
Revealing the Changes in Saliva and Serum Proteins of Pigs with Meningitis Caused by Streptococcus Suis: A Proteomic Approach
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
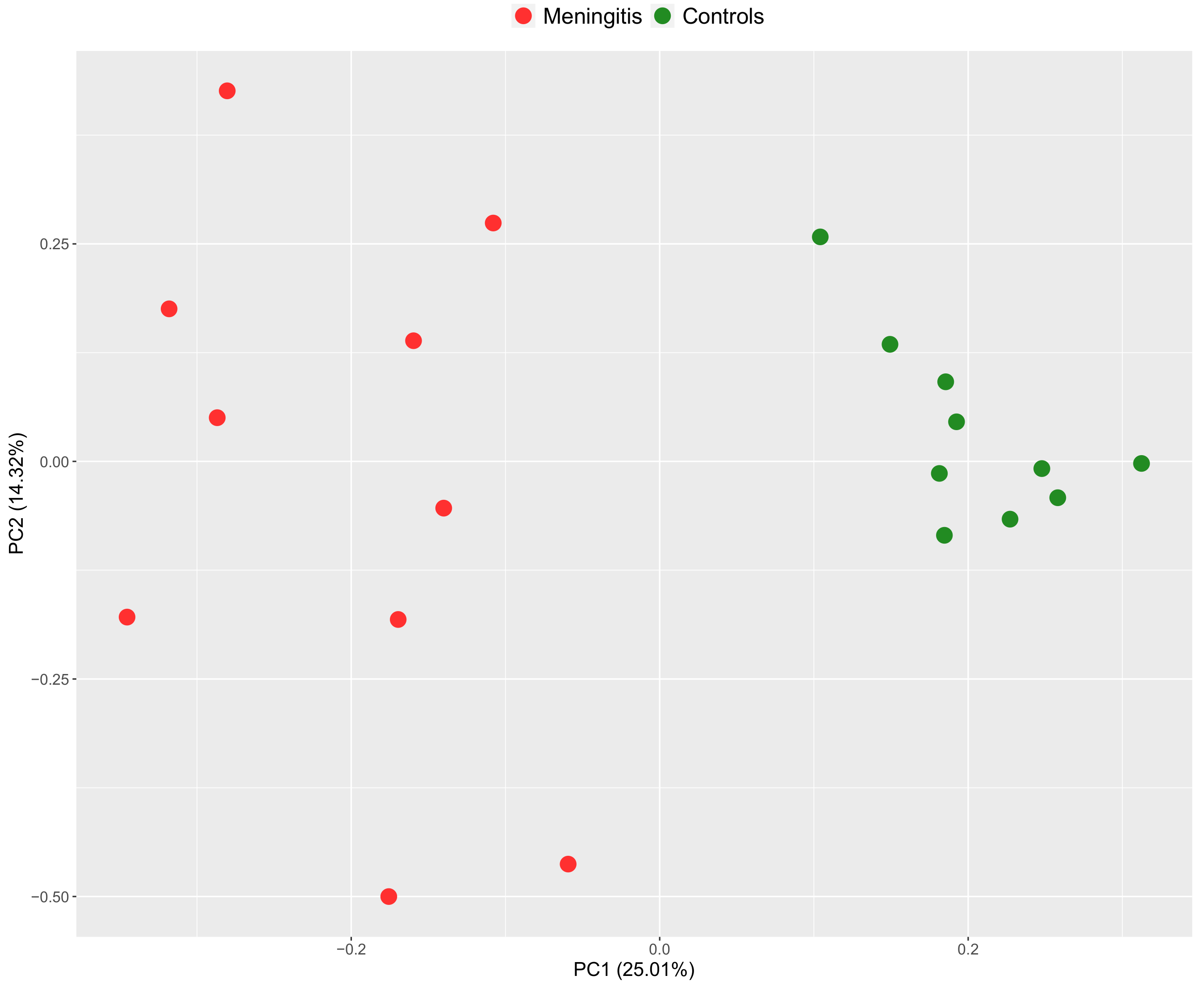
2.1. Salivary Proteomic Profile in Piglets with Meningitis
2.2. Serum Proteomic Profile in Piglets with Meningitis
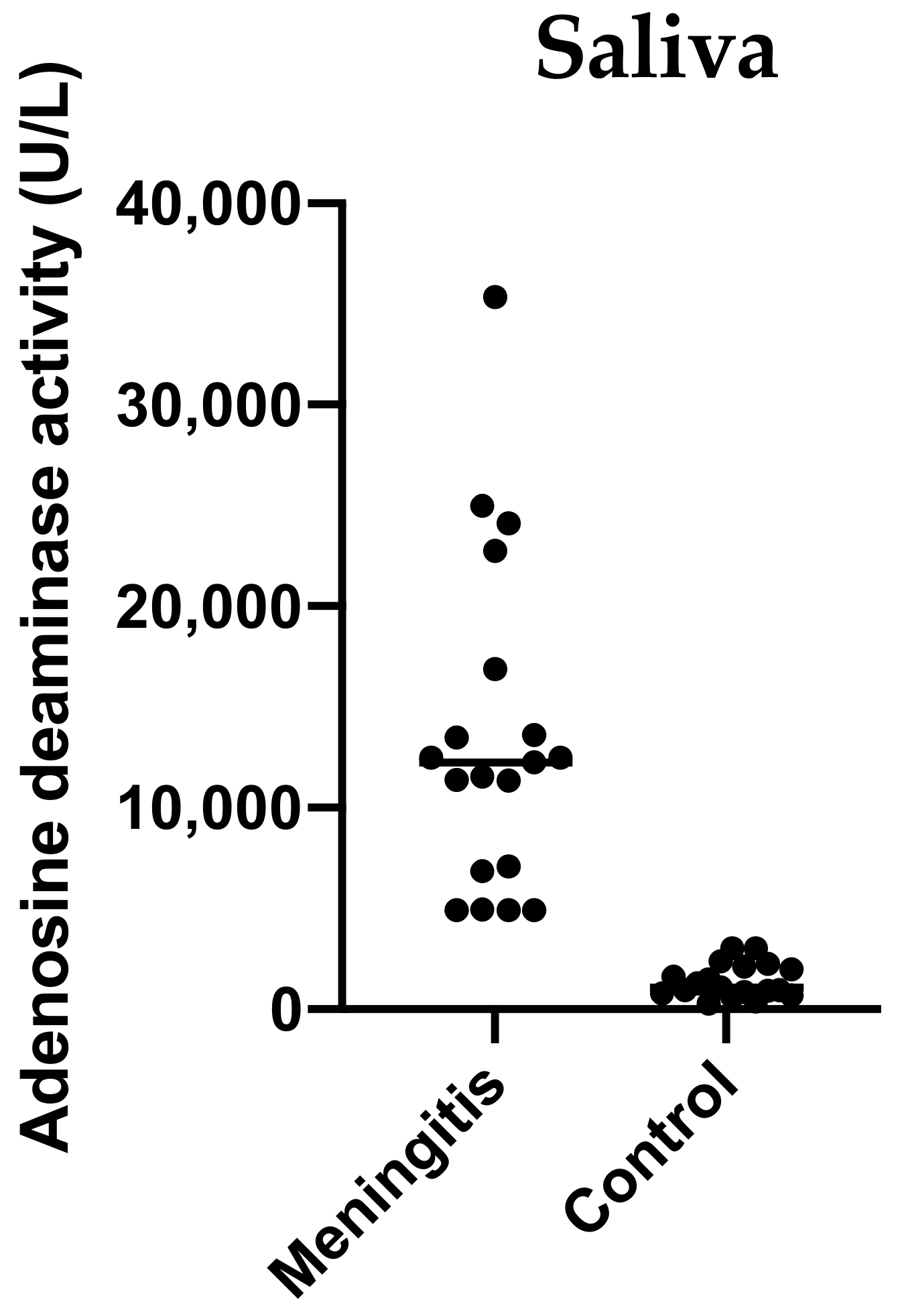
2.3. Validation Study: Adenosine Deaminase (ADA) Activity of Saliva in Pigs with Meningitis
3. Discussion
3.1. Changes in Salivary Proteins in Pigs with Meningitis
3.2. Changes in Serum Proteins in Pigs with Meningitis
3.3. Clinical Implications of Differentially Expressed Salivary Proteins in Pigs with Meningitis
4. Materials and Methods
4.1. Animals
4.2. Saliva and Serum Collection
4.3. Streptococcus suis Isolation and Typification
4.4. Proteomic Analysis
4.5. Statistics and Bioinformatics Analyses
4.6. Validation Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hopkins, D.; Poljak, Z.; Farzan, A.; Friendship, R. Factors Contributing to Mortality during a Streptoccocus Suis Outbreak in Nursery Pigs. Can. Vet. J. 2018, 59, 623–630. [Google Scholar] [PubMed]
- Madsen, L.W.; Svensmark, B.; Elvestad, K.; Aalbaek, B.; Jensen, H.E. Streptococcus Suis Serotype 2 Infection in Pigs: New Diagnostic and Pathogenetic Aspects. J. Comp. Pathol. 2002, 126, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Fittipaldi, N.; Segura, M.; Grenier, D.; Gottschalk, M. Virulence Factors Involved in the Pathogenesis of the Infection Caused by the Swine Pathogen and Zoonotic Agent Streptococcus Suis. Future Microbiol. 2012, 7, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.; Grenier, D. Understanding the Virulence of Streptococcus Suis: A Veterinary, Medical, and Economic Challenge. Med. Mal. Infect. 2018, 48, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Zhang, H.; Wu, Z.; Wang, S.; Cao, M.; Hu, D.; Wang, C. Streptococcus Suis Infection: An Emerging/Reemerging Challenge of Bacterial Infectious Diseases? Virulence 2014, 5, 477–497. [Google Scholar] [CrossRef] [Green Version]
- Wertheim, H.F.L.; Nghia, H.D.T.; Taylor, W.; Schultsz, C. Streptococcus Suis: An Emerging Human Pathogen. Clin. Infect. Dis. 2009, 48, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Aslam, B.; Basit, M.; Nisar, M.A.; Khurshid, M.; Rasool, M.H. Proteomics: Technologies and Their Applications. J. Chromatogr. Sci. 2017, 55, 182–196. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Liu, J.; Wu, T.; Liu, M.; Sun, Q.; Wang, J.; Zhu, R.; Qu, G.; Li, S.; Liu, H.; et al. Proteomics Analysis of Important Molecules in Serum from Meningitic Piglets Caused by Streptococcus Suis Serotype 2. J. Infect. Dev. Ctries. 2020, 14, 502–510. [Google Scholar] [CrossRef]
