Carotenoids, phenolics, tocopherol (a form of vitamin E) | [163] | ||
| HPLC-fluorescence detection | Carotenoids (i.e., tocopherols and tocotrienols: vitamin E) | [164] | ||
| HPLC-PDA | Carotenoids (α-carotene, β-carotene, β-cryptoxanthin, lutein, phytofluene, zeaxanthin, zeinoxanthin) | [165] | ||
| HPLC-UV/Vis | Anthocyanins | [166] | ||
| Potato | S.tuberosum | UPLC-Q-TOF-MS | Alkaloids, amino acids | [167] |
| Rice | O.sativa | LC-ESI-Q TRAP-MS/MS | Phenolamides | [168] |
| GC-TOF-MS | Amino acids, flavonoids, organic acids | [169] | ||
| LC-ESI-MS/MS | Amino acids, fatty acids, flavonoids | [170] | ||
| HPLC-ESI-QTOF/MS | Amino acids, flavonoids, phenolamines, terpenoids | [171] | ||
| HPLC-ESI-(QTRAP or QqTOF)-MS | Amino acids, flavonoids, phenolamines, terpenoids | [70] | ||
| LC-ESI-Q TRAP-MS/MS | Flavonoids | [172] | ||
| Soybean | Glycinemax L. | GC | Fatty acids | [173] |
| HPLC-DAD | Isoflavones | [174] | ||
| HPLC-MS | Aminoacids, isoflavones, lipids, organic acids | [175] | ||
| Tea | Camelliasinensis L. | HPLC | Theanine, caffeine, catechins | [176] |
| HPLC-PDA | Amino acids, caffeine, catechins | [177] | ||
| Tomato | S.lycopersicum | GC-MS | Organic acids, sugars | [178] |
| GC-MS | Amino acids, organic acid ‡‡, sugars | [179] | ||
| HPLC-MS/MS | Alkaloids ‡‡‡ | [180] | ||
| GC-MS | Fatty acids, lipids, carotenoids (i.e., tocopherols and tocotrienols: vitamin E) | [181] | ||
| Wheat | T.aestivum | HPLC-ESI-QTRAP-MS/MS | Amino acids, (poly)amines, flavonoids, organic acids, sugars, vitamins, etc. | [182] |
| Technique | Crop | Method | Biofortified Trait | Reference |
|---|---|---|---|---|
| Conventional breeding | Rice | Backcrosses between a high-yielding cultivar and the IR68144 line | A 2.54-fold increase in iron and 1.54-fold increase in zinc | [4] |
| Maize | Backcrosses involving diverse exotic donor lines | Lines with high provitamin A content by accumulating mainly high β-carotene and lines with high provitamin A by promoting accumulation of high levels of both carotenes and xanthophylls | [13] | |
| Marker-assisted introgression of lpa1-1 and lpa2-1 alleles in elite lines of provitamin A-enriched quality protein maize (QPM) | A reduction in phytic acid content and improvement in the mineral bioavailability in lines of QPM rich in provitamin A | [197] | ||
| Introgression of VTE4 (γ-tocopherol methyl transferase) allele into four provitamin-A rich QPM elite inbreds using marker-assisted backcross breeding | An increase in α-tocopherol to 15.2 ppm over 8.0 ppm in the original inbreds | [14] | ||
| Wheat | Marker-assisted introgression of group 4 and 7 chromosomes of the wild ancestor Aegilops peregrina in a commercial variety of wheat | Higher content in iron and zinc in wheat grains | [5] | |
| Backcrosses between low-yielding exotic donor lines and commercial varieties | Black, purple, and blue lines with high content in anthocyanins | [38] | ||
| Cassava | Rapid cycling recurrent selection | Significant gains for total carotenoid content and total β-carotene | [15] | |
| Potato | ‘Atlantic’ and 17 4x-2x hybrids between S. tuberosum and diploid hybrids of Solanum phureja-Solanum stenotomum | Higher contents of copper, iron, manganese, and zinc | [6] | |
| Tomato | Backcrosses between landraces of tomato | Hybrid with increased concentration of polyphenols and high antioxidant activity in pink ripeness stage | [39] | |
| Bean | Backcrosses between low and high mineral genotypes using a QTL mapping approach | Increased iron and zinc content | [7] | |
| Chickpea | Crosses between different cultivars | Higher content of carotenoids | [16] | |
| Transgenesis | Rice | Endosperm-specific overexpression of Arabidopsis thaliana GTP cyclohydrolase I (GTPCHI) and aminodeoxychorismate synthase (ADCS) genes | An enhancement of 100 times in folate | [198] |
| Overexpression of phytoene synthase | Higher content in β-carotene | [17] | ||
