
Antiviral Activity and Crystal Structures of HIV-1 gp120 Antagonists

 ,
,
Abstract
:1. Introduction
2. Results & Discussion
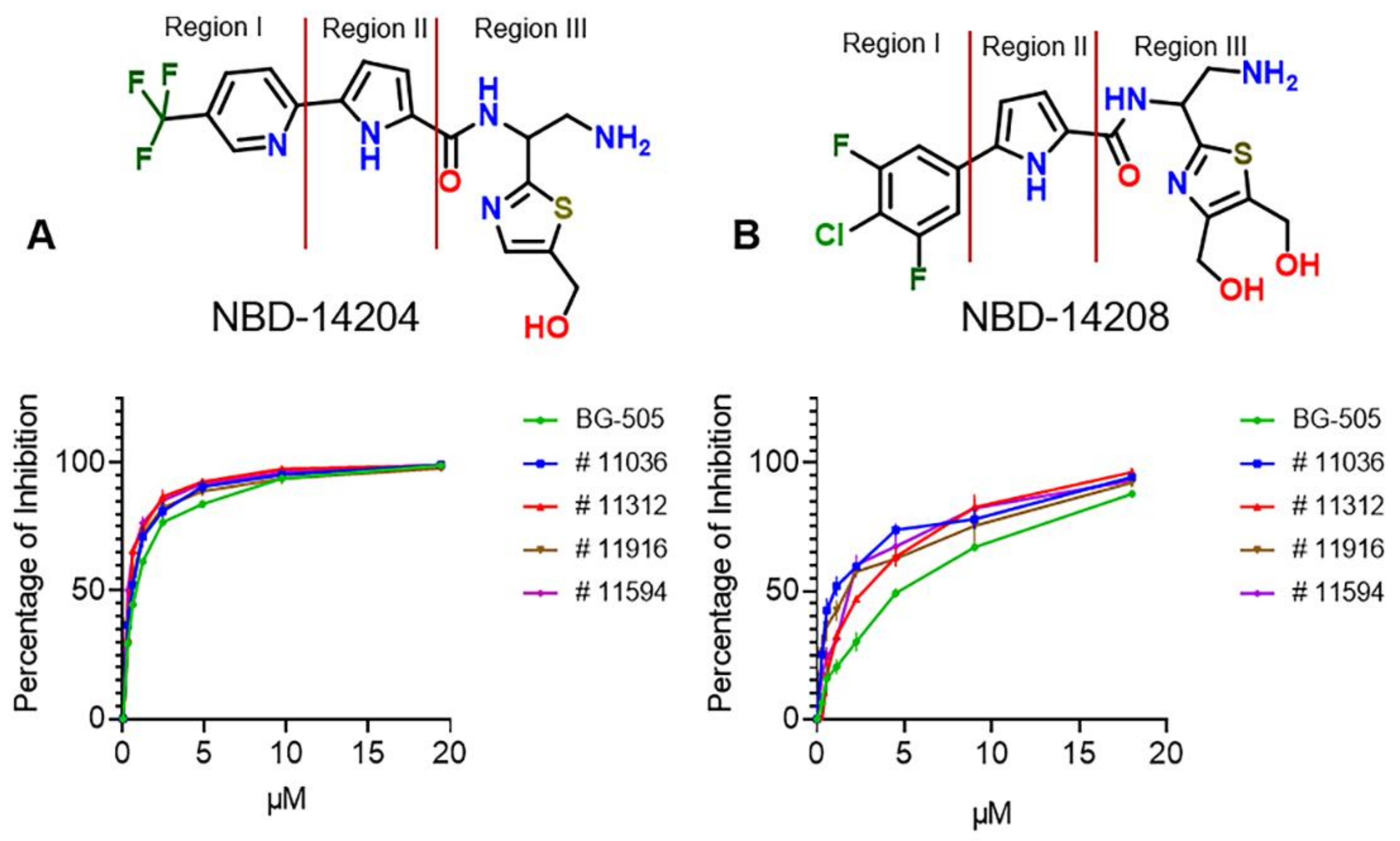
2.1. Evaluation of the Antiviral Activity of NBD-14204 and NBD-14208 against Env-Pseudotyped Clinical Isolates
2.2. S375Y and S375H Mutants Are Not Resistant to NBD-14204 and NBD-14208
2.3. NBD Compounds Showed Activity against HIV-1 Reverse Transcriptase (RT)
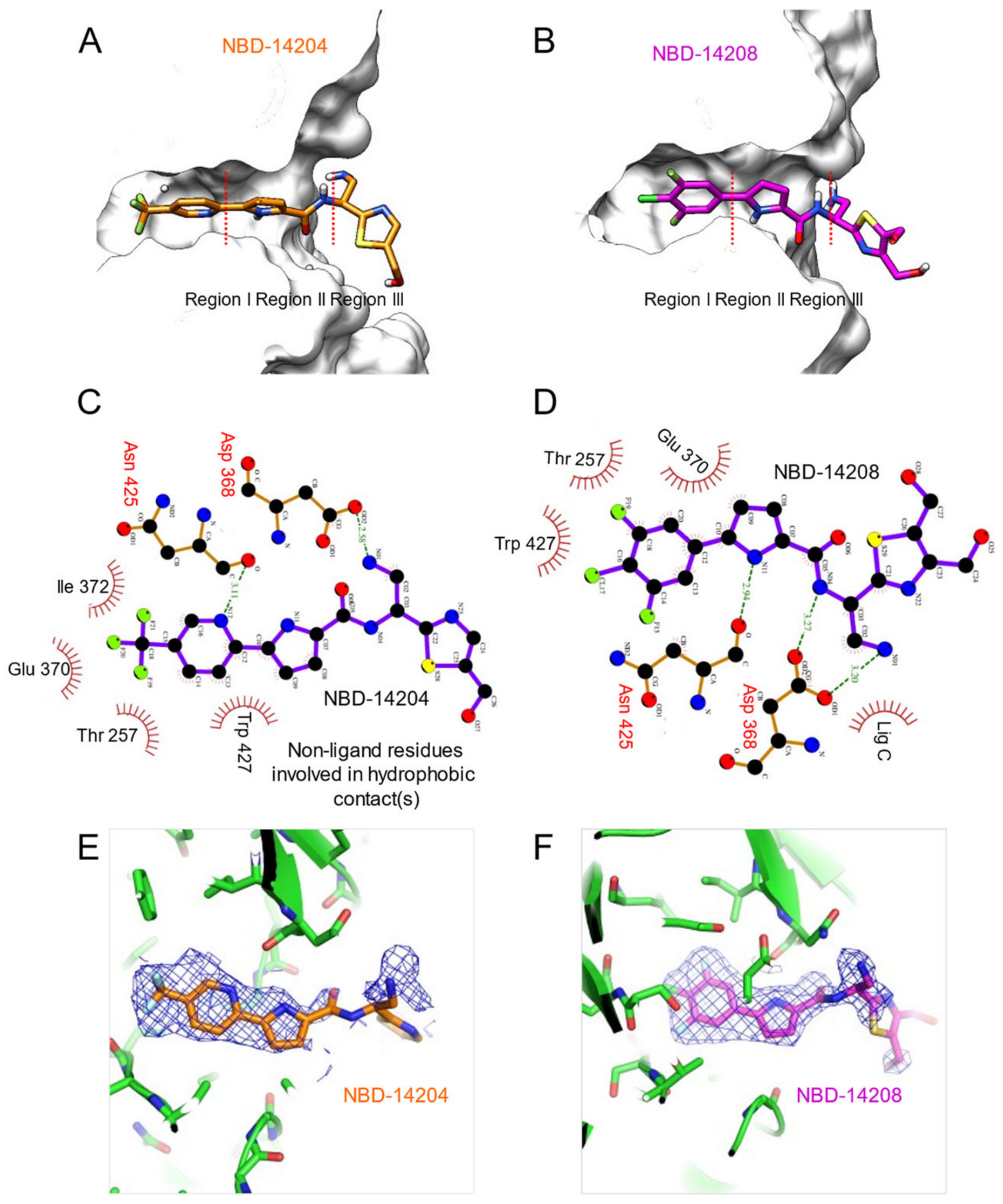
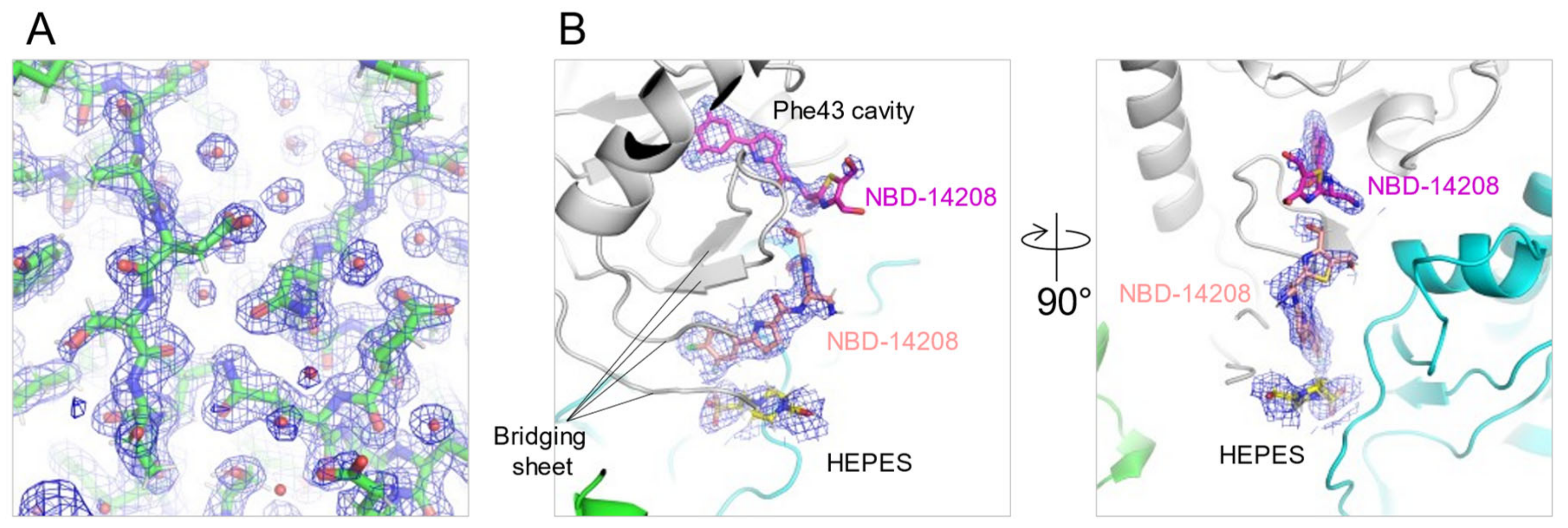
2.4. Crystal Structures of NBD-14204 and NBD-14208-Bound HIV-1 gp120 Core
3. Materials and Methods
3.1. Cells and Viruses
3.2. Pseudovirus Preparation
3.3. Measurement of Antiviral Activity
3.4. Evaluation of Cytotoxicity
3.5. Reverse Transcriptase Inhibition Assay
3.6. Crystallization, Data Collection, and Structural Refinement
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, T.; Zhang, Z.; Wallace, O.B.; Deshpande, M.; Fang, H.; Yang, Z.; Zadjura, L.M.; Tweedie, D.L.; Huang, S.; Zhao, F.; et al. Discovery of 4-benzoyl-1-[(4-methoxy-1H- pyrrolo[2,3-b]pyridin-3-yl)oxoacetyl]-2- (R)-methylpiperazine (BMS-378806): A novel HIV-1 attachment inhibitor that interferes with CD4-gp120 interactions. J. Med. Chem. 2003, 46, 4236–4239. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Ueda, Y.; Zhang, Z.; Yin, Z.; Matiskella, J.; Pearce, B.C.; Yang, Z.; Zheng, M.; Parker, D.D.; Yamanaka, G.A.; et al. Discovery of the Human Immunodeficiency Virus Type 1 (HIV-1) Attachment Inhibitor Temsavir and Its Phosphonooxymethyl Prodrug Fostemsavir. J. Med. Chem. 2018, 61, 6308–6327. [Google Scholar] [CrossRef] [PubMed]
- Curreli, F.; Choudhury, S.; Pyatkin, I.; Zagorodnikov, V.P.; Bulay, A.K.; Altieri, A.; Kwon, Y.D.; Kwong, P.D.; Debnath, A.K. Design, synthesis and antiviral activity of entry inhibitors that target the CD4-binding site of HIV-1. J. Med. Chem. 2012, 55, 4764–4775. [Google Scholar] [CrossRef] [PubMed]