- Cerón, J.J. Acute Phase Proteins, Saliva and Education in Laboratory Science: An Update and Some Reflections. BMC Vet. Res. 2019, 15, 197. [Google Scholar] [CrossRef]
- Muñoz-Prieto, A.; Contreras-Aguilar, M.D.; Cerón, J.J.; Ayala, I.; Martin-Cuervo, M.; Gonzalez-Sanchez, J.C.; Jacobsen, S.; Kuleš, J.; Beletić, A.; Rubić, I.; et al. Changes in Proteins in Saliva and Serum in Equine Gastric Ulcer Syndrome Using a Proteomic Approach. Animals 2022, 12, 1169. [Google Scholar] [CrossRef]
- Franco-Martínez, L.; Muñoz-Prieto, A.; Contreras-Aguilar, M.D.; Želvytė, R.; Monkevičienė, I.; Horvatić, A.; Kuleš, J.; Mrljak, V.; Cerón, J.J.; Escribano, D. Changes in Saliva Proteins in Cows with Mastitis: A Proteomic Approach. Res. Vet. Sci. 2021, 140, 91–99. [Google Scholar] [CrossRef]
- López-Martínez, M.J.; Cerón, J.J.; Ortín-Bustillo, A.; Escribano, D.; Kuleš, J.; Beletić, A.; Rubić, I.; González-Sánchez, J.C.; Mrljak, V.; Martínez-Subiela, S.; et al. A Proteomic Approach to Elucidate the Changes in Saliva and Serum Proteins of Pigs with Septic and Non-Septic Inflammation. Int. J. Mol. Sci. 2022, 23, 6738. [Google Scholar] [CrossRef]
- Neilson, K.A.; Ali, N.A.; Muralidharan, S.; Mirzaei, M.; Mariani, M.; Assadourian, G.; Lee, A.; van Sluyter, S.C.; Haynes, P.A. Less Label, More Free: Approaches in Label-Free Quantitative Mass Spectrometry. Proteomics 2011, 11, 535–553. [Google Scholar] [CrossRef]
- Lee, H.T.; Sharek, L.; Timothy O’Brien, E.; Urbina, F.L.; Gupton, S.L.; Superfine, R.; Burridge, K.; Campbell, S.L. Vinculin and Metavinculin Exhibit Distinct Effects on Focal Adhesion Properties, Cell Migration, and Mechanotransduction. PLoS ONE 2019, 14, e0221962. [Google Scholar] [CrossRef] [Green Version]
- Belkin, A.M.; Ornatsky, O.I.; Glukhova, M.A.; Koteliansky, V.E. Immunolocalization of Meta-Vinculin in Human Smooth and Cardiac Muscles. J. Cell. Biol. 1988, 107, 545–553. [Google Scholar] [CrossRef]
- Zemljic-Harpf, A.E.; Miller, J.C.; Henderson, S.A.; Wright, A.T.; Manso, A.M.; Elsherif, L.; Dalton, N.D.; Thor, A.K.; Perkins, G.A.; McCulloch, A.D.; et al. Cardiac-Myocyte-Specific Excision of the Vinculin Gene Disrupts Cellular Junctions, Causing Sudden Death or Dilated Cardiomyopathy. Mol. Cell. Biol. 2007, 27, 7522–7537. [Google Scholar] [CrossRef] [Green Version]