| Expression of four synthetic genes: sZmPSY1, sPaCrtI, sCrBKT, and sHpBHY (for phytoene synthase, phytoene desaturase, β-carotene ketolase, and β-carotene hydroxylase, respectively) | Synthesis de novo of the carotenoid astaxanthin | [40] | ||
| Coexpression of an Arabidopsis nicotianamine synthase (AtNAS1), bean ferritin (PvFerritin), bacterial carotene desaturase (CRTI), and maize phytoene synthase (ZmPSY) | Simultaneous increase in iron, zinc, and β-carotene content in the rice endosperm | [29] | ||
| Constitutive overexpression of the rice GDP-L-galactose phosphorylase (35S-OsGGP) gene | Increase in ascorbate concentrations in germinated brown rice | [18] | ||
| Expression bacterial aspartate kinase (AK) and dihydrodipicolinate synthase (DHPS), downregulation of rice lysine ketoglutarate reductase/saccharopine dehydrogenase (LKR/SD) and selection of marker-free transgenic lines | Up to 25-fold increase in free lysine levels | [36] | ||
| Expression of an AmA1 gene from Amaranthus hypochondriacus | A significant increase in the content of several EAAs, including lysine, threonine, and valine, as well as a 1.06~12.87% increase in the total protein content | [37] | ||
| Maize | Overexpression of the bacterial genes crtB (for phytoene synthase) and crtI (for the four desaturation steps of the carotenoid pathway) under the control of a endosperm-specific promoter | An increase in total carotenoids of up to 34-fold with a preferential accumulation of β-carotene in the maize endosperm | [19] | |
| Endosperm-specific overexpression of soybean ferritin | A 2-fold improvement in seed iron bioavailability | [8] | ||
| Coexpression of Gm8gGCHI and GmADCS genes driven by endosperm-specific promoters | A 4.2-fold increase in folate (vitamin B9) level in transgenic maize grains | [20] | ||
| Insertion of the lysine-rich sb401 gene | Significantly higher levels of lysine total protein in maize seeds | [35] | ||
| Wheat | Constitutive expression of the rice nicotianamine synthase 2 (OsNAS2) gene | Higher concentrations of grain iron and zinc, and enhanced localization of iron and zinc in endosperm and crease tissues, respectively | [9] | |
| Cassava | Coexpression of ferritin (FER1) and mutated Iron transporter (IRT1) from A. thaliana | Accumulation of iron levels 7–18 times higher and zinc levels 3–10 times higher | [10] | |
| Potato | Overexpression of AtGTPCHI, AtADCS, OsHPPK/DHPS and AtFPGS genes | A 2-fold increase in folate content in mature tubers and stable accumulation of folates for up to 9 months of storage | [21] | |
| Simultaneous expression of Wrinkled 1 (WRI1), Diacylglycerol acyltransferase 1 (DGAT1) and Oleosin under the transcriptional control of tuber-specific (patatin) and constitutive (CaMV-35S) promoters. | Over a 100-fold increase in triacylglycerol accumulation to levels up to 3.3% of tuber dry weigh | [33] | ||
| Sweet Potato | Expression of a barley NA synthase 1 (HvNAS1) gene | A 3- and 2.9-fold increase in the concentrations of iron and zinc, respectively | [11] | |
| Tomato | Cross between GTPCHI and ADCS overexpressing plants | A 25-fold more in folate (Vitamin B9) level in fruits | [199] | |
| Overexpression of an A. thaliana Orange (AtOR) gene | An increase in total carotenoids in fruits | [22] | ||
| Overexpression of GDP-l-galactose phosphorylase (GGP) gene from Actinidia chinensis under the control of the 35S promoter | A 3- to 6-fold higher content in ascorbic acid in fruits | [200] | ||
| Fruit-specific expression of the transcription factor AtMYB12 | Increased content of different phenylpropanoids | [23] | ||
| Strawberry | Overexpression of a GDP-l-galactose phosphorylase (GGP) gene from Actinidia chinensis under the control of the 35S promoter | A 2-fold higher content in ascorbic acid in fruits | [200] | |
| Banana | Expression of a Fe’i banana-derived phytoene synthase (MtPsy2a) gene under the maize polyubiquitin promoter | Enhanced β-carotene content in fruit | [24] | |
| Soybean | Overexpression of the bacterial genes crtB (for phytoene synthase) and crtW and bkt1 (ketolase genes) under the control of seed-specific promoters | Enhanced accumulation of ketocarotenoids in seeds | [201] | |