- Curreli, F.; Kwon, Y.D.; Zhang, H.; Yang, Y.; Scacalossi, D.; Kwong, P.D.; Debnath, A.K. Binding mode characterization of NBD series CD4-mimetic HIV-1 entry inhibitors by X-ray structure and resistance study. Antimicrob. Agents Chemother. 2014, 58, 5478–5491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Curreli, F.; Haque, K.; Xie, L.; Qiu, Q.; Xu, J.; Yong, W.; Tong, X.; Debnath, A.K. Synthesis, antiviral activity and resistance of a novel small molecule HIV-1 entry inhibitor. Bioorg. Med. Chem. 2015, 23, 7618–7628. [Google Scholar] [CrossRef] [Green Version]
- Curreli, F.; Kwon, Y.D.; Zhang, H.; Scacalossi, D.; Belov, D.S.; Tikhonov, A.A.; Andreev, I.A.; Altieri, A.; Kurkin, A.V.; Kwong, P.D.; et al. Structure-based design of a small molecule CD4-antagonist with broad spectrum anti-HIV-1 activity. J. Med. Chem. 2015, 58, 6909–6927. [Google Scholar] [CrossRef] [Green Version]
- Curreli, F.; Belov, D.S.; Ramesh, R.R.; Patel, N.; Altieri, A.; Kurkin, A.V.; Debnath, A.K. Design, synthesis and evaluation of small molecule CD4-mimics as entry inhibitors possessing broad spectrum anti-HIV-1 activity. Bioorg. Med. Chem. 2016, 24, 5988–6003. [Google Scholar] [CrossRef] [Green Version]
- Belov, D.S.; Ivanov, V.N.; Curreli, F.; Kurkin, A.V.; Altieri, A.; Debnath, A.K. Synthesis of 5-arylpyrrole-2-carboxylic acids as key intermediates for NBD series HIV-1 entry inhibitors. Synthesis 2017, 49, 3692–3699. [Google Scholar] [CrossRef] [Green Version]
- Curreli, F.; Kwon, Y.D.; Belov, D.S.; Ramesh, R.R.; Kurkin, A.V.; Altieri, A.; Kwong, P.D.; Debnath, A.K. Synthesis, antiviral potency, in vitro ADMET and X-ray structure of potent CD4-mimics as entry inhibitors that target the Phe43 cavity of HIV-1 gp120. J. Med. Chem. 2017, 60, 3124–3153. [Google Scholar] [CrossRef] [PubMed]
- Belov, D.S.; Curreli, F.; Kurkin, A.V.; Altieri, A.; Debnath, A.K. Guanidine-containing phenyl-pyrrole compounds as probes for generating HIV nntry inhibitors targeted to gp120. ChemistrySelect 2018, 3, 6450–6453. [Google Scholar] [CrossRef]
- Curreli, F.; Belov, D.S.; Ahmed, S.; Ramesh, R.R.; Kurkin, A.V.; Altieri, A.; Debnath, A.K. Synthesis, antiviral activity, and structure-activity relationship of 1,3-benzodioxolyl pyrrole-based entry inhibitors targeting the Phe43 cavity in HIV-1 gp120. ChemMedChem 2018, 13, 2332–2348. [Google Scholar] [CrossRef] [PubMed]