- Yin, G.; Zeng, W.; Li, R.; Zeng, M.; Chen, R.; Liu, Y.; Jiang, R.; Wang, Y. Glia Maturation Factor- b Supports Liver Regeneration By Remodeling Actin Network to Enhance STAT3 Proliferative Signals. Cell. Mol. Gastroenterol. Hepatol. 2022, 14, 1123–1145. [Google Scholar] [CrossRef]
- Greenwood, M.D.; Marsden, M.D.; Cowley, C.M.E.; Sahota, V.K.; Buxton, R.S. Exon-Intron Organization of the Human Type 2 Desmocollin Gene (DSC2): Desmocollin Gene Structure Is Closer to “‘Classical’” Cadherins Than to Desmogleins. Genomics 1997, 44, 330–335. [Google Scholar] [CrossRef]
- Brodehl, A.; Belke, D.D.; Garnett, L.; Martens, K.; Abdelfatah, N.; Rodriguez, M.; Diao, C.; Chen, Y.-X.; Gordon, P.M.K.; Nygren, A.; et al. Transgenic Mice Overexpressing Desmocollin-2 (DSC2) Develop Cardiomyopathy Associated with Myocardial Inflammation and Fibrotic Remodeling. PLoS ONE 2017, 12, e0174019. [Google Scholar] [CrossRef] [Green Version]
- Bhuiyan, Z.A.; Jongbloed, J.D.H.; van der Smagt, J.; Lombardi, P.M.; Wiesfeld, A.C.P.; Nelen, M.; Schouten, M.; Jongbloed, R.; Cox, M.G.P.J.; van Wolferen, M.; et al. Desmoglein-2 and Desmocollin-2 Mutations in Dutch Arrhythmogenic Right Ventricular Dysplasia/Cardiomypathy Patients. Circ. Cardiovasc. Genet. 2009, 2, 418–427. [Google Scholar] [CrossRef]
- Reams, R.Y.; Glickman, L.T.; Harrington, D.D.; Thacker, H.L.; Bowersock, T.L. Streptococclcs Suis Infection in Swine: A Retrospective Study of 256 Cases. Part II. Clinical Signs, Gross and Microscopic Lesions, and Coexisting Microorganisms. J. Vet. Diagn. Investig. 1994, 6, 326–334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tecles, F.; Rubio, C.P.; Contreras-Aguilar, M.D.; Lopez-Arjona, M.; Martinez-Miro, S.; Martinez-Subiela, S.; Ceron, J.J. Adenosine Deaminase Activity in Pig Saliva: Analytical Validation of Two Spectrophotometric Assays. J. Vet. Diagn. Investig. 2018, 30, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, M.; Dahl, J.; Jacobsen, S.; Jacobson, M.; Andersen, P.H.; Bækbo, P.; Escribano, D.; Cerón, J.J.; Tecles, F. Changes of Adenosine Deaminase Activity in Serum and Saliva around Parturition in Sows with and without Postpartum Dysgalactia Syndrome. BMC Vet. Res. 2021, 17, 352. [Google Scholar] [CrossRef] [PubMed]
