
The Plastome Sequences of Triticum sphaerococcum (ABD) and Triticum turgidum subsp. durum (AB) Exhibit Evolutionary Changes, Structural Characterization, Comparative Analysis, Phylogenomics and Time Divergence

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Material and Methods
2.1. Sequencing, Assembly and Annotation
2.2. Characterization of Repetitive Sequences and SSRs
2.3. Sequence Divergence, Phylogenetic Analyses, and Divergence Time
3. Results
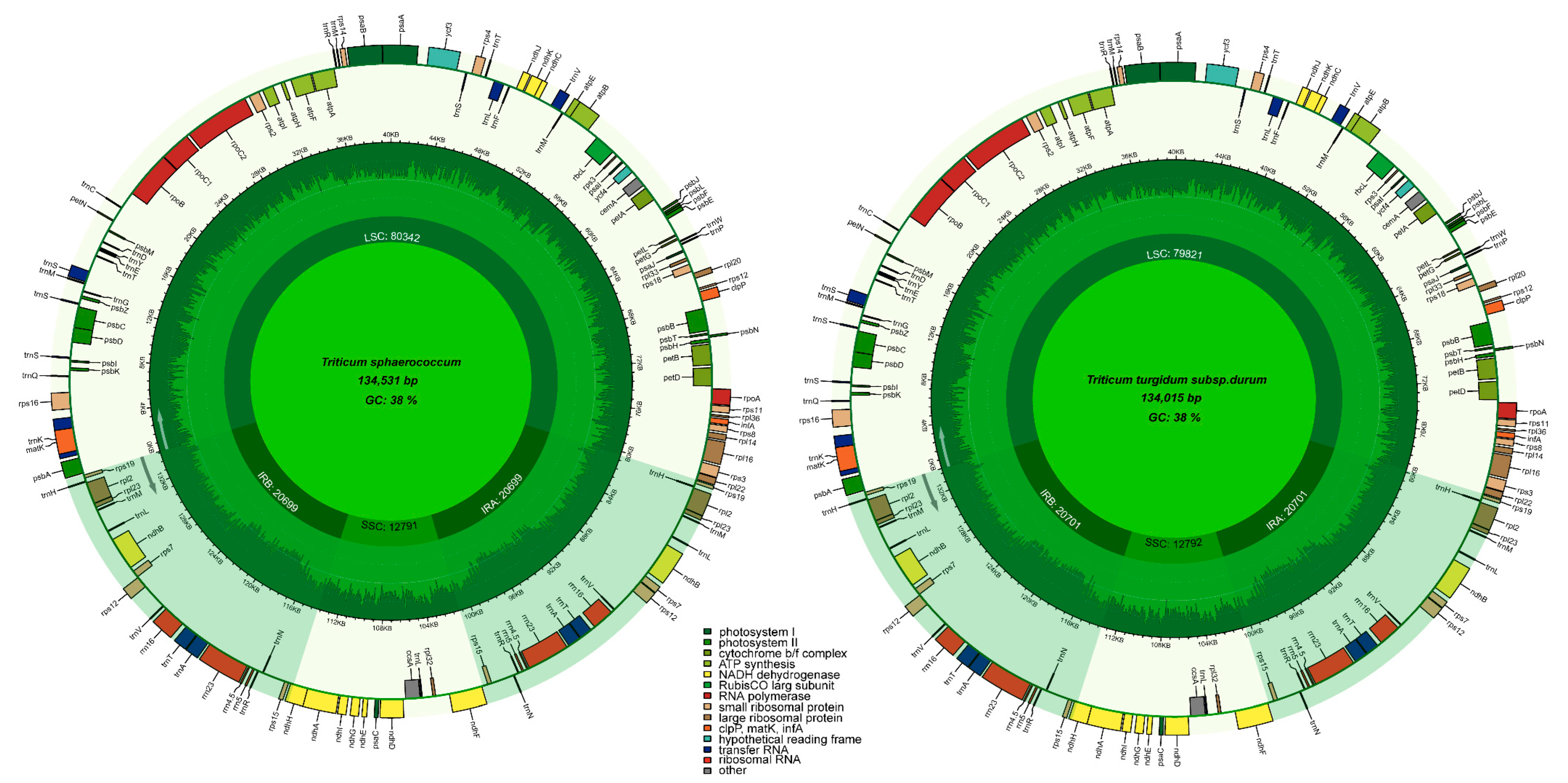
3.1. Triticum Plastome Characteristics (Genome Size and GC Content Variation)
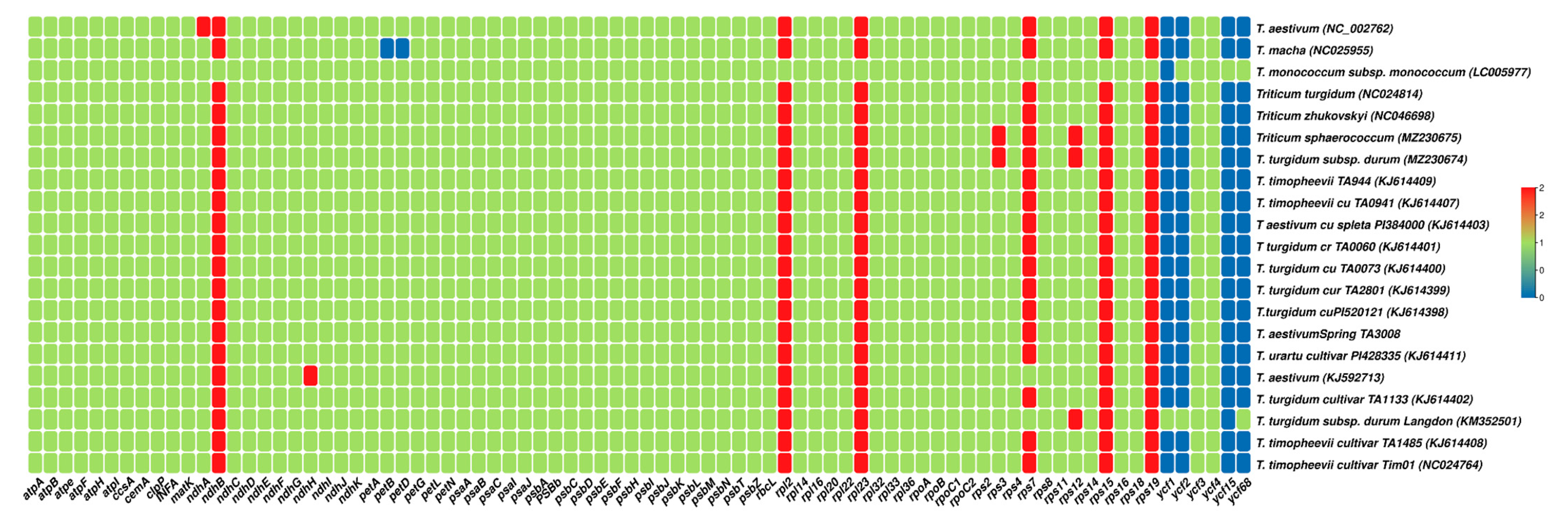
3.2. Gene Content and Gene Loss in Triticum Plastomes
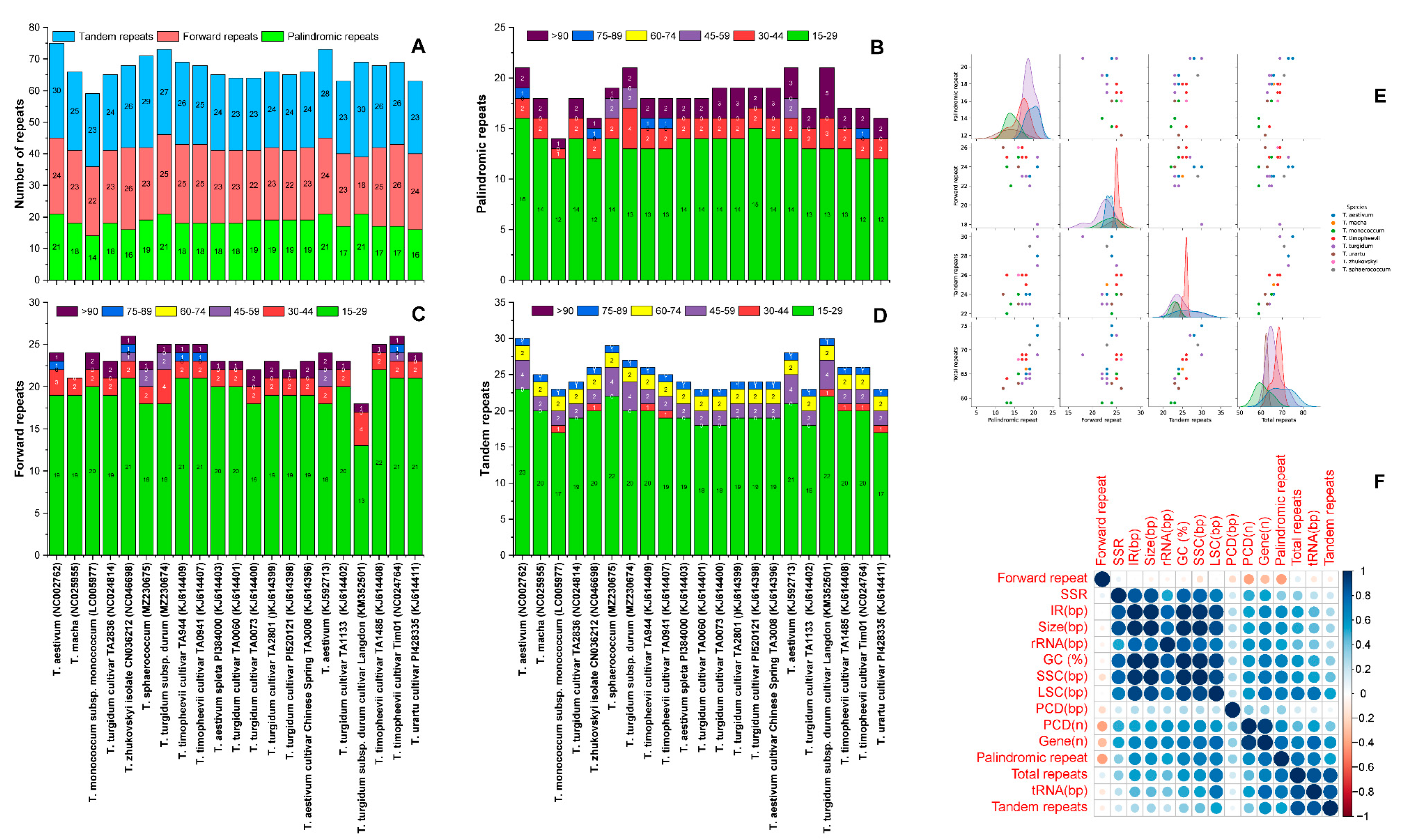
3.3. Functional Repeats within Triticum Plastomes
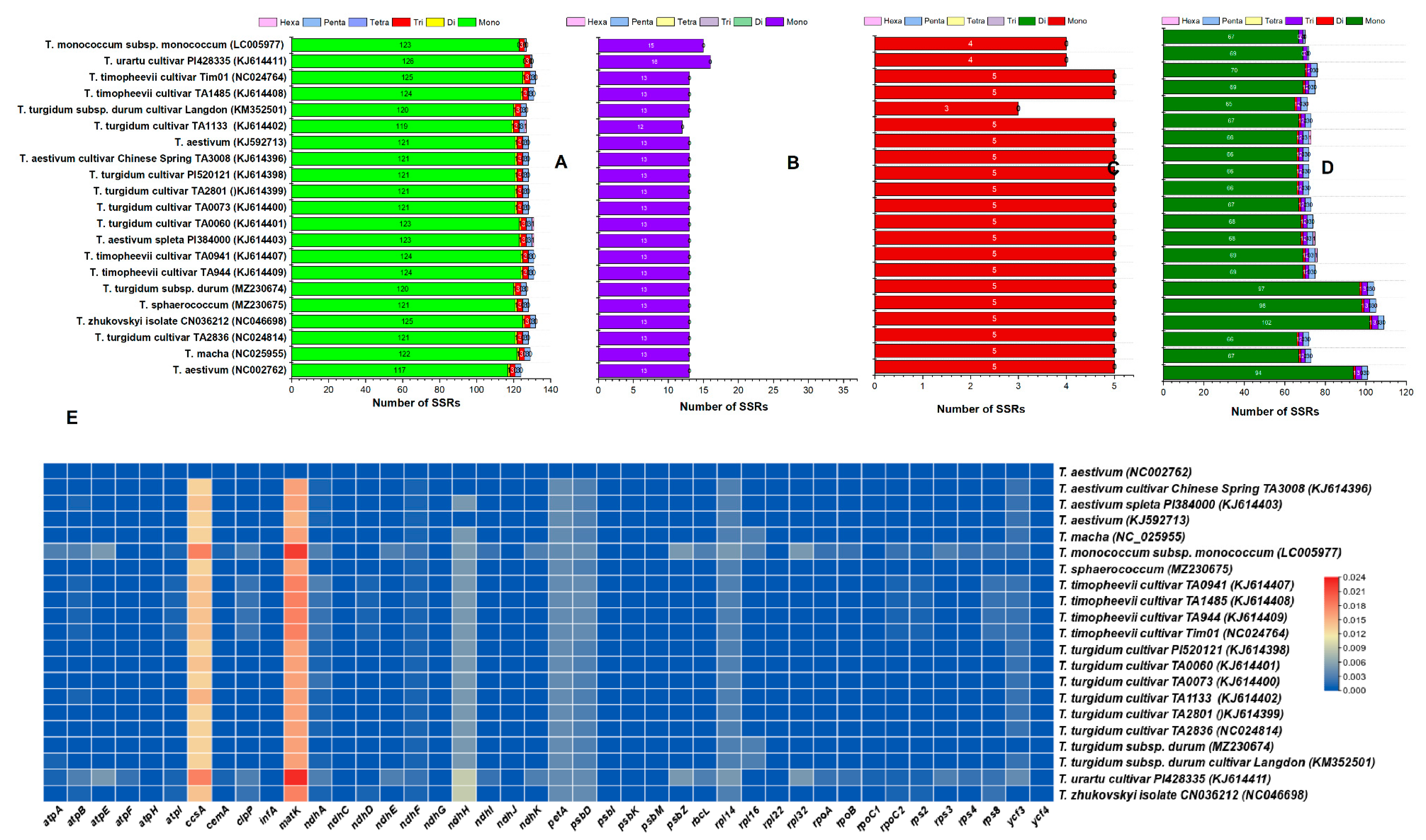
3.4. Simple Sequence Repeat (SSR) Analysis in Triticum Plastomes
3.5. Comparative Analysis and Divergence of Triticum Plastomes
3.6. Evolution and Origin of IRs in Triticum Plastomes
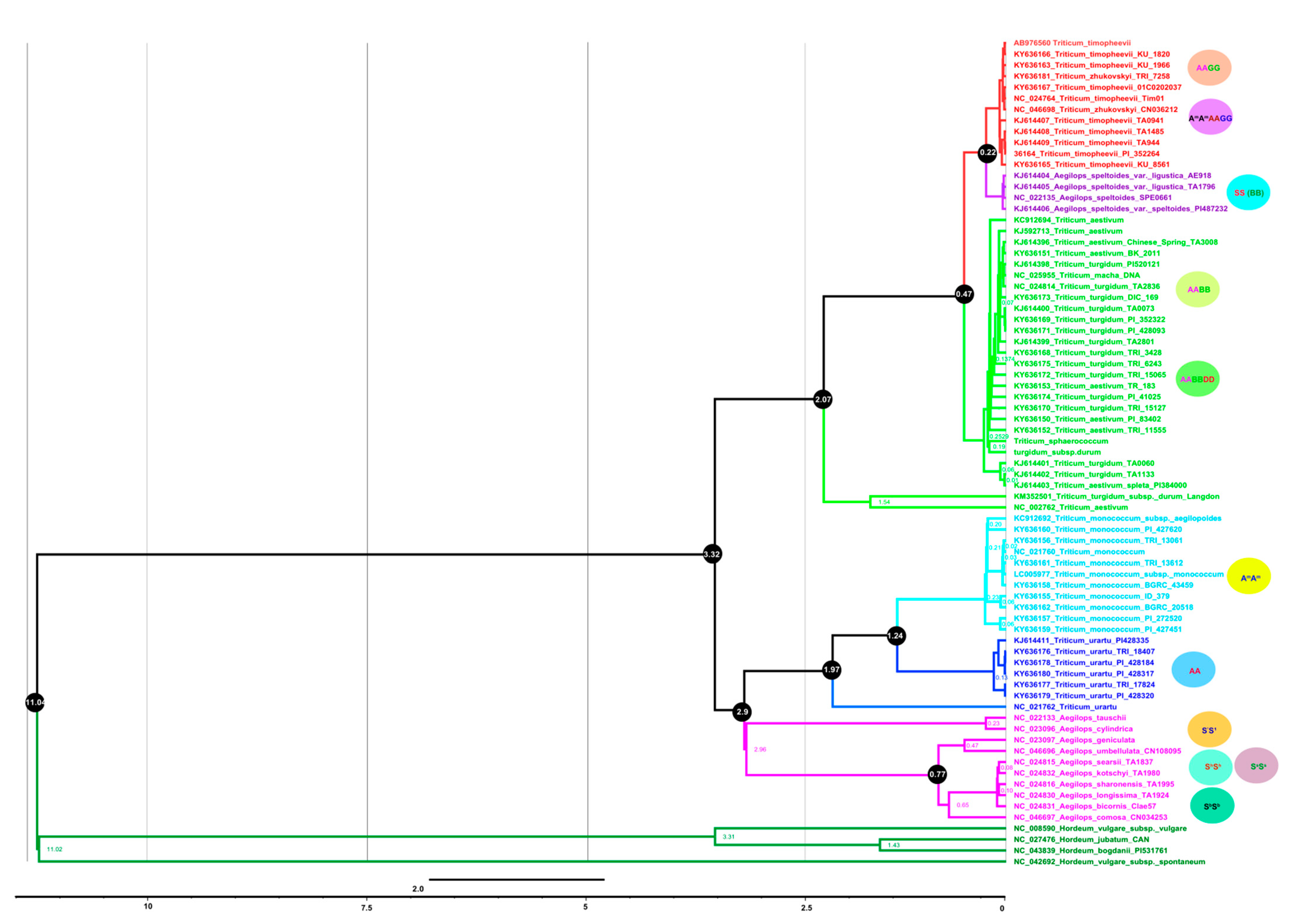
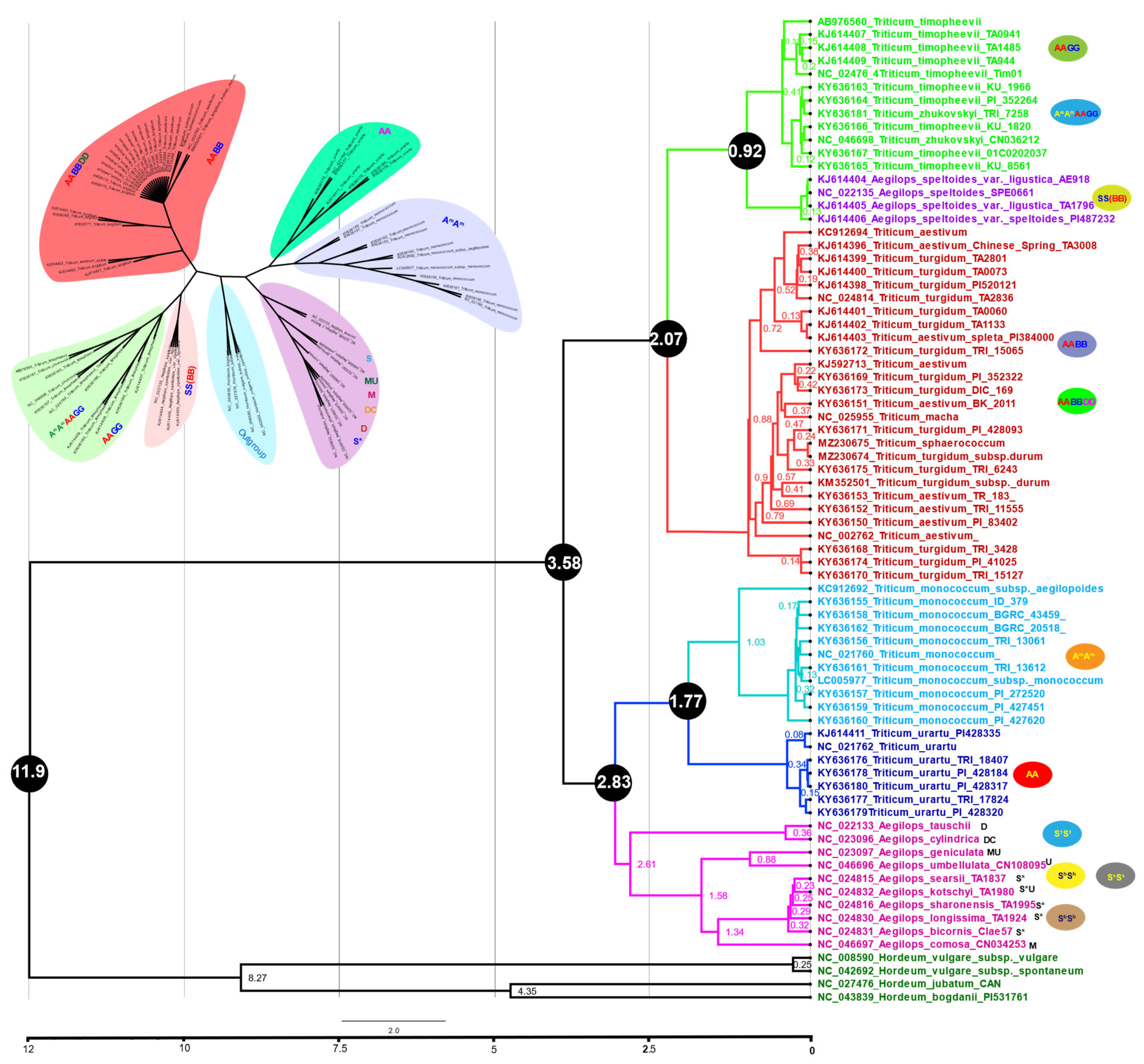
3.7. Plastome Phylogenomics and Diversification of Triticum Plastomes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salamini, F.; Özkan, H.; Brandolini, A.; Schäfer-Pregl, R.; Martin, W. Genetics and geography of wild cereal domestication in the near east. Nat. Rev. Genet. 2002, 3, 429–441. [Google Scholar] [CrossRef]
- Gill, B.; Friebe, B. Cytogenetics, Phylogeny and Evolution of Cultivated Wheats. Bread Wheat: Improvement and Production; Food and Agriculture Organization of the United Nations: Rome, Italy, 2002; pp. 71–88. [Google Scholar]
- Dvorak, J.; McGuire, P.E.; Cassidy, B. Apparent sources of the A genomes of wheats inferred from polymorphism in abundance and restriction fragment length of repeated nucleotide sequences. Genome 1988, 30, 680–689. [Google Scholar] [CrossRef]
- Kerby, K.; Kuspira, J.; Jones, B. Biochemical data bearing on the relationship between the genome of Triticum urartu and the A and B genomes of the polyploid wheats. Genome 1988, 30, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-Z.; Miyashita, N.T.; Tsunewaki, K. Plasmon analyses of Triticum (wheat) and Aegilops: PCR–single-strand conformational polymorphism (PCR-SSCP) analyses of organellar DNAs. Proc. Natl. Acad. Sci. USA 1997, 94, 14570–14577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimber, G. A reassessment of the origin of the polyploid wheats. Genetics 1974, 78, 487–492. [Google Scholar] [CrossRef]