- Curreli, F.; Belov, D.S.; Kwon, Y.D.; Ramesh, R.; Furimsky, A.M.; O’Loughlin, K.; Byrge, P.C.; Iyer, L.V.; Mirsalis, J.C.; Kurkin, A.V.; et al. Structure-based lead optimization to improve antiviral potency and ADMET properties of phenyl-1H-pyrrole-carboxamide entry inhibitors targeted to HIV-1 gp120. Eur. J. Med. Chem. 2018, 154, 367–391. [Google Scholar] [CrossRef] [PubMed]
- Curreli, F.; Ahmed, S.; Benedict Victor, S.M.; Iusupov, I.R.; Belov, D.S.; Markov, P.O.; Kurkin, A.V.; Altieri, A.; Debnath, A.K. Preclinical Optimization of gp120 Entry Antagonists as anti-HIV-1 Agents with Improved Cytotoxicity and ADME Properties through Rational Design, Synthesis, and Antiviral Evaluation. J. Med. Chem. 2020, 63, 1724–1749. [Google Scholar] [CrossRef] [PubMed]
- Curreli, F.; Ahmed, S.; Benedict Victor, S.M.; Iusupov, I.R.; Spiridonov, E.A.; Belov, D.S.; Altieri, A.; Kurkin, A.V.; Debnath, A.K. Design, synthesis, and antiviral activity of a series of CD4-mimetic small-molecule HIV-1 entry inhibitors. Bioorg. Med. Chem. 2021, 32, 116000. [Google Scholar] [CrossRef] [PubMed]
- Losada, N.; Ruiz, F.X.; Curreli, F.; Gruber, K.; Pilch, A.; Das, K.; Debnath, A.K.; Arnold, E. HIV-1 gp120 Antagonists Also Inhibit HIV-1 Reverse Transcriptase by Bridging the NNRTI and NRTI Sites. J. Med. Chem. 2021, 64, 16530–16540. [Google Scholar] [CrossRef]
- Stoddart, C.A.; Curreli, F.; Horrigan, S.; Altieri, A.; Kurkin, A.V.; Debnath, A.K. Comparative Pharmacokinetics of a Dual Inhibitor of HIV-1, NBD-14189, in Rats and Dogs with a Proof-of-Concept Evaluation of Antiviral Potency in SCID-hu Mouse Model. Viruses 2022, 14, 2268. [Google Scholar] [CrossRef]
- Zhou, N.; Nowicka-Sans, B.; Zhang, S.; Fan, L.; Fang, J.; Fang, H.; Gong, Y.F.; Eggers, B.; Langley, D.R.; Wang, T.; et al. In vivo patterns of resistance to the HIV attachment inhibitor BMS-488043. Antimicrob. Agents Chemother. 2011, 55, 729–737. [Google Scholar] [CrossRef] [Green Version]