- Contreras-Aguilar, M.D.; Tvarijonaviciute, A.; Monkeviciene, I.; Martín-Cuervo, M.; González-Arostegui, L.G.; Franco-Martínez, L.; Cerón, J.J.; Tecles, F.; Escribano, D. Characterization of Total Adenosine Deaminase Activity (ADA) and Its Isoenzymes in Saliva and Serum in Health and Inflammatory Conditions in Four Different Species: An Analytical and Clinical Validation Pilot Study. BMC Vet. Res. 2020, 16, 384. [Google Scholar] [CrossRef] [PubMed]
- Prims, S.; Van Raemdonck, G.; Vanden Hole, C.; Van Cruchten, S.; Van Ginneken, C.; Van Ostade, X.; Casteleyn, C. On the Characterisation of the Porcine Gland-Specific Salivary Proteome. J. Proteom. 2019, 196, 92–105. [Google Scholar] [CrossRef]
- Kang, S.S.; Ren, Y.; Liu, C.C.; Kurti, A.; Baker, K.E.; Bu, G.; Asmann, Y.; Fryer, J.D. Lipocalin-2 Protects the Brain during Inflammatory Conditions. Mol. Psychiatry 2018, 23, 344–350. [Google Scholar] [CrossRef]
- Srinivasan, G.; Aitken, J.D.; Zhang, B.; Carvalho, F.A.; Chassaing, B.; Shashidharamurthy, R.; Borregaard, N.; Jones, D.P.; Gewirtz, A.T.; Vijay-Kumar, M. Lipocalin 2 Deficiency Dysregulates Iron Homeostasis and Exacerbates Endotoxin-Induced Sepsis. J. Immunol. 2012, 189, 1911–1919. [Google Scholar] [CrossRef] [Green Version]
- Nikinmaa, M. Oxygen and Carbon Dioxide Transport in Vertebrate Erythrocytes: An Evolutionary Change in the Role of Membrane Transport. J. Exp. Biol. 1997, 200, 369–380. [Google Scholar] [CrossRef]
- Yoo, H.; Ku, S.-K.; Kim, S.-W.; Bae, J.-S. Early Diagnosis of Sepsis Using Serum Hemoglobin Subunit Beta. Inflammation 2015, 38, 394–399. [Google Scholar] [CrossRef]
- Olumee-Shabon, Z.; Chattopadhaya, C.; Myers, M.J. Proteomics Profiling of Swine Serum Following Lipopolysaccharide Stimulation. Rapid Commun. Mass Spectrom. 2020, 34, e8639. [Google Scholar] [CrossRef]
- Muñoz-Prieto, A.; Escribano, D.; Contreras-Aguilar, M.D.; Horvatić, A.; Guillemin, N.; Jacobsen, S.; Cerón, J.J.; Mrljak, V. Tandem Mass Tag (TMT) Proteomic Analysis of Saliva in Horses with Acute Abdominal Disease. Animals 2021, 11, 1304. [Google Scholar] [CrossRef]
- Borges-Rodriguez, M.; Shields, C.A.; Travis, O.K.; Tramel, R.W.; Baik, C.H.; Giachelli, C.A.; Tardo, G.A.; Williams, J.M.; Cornelius, D.C. Platelet Inhibition Prevents Nlrp3 Inflammasome Activation and Sepsis-Induced Kidney Injury. Int. J. Mol. Sci. 2021, 22, 10330. [Google Scholar] [CrossRef]
- Wang, Y.; Ouyang, Y.; Liu, B.; Ma, X.; Ding, R. Platelet Activation and Antiplatelet Therapy in Sepsis: A Narrative Review. Thromb. Res. 2018, 166, 28–36. [Google Scholar] [CrossRef]