- Dvořák, J.; Zhang, H.-B. Reconstruction of the phylogeny of the genus Triticum from variation in repeated nucleotide sequences. Theor. Appl. Genet. 1992, 84, 419–429. [Google Scholar] [CrossRef]
- Jenkins, J. Chromosome homologies in wheat and Aegilops. Am. J. Bot. 1929, 16, 238–245. [Google Scholar] [CrossRef]
- Haider, N. The origin of the B-genome of bread wheat (Triticum aestivum L.). Russ. J. Genet. 2013, 49, 263–274. [Google Scholar] [CrossRef]
- Escobar, J.S.; Scornavacca, C.; Cenci, A.; Guilhaumon, C.; Santoni, S.; Douzery, E.J.; Ranwez, V.; Glémin, S.; David, J. Multigenic phylogeny and analysis of tree incongruences in Triticeae (Poaceae). BMC Evol. Biol. 2011, 11, 181. [Google Scholar] [CrossRef] [Green Version]
- Marcussen, T.; Sandve, S.R.; Heier, L.; Spannagl, M.; Pfeifer, M.; Jakobsen, K.S.; Wulff, B.B.; Steuernagel, B.; Mayer, K.F.; Olsen, O.-A. Ancient hybridizations among the ancestral genomes of bread wheat. Science 2014, 345, 1250092. [Google Scholar] [CrossRef]
- Tsunewaki, K.; Ogihara, Y. The molecular basis of genetic diversity among cytoplasms of Triticum and Aegilops species. II. On the origin of polyploid wheat cytoplasms as suggested by chloroplast DNA restriction fragment patterns. Genetics 1983, 104, 155–171. [Google Scholar] [CrossRef] [PubMed]
- Tsunewaki, K. Plasmon analysis in the Triticum-Aegilops complex. Breed. Sci. 2009, 59, 455–470. [Google Scholar] [CrossRef] [Green Version]
- Sang, T. Utility of low-copy nuclear gene sequences in plant phylogenetics. Crit. Rev. Biochem. Mol. Biol. 2002, 37, 121–147. [Google Scholar] [CrossRef]
- Baenfer, G.; Moog, U.; Fiala, B.; Mohamed, M.; Weising, K.; Blattner, F.R. A chloroplast genealogy of myrmecophytic Macaranga species (Euphorbiaceae) in Southeast Asia reveals hybridization, vicariance and long-distance dispersals. Mol. Ecol. 2006, 15, 4409–4424. [Google Scholar] [CrossRef] [PubMed]
- Bordbar, F.; Rahiminejad, M.R.; Saeidi, H.; Blattner, F.R. Phylogeny and genetic diversity of D-genome species of Aegilops and Triticum (Triticeae, Poaceae) from Iran based on microsatellites, ITS, and trn LF. Plant Syst. Evol. 2011, 291, 117–131. [Google Scholar] [CrossRef]
- Petersen, G.; Seberg, O. Phylogenetic Analysis of the Triticeae (Poaceae) Based onrpoA Sequence Data. Mol. Phylogenetics Evol. 1997, 7, 217–230. [Google Scholar] [CrossRef] [PubMed]
- Golovnina, K.; Glushkov, S.; Blinov, A.; Mayorov, V.; Adkison, L.; Goncharov, N. Molecular phylogeny of the genus Triticum L. Plant Syst. Evol. 2007, 264, 195–216. [Google Scholar] [CrossRef]
- Seberg, O.; Petersen, G. Phylogeny of Triticeae (Poaceae) based on three organelle genes, two single-copy nuclear genes, and morphology. Aliso J. Syst. Evol. Bot. 2007, 23, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Bernhardt, N.; Brassac, J.; Kilian, B.; Blattner, F.R. Dated tribe-wide whole chloroplast genome phylogeny indicates recurrent hybridizations within Triticeae. BMC Evol. Biol. 2017, 17, 141. [Google Scholar] [CrossRef] [Green Version]
- Ogihara, Y.; Isono, K.; Kojima, T.; Endo, A.; Hanaoka, M.; Shiina, T.; Terachi, T.; Utsugi, S.; Murata, M.; Mori, N. Chinese spring wheat (Triticum aestivum L.) chloroplast genome: Complete sequence and contig clones. Plant Mol. Biol. Report. 2000, 18, 243–253. [Google Scholar] [CrossRef]
- Gornicki, P.; Zhu, H.; Wang, J.; Challa, G.S.; Zhang, Z.; Gill, B.S.; Li, W. The chloroplast view of the evolution of polyploid wheat. New Phytol. 2014, 204, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Middleton, C.P.; Senerchia, N.; Stein, N.; Akhunov, E.D.; Keller, B.; Wicker, T.; Kilian, B. Sequencing of chloroplast genomes from wheat, barley, rye and their relatives provides a detailed insight into the evolution of the Triticeae tribe. PLoS ONE 2014, 9, e85761. [Google Scholar] [CrossRef] [Green Version]
- Saarela, J.M.; Wysocki, W.P.; Barrett, C.F.; Soreng, R.J.; Davis, J.I.; Clark, L.G.; Kelchner, S.A.; Pires, J.C.; Edger, P.P.; Mayfield, D.R. Plastid phylogenomics of the cool-season grass subfamily: Clarification of relationships among early-diverging tribes. AoB Plants 2015, 7, plv046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Q.X.; Yuan, J.H.; Wang, L.Y.; Xu, J.Q.; Nyima, T. The complete chloroplast genome of Tibetan hulless barley. Mitochondrial DNA Part A 2017, 28, 324–325. [Google Scholar] [CrossRef] [PubMed]
- Gogniashvili, M.; Jinjikhadze, T.; Maisaia, I.; Akhalkatsi, M.; Kotorashvili, A.; Kotaria, N.; Beridze, T.; Dudnikov, A.J. Complete chloroplast genomes of Aegilops tauschii Coss. and Ae. cylindrica Host sheds light on plasmon D evolution. Curr. Genet. 2016, 62, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Shizuka, T.; Mori, N.; Ozkan, H.; Ohta, S. Chloroplast DNA haplotype variation within two natural populations of wild emmer wheat (Triticum turgidum ssp. dicoccoides) in southern Turkey. Biotechnol. Biotechnol. Equip. 2015, 29, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Gogniashvili, M.; Naskidashvili, P.; Bedoshvili, D.; Kotorashvili, A.; Kotaria, N.; Beridze, T. Complete chloroplast DNA sequences of Zanduri wheat (Triticum spp.). Genet. Resour. Crop Evol. 2015, 62, 1269–1277. [Google Scholar] [CrossRef]
- Catalán, P.; Kellogg, E.A.; Olmstead, R.G. Phylogeny of Poaceae Subfamily Pooideae Based on ChloroplastndhF Gene Sequences. Mol. Phylogenetics Evol. 1997, 8, 150–166. [Google Scholar] [CrossRef]
- Jansen, R.K.; Kaittanis, C.; Saski, C.; Lee, S.-B.; Tomkins, J.; Alverson, A.J.; Daniell, H. Phylogenetic analyses of Vitis (Vitaceae) based on complete chloroplast genome sequences: Effects of taxon sampling and phylogenetic methods on resolving relationships among rosids. BMC Evol. Biol. 2006, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Ogihara, Y.; Isono, K.; Kojima, T.; Endo, A.; Hanaoka, M.; Shiina, T.; Terachi, T.; Utsugi, S.; Murata, M.; Mori, N. Structural features of a wheat plastome as revealed by complete sequencing of chloroplast DNA. Mol. Genet. Genom. 2002, 266, 740–746. [Google Scholar] [CrossRef]
- Saski, C.; Lee, S.-B.; Fjellheim, S.; Guda, C.; Jansen, R.K.; Luo, H.; Tomkins, J.; Rognli, O.A.; Daniell, H.; Clarke, J.L. Complete chloroplast genome sequences of Hordeum vulgare, Sorghum bicolor and Agrostis stolonifera, and comparative analyses with other grass genomes. Theor. Appl. Genet. 2007, 115, 571–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaf, S.; Waqas, M.; Khan, A.L.; Khan, M.A.; Kang, S.-M.; Imran, Q.M.; Shahzad, R.; Bilal, S.; Yun, B.-W.; Lee, I.-J. The complete chloroplast genome of wild rice (Oryza minuta) and its comparison to related species. Front. Plant Sci. 2017, 8, 304. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Hu, N.; Huang, H.; Gao, J.; Zhao, Y.-J.; Gao, L.-Z. An improved chloroplast DNA extraction procedure for whole plastid genome sequencing. PLoS ONE 2012, 7, e31468. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.-J.; Yu, W.-B.; Yang, J.-B.; Song, Y.; Yi, T.-S.; Li, D.-Z. GetOrganelle: A simple and fast pipeline for de novo assembly of a complete circular chloroplast genome using genome skimming data. bioRxiv 2018, 4, 256479. [Google Scholar]
- Liu, C.; Shi, L.; Zhu, Y.; Chen, H.; Zhang, J.; Lin, X.; Guan, X. CpGAVAS, an integrated web server for the annotation, visualization, analysis, and GenBank submission of completely sequenced chloroplast genome sequences. BMC Genom. 2012, 13, 715. [Google Scholar] [CrossRef] [Green Version]
- Wyman, S.K.; Jansen, R.K.; Boore, J.L. Automatic annotation of organellar genomes with DOGMA. Bioinformatics 2004, 20, 3252–3255. [Google Scholar] [CrossRef] [Green Version]
- Lowe, T.M.; Eddy, S.R. tRNAscan-SE: A program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res. 1997, 25, 955–964. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis (Use R!); Springer: New York, NY, USA, 2009; Volume 10, pp. 970–978. [Google Scholar]
- Kurtz, S.; Choudhuri, J.V.; Ohlebusch, E.; Schleiermacher, C.; Stoye, J.; Giegerich, R. REPuter: The manifold applications of repeat analysis on a genomic scale. Nucleic Acids Res. 2001, 29, 4633–4642. [Google Scholar] [CrossRef] [Green Version]
- Benson, G. Tandem repeats finder: A program to analyze DNA sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [Green Version]
- Beier, S.; Thiel, T.; Münch, T.; Scholz, U.; Mascher, M. MISA-web: A web server for microsatellite prediction. Bioinformatics 2017, 33, 2583–2585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frazer, K.A.; Pachter, L.; Poliakov, A.; Rubin, E.M.; Dubchak, I. VISTA: Computational tools for comparative genomics. Nucleic Acids Res. 2004, 32, W273–W279. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Crisp, M.D.; Cook, L.G. Cenozoic extinctions account for the low diversity of extant gymnosperms compared with angiosperms. New Phytol. 2011, 192, 997–1009. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1. 3.1. 2009. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 25 February 2022).
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.-H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Asaf, S.; Khan, A.L.; Khan, A.; Khan, G.; Lee, I.-J.; Al-Harrasi, A. Expanded inverted repeat region with large scale inversion in the first complete plastid genome sequence of Plantago ovata. Sci. Rep. 2020, 10, 3881. [Google Scholar] [CrossRef] [Green Version]
- Chalupska, D.; Lee, H.; Faris, J.; Evrard, A.; Chalhoub, B.; Haselkorn, R.; Gornicki, P. Acc homoeoloci and the evolution of wheat genomes. Proc. Natl. Acad. Sci. USA 2008, 105, 9691–9696. [Google Scholar] [CrossRef] [Green Version]
- Won, H.; Renner, S.S. Dating dispersal and radiation in the gymnosperm Gnetum (Gnetales)—Clock calibration when outgroup relationships are uncertain. Syst. Biol. 2006, 55, 610–622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, S.; Sirikhachornkit, A.; Su, X.; Faris, J.; Gill, B.; Haselkorn, R.; Gornicki, P. Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of the Triticum/Aegilops complex and the evolutionary history of polyploid wheat. Proc. Natl. Acad. Sci. USA 2002, 99, 8133–8138. [Google Scholar] [CrossRef] [Green Version]
- Wolfe, K.H.; Morden, C.W.; Palmer, J.D. Function and evolution of a minimal plastid genome from a nonphotosynthetic parasitic plant. Proc. Natl. Acad. Sci. USA 1992, 89, 10648–10652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.-J.; Fan, S.-J.; Wicke, S.; Yi, T.-S. Plastome reduction in the only parasitic gymnosperm Parasitaxus is due to losses of photosynthesis but not housekeeping genes and apparently involves the secondary gain of a large inverted repeat. Genome Biol. Evol. 2019, 11, 2789–2796. [Google Scholar] [CrossRef]
- Huang, Y.-Y.; Cho, S.-T.; Haryono, M.; Kuo, C.-H. Complete chloroplast genome sequence of common bermudagrass (Cynodon dactylon (L.) Pers.) and comparative analysis within the family Poaceae. PLoS ONE 2017, 12, e0179055. [Google Scholar]
- Khan, A.L.; Al-Harrasi, A.; Asaf, S.; Park, C.E.; Park, G.-S.; Khan, A.R.; Lee, I.-J.; Al-Rawahi, A.; Shin, J.-H. The first chloroplast genome sequence of Boswellia sacra, a resin-producing plant in Oman. PLoS ONE 2017, 12, e0169794. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.L.; Asaf, S.; Al-Rawahi, A.; Al-Harrasi, A. Decoding first complete chloroplast genome of toothbrush tree (Salvadora persica L.): Insight into genome evolution, sequence divergence and phylogenetic relationship within Brassicales. BMC Genom. 2021, 22, 312. [Google Scholar] [CrossRef]
- Nie, X.; Lv, S.; Zhang, Y.; Du, X.; Wang, L.; Biradar, S.S.; Tan, X.; Wan, F.; Weining, S. Complete chloroplast genome sequence of a major invasive species, crofton weed (Ageratina adenophora). PLoS ONE 2012, 7, e36869. [Google Scholar] [CrossRef] [Green Version]
- Raubeson, L.A.; Peery, R.; Chumley, T.W.; Dziubek, C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Comparative chloroplast genomics: Analyses including new sequences from the angiosperms Nuphar advena and Ranunculus macranthus. BMC Genom. 2007, 8, 174. [Google Scholar] [CrossRef] [Green Version]
- Frailey, D.C.; Chaluvadi, S.R.; Vaughn, J.N.; Coatney, C.G.; Bennetzen, J.L. Gene loss and genome rearrangement in the plastids of five Hemiparasites in the family Orobanchaceae. BMC Plant Biol. 2018, 18, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oyebanji, O.; Zhang, R.; Chen, S.-Y.; Yi, T.-S. Corrigendum: New Insights into the Plastome Evolution of the Millettioid/Phaseoloid Clade (Papilionoideae, Leguminosae). Front. Plant Sci. 2021, 12, 652483. [Google Scholar] [CrossRef] [PubMed]
- Millen, R.S.; Olmstead, R.G.; Adams, K.L.; Palmer, J.D.; Lao, N.T.; Heggie, L.; Kavanagh, T.A.; Hibberd, J.M.; Gray, J.C.; Morden, C.W. Many parallel losses of infA from chloroplast DNA during angiosperm evolution with multiple independent transfers to the nucleus. Plant Cell 2001, 13, 645–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magee, A.M.; Aspinall, S.; Rice, D.W.; Cusack, B.P.; Sémon, M.; Perry, A.S.; Stefanović, S.; Milbourne, D.; Barth, S.; Palmer, J.D. Localized hypermutation and associated gene losses in legume chloroplast genomes. Genome Res. 2010, 20, 1700–1710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haberle, R.C.; Fourcade, H.M.; Boore, J.L.; Jansen, R.K. Extensive rearrangements in the chloroplast genome of Trachelium caeruleum are associated with repeats and tRNA genes. J. Mol. Evol. 2008, 66, 350–361. [Google Scholar] [CrossRef] [PubMed]
- Cheon, K.-S.; Kim, K.-A.; Jang, S.-K.; Yoo, K.-O. Complete chloroplast genome sequence of Campanula takesimana (Campanulaceae), an endemic to Korea. Mitochondrial DNA Part A 2016, 27, 2169–2171. [Google Scholar]
- Blazier, J.C.; Guisinger, M.M.; Jansen, R.K. Recent loss of plastid-encoded ndh genes within Erodium (Geraniaceae). Plant Mol. Biol. 2011, 76, 263–272. [Google Scholar] [CrossRef]
- Lee, H.-L.; Jansen, R.K.; Chumley, T.W.; Kim, K.-J. Gene relocations within chloroplast genomes of Jasminum and Menodora (Oleaceae) are due to multiple, overlapping inversions. Mol. Biol. Evol. 2007, 24, 1161–1180. [Google Scholar] [CrossRef] [Green Version]
- Konishi, T.; Shinohara, K.; Yamada, K.; Sasaki, Y. Acetyl-CoA carboxylase in higher plants: Most plants other than gramineae have both the prokaryotic and the eukaryotic forms of this enzyme. Plant Cell Physiol. 1996, 37, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Palmer, J.D.; Nugent, J.M.; Herbon, L.A. Unusual structure of geranium chloroplast DNA: A triple-sized inverted repeat, extensive gene duplications, multiple inversions, and two repeat families. Proc. Natl. Acad. Sci. USA 1987, 84, 769–773. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.-Z.; Liu, Y.-L.; Zhang, D.; Li, W.; Gao, J.; Liu, Y.; Li, K.; Shi, C.; Zhao, Y.; Zhao, Y.-J. Evolution of Oryza chloroplast genomes promoted adaptation to diverse ecological habitats. Commun. Biol. 2019, 2, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, A.; Asaf, S.; Khan, A.L.; Al-Harrasi, A.; Al-Sudairy, O.; AbdulKareem, N.M.; Khan, A.; Shehzad, T.; Alsaady, N.; Al-Lawati, A. First complete chloroplast genomics and comparative phylogenetic analysis of Commiphora gileadensis and C. foliacea: Myrrh producing trees. PLoS ONE 2019, 14, e0208511. [Google Scholar]
- Hirao, T.; Watanabe, A.; Kurita, M.; Kondo, T.; Takata, K. Complete nucleotide sequence of the Cryptomeria japonica D. Don. chloroplast genome and comparative chloroplast genomics: Diversified genomic structure of coniferous species. BMC Plant Biol. 2008, 8, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, X.; Gao, L.; Wang, B.; Su, Y.-J.; Wang, T. The complete chloroplast genome sequence of Cephalotaxus oliveri (Cephalotaxaceae): Evolutionary comparison of Cephalotaxus chloroplast DNAs and insights into the loss of inverted repeat copies in gymnosperms. Genome Biol. Evol. 2013, 5, 688–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Ranade, S.S.; Lin, Y.-C.; Zuccolo, A.; Van de Peer, Y.; del Rosario García-Gil, M. Comparative in silico analysis of EST-SSRs in angiosperm and gymnosperm tree genera. BMC Plant Biol. 2014, 14, 220. [Google Scholar] [CrossRef] [Green Version]
- Provan, J.; Powell, W.; Hollingsworth, P.M. Chloroplast microsatellites: New tools for studies in plant ecology and evolution. Trends Ecol. Evol. 2001, 16, 142–147. [Google Scholar] [CrossRef]
- Jakobsson, M.; Säll, T.; Lind-Halldén, C.; Halldén, C. Evolution of chloroplast mononucleotide microsatellites in Arabidopsis thaliana. Theor. Appl. Genet. 2007, 114, 223. [Google Scholar] [CrossRef]
- Timme, R.E.; Kuehl, J.V.; Boore, J.L.; Jansen, R.K. A comparative analysis of the Lactuca and Helianthus (Asteraceae) plastid genomes: Identification of divergent regions and categorization of shared repeats. Am. J. Bot. 2007, 94, 302–312. [Google Scholar] [CrossRef]
- Gao, L.; Yi, X.; Yang, Y.-X.; Su, Y.-J.; Wang, T. Complete chloroplast genome sequence of a tree fern Alsophila spinulosa: Insights into evolutionary changes in fern chloroplast genomes. BMC Evol. Biol. 2009, 9, 130. [Google Scholar] [CrossRef] [Green Version]
- Menezes, A.P.; Resende-Moreira, L.C.; Buzatti, R.S.; Nazareno, A.G.; Carlsen, M.; Lobo, F.P.; Kalapothakis, E.; Lovato, M.B. Chloroplast genomes of Byrsonima species (Malpighiaceae): Comparative analysis and screening of high divergence sequences. Sci. Rep. 2018, 8, 2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khakhlova, O.; Bock, R. Elimination of deleterious mutations in plastid genomes by gene conversion. Plant J. 2006, 46, 85–94. [Google Scholar] [CrossRef]
- Yue, F.; Cui, L.; Claude, W.d.; Moret, B.M.; Tang, J. Gene rearrangement analysis and ancestral order inference from chloroplast genomes with inverted repeat. BMC Genom. 2008, 9, S25. [Google Scholar] [CrossRef] [Green Version]
- Asaf, S.; Khan, A.L.; Khan, A.R.; Waqas, M.; Kang, S.-M.; Khan, M.A.; Lee, S.-M.; Lee, I.-J. Complete chloroplast genome of Nicotiana otophora and its comparison with related species. Front. Plant Sci. 2016, 7, 843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsiao, C.; Chatterton, N.; Asay, K.; Jensen, K. Phylogenetic relationships of the monogenomic species of the wheat tribe, Triticeae (Poaceae), inferred from nuclear rDNA (internal transcribed spacer) sequences. Genome 1995, 38, 211–223. [Google Scholar] [CrossRef] [PubMed]
- Kellogg, E.A.; Appels, R.; Mason-Gamer, R.J. When genes tell different stories: The diploid genera of Triticeae (Gramineae). Syst. Bot. 1996, 321–347. [Google Scholar] [CrossRef]
- De Bustos, A.; Jouve, N. Phylogenetic relationships of the genus Secale based on the characterisation of rDNA ITS sequences. Plant Syst. Evol. 2002, 235, 147–154. [Google Scholar] [CrossRef]
- Mason-Gamer, R.J.; Orme, N.L.; Anderson, C.M. Phylogenetic analysis of North American Elymus and the monogenomic Triticeae (Poaceae) using three chloroplast DNA data sets. Genome 2002, 45, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Petersen, G.; Seberg, O.; Yde, M.; Berthelsen, K. Phylogenetic relationships of Triticum and Aegilops and evidence for the origin of the A, B, and D genomes of common wheat (Triticum aestivum). Mol. Phylogenetics Evol. 2006, 39, 70–82. [Google Scholar] [CrossRef]
- Bernhardt, N. Taxonomic treatments of Triticeae and the wheat genus Triticum. In Alien Introgression in Wheat; Molnár-Láng, M., Ceoloni, C., Doležel, J., Eds.; Springer: Cham, Switzerland, 2015; pp. 1–19. [Google Scholar]
- Kellogg, E.A. Flowering plants. In Monocots; Springer International Publishing: Cham, Switzerland, 2015; Volume 10, pp. 973–978. [Google Scholar]
- Soltis, D.E.; Soltis, P.S. Polyploidy: Recurrent formation and genome evolution. Trends Ecol. Evol. 1999, 14, 348–352. [Google Scholar] [CrossRef]
- Jakob, S.S.; Blattner, F.R. Two extinct diploid progenitors were involved in allopolyploid formation in the Hordeum murinum (Poaceae: Triticeae) taxon complex. Mol. Phylogenetics Evol. 2010, 55, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Brassac, J.; Blattner, F.R. Species-level phylogeny and polyploid relationships in Hordeum (Poaceae) inferred by next-generation sequencing and in silico cloning of multiple nuclear loci. Syst. Biol. 2015, 64, 792–808. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, E.A. Comments on genomic genera in the Triticeae (Poaceae). Am. J. Bot. 1989, 76, 796–805. [Google Scholar] [CrossRef]
- Seberg, O.; Frederiksen, S. A phylogenetic analysis of the monogenomic Triticeae (Poaceae) based on morphology. Bot. J. Linn. Soc. 2001, 136, 75–97. [Google Scholar] [CrossRef]
- Dvorak, J.; Akhunov, E.D. Tempos of gene locus deletions and duplications and their relationship to recombination rate during diploid and polyploid evolution in the Aegilops-Triticum alliance. Genetics 2005, 171, 323–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Baidouri, M.; Murat, F.; Veyssiere, M.; Molinier, M.; Flores, R.; Burlot, L.; Alaux, M.; Quesneville, H.; Pont, C.; Salse, J. Reconciling the evolutionary origin of bread wheat (Triticum aestivum). New Phytol. 2017, 213, 1477–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kilian, B.; Mammen, K.; Millet, E.; Sharma, R.; Graner, A.; Salamini, F.; Hammer, K.; Özkan, H. Aegilops. In Wild Crop Relatives: Genomic and Breeding Resources; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 1–76. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Size (bp) | GC (%) | LSC (bp) | SSC (bp) | IR (bp) | PCD (bp) | tRNA (bp) | rRNA (bp) | Gene (n) | PCD (n) | tRNA (n) | Accesion No. | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| T. sphaerococcum | 134,531 | 38.3 | 80,342 | 12,791 | 20,699 | 59,538 | 3004 | 9192 | 131 | 84 | 39 | MZ230675 |
| T. turgidum subsp. durum | 134,015 | 38.3 | 79,817 | 12,788 | 20,701 | 59,538 | 3004 | 9192 | 131 | 84 | 39 | MZ230674 |
| T. aestivum (NC_002762) | 134,545 | 38.3 | 80,348 | 12,790 | 20,703 | 59,946 | 3264 | 9191 | 133 | 83 | 42 | NC002762 |
| T. aestivum spleta PI384000 | 135,919 | 38.3 | 56,296 | 13,144 | 21,541 | 58,701 | 2469 | 9192 | 123 | 82 | 33 | KJ614403 |
| T. aestivum (KJ592713) | 133,873 | 38.3 | 56,228 | 12,791 | 20,573 | 59,664 | 2943 | 9192 | 131 | 84 | 39 | KJ592713 |
| T. aestivum cultivar Chinese Spring TA3008 | 135,835 | 38.3 | 56,254 | 12,792 | 21,541 | 58,683 | 2469 | 9192 | 123 | 82 | 33 | KJ614396 |
| T. macha | 135,899 | 38.3 | 56,298 | 12,791 | 21,522 | 54,723 | 2480 | 9190 | 112 | 72 | 32 | NC_025955 |
| T. monococcum subsp. monococcum | 136,886 | 38.3 | 56,492 | 12,806 | 21,547 | 54,723 | 2480 | 9192 | 112 | 72 | 32 | LC005977 |
| T. timopheevii cultivar Tim01 | 136,157 | 38.3 | 56,550 | 12,795 | 21,553 | 58,686 | 2469 | 9192 | 123 | 82 | 33 | NC024764 |
| T. timopheevii cultivar TA0941 | 136,074 | 38.3 | 56,474 | 12,789 | 21,553 | 58,686 | 2469 | 9192 | 123 | 82 | 33 | KJ614407 |
| T. timopheevii cultivar TA944 | 136,124 | 38.3 | 56,523 | 12,789 | 21,553 | 58,686 | 2469 | 9192 | 123 | 82 | 33 | KJ614409 |
| T. timopheevii cultivar TA1485 | 136,119 | 38.3 | 56,518 | 12,789 | 21,553 | 58,686 | 2469 | 9192 | 123 | 82 | 33 | KJ614408 |
| T. turgidum cultivar TA0060 | 135,926 | 38.3 | 56,284 | 12,792 | 21,541 | 58,701 | 2469 | 9192 | 123 | 82 | 33 | KJ614401 |
| T. turgidum cultivar TA0073 | 135,865 | 38.3 | 56,266 | 12,792 | 21,541 | 58,683 | 2469 | 9192 | 123 | 82 | 33 | KJ614400 |
| T. turgidum cultivar TA1133 | 135,889 | 38.3 | 56,287 | 12,793 | 21,541 | 58,701 | 2469 | 9192 | 123 | 82 | 33 | KJ614402 |
| T. turgidum cultivar TA2801 | 135,835 | 38.3 | 56,254 | 12,792 | 21,541 | 85,683 | 2469 | 9192 | 123 | 82 | 33 | KJ614399 |
| T. turgidum cultivar TA2836 | 135,835 | 38.3 | 56,254 | 12,792 | 21,541 | 58,683 | 2469 | 9192 | 123 | 82 | 33 | NC_024814 |
| T. turgidum cultivar PI520121 | 135,836 | 38.3 | 56,255 | 12,792 | 21,541 | 58,683 | 2469 | 9192 | 123 | 82 | 33 | KJ614398 |
| T. turgidum subsp. durum cultivar Langdon | 135,948 | 38.3 | 56,293 | 15,992 | 17,066 | 61,746 | 2913 | 9192 | 136 | 89 | 39 | KM352501 |
| T. urartu cultivar PI428335 | 136,865 | 38.3 | 56,454 | 12,824 | 21,547 | 58,683 | 2469 | 9192 | 123 | 82 | 33 | KJ614411 |
| T. zhukovskyi isolate CN036212 | 136,028 | 38.3 | 80,257 | 12,790 | 21,495 | 59,208 | 2871 | 9061 | 128 | 82 | 38 | NC_046698 |
| Category of Genes | Group of Genes | Name of Genes |
|---|---|---|
| Genes for photosynthesis | Subunits of ATP synthase | atpA, atpB, atpE, atpF, atpH, atpI |
| Subunits of photosystem II | psbA, psbB, psbC, psbD, psbE, psbF, psbI, psbJ, psbK, psbL, psbM, psbN, psbT, psbZ, ycf3 | |
| Subunits of NADH-dehydrogenase | ndhA, ndhB*, ndhC, ndhD, ndhE, ndhF, ndhG, ndhH, ndhI, ndhJ, ndhK | |
| Subunits of cytochrome b/f complex | petA, petB, petD, petG, petL, petN | |
| Subunits of photosystem I | psaA, psaB, psaC, psaI, psaJ | |
| Subunit of rubisco | rbcL | |
| Self-replication | Large subunit of ribosome | rpl14, rpl16, rpl2*, rpl20, rpl22, rpl23*, rpl32, rpl33, rpl36 |
| DNA dependent RNA polymerase | rpoA, rpoB, rpoC1, rpoC2 | |
| Small subunit of ribosome | rps11, rps12*, rps14, rps15*, rps16, rps18, rps19*, rps2, rps3, rps3, rps4, rps7*, rps8 | |
| Other genes | c-type cytochrom synthesis gene | ccsA |
| Envelop membrane protein | cemA | |
| Protease | clpP | |
| Translational initiation factor | infA | |
| Maturase | matK | |
| Unknown | Conserved open reading frames | ycf4 |
| Gene | Strand | ExonI | IntronI | ExonII | IntronII | ExonIII | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| T.S | T.T | T.S | T.T | T.S | T.T | T.S | T.T | T.S | T.T | ||
| trnK-UUU | − | 38 | 38 | 2486 | 2490 | 36 | 36 | ||||
| rps16 | − | 40 | 40 | 843 | 845 | 218 | 218 | ||||
| trnS-CGA | − | 31 | 31 | 658 | 658 | 62 | 62 | ||||
| atpF | + | 145 | 145 | 825 | 825 | 407 | 407 | ||||
| ycf3 | − | 124 | 124 | 756 | 756 | 230 | 230 | 731 | 731 | 159 | 159 |
| rps12 | + | 114 | 114 | 223 | 223 | 540 | 540 | 32 | 32 | ||
| trnL-UAA | + | 36 | 36 | 588 | 588 | 51 | 51 | ||||
| trnV-UAC | − | 38 | 38 | 581 | 581 | 53 | 53 | ||||
| petB | + | 6 | 6 | 748 | 748 | 642 | 642 | ||||
| petD | + | 8 | 8 | 749 | 749 | 475 | 475 | ||||
| rpl16 | − | 9 | 9 | 1043 | 1043 | 402 | 402 | ||||
| rpl2 * | − | 385 | 388 | 663 | 663 | 431 | 431 | ||||
| ndhB * | − | 775 | 775 | 712 | 712 | 758 | 758 | ||||
| trnT-CGU | + | 32 | 32 | 787 | 787 | 59 | 59 | ||||
| trnA-UGC | + | 37 | 37 | 805 | 805 | 36 | 36 | ||||
| ndhA | − | 550 | 550 | 1036 | 1036 | 539 | 539 | ||||
| trnA-UGC | − | 37 | 37 | 805 | 805 | 36 | 36 | ||||
| trnT-CGU | − | 32 | 32 | 787 | 787 | 59 | 59 | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lubna; Asaf, S.; Jan, R.; Khan, A.L.; Ahmad, W.; Asif, S.; Al-Harrasi, A.; Kim, K.-M.; Lee, I.-J. The Plastome Sequences of Triticum sphaerococcum (ABD) and Triticum turgidum subsp. durum (AB) Exhibit Evolutionary Changes, Structural Characterization, Comparative Analysis, Phylogenomics and Time Divergence. Int. J. Mol. Sci. 2022, 23, 2783. https://doi.org/10.3390/ijms23052783
Lubna, Asaf S, Jan R, Khan AL, Ahmad W, Asif S, Al-Harrasi A, Kim K-M, Lee I-J. The Plastome Sequences of Triticum sphaerococcum (ABD) and Triticum turgidum subsp. durum (AB) Exhibit Evolutionary Changes, Structural Characterization, Comparative Analysis, Phylogenomics and Time Divergence. International Journal of Molecular Sciences. 2022; 23(5):2783. https://doi.org/10.3390/ijms23052783
Chicago/Turabian StyleLubna, Sajjad Asaf, Rahmatullah Jan, Abdul Latif Khan, Waqar Ahmad, Saleem Asif, Ahmed Al-Harrasi, Kyung-Min Kim, and In-Jung Lee. 2022. "The Plastome Sequences of Triticum sphaerococcum (ABD) and Triticum turgidum subsp. durum (AB) Exhibit Evolutionary Changes, Structural Characterization, Comparative Analysis, Phylogenomics and Time Divergence" International Journal of Molecular Sciences 23, no. 5: 2783. https://doi.org/10.3390/ijms23052783