- Lataillade, M.; Zhou, N.; Joshi, S.R.; Lee, S.; Stock, D.A.; Hanna, G.J.; Krystal, M.; AI438011 Study Team. Viral Drug Resistance Through 48 Weeks, in a Phase 2b, Randomized, Controlled Trial of the HIV-1 Attachment Inhibitor Prodrug, Fostemsavir. J. Acquir. Immune Defic. Syndr. 2018, 77, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Himmel, D.M.; Sarafianos, S.G.; Dharmasena, S.; Hossain, M.M.; McCoy-Simandle, K.; Ilina, T.; Clark, A.D., Jr.; Knight, J.L.; Julias, J.G.; Clark, P.K.; et al. HIV-1 reverse transcriptase structure with RNase H inhibitor dihydroxy benzoyl naphthyl hydrazone bound at a novel site. ACS Chem. Biol. 2006, 1, 702–712. [Google Scholar] [CrossRef] [Green Version]
- Lai, M.T.; Tawa, P.; Auger, A.; Wang, D.; Su, H.P.; Yan, Y.; Hazuda, D.J.; Miller, M.D.; Asante-Appiah, E.; Melnyk, R.A. Identification of novel bifunctional HIV-1 reverse transcriptase inhibitors. J. Antimicrob. Chemother. 2018, 73, 109–117. [Google Scholar] [CrossRef]
- Lalonde, J.M.; Kwon, Y.D.; Jones, D.M.; Sun, A.W.; Courter, J.R.; Soeta, T.; Kobayashi, T.; Princiotto, A.M.; Wu, X.; Schon, A.; et al. Structure-based design, synthesis, and characterization of dual hotspot small-molecule HIV-1 entry inhibitors. J. Med. Chem. 2012, 55, 4382–4396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, Y.D.; Lalonde, J.M.; Yang, Y.; Elban, M.A.; Sugawara, A.; Courter, J.R.; Jones, D.M.; Smith, A.B., III; Debnath, A.K.; Kwong, P.D. Crystal structures of HIV-1 gp120 envelope glycoprotein in complex with NBD analogues that target the CD4-binding site. PLoS One 2014, 9, e85940. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.D.; Finzi, A.; Wu, X.; Dogo-Isonagie, C.; Lee, L.K.; Moore, L.R.; Schmidt, S.D.; Stuckey, J.; Yang, Y.; Zhou, T.; et al. Unliganded HIV-1 gp120 core structures assume the CD4-bound conformation with regulation by quaternary interactions and variable loops. Proc. Natl. Acad. Sci. USA 2012, 109, 5663–5668. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Decker, J.M.; Liu, H.; Zhang, Z.; Arani, R.B.; Kilby, J.M.; Saag, M.S.; Wu, X.; Shaw, G.M.; Kappes, J.C. Emergence of resistant human immunodeficiency virus type 1 in patients receiving fusion inhibitor (T-20) monotherapy. Antimicrob. Agents Chemother. 2002, 46, 1896–1905. [Google Scholar] [CrossRef] [Green Version]
- Page, K.A.; Landau, N.R.; Littman, D.R. Construction and use of a human immunodeficiency virus vector for analysis of virus infectivity. J. Virol. 1990, 64, 5270–5276. [Google Scholar] [CrossRef] [Green Version]