| Overexpression of adenosine 5’-phosphosulfate sulfurylase 1 | Higher amounts of sulfate, cysteine, and some sulfur-containing secondary metabolites in seeds | [34] | ||
| Overexpression of a GmDGAT2A gene driven by a seed-specific promoter of Gmole1 | Significantly increased linoleic acid content specifically and total oil content | [32] | ||
| Bean | Seed-specific overexpression of a GTP cyclohydrolase I gene from Arabidopsis (AtGchI) | Increased folate levels in raw desiccated seeds by up to 3-fold | [25] | |
| Canola | Downregulation of lycopene ε-cyclase (ε-CYC) | Increased levels of β-carotene, zeaxanthin, violaxanthin, and lutein | [26] | |
| Brassicacarinata | Expression of an 18-carbon ω3 desaturase (CpDesX) gene from Claviceps purpurea and a 20-carbon ω3 desaturase (Pir-ω3) gene from Pythium irregulare | Up to 25% increase in eicosapentaenoic acid | [31] | |
| Linseed | Expression of a Δ6-desaturase from Primula vialii | Transgenic lines that accumulate the omega-3 fatty acid stearidonic acid | [30] | |
| Cisgenesis | Barley | Expression of a barley phytase gene (HvPAPhy_a) | Decrease in phytate concentration, which then increases phosphate bioavailability | [202] |
| Potato | Suppression of a starch phosphorylase L. gene through dsRNAi technology | Decrease in starch degradation what reduces the accumulation of reducing (glucose, fructose) and non-reducing (sucrose) sugars in tubers stored at 4 °C | [203] | |
| Apple | Expression of MdMYB10 transcription factor | Red-fleshed ‘Gala’ apples rich in anthocyanins | [41] | |
| Intragenesis | Potato | Silencing of a granule-bound starch synthase (GBSS) gene | An increase in amylopectin content | [204] |
| Silencing of an asparagine synthase gene (StAs1) | Reduced free asparagine concentration by up to 80% and consequent decrease in acrylamide content in processed potato | [205] | ||
| Overexpression of a lycopene b-cyclase (StLYCb) gene under the GBSS promoter | An increase in β-carotene accumulation in potato tubers | [27] | ||
| Tomato | Suppression of a DE-ETIOLATED1 (DET1) gene through RNAi technology | Enhanced carotenoid and flavonoid content | [28] | |
| Wheat | Suppression of a γ-gliadin gene by using RNAi technology | Gluten-free wheat | [206] | |
| Overexpression of a vacuolar Iron transporter (TaVIT2) under the control of a wheat endosperm-specific promoter | An increase in more than 2-fold of iron in white flour fractions | [12] | ||
| Soybean | RNAi technology | Plenish® high oleic | Dupont-Pioneer (Johnston, IA, USA) | |
| Vistive® Gold low saturated high oleic | Monsanto (St. Louis, MO, USA) |
| Characteristic | Conventional Breeding | Transgenesis | Cisgenesis | Intragenesis |
|---|---|---|---|---|
| Variability source | Sexually compatible group | Any organism | Sexually compatible group | Sexually compatible group |
| Method | Crosses and selection | Recombinant DNA | By Agrobacterium | By Agrobacterium (recombinant DNA) |
| Introducing DNA | Natural | Natural and/or artificial | Natural | Natural and/or artificial |
| Gene pool | Unaltered | Altered | Unaltered | Altered |
| Borders | - | T-DNA | T-DNA (to be eliminated) | T-DNA or P-DNA |
| Linkage drag | Yes | No | No | No |
| Expression modulation | No | Yes | Yes | Yes |
| Time | High | Medium | Medium | Medium |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina-Lozano, I.; Díaz, A. Applications of Genomic Tools in Plant Breeding: Crop Biofortification. Int. J. Mol. Sci. 2022, 23, 3086. https://doi.org/10.3390/ijms23063086
Medina-Lozano I, Díaz A. Applications of Genomic Tools in Plant Breeding: Crop Biofortification. International Journal of Molecular Sciences. 2022; 23(6):3086. https://doi.org/10.3390/ijms23063086
Chicago/Turabian StyleMedina-Lozano, Inés, and Aurora Díaz. 2022. "Applications of Genomic Tools in Plant Breeding: Crop Biofortification" International Journal of Molecular Sciences 23, no. 6: 3086. https://doi.org/10.3390/ijms23063086