- Franco-Martínez, L.; Horvatić, A.; Gelemanović, A.; Samardžija, M.; Mrljak, V.; Contreras-Aguilar, M.D.; Martínez-Subiela, S.; Dąbrowski, R.; Tvarijonaviciute, A. Changes in the Salivary Proteome Associated With Canine Pyometra. Front. Vet. Sci. 2020, 7, 277. [Google Scholar] [CrossRef]
- Cerón, J.J.; Contreras-Aguilar, M.D.; Escribano, D.; Martínez-Miró, S.; López-Martínez, M.J.; Ortín-Bustillo, A.; Franco-Martínez, L.; Rubio, C.P.; Muñoz-Prieto, A.; Tvarijonaviciute, A.; et al. Basics for the Potential Use of Saliva to Evaluate Stress, Inflammation, Immune System, and Redox Homeostasis in Pigs. BMC Vet. Res. 2022, 18, 81. [Google Scholar] [CrossRef]
- Andersen, C.B.F.; Stødkilde, K.; Sæderup, K.L.; Kuhlee, A.; Raunser, S.; Graversen, J.H.; Moestrup, S.K. Haptoglobin. Antioxid. Redox. Signal. 2017, 26, 814–831. [Google Scholar] [CrossRef]
- López-Martínez, M.J.; Franco-Martínez, L.; Martínez-Subiela, S.; Cerón, J.J. Biomarkers of Sepsis in Pigs, Horses and Cattle: From Acute Phase Proteins to Procalcitonin. Anim. Health Res. Rev. 2022, 23, 82–99. [Google Scholar] [CrossRef]
- Cerón, J.J.; Tecles, F.; Escribano, D.; Fuentes-Pardo, P.; Martínez-Subiela, S. Acute Phase Proteins in Pigs: From Theory to Practice. Suis 2016, 127. Available online: https://www.portalveterinaria.com/porcino/articulos/12951/proteinas-de-fase-aguda-en-el-ganado-porcino-de-la-teoria-a-la-practica.html (accessed on 7 July 2022).
- Yin, C.; Liu, W.; Liu, Z.; Huang, Y.; Ci, L.; Zhao, R.; Yang, X. Identification of Potential Serum Biomarkers in Pigs at Early Stage after Lipopolysaccharide Injection. Res. Vet. Sci. 2017, 111, 140–146. [Google Scholar] [CrossRef]
- Sali, V.; Veit, C.; Valros, A.; Junnikkala, S.; Heinonen, M.; Nordgreen, J. Dynamics of Salivary Adenosine Deaminase, Haptoglobin, and Cortisol in Lipopolysaccharide-Challenged Growing Pigs. Front. Vet. Sci. 2021, 8, 698628. [Google Scholar] [CrossRef]
- Gomez-Laguna, J.; Gutierrez, A.; Pallares, F.J.; Salguero, F.J.; Ceron, J.J.; Carrasco, L. Haptoglobin and C-Reactive Protein as Biomarkers in the Serum, Saliva and Meat Juice of Pigs Experimentally Infected with Porcine Reproductive and Respiratory Syndrome Virus. Vet. J. 2010, 185, 83–87. [Google Scholar] [CrossRef] [PubMed]
- De Haas, C.J.C. New Insights into the Role of Serum Amyloid P Component, a Novel Lipopolysaccharide-Binding Protein. FEMS Immunol. Med. Microbiol. 1999, 26, 197–202. [Google Scholar] [CrossRef]