- Blish, C.A.; Jalalian-Lechak, Z.; Rainwater, S.; Nguyen, M.A.; Dogan, O.C.; Overbaugh, J. Cross-subtype neutralization sensitivity despite monoclonal antibody resistance among early subtype A, C, and D envelope variants of human immunodeficiency virus type 1. J. Virol. 2009, 83, 7783–7788. [Google Scholar] [CrossRef] [Green Version]
- Long, E.M.; Rainwater, S.M.; Lavreys, L.; Mandaliya, K.; Overbaugh, J. HIV type 1 variants transmitted to women in Kenya require the CCR5 coreceptor for entry, regardless of the genetic complexity of the infecting virus. AIDS Res. Hum. Retrov. 2002, 18, 567–576. [Google Scholar] [CrossRef]
- Wu, X.; Parast, A.B.; Richardson, B.A.; Nduati, R.; John-Stewart, G.; Mbori-Ngacha, D.; Rainwater, S.M.J.; Overbaugh, J. Neutralization Escape Variants of Human Immunodeficiency Virus Type 1 Are Transmitted from Mother to Infant. J. Virol. 2006, 80, 835–844. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.S.; Lapedes, A.; Tang, H.; Gnanakaran, S.; Daniels, M.G.; Zhang, M.; Bhattacharya, T.; Li, M.; Polonis, V.R.; McCutchan, F.E.; et al. Highly complex neutralization determinants on a monophyletic lineage of newly transmitted subtype C HIV-1 Env clones from India. Virology 2009, 385, 505–520. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Gao, F.; Mascola, J.R.; Stamatatos, L.; Polonis, V.R.; Koutsoukos, M.; Voss, G.; Goepfert, P.; Gilbert, P.; Greene, K.M.; et al. Human immunodeficiency virus type 1 env clones from acute and early subtype B infections for standardized assessments of vaccine-elicited neutralizing antibodies. J. Virol. 2005, 79, 10108–10125. [Google Scholar] [CrossRef]
- Li, M.; Salazar-Gonzalez, J.F.; Derdeyn, C.A.; Morris, L.; Williamson, C.; Robinson, J.E.; Decker, J.M.; Li, Y.; Salazar, M.G.; Polonis, V.R.; et al. Genetic and neutralization properties of subtype C human immunodeficiency virus type 1 molecular env clones from acute and early heterosexually acquired infections in Southern Africa. J. Virol. 2006, 80, 11776–11790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keele, B.F.; Giorgi, E.E.; Salazar-Gonzalez, J.F.; Decker, J.M.; Pham, K.T.; Salazar, M.G.; Sun, C.; Grayson, T.; Wang, S.; Li, H.; et al. Identification and characterization of transmitted and early founder virus envelopes in primary HIV-1 infection. Proc. Natl. Acad. Sci. USA 2008, 105, 7552–7557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derdeyn, C.A.; Decker, J.M.; Bibollet-Ruche, F.; Mokili, J.L.; Muldoon, M.; Denham, S.A.; Heil, M.L.; Kasolo, F.; Musonda, R.; Hahn, B.H.; et al. Envelope-constrained neutralization-sensitive HIV-1 after heterosexual transmission. Science 2004, 303, 2019–2022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, X.; Decker, J.M.; Wang, S.; Hui, H.; Kappes, J.C.; Wu, X.; Salazar-Gonzalez, J.F.; Salazar, M.G.; Kilby, J.M.; Saag, M.S.; et al. Antibody neutralization and escape by HIV-1. Nature 2003, 422, 307–312. [Google Scholar] [CrossRef]
- Zhao, Q.; Ma, L.; Jiang, S.; Lu, H.; Liu, S.; He, Y.; Strick, N.; Neamati, N.; Debnath, A.K. Identification of N-phenyl-N’-(2,2,6,6-tetramethyl-piperidin-4-yl)-oxalamides as a new class of HIV-1 entry inhibitors that prevent gp120 binding to CD4. Virology 2005, 339, 213–225. [Google Scholar] [CrossRef] [PubMed]
- Otwinowski, Z.; Minor, W. Processing of X-ray Diffraction Data Collected in Oscillation Method. In Methods Enzymol; Otwinowski, Z., Minor, W., Eds.; Elsevier: Amsterdam, The Netherlands, 1997; Volume 276, pp. 307–326. [Google Scholar]
- McCoy, A.J.; Grosse-Kunstleve, R.W.; Adams, P.D.; Winn, M.D.; Storoni, L.C.; Read, R.J. Phaser crystallographic software. J. Appl. Crystallogr. 2007, 40, 658–674. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. Sect. D 2004, 60, 2126–2132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkoczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D 2010, 66, 213–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, A.C.; Laskowski, R.A.; Thornton, J.M. LIGPLOT: A program to generate schematic diagrams of protein-ligand interactions. Protein Eng. 1995, 8, 127–134. [Google Scholar] [CrossRef]
- Schrodinger, L.D.W. Incentive PyMOL Software Packag. Available online: http://www.pymol.org/pymol (accessed on 22 November 2022).
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| IC50 (µM) a | ||||
|---|---|---|---|---|
| Subtype | ARP # | ENVs | NBD-14204 | NBD-14208 |
| A | 11895 | QH209.14M.ENV.A2 | 0.27 ± 0.1 | 4.4 ± 0.1 |
| A | 11888 | QB726.70M.ENV.C4 | 0.4 ± 0.2 | 2.87 ± 0.2 |
| A | BG505-T332N | 0.8 ± 0.04 | 4.7 ± 0.4 | |
| A | KNH1144 | 0.48 ± 0.05 | 3.6 ± 0.1 | |
| AD | 11901 | QA790.204I.ENV.A4 | 0.24 ± 0.01 | 3.4 ± 1 |
| AE | 11603 | CRF01_AE clone 269 | 0.53 ± 0.04 | 5.7 ± 1.3 |
| AG | 11594 | CRF02_AG clone 250 | 0.32 ± 0.01 | 1.87 ± 0.08 |
| AG | 11605 | CRF02_AG Clone 278 | 0.53 ± 0.04 | 4.4 ± 0.16 |
| B | B41 | 0.9 ± 0.1 | 4.2 ± 0.1 | |
| B | 11018 | QH0692, clone 42 | 0.48 ± 0.05 | 3.6 ± 0.1 |
| B | 11022 | PVO, clone 4 | 0.5 ± 0.04 | 0.66 ± 0.03 |
| B | 11024 | AC10.0.29 (SVPB13) | 0.33 ± 0.03 | 2.3 ± 0.1 |
| B | 11036 | pRHPA4259 clone7 (SVPB14) | 0.56 ± 0.05 | 1.1 ± 0.2 |
| B | 11037 | pTHRO4156 clone 18 (SVPB15) | 0.29 ± 0.01 | 1.9 ± 0.04 |
| B | 11563 | 1058_11.B11.1550 b | 0.58 ± 0.02 | 3.7 ± 0.09 |
| C | 11307 | Du172, clone 17 | 0.6 ± 0.15 | 3.7 ± 0.3 |
| C | 11308 | Du422, clone 1 | 0.65 ± 0.03 | 3.8 ± 0.2 |
| C | 11312 | ZM249M.PL1, SVPC10 | 0.46 ± 0.05 | 2.8 ± 0.3 |
| C | 11313 | ZM53M.PB12, SVPC11 | 0.7 ± 0.09 | 3.8 ± 0.3 |
| C | 11507 | HIV-25925-2, clone22 | 0.29 ± 0.01 | 1.85 ± 0.5 |
| C | 11908 | QB099.391M.ENV.B1 | 0.3 ± 0.02 | 1.5 ± 0.08 |
| D | 11911 | QA013.70I.ENV.H1 | 0.35 ± 0.1 | 1.7 ± 0.1 |
| D | 11912 | QA013.70I.ENV.M12 | 0.24 ± 0.1 | 3.3 ± 0.5 |
| D | 11916 | QD435.100M.ENV.B5 | 0.54 ± 0.06 | 1.7 ± 0.2 |
| D/A | 11526 | MF535.WOM.ENV.C1 | 0.48 ± 0.07 | 2.8 ± 0.6 |
| Mean ± SEM (µM): Overall (n = 25) SI | 0.47 ± 0.03 310 | 3 ± 0.25 27.5 | ||
| Subtype A (n = 4) | 0.49 ± 0.11 | 3.9 ± 0.4 | ||
| Subtype B (n = 7) | 0.52 ± 0.08 | 2.5 ± 0.5 | ||
| Subtype C (n = 6) | 0.5 ± 0.07 | 2.9 ± 0.4 | ||
| Subtype D (n = 3) | 0.38 ± 0.09 | 2.2 ± 0.5 | ||
| Subtype Rec (n = 5) | 0.39 ± 0.07 | 3.4 ± 0.8 | ||
| Toxicity CC50 (µM) a | 146 ± 3 | 82.5 ± 1.5 | ||
| Inhibitors | RT IC50 (μM) a | HXB2 | Clinical Isolates (CI) | ||
|---|---|---|---|---|---|
| IC50 (μM) | Ratio RT/HXB2 | Mean IC50 (μM) | Ratio RT/CI | ||
| NBD-14204 | 8.3 ± 1.2 | 0.96 ± 0.1 | 8.6 | 0.47 ± 0.03 | 17.7 |
| NBD-14208 | 5 ± 0.5 | 2.3 ± 0.1 | 2.2 | 3 ± 0.25 | 1.7 |
| NBD-556 | >300 | - | - | - | - |
| Nevirapine | 0.2 | - | - | - | - |
| NBD-14204:gp120 | NBD-14208:gp120 | |
|---|---|---|
| Data collection | ||
| Space group | P212121 | P212121 |
| Cell dimensions | ||
| a, b, c (Å) | 65.2, 67.6, 87.9 | 61.2, 66.9, 90.4 |
| α, β, γ (°) | 90, 90, 90 | 90.0, 90, 90 |
| Resolution (Å) | 50–1.90 (1.93–1.90) * | 50–2.09 (2.18–2.09) * |
| Rsym | 0.076 (0.82) | 0.14 (0.56) |
| I/sI | 13.5 (2.4) | 14.5 (1.1) |
| Completeness (%) | 92.2 (74.1) | 94.2 (71.0) |
| Redundancy | 4.3 (3.1) | 6.4 (2.9) |
| Refinement | ||
| Resolution (Å) | 36.4–1.94 | 36.4–2.09 |
| No. reflections | 24,631 | 21,346 |
| Rwork/Rfree | 18.2/21.6 | 24.2/29.7 |
| No. atoms | ||
| Protein | 5308 | 5278 |
| Ligand/ion | 254 | 370 |
| Water | 277 | 49 |
| B-factors (Å2) | ||
| Protein | 26.4 | 59.6 |
| Ligand/ion | 40.8 | 76.8 |
| Water | 32.9 | 59.5 |
| R.m.s. deviations | ||
| Bond lengths (Å) | 0.008 | 0.011 |
| Bond angles (°) | 0.767 | 0.760 |
| Ramachandran plot (%) | ||
| Favored | 97.3 | 96.65 |
| Allowed | 2.70 | 3.35 |
| Outliers | 0.00 | 0.00 |
| PDB ID | 8F9Z | 8FA0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Curreli, F.; Kwon, Y.D.; Nicolau, I.; Burgos, G.; Altieri, A.; Kurkin, A.V.; Verardi, R.; Kwong, P.D.; Debnath, A.K. Antiviral Activity and Crystal Structures of HIV-1 gp120 Antagonists. Int. J. Mol. Sci. 2022, 23, 15999. https://doi.org/10.3390/ijms232415999
Curreli F, Kwon YD, Nicolau I, Burgos G, Altieri A, Kurkin AV, Verardi R, Kwong PD, Debnath AK. Antiviral Activity and Crystal Structures of HIV-1 gp120 Antagonists. International Journal of Molecular Sciences. 2022; 23(24):15999. https://doi.org/10.3390/ijms232415999
Chicago/Turabian StyleCurreli, Francesca, Young D. Kwon, Isabella Nicolau, Giancarla Burgos, Andrea Altieri, Alexander V. Kurkin, Raffaello Verardi, Peter D. Kwong, and Asim K. Debnath. 2022. "Antiviral Activity and Crystal Structures of HIV-1 gp120 Antagonists" International Journal of Molecular Sciences 23, no. 24: 15999. https://doi.org/10.3390/ijms232415999