- Pilling, D.; Gomer, R.H. The development of serum amyloid P as a possible therapeutic. Front. Immunol. 2018, 9, 2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poulsen, E.T.; Pedersen, K.W.; Marzeda, A.M.; Enghild, J.J. Serum Amyloid P Component (SAP) Interactome in Human Plasma Containing Physiological Calcium Levels. Biochemistry 2017, 56, 896–902. [Google Scholar] [CrossRef] [PubMed]
- Wake, H. Histidine-Rich Glycoprotein Modulates the Blood-Vascular System in Septic Condition. Acta Med. Okayama 2019, 73, 379–382. [Google Scholar]
- Kuroda, K.; Wake, H.; Mori, S.; Hinotsu, S.; Nishibori, M.; Morimatsu, H. Decrease in Histidine-Rich Glycoprotein as a Novel Biomarker to Predict Sepsis Among Systemic Inflammatory Response Syndrome. Crit. Care Med 2018, 46, 570–576. [Google Scholar] [CrossRef] [Green Version]
- Kacprzyk, L.; Rydengård, V.; Mörgelin, M.; Davoudi, M.; Pasupuleti, M.; Malmsten, M.; Schmidtchen, A. Antimicrobial Activity of Histidine-Rich Peptides Is Dependent on Acidic Conditions. Biochim. Biophys. Acta 2007, 1768, 2667–2680. [Google Scholar] [CrossRef] [Green Version]
- Burger, D.; Dayer, J.-M. High-Density Lipoprotein-Associated Apolipoprotein A-I: The Missing Link between Infection and Chronic Inflammation? Autoimmun. Rev. 2002, 1, 111–117. [Google Scholar] [CrossRef]
- Sorensen, N.S.; Tegtmeier, C.; Andresen, L.O.; Piñeiro, M.; Toussaint, M.J.M.; Campbell, F.M.; Lampreave, F.; Heegaard, P.M.H. The Porcine Acute Phase Protein Response to Acute Clinical and Subclinical Experimental Infection with Streptococcus Suis. Vet. Immunol. Immunopathol. 2006, 113, 157–168. [Google Scholar] [CrossRef]
- Heegaard, P.M.; Stockmarr, A.; Piñeiro, M.; Carpintero, R.; Lampreave, F.; Campbell, F.M.; Eckersall, P.; Toussaint, M.J.; Gruys, E.; Sorensen, N. Optimal Combinations of Acute Phase Proteins for Detecting Infectious Disease in Pigs. Vet. Res. 2011, 42, 50. [Google Scholar] [CrossRef] [Green Version]
- Obradovic, M.R.; Segura, M.; Segalés, J.; Gottschalk, M. Review of the Speculative Role of Co-Infections in Streptococcus Suis-Associated Diseases in Pigs. Vet. Res. 2021, 52, 49. [Google Scholar] [CrossRef]
- Petrocchi-Rilo, M.; Martínez-Martínez, S.; Aguarón-Turrientes, Á.; Roca-Martínez, E.; García-Iglesias, M.-J.; Pérez-Fernández, E.; González-Fernández, A.; Herencia-Lagunar, E.; Gutiérrez-Martín, C.-B. Anatomical Site, Typing, Virulence Gene Profiling, Antimicrobial Susceptibility and Resistance Genes of Streptococcus Suis Isolates Recovered from Pigs in Spain. Antibiotics 2021, 10, 707. [Google Scholar] [CrossRef]
- Okwumabua, O.; O’Connor, M.; Shull, E. A Polymerase Chain Reaction (PCR) Assay Specific for Streptococcus Suis Based on the Gene Encoding the Glutamate Dehydrogenase. FEMS Microbiol. Lett. 2003, 218, 79–84. [Google Scholar] [CrossRef]
- Horvatic, A.; Guillemin, N.; Kaab, H.; McKeegan, D.; O’Reilly, E.; Bain, M.; Kules, J.; Eckersall, P.D. Integrated Dataset on Acute Phase Protein Response in Chicken Challenged with Escherichia Coli Lipopolysaccharide Endotoxin. Data Brief 2018, 21, 684–699. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: http://www.r-project.org/index.html (accessed on 26 September 2022).
- Kuleš, J.; Lovrić, L.; Gelemanović, A.; Beer Ljubić, B.; Rubić, I.; Bujanić, M.; Konjević, D. Complementary Liver and Serum Protein Profile in Wild Boars Infected by the Giant Liver Fluke Fascioloides Magna Using Tandem Mass Tags Quantitative Approach. J. Proteom. 2021, 247, 104332. [Google Scholar] [CrossRef]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.-P.; Mushayamaha, T.; Thomas, P.D. PANTHER Version 16: A Revised Family Classification, Tree-Based Classification Tool, Enhancer Regions and Extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Gillespie, M.; Jassal, B.; Stephan, R.; Milacic, M.; Rothfels, K.; Senff-Ribeiro, A.; Griss, J.; Sevilla, C.; Matthews, L.; Gong, C.; et al. The Reactome Pathway Knowledgebase 2022. Nucleic Acids Res. 2022, 50, D687–D692. [Google Scholar] [CrossRef]
- Tecles, F.; Escribano, D.; Contreras-Aguilar, M.D.; Rubio, C.P.; Szczubial, M.; Ceron, J.J.; Dabrowski, R.; Tvarijonaviciute, A. Evaluation of Adenosine Deaminase in Saliva and Serum, and Salivary Alpha-Amylase, in Canine Pyometra at Diagnosis and after Ovariohysterectomy. Vet. J. 2018, 236, 102–110. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathway | FDR | Count | Genes | |
|---|---|---|---|---|
| Observed | Background | |||
| Platelet degranulation | <0.001 | 7 | 141 | TF, ECM1, APOA1, A1BG, ALDOA, HRG, VCL |
| Response to elevated platelet cytosolic Ca2+ | <0.001 | 7 | 148 | TF, ECM1, APOA1, A1BG, ALDOA, HRG, VCL |
| Hemostasis | <0.001 | 10 | 803 | IGHM, TF, ECM1, HBB, APOA1, A1BG, ALDOA, HRG, VCL |
| Platelet activation, signaling and aggregation | <0.001 | 7 | 293 | TF, ECM1, APOA1, A1BG, ALDOA, HRG, VCL |
| Neutrophil degranulation | <0.001 | 8 | 480 | TF, SERPINB12, GSN, KRT1, HBB, A1BG, ALDOA, VCL |
| Amyloid fiber formation | 0.010 | 3 | 89 | TF, GSN, APOA1 |
| Innate immune system | 0.026 | 8 | 1345 | TF, SERPINB12, GSN, KRT1, HBB, A1BG, ALDOA, VCL |
| Formation of the cornified envelope | 0.027 | 3 | 138 | KRT1, DSG3, DSC2 |
| Apoptotic cleavage of cellular proteins | 0.029 | 2 | 38 | GSN, DSG3 |
| Pathway | FDR | Count | Genes | |
|---|---|---|---|---|
| Observed | Background | |||
| Platelet degranulation | 6.305 × 105 | 6 | 141 | LOC100156325, TF, ALB, APOA1, HRG, CLU |
| Response to elevated platelet cytosolic Ca2+ | 1.859 × 104 | 6 | 148 | LOC100156325, TF, ALB, APOA1, HRG, CLU |
| Post-translational protein phosphorylation | 0.002 | 4 | 109 | TF, ITIH2, ALB, APOA1 |
| Complement cascade | 0.002 | 5 | 156 | APCS, VTN, CLU, CPN2, CFB |
| Regulation of complement cascade | 0.017 | 4 | 139 | VTN, CLU, CPN2, CFB |
| Regulation of insulin-like growth factor (IGF) transport and uptake by insulin-like growth factor binding proteins (IGFBPs) | 0.020 | 4 | 127 | TF, ITIH2, ALB, APOA1 |
| Neutrophil degranulation | 0.020 | 5 | 480 | LOC100156325, TF, MGAM, HP, LBP |
| Scavenging of heme from plasma | 0.023 | 3 | 106 | ALB, HP, APOA1 |
| Platelet activation, signaling and aggregation | 0.030 | 6 | 293 | LOC100156325, TF, ALB, APOA1, HRG, CLU |
| Antimicrobial peptides | 0.042 | 3 | 123 | TF, LBP, CLU |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Martínez, M.J.; Beletić, A.; Kuleš, J.; Rešetar-Maslov, D.; Rubić, I.; Mrljak, V.; Manzanilla, E.G.; Goyena, E.; Martínez-Subiela, S.; Cerón, J.J.; et al. Revealing the Changes in Saliva and Serum Proteins of Pigs with Meningitis Caused by Streptococcus Suis: A Proteomic Approach. Int. J. Mol. Sci. 2022, 23, 13700. https://doi.org/10.3390/ijms232213700
López-Martínez MJ, Beletić A, Kuleš J, Rešetar-Maslov D, Rubić I, Mrljak V, Manzanilla EG, Goyena E, Martínez-Subiela S, Cerón JJ, et al. Revealing the Changes in Saliva and Serum Proteins of Pigs with Meningitis Caused by Streptococcus Suis: A Proteomic Approach. International Journal of Molecular Sciences. 2022; 23(22):13700. https://doi.org/10.3390/ijms232213700
Chicago/Turabian StyleLópez-Martínez, María José, Anđelo Beletić, Josipa Kuleš, Dina Rešetar-Maslov, Ivana Rubić, Vladimir Mrljak, Edgar Garcia Manzanilla, Elena Goyena, Silvia Martínez-Subiela, José Joaquín Cerón, and et al. 2022. "Revealing the Changes in Saliva and Serum Proteins of Pigs with Meningitis Caused by Streptococcus Suis: A Proteomic Approach" International Journal of Molecular Sciences 23, no. 22: 13700. https://doi.org/10.3390/ijms